
Transcriptomic Revealed That Selenium-Rich Lactobacillus plantarum Alleviated Cadmium-Induced Immune Responses in Bulatmai barbel Luciobarbus capito Kidneys

Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs and Reagents
2.2. Preparation of Se-Enriched Lactic Acid Bacteria
2.3. Scanning Electron Microscopy Observation of Se and Morphological Distribution of Bacteria
2.4. Experimental Design
2.5. Blood and Serum Immunological Tests
2.6. Kidney Tissue Sections
2.7. Transcriptome
2.8. Gene Ontology and Enrichment Analysis
2.9. qRT-PCR
2.10. Data Analysis
3. Results
3.1. Se-Enriched Lactic Acid Bacteria
3.2. Kidney Histology and Morphology
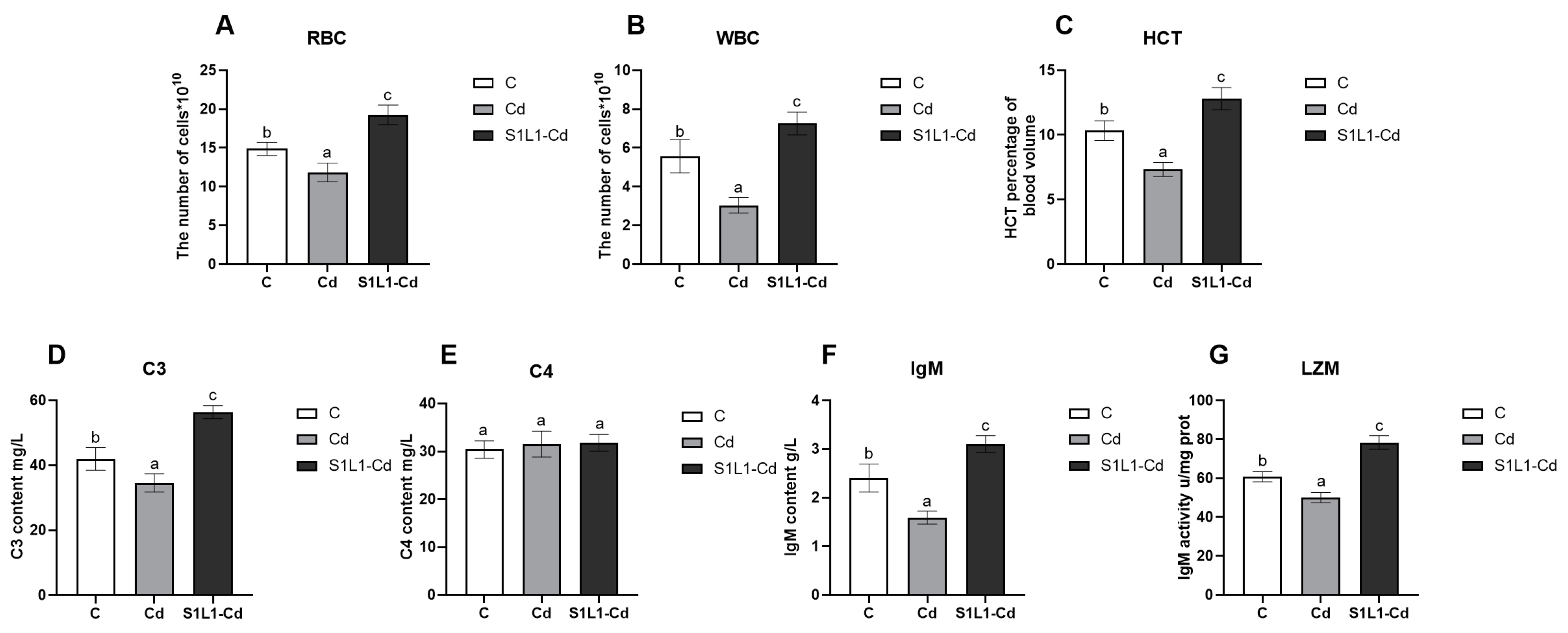
3.3. Haemocyte Parameters and Serum Immune Responses
3.4. Kidney Damage Indicators
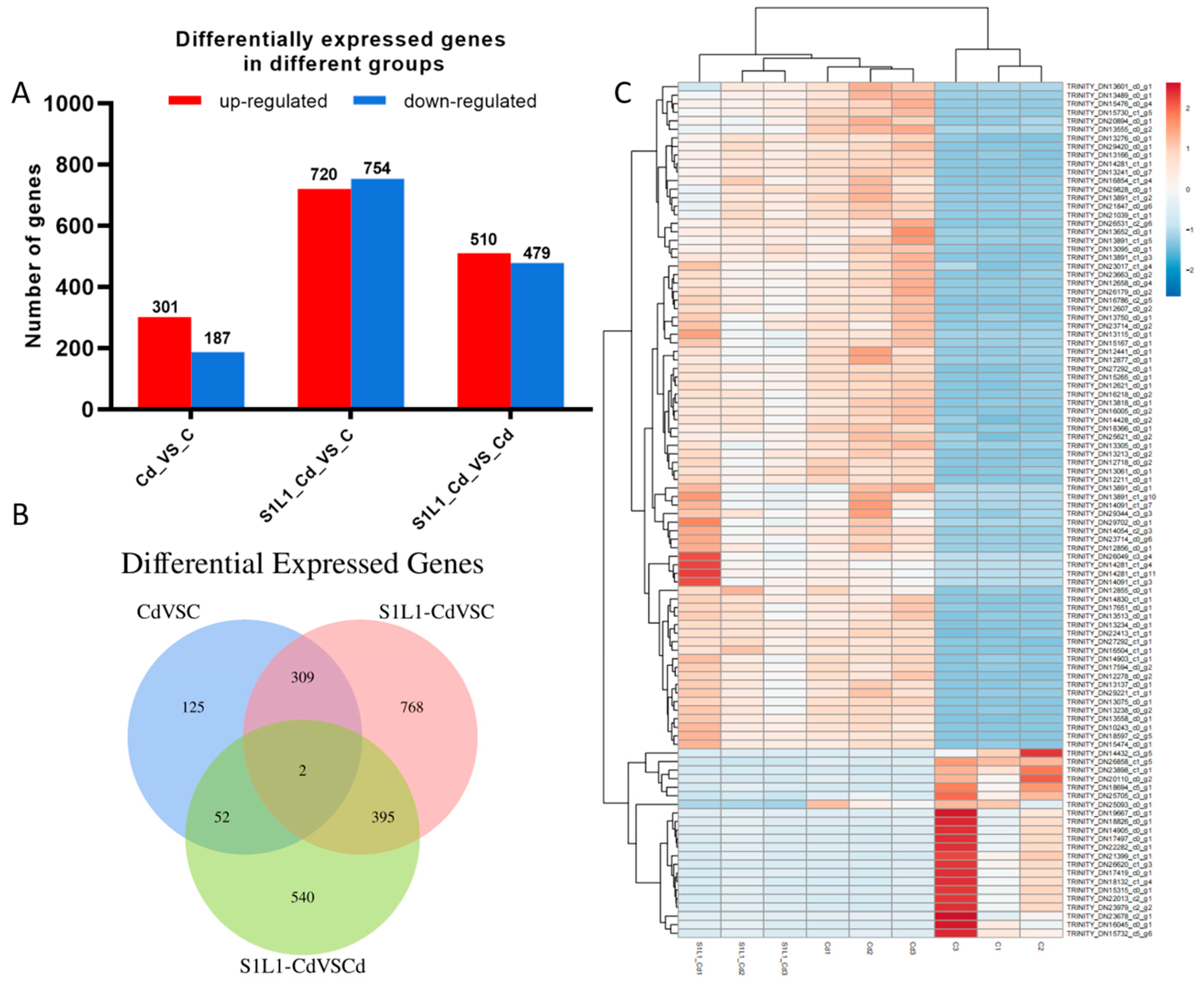
3.5. Transcriptome
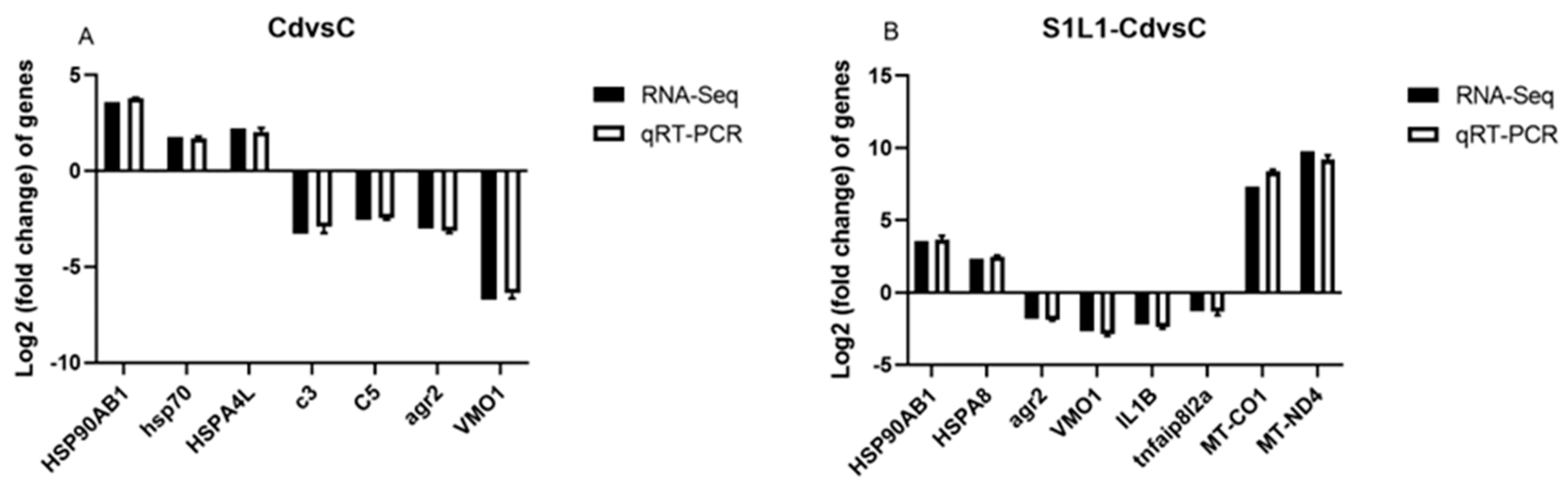
3.6. qRT-PCR Validation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.L.; Xu, W.; Wei, Y.Y.; Xu, Q.Y. The complete mitochondrial genome of Barbus capito (Cypriniformes, Cyprinidae). Mitochondr Dna. 2013, 24, 326–327. [Google Scholar] [CrossRef] [PubMed]
- Canli, M.A.Y.O.; Ay, Ö.; Kalay, M. Levels of Heavy Metals (Cd, Pb, Cu, Cr and Ni) in Tissue of Cyprinus carpio, Barbus capitoand Chondrostoma regiumfrom the Seyhan River, Turkey. Turk. J. Zool. 1998, 22, 149–158. [Google Scholar]
- Koca, S.; Koca, Y.B.; Yildiz, Ş.; Gürcü, B. Genotoxic and histopathological effects of water pollution on two fish species, Barbus capito pectoralis and Chondrostoma nasus in the Büyük Menderes River, Turkey. Biol. Trace Element. Res. 2008, 122, 276–291. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Xu, W.; Zhang, Y.; Sun, Q.; Li, Z.; Geng, L.; Teng, X. Transcriptome analysis revealed the mechanism of Luciobarbus capito (L. capito) adapting high salinity: Antioxidant capacity, heat shock proteins, immunity. Mar. Pollut. Bull. 2023, 192, 115017. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Xu, W.; Che, X.; Cui, J.; Shang, X.; Teng, X.; Jia, Z. Effect of arsenic stress on the intestinal structural integrity and intestinal flora abundance of Cyprinus carpio. Front. Microbiol. 2023, 14, 1179397. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Garrett, S.H.; Sens, M.A.; Sens, D.A. Cadmium, environmental exposure, and health outcomes. Environ. Health Persp. 2010, 118, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Rani, A.; Kumar, A.; Lal, A.; Pant, M. Cellular mechanisms of cadmium-induced toxicity: A review. Int. J. Environ. Heal. Res. 2014, 24, 378–399. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, Z.; Hu, R.; Pi, S.; Wei, Z.; Wang, C.; Yang, F.; Xing, C.; Nie, G.; Hu, G. New insights into crosstalk between pyroptosis and autophagy co-induced by molybdenum and cadmium in duck renal tubular epithelial cells. J. Hazard. Mater. 2021, 416, 126138. [Google Scholar] [CrossRef]
- Zhang, N.; Bevan, M.J. CD8+ T cells: Foot soldiers of the immune system. Immunity 2011, 35, 161–168. [Google Scholar] [CrossRef]
- Castellino, F.; Germain, R.N. Cooperation between CD4+ and CD8+ T cells: When, where, and how. Annu. Rev. Immunol. 2006, 24, 519–540. [Google Scholar] [CrossRef] [PubMed]
- Goyal, T.; Mitra, P.; Singh, P.; Sharma, S.; Purohit, P.; Sharma, P. Effect of occupational co-exposure to lead and cadmium on selected immunomodulatory cytokines. Toxicol. Ind. Health 2022, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tucovic, D.; Aleksandrov, A.P.; Mirkov, I.; Ninkov, M.; Kulas, J.; Zolotarevski, L.; Vukojevic, V.; Mutic, J.; Tatalovic, N.; Kataranovski, M. Oral cadmium exposure affects skin immune reactivity in rats. Ecotoxicol. Environ. Safe. 2018, 164, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Minetti, G. Mevalonate pathway, selenoproteins, redox balance, immune system, Covid-19: Reasoning about connections. Med. Hypotheses 2020, 144, 110128. [Google Scholar] [CrossRef]
- Shang, X.; Xu, W.; Zhao, Z.; Luo, L.; Zhang, Q.; Li, M.; Geng, L. Effects of exposure to cadmium (Cd) and selenium-enriched Lactobacillus plantarum in Luciobarbus capito: Bioaccumulation, antioxidant responses and intestinal microflora. COMP Biochem. Phys. C 2022, 257, 109352. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Wu, Q.; Deng, H.; Yu, Y.; Tang, W.; Deng, Y.; Deng, J. Effects of selenium on the immunotoxicity of subacute arsenic poisoning in chickens. Biol. Trace Element. Res. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, J.; Jiang, C.; Cui, J.; Hong, L.; Hao, Z.; Teng, X. Se alleviated Pb-caused neurotoxicity in chickens: SPS2-GPx1-GSH-IL-2/IL-17-NO pathway, selenoprotein suppression, oxidative stress, and inflammatory injury. Antioxidants 2024, 13, 370. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yu, W.; Hao, Z.; Qiu, M.; Cui, J.; Tang, Y.; Liu, H. Molecular mechanism of selenium against lead-induced apoptosis in chicken brainstem relating to heat shock protein, selenoproteins, and inflammatory cytokines. Ecotox Environ Safe. 2024, 272, 116028. [Google Scholar] [CrossRef] [PubMed]
- Banni, M.; Chouchene, L.; Said, K.; Kerkeni, A.; Messaoudi, I. Mechanisms underlying the protective effect of zinc and selenium against cadmium-induced oxidative stress in zebrafish Danio rerio. Biometals 2011, 24, 981–992. [Google Scholar] [CrossRef]
- Reid, G. Probiotics: Definition, scope and mechanisms of action. Best. Pract. Res. Clin. Gastroenterol. 2016, 30, 17–25. [Google Scholar] [CrossRef]
- Zhang, Q.; Shang, X.; Geng, L.; Che, X.; Wei, H.; Tang, S.; Xu, W. Dietary Selenium-Rich Lactobacillus plantarum Alleviates Cadmium-Induced Oxidative Stress and Inflammation in Bulatmai barbel Luciobarbus capito. Fishes 2023, 8, 136. [Google Scholar] [CrossRef]
- Shang, X.; Sun, Q.; Yin, Y.; Zhang, Y.; Zhang, P.; Mao, Q.; Li, Y. Reducingmercury accumulation in common carp using selenium-enriched Bacillus subtilis. Aquacult Rep. 2021, 19, 100609. [Google Scholar] [CrossRef]
- Ibrahim, F.; Halttunen, T.; Tahvonen, R.; Salminen, S. Probiotic bacteria as potential detoxification tools: Assessing their heavy metal binding isotherms. Can. J. Microbiol. 2006, 52, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Teemu, H.; Seppo, S.; Jussi, M.; Raija, T.; Kalle, L. Reversible surface binding of cadmium and lead by lactic acid and bifidobacteria. Int. J. Food Microbiol. 2008, 125, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kumar, V.; Panwar, R.; Ram, C. Efficacy of indigenous probiotic Lactobacillus strains to reduce cadmium bioaccessibility-an in vitro digestion model. Environ. Sci. Pollut. Res. 2017, 24, 1241–1250. [Google Scholar] [CrossRef]
- Zhai, Q.; Liu, Y.; Wang, C.; Qu, D.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus plantarum CCFM8661 modulates bile acid enterohepatic circulation and increases lead excretion in mice. Food Funct. 2019, 10, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.M.; Hodson, P.V. Field studies using fish biomarkers–How many fish are enough? Mar. Pollut. Bull. 2012, 64, 2871–2876. [Google Scholar] [CrossRef]
- Tanaka, N.; Izawa, T.; Kuwamura, M.; Higashiguchi, N.; Kezuka, C.; Kurata, O.; Yamate, J. The first case of infectious spleen and kidney necrosis virus (ISKNV) infection in aquarium-maintained mandarin fish, S iniperca chuatsi (B asilewsky), in Japan. J. Fish. Dis. 2014, 37, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhong, M.; Zhou, R.; Qin, W.; Si, Y. Se (IV) reduction and extracellular biosynthesis of Nano-Se (0) by Shewanella oneidensis MR-1 and Shewanella putrefaciens. Process Biochem. 2023, 130, 481–491. [Google Scholar] [CrossRef]
- McRae, N.K.; Gaw, S.; Brooks, B.W.; Glover, C.N. Oxidative stress in the galaxiid fish, Galaxias maculatus, exposed to binary waterborne mixtures of the pro-oxidant cadmium and the anti-oxidant diclofenac. Environ. Pollt. 2019, 247, 638–646. [Google Scholar] [CrossRef]
- Hernández-Cruz, E.Y.; Amador-Martínez, I.; Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Chaverri, J.P. Renal damage induced by cadmium and its possible therapy by mitochondrial transplantation. Chem-Biol. Interact. 2022, 361, 109961. [Google Scholar] [CrossRef]
- Imed, M.; Fatima, H.; Abdelhamid, K. Protective effects of selenium (Se) and zinc (Zn) on cadmium (Cd) toxicity in the liver of the rat: Effects on the oxidative stress. Ecotox Environ. Safe 2009, 72, 1559–1564. [Google Scholar] [CrossRef]
- Has-Schön, E.; Bogut, I.; Vuković, R.; Galović, D.; Bogut, A.; Horvatić, J. Distribution and age-related bioaccumulation of lead (Pb), mercury (Hg), cadmium (Cd), and arsenic (As) in tissues of common carp (Cyprinus carpio) and European catfish (Sylurus glanis) from the Buško Blato reservoir (Bosnia and Herzegovina). Chemosphere 2015, 135, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Buha, A.; Wallace, D.; Matovic, V.; Schweitzer, A.; Oluic, B.; Micic, D.; Djordjevic, V. Cadmium exposure as a putative risk factor for the development of pancreatic cancer: Three different lines of evidence. Biomed. Res. Int. 2017, 2017, 1981837. [Google Scholar] [CrossRef]
- Andjelkovic, M.; Buha Djordjevic, A.; Antonijevic, E.; Antonijevic, B.; Stanic, M.; Kotur-Stevuljevic, J.; Bulat, Z. Toxic effect of acute cadmium and lead exposure in rat blood, liver, and kidney. Int. J. Environ. Res. Public Health 2019, 16, 274. [Google Scholar] [CrossRef]
- Remyla, S.R.; Ramesh, M.; Sajwan, K.S.; Senthil Kumar, K. Influence of zinc on cadmium induced haematological and bio-chemical responses in a freshwater teleost fish Catla catla. Fish. Physiol. Biochem. 2008, 34, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.A.; Nisa, Z.U.; Mehmood, A.; Anjum, M.S.; Shahzad, K. Metal-induced nephrotoxicity to diabetic and non-diabetic Wistar rats. Environ. Sci. Pollut. Res. 2019, 26, 31111–31118. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Verma, P.K.; Sood, S.; Yousuf, R.; Raina, R. Oxidative renal damage induced by fluoride and dimethoate and its mitigation by Zingiber officinale in Wistar rats. Research Square 2022, preprint. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, H.; Hao, D.; Wang, J.; Zhu, R.; Liu, W.; Liu, C. Selenium regulates the mitogen-activated protein kinase pathway to protect broilers from hexavalent chromium-induced kidney dysfunction and apoptosis. Ecotox Environ. Safe. 2022, 239, 113629. [Google Scholar] [CrossRef]
- Magnadóttir, B. Innate immunity of fish (overview). Fish. Shellfish. Immun. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Gao, X.Q.; Fei, F.; Huo, H.H.; Huang, B.; Meng, X.S.; Zhang, T.; Liu, B.L. Impact of nitrite exposure on plasma biochemical parameters and immune-related responses in Takifugu rubripes. Aquat. Toxicol. 2020, 218, 105362. [Google Scholar] [CrossRef]
- Jia, R.; Liu, B.L.; Han, C.; Huang, B.; Lei, J.L. Effects of ammonia exposure on stress and immune response in juvenile turbot (Scophthalmus maximus). Aquac. Res. 2017, 48, 3149–3162. [Google Scholar] [CrossRef]
- Yazdi, M.H.; Mahdavi, M.; Setayesh, N.; Esfandyar, M.; Shahverdi, A.R. Selenium nanoparticle-enriched Lactobacillus brevis causes more efficient immune responses in vivo and reduces the liver metastasis in metastatic form of mouse breast cancer. J. Pharm. Sci. 2013, 21, 1–9. [Google Scholar] [CrossRef]
- Kong, Y.; Gao, C.; Du, X.; Zhao, J.; Li, M.; Shan, X.; Wang, G. Effects of single or conjoint administration of lactic acid bacteria as potential probiotics on growth, immune response and disease resistance of snakehead fish (Channa argus). Fish. Shellfish. Immun. 2020, 102, 412–421. [Google Scholar] [CrossRef]
- Muller, W.A. Mechanisms of transendothelial migration of leukocytes. Circ. Res. 2009, 105, 223–230. [Google Scholar] [CrossRef]
- Muller, W.A. How endothelial cells regulate transmigration of leukocytes in the inflammatory response. Am. J. Pathol. 2014, 184, 886–896. [Google Scholar] [CrossRef]
- Ferrante, A.W., Jr. The immune cells in adipose tissue. Diabetes Obes. Metab. 2013, 15, 34–38. [Google Scholar] [CrossRef]
- Gaffen, S.L. An overview of IL-17 function and signaling. Cytokine. 2008, 43, 402–407. [Google Scholar] [CrossRef]
- Zhao, L.; Tang, Y.; You, Z.; Wang, Q.; Liang, S.; Han, X.; Ma, X. Interleukin-17 contributes to the pathogenesis of autoimmune hepatitis through inducing hepatic interleukin-6 expression. PLoS ONE 2011, 6, e18909. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, S.; Song, X.; Hu, Q.; Pan, W. IL-17 enhances oxidative stress in hepatocytes through Nrf2/keap1 signal pathway activation. Int. J. Clin. Exp. Patho. 2018, 11, 3318. [Google Scholar]
- Chen, D.; Shen, F.; Liu, J.; Tang, H.; Teng, X.; Yang, F.; Liu, H. Luteolin enhanced antioxidant capability and induced pyroptosis through NF-κB/NLRP3/Caspase-1 in splenic lymphocytes exposure to ammonia. Sci. Total Environ. 2024, 919, 170699. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | Nucleotide Sequence (5′–3′) | Accession No. | |
|---|---|---|---|---|
| β-Actin | Beta-Actin | Forward Reverse | CATTGGAGGGCAAGTCTGGT CCCGAGATCCAACTACGAGC | NR145818.1 |
| HSP90AB1 | Heat shock protein 90ab1 | Forward Reverse | ACCACCCTGCTCTGTACTACT TCCTGAGGGTTGGGGATGAT | |
| HSP70 | Heat shock protein 70 | Forward Reverse | GCCATGAACCCCAACAACAC TTTCGGCTTCCCTCCATCAC | AY120894.1 |
| IL-1β | Interleukin-1β | Forward Reverse | ACCAGCTGGATTTGTCAGAAG ACATACTGAATTGAACTTTG | AB010701 |
| HSPA4L | Heat shock 70-kDa protein 4-like | Forward Reverse | AACAGGACTTGCCAAACCCA CAAAGTTGCATCCGCCCAAA | |
| agr2 | Anterior gradient 2 | Forward Reverse | AGCACTTGTCTCCTGATGGC TACAGGCGGTTAGCATAGCG | |
| MT-CO1 | Mitochondrially encoded cytochrome c oxidase I | Forward Reverse | TTAGCTGACTCGCCACACTC TACAATGCCAGTCAGGCCAC | |
| MT-ND4 | Mitochondrially encoded NADH dehydrogenase 4 | Forward Reverse | ACAAGCTCCATCTGCCTACG GAAGCTTCAGGGGGTTTGGA | |
| VMO1 | Vitelline membrane outer layer 1 | Forward Reverse | TGGGGTCACAGGGAAATGTG TGAACTGAGGCGTAGCTGTG | |
| HSPA8 | Heat shock protein family A member 8 | Forward Reverse | AGTGGACAAAAGCACTGGCA TTAAAGGCCAGCGACTCCAA | |
| tnfaip8l2a | Tumour necrosis factor alpha-induced protein 8-like protein 2 | Forward Reverse | GACAACAGCGGTGAGGTTCT AAGTCCTGTGCCACACTCAG | |
| C3 | complement component 3 | Forward Reverse | GTACGTGGGAAAGACGCTGA CAGCGTAAGTGCTGGTCAGA | |
| C5 | complement component 4 | Forward Reverse | GGAGGAACACACCAGCAAGT GCTCTCCACGAACCATGGAA |
| Sample | Raw_Reads | Clean_Reads |
|---|---|---|
| C1 | 39,890,490 | 38,813,784 |
| C2 | 43,258,692 | 42,393,916 |
| C3 | 43,489,474 | 42,578,024 |
| Cd1 | 39,470,924 | 37,924,708 |
| Cd2 | 42,786,912 | 41,503,984 |
| Cd3 | 43,699,384 | 41,839,558 |
| S1L1-Cd1 | 47,325,678 | 46,031,170 |
| S1L1-Cd2 | 44,356,428 | 42,993,436 |
| S1L1-Cd3 | 40,541,038 | 39,382,828 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Pang, Y.; Qin, Y.; Dong, Z.; Ma, Y.; Zhao, Y.; Zhang, Z.; Liu, J.; Mao, B.; Yin, B. Transcriptomic Revealed That Selenium-Rich Lactobacillus plantarum Alleviated Cadmium-Induced Immune Responses in Bulatmai barbel Luciobarbus capito Kidneys. Fishes 2024, 9, 230. https://doi.org/10.3390/fishes9060230
Sun Q, Pang Y, Qin Y, Dong Z, Ma Y, Zhao Y, Zhang Z, Liu J, Mao B, Yin B. Transcriptomic Revealed That Selenium-Rich Lactobacillus plantarum Alleviated Cadmium-Induced Immune Responses in Bulatmai barbel Luciobarbus capito Kidneys. Fishes. 2024; 9(6):230. https://doi.org/10.3390/fishes9060230
Chicago/Turabian StyleSun, Qingsong, Yuran Pang, Yuhan Qin, Ziting Dong, Yanling Ma, Yuan Zhao, Zhanning Zhang, Jinmei Liu, Binghui Mao, and Baishuang Yin. 2024. "Transcriptomic Revealed That Selenium-Rich Lactobacillus plantarum Alleviated Cadmium-Induced Immune Responses in Bulatmai barbel Luciobarbus capito Kidneys" Fishes 9, no. 6: 230. https://doi.org/10.3390/fishes9060230
APA StyleSun, Q., Pang, Y., Qin, Y., Dong, Z., Ma, Y., Zhao, Y., Zhang, Z., Liu, J., Mao, B., & Yin, B. (2024). Transcriptomic Revealed That Selenium-Rich Lactobacillus plantarum Alleviated Cadmium-Induced Immune Responses in Bulatmai barbel Luciobarbus capito Kidneys. Fishes, 9(6), 230. https://doi.org/10.3390/fishes9060230