1. Introduction
Muskellunge
Esox masquinongy is a highly prized sportfish throughout much of North America, and many state agencies have developed stocking programs with the intent to establish or augment natural Muskellunge populations to meet angler demand [
1]. A common concern among fisheries managers is that Muskellunge poststocking mortality may be highly variable among systems and years [
2,
3,
4,
5,
6]. Some of this variability can be explained by understanding the biotic and abiotic components of the fishery [
2,
6,
7,
8,
9], while others found that stocked Muskellunge movement and dispersal patterns [
2,
10] can influence Muskellunge poststocking success. Regardless of the mechanisms, Muskellunge poststocking survival is generally less than desired, and lapses in Muskellunge year-class development may occur and lead to diminishing Muskellunge populations, especially in systems where emigration can contribute to high Muskellunge mortality rates [
11,
12,
13] and/or where habitat degradation has limited Muskellunge natural recruitment [
14].
Because of the complexity and variability within aquatic ecosystems, little can be done to enhance stocking survival once fish leave the confines of advanced husbandry. Accordingly, agencies have spent considerable time evaluating the intrinsic characteristics of rearing Muskellunge [
15]. Culture practices such as rearing medium (i.e., line pond, trough or concrete raceways, earthen pond [
16,
17,
18]) and Muskellunge diet (i.e., artificial or live forage) can significantly influence fish physical characteristics, such as the coloration, health, and size of reared Muskellunge [
3,
19,
20,
21], which collectively have been associated with poor poststocking performance [
3,
21]. Thus, agencies strive to mitigate these shortcomings via the inclusion of several rearing strategies within their culture procedures. Subsequently, nearly continuous research has been conducted with the intent to evaluate different hatchery products and their poststocking survival and contribution to wild populations. Collectively, these studies consistently document that Muskellunge size at stocking substantially influences poststocking survival, primarily through minimizing predation risks [
2,
4,
15,
18,
22,
23,
24,
25,
26,
27,
28,
29,
30]. A better understanding of the influence of fish size and behavior of stocked Muskellunge may provide insight for improving the success of stocking.
While the size at stocking relationship is well known among managers, there is little consensus on the size at which stocked Muskellunge may overcome most predation risks. Previous research has consistently identified that Muskellunge stocked > 200 mm have higher survival probabilities than their smaller stocked counterparts [
4,
7,
18,
21,
23], but depending on stocking objectives, the interpretation of the success or failure of each stocked cohort is rather vague. Moreover, fish stockings are often assigned as a success or failure based on the manager’s perception of the outcome and not in context to the fish’s contribution to the overall fishery. Alternatively, many modern-day poststocking evaluations incorporate telemetry to examine individual fish behavior and identify biotic or abiotic interactions that may be resulting in recruitment bottlenecks [
5,
6,
10,
29,
31,
32]. One of the first evaluations of this type was performed on fall-stocked Muskellunge fingerlings (307–333 mm total length [TL]) in two Wisconsin natural lakes and documented variable survival of tagged fish over 34 d (43–85%) and that fish moved from the stocking location within 14 d [
2]. Other radiotelemetry evaluations examining age-0 or age-1 Muskellunge poststocking movement and survival are limited to those conducted in Southern US rivers and Midwestern reservoirs, but these studies documented poor survival despite high poststocking dispersal rates and larger stocking sizes (≥255 mm [
5,
6,
10]). These studies further document that poststocking survival is highly variable among systems and that poststocking predation risks may not be minimized at stocking size ranges up to at least 335 mm. More recently, age-1 Muskellunge telemetry data collected from Spirit Lake, Iowa, documented that stocked Muskellunge need to be reared to sizes beyond 375 mm for poststocking survival to be high (≥90% [
29]). Few observations of fish reared, radio-tagged, and stocked at TLs ≥ 375 mm exist; thus, it is relatively unknown if models relating to increased Muskellunge size at stocking accurately represent improvements in Muskellunge poststocking survival in the wild. Furthermore, the emergence of new hatchery production techniques has allowed more consistent production of large Muskellunge in Iowa [
31], yet the success of these stockings has not been validated.
In lakes where artificial propagation is needed to enhance or maintain Muskellunge populations, identifying cost-effective stocking strategies that augment the adult population may be the most important Muskellunge management tool available to managers. If stocking large yearling Muskellunge results in more predictable stocking success across a gradient of lake types, then more cost-effective stocking programs could be developed, and adult Muskellunge populations would benefit. Thus, the objective of my study was to evaluate poststocking survival, dispersal, movement, proximity to shore, depth, and vegetation usage of large (≥375 mm) yearling Muskellunge stocked in two Iowa natural lakes of varying complexity.
2. Materials and Methods
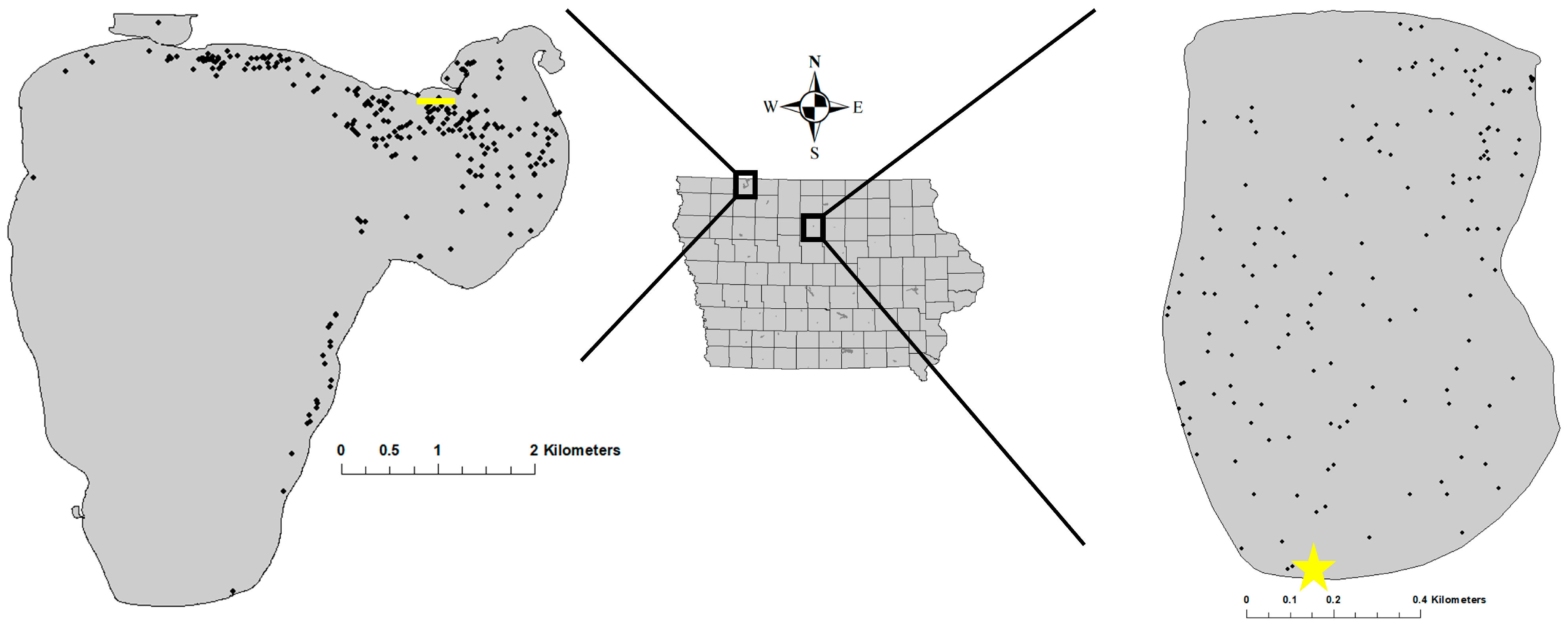
Study systems—Spirit Lake (2300 ha) is located in Dickinson County (northwest Iowa;
Figure 1) and has a watershed size of 15,349 ha that consists primarily of agriculture. Spirit Lake is a large bowl-shaped, eutrophic, wind-swept natural lake with few rock reefs and several shallow bays fringed with emergent vegetation. The lake’s basin slopes gradually with a mean depth of 4.8 m, a maximum depth of 7.4 m, and a water conductivity of 357 ± 9.9 µS/cm (mean ± standard error [SE]). Submersed aquatic vegetation is abundant during the open water season in Spirit Lake. Extensive curly leaf pondweed
Potamogeton crispus stands are common in shallow (<6.0 m) habitats from April to early July, and many native species (e.g., wild celery
Vallisneria americana; bushy pondweed
Najas flexilis; clasping-leaf pondweed
Potamogeton perfoliatus; coontail
Ceratophyllum demersum) are present during the remainder of the growing season. The Spirit Lake fishery is diverse, with several species of top-level predators managed both as sport fisheries and broodstock population sources for the state fish hatchery. Piscivores assemblage includes Muskellunge, Northern Pike
Esox lucius, Walleye
Sander vitreus, Largemouth Bass
Micropterus salmoides, and Smallmouth Bass
Micropterus dolomieu.
Lake Cornelia (Wright County;
Figure 1) has a similar basin shape to Spirit Lake but is a much smaller (99 ha; 399 ha watershed) and shallower (mean depth of 2.9 m; maximum depth of 6.3 m) natural lake located in north-central Iowa. Lake Cornelia has limited basin and habitat complexity, and water conductivity averages 321 ± 8.8 µS/cm (mean ± SE). A very small remnant of emergent vegetation exists on the northwest and south shores. Submergent vegetation consists of curly leaf pondweed early in the growing season and moderate to low amounts of sago pondweed
Potamogeton pectinatus, clasping-leaf pondweed, bushy pondweed, and longleaf pondweed
Potamogeton nodosus persisting in shallow (<2.7 m) water habitats throughout the summer months. In the fall of 2022, a whole-lake low-dose (8 ppb) rotenone experiment was conducted at Lake Cornelia in an attempt to manage non-native Yellow Bass
Morone mississippiensis. As part of this treatment, piscivore species that were less tolerant to these concentrations of rotenone were eliminated from the fishery (e.g., Northern Pike and Walleye). Restocking of the lake with both juvenile and adult fish began in April 2023 and included Walleye, Muskellunge, Largemouth Bass, and other species of piscivores that are not commonly managed in natural lakes (i.e., Flathead Catfish
Pylodictis olivaris; Blue Catfish
Ictalurus furcatus).
Fish tagging and stocking—On 18 May–19 May 2023, 25 large yearling Muskellunge (hereafter referred to as Muskellunge; mean TL ± SE = 418 ± 3.4 mm;
n = 10–15 per lake;
Table 1) were surgically implanted with a radio tag (Advanced Telemetry Systems, Isanti, MN, USA; Model F1440; 2.1 g in water; 158-d battery life; 0.4–0.9% of body weight) at the Spirit Lake Fish Hatchery, Spirit Lake, IA, USA. The average size of Muskellunge was similar between the two lakes (mean TL ± SE, Spirit Lake: 416 ± 4.8 mm; Lake Cornelia: 420 ± 5.0 mm). Fish were anesthetized with tricaine methanesulfonate-laden water (approximately 75 mg/L) and then measured (TL, mm), weighed (g), and a 12.5 mm Passive Integrated Transponder (PIT; Electronic Identification Devices, Ltd., Santa Barbara, CA, USA, Model FDX; 134.2 KHZ frequency) tag was implanted in the dorsal musculature. Prior to surgery, surgical tools and the transmitter were disinfected in a chlorhexidine solution (2% Chlorhexidine Gluconate), and fish were placed ventral side up on a homemade surgery cradle (sponge mounted on plastic tote [
33]. A scalpel was used to make an initial incision (approximately 25 mm in length) anterior to the pelvic girdle. Then, an exit hole for the transmitter whip antenna was created by inserting a 16-gauge hypodermic needle into the incision and exiting anterior to the anus. The whip antenna was threaded through the 16-gauge hypodermic needle, and the tag was carefully inserted into the fish [
33]. Nylon monofilament sutures (Scientify Global, Danbury, CT, USA) were used to close the incision (2 sutures per incision). The incision was sprayed with iodine prior to the fish being placed into covered indoor concrete raceways (4.8 m × 1.3 m × 1.0 m). On 19 May 2023, radio-tagged yearling Muskellunge scheduled to be stocked at either Spirit Lake (
n = 15) or Lake Cornelia (
n = 10) were loaded onto separate hatchery transportation trucks with untagged conspecifics. For Lake Cornelia, 265 yearling Muskellunge were transported approximately 183 km to a boat ramp where they were netted from the hatchery truck and released into the lake at the shore. At Spirit Lake, 439 yearling Muskellunge were transported 8.7 km to a boat ramp where they were then off-loaded from the transportation truck to a boat containing a large holding tank. Fish were then transported by boat where they were distributed haphazardly while slowly navigating a 0.5 km stretch of shoreline approximately 100 m from shore (
Figure 1).
Radiotelemetry and survival—At Spirit Lake, telemetry began the day after stocking and was continued daily for 5 days. At Lake Cornelia, telemetry was conducted on the second and sixth days poststocking. After the first week poststocking, telemetry was conducted on a weekly basis until all fish had been located or the entire lake had been searched. At Spirit Lake, it was impractical to search the entire lake basin each week for missing fish due to the size of the lake. Thus, weekly tracking was limited to the north shore and northeast basin (near fish stocking location) and occasionally the entire littoral (≤5.5 m water depth) periphery of the lake. Muskellunge tracking consisted of using an Advanced Telemetry Systems Model R4000 receiver connected to a three-element folding Yagi antenna that was affixed to a 4.3 m aluminum rod. The rod was placed in a custom-made stand within the boat which elevated the Yagi antenna and allowed the worker to efficiently scan for radio frequencies while traversing the lake. More specifically, the worker set the receiver to user-specific volume and gain (either maximum volume and gain or set at a lower level to avoid interference) to scan for each tag frequency. Each tag frequency was scanned for two seconds before advancing to the next tag frequency. Tracking first consisted of slowly (<16 kph) scanning the perimeter of the lake (antenna pointed to shore, approximately 100 m from shore) until a fish was detected. If a fish was not detected in nearshore habitats during a tracking event, transects (typically 100–150 m apart aided via the use of a Lowrance™ Gen 2 HDS-7 unit; Navico corp., Tulsa, OK, USA) were run in open water habitats and along the periphery (approximately 100 m from shore) of the lake with the antenna pointed towards offshore habitats. After detection, the direction of the tagged fish was determined by rotating the antenna until the signal was maximized. As the worker approached the tagged fish, the gain and volume were slowly reduced until the fish signal was faint. Then, a hand-held three-element folding Yagi antenna was used to pinpoint the location of the fish by reducing the receiver gain to the lowest achievable setting, while maintaining strong signal strength. Fish locations were recorded at these locations with a handheld Garmin GPSMAP 78SC (Garmin Ltd., Olathe, KS, USA). Prior individual fish locations were evaluated in the field, and when no movement occurred for a given fish between three or more tracking events and the radio tag would not move when disturbed or was recovered, the fish was considered dead. In these instances, the data were reviewed to determine the fish’s last live location [
34]. Due to the high detection rates observed in this study, survival was simply expressed using the percentage of fish known to survive during the duration of the study.
Dispersal and movement—Individual Muskellunge dispersal was calculated for the first 25 days poststocking (DPS). Only data collected within the first 25 DPS were used for dispersal analysis because most Muskellunge poststocking mortality occurs within this period [
29]. Dispersal was defined as the minimum straight-line, in-water distance (m) between the stocking site and the fish’s location during each tracking event. Since fish stocked at Spirit Lake were not stocked at a single location, individual fish dispersal was calculated as the minimum distance (m) from the fish’s location to the nearest point of the stocking transect (
Figure 1). Dispersal data for known alive fish were pooled by lake and DPS (11 tracking events at Spirit Lake; 5 tracking events at Lake Cornelia). Daily movement rates (m day
−1) for individual fish were calculated by dividing the minimum in-water, straight-line distance traversed between tracking events by the number of lapsed days. If the number of lapsed days between two consecutive individual fish detections was ≥14 days or the fish was determined to be dead, daily movement rates were excluded from the analyses. Daily movement rates observed within the first seven DPS were pooled by lake to examine mean movement rates within the first week poststocking. Subsequent mean daily movement rates were calculated and pooled by each week poststocking to examine the movement behavior of Muskellunge throughout the study.
Muskellunge proximity to shore, depth, and aquatic vegetation usage—Individual Muskellunge proximity (m) to shore was calculated by converting existing lake boundary polygons to polylines and then using the near analysis tool in ArcMap 10.7.1 to generate minimum distances between Muskellunge locations and lake boundaries. The exact depth of Muskellunge at observed locations could not be determined; instead, the maximum water depth at each observed Muskellunge location was recorded using a boat-mounted depth finder (LowranceTM Gen 2 HDS-7) and used as a proxy to classify Muskellunge depth usage. Once depth was recorded, the worker examined the aquatic plant signature displayed via the sonar unit to estimate the height of aquatic plants within the water column. This number was recorded and later converted to a percentage to calculate submersed aquatic vegetation (SAV) biovolume (plant height within the water column divided by maximum water depth) for each point where Muskellunge were located.
Data analysis—Fish dispersal, movement, proximity to shore, and depth were natural logarithm transformed, and an arcsine transformation was used for SAV biovolume data to meet data normality assumptions. The effects of lake, week poststocking, and the lake×week poststocking interaction term on Muskellunge movement, proximity to shore, depth usage, and SAV biovolume were determined using repeated-measures analysis of variance (ANOVA; with a first-order autoregressive covariance structure [
34]) with individual fish specified as the repeated variable. If differences in the main effects or interaction term were detected within a lake, Tukey’s multiple comparison test was used to determine where differences existed through time. Similarly, 25-day dispersal rates were examined within a lake using repeated-measure ANOVA, and differences in mean separation were compared using Tukey’s multiple comparison test. To determine if dispersal rates differed among lakes within the first 7 DPS, dispersal distances for individual fish were pooled by lake and DPS, and a t-test was used to compare dispersal rates among systems. All analyses were conducted within Statistical Analysis System (SAS Institute, Cary, NC, USA) software, and significance was set at α = 0.05.
3. Results
Survival—Muskellunge were detected 245 times (mean ± standard error [SE] = 16.3 ± 3.9 times fish
−1) during 45 tracking events at Spirit Lake and 156 times (mean ± SE = 15.6 ± 4.4 times fish
−1) during 17 tracking events at Lake Cornelia between 20 May 2023 and 8 September 2023 (
Table 1). Only three Muskellunge (two at Spirit Lake and one at Lake Cornelia) were confirmed dead by 8 September 2023. Of these, one fish (Fish 8.071) was not located after 12 DPS and went missing until 102 DPS when it was found deposited (no movement on five separate tracking events) in the middle of the northeast bay of Spirit Lake in 5.1 m of water. Fish 8.152 was first located 12 DPS, 1093 m from the stocking location in Spirit Lake, and had movement patterns consistent with a live fish up to 101 DPS. At this time, the fish had moved nearshore (within 25 m of shore), and the tag was subsequently recovered on the bank (approximately 10 m from shoreline) at 112 DPS. At Lake Cornelia, Fish 9.332 was located alive during each tracking event between 2 and 11 DPS in nearshore (<15 m of shoreline) habitats (e.g., docks with aquatic vegetation) but went missing during the tracking event on 17 DPS. A social media post confirmed the capture and harvest of a radio-tagged fish that was misidentified as a Northern Pike, and the fish was considered dead thereafter. All other radio-tagged Muskellunge displayed frequent movement and were considered alive at the end of the 111–112-day study. Therefore, observed Muskellunge survival was 86.7% (13 of 15 fish) in Spirit Lake and 90.0% (9 of 10 fish) in Lake Cornelia, with a cumulative survival rate of 88% (22 of 25 fish).
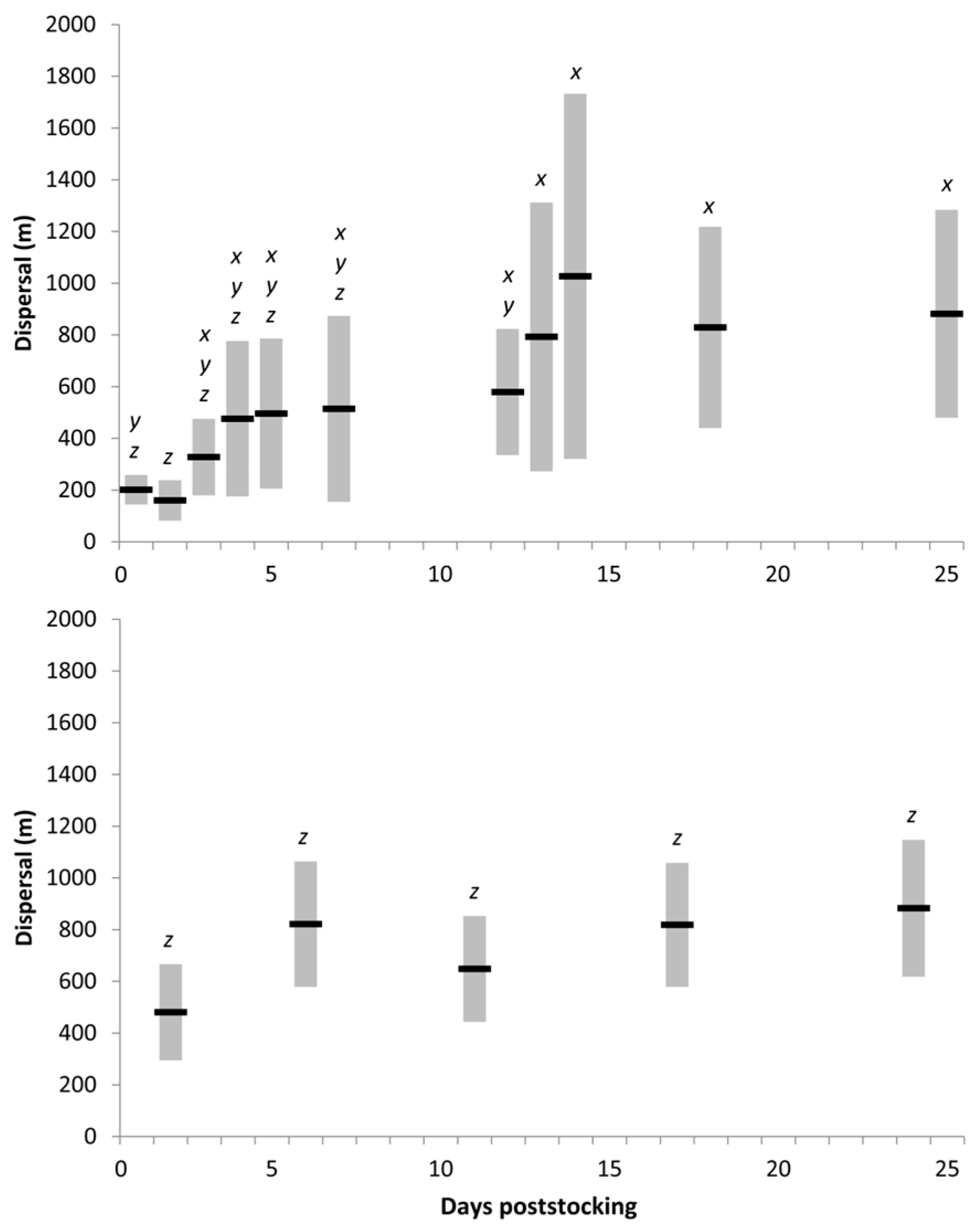
Dispersal and movement—Muskellunge dispersal with the first 25 DPS ranged from 55 to 2436 m (mean ± SE = 606 ± 41 m fish
−1). Muskellunge dispersed quickly after stocking, as fish were located more than 1090 m from stocking locations by 2 DPS (mean ± SE = 289 ± 44 m fish
−1), but dispersal was higher at Lake Cornelia than at Spirit Lake within the first week poststocking (
t = −2.78, pooled df = 78,
p = < 0.01). Muskellunge dispersal at Spirit Lake (
n = 104 observations; mean ± SE = 552 ± 552 m fish
−1) differed among DPS (
df = 10; F = 5.82;
p < 0.001), with increased dispersal rates observed at ≥13 DPS compared to those within the first 2 DPS (
p < 0.05;
Figure 2). Dispersal at Lake Cornelia (
n = 48 observations; mean ± SD = 725 ± 376 m fish
−1) did not differ among DPS (
df = 4; F = 1.16;
p 0.3429;
Figure 2).
Muskellunge mean daily movement ranged from 0.48 to 689.46 m day
−1 (mean ± SE = 81 ± 4.9 m day
−1) and was generally higher at Spirit Lake (mean ± SE = 92 ± 7.2 m day
−1) than at Lake Cornelia (mean ± SE = 64 ± 5.9 m day
−1). Muskellunge movement rates differed among weeks (
p < 0.001) but did not differ among lakes (
p = 0.745), and the lake × week interaction term was insignificant (
p = 0.147; see
F values,
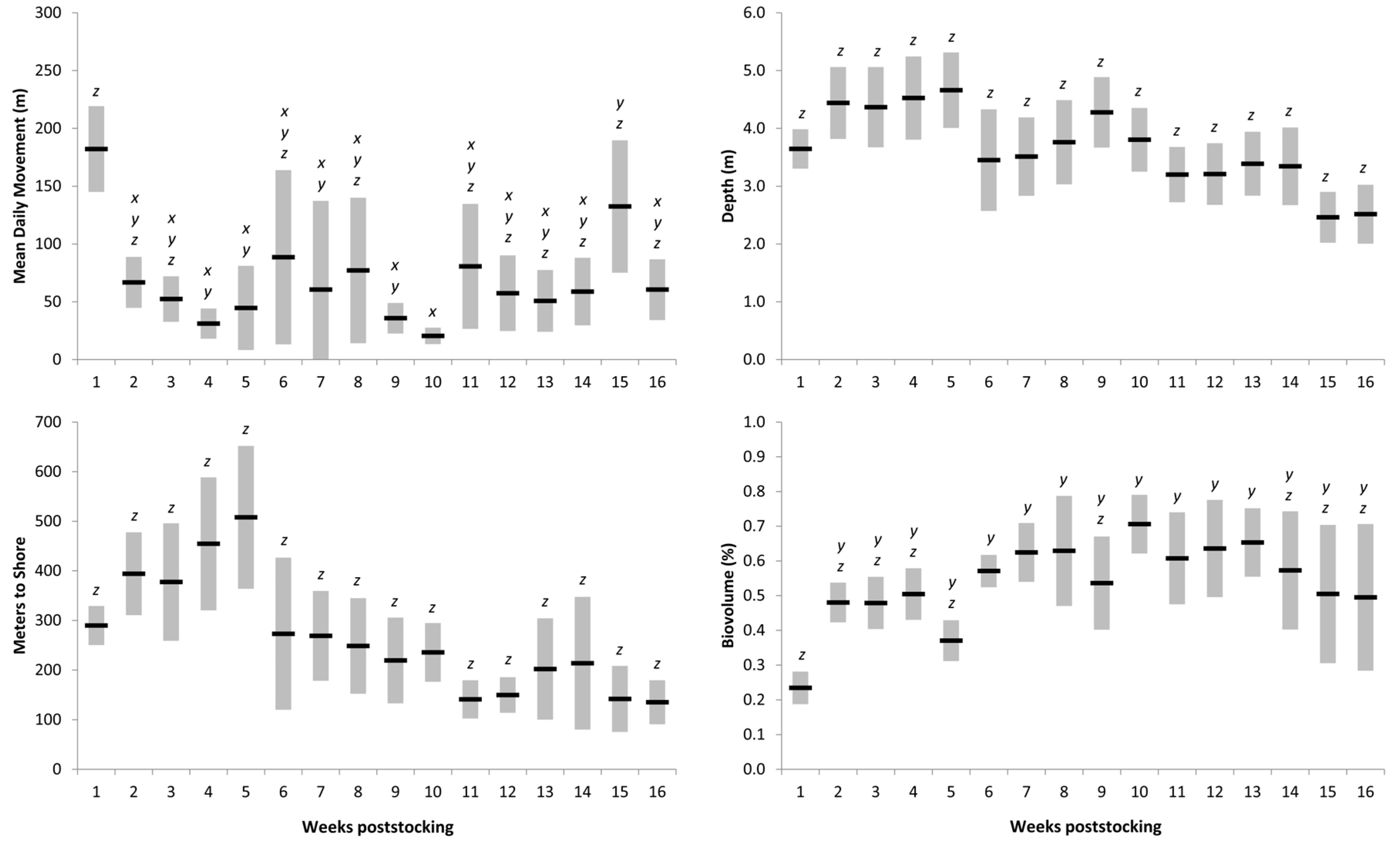
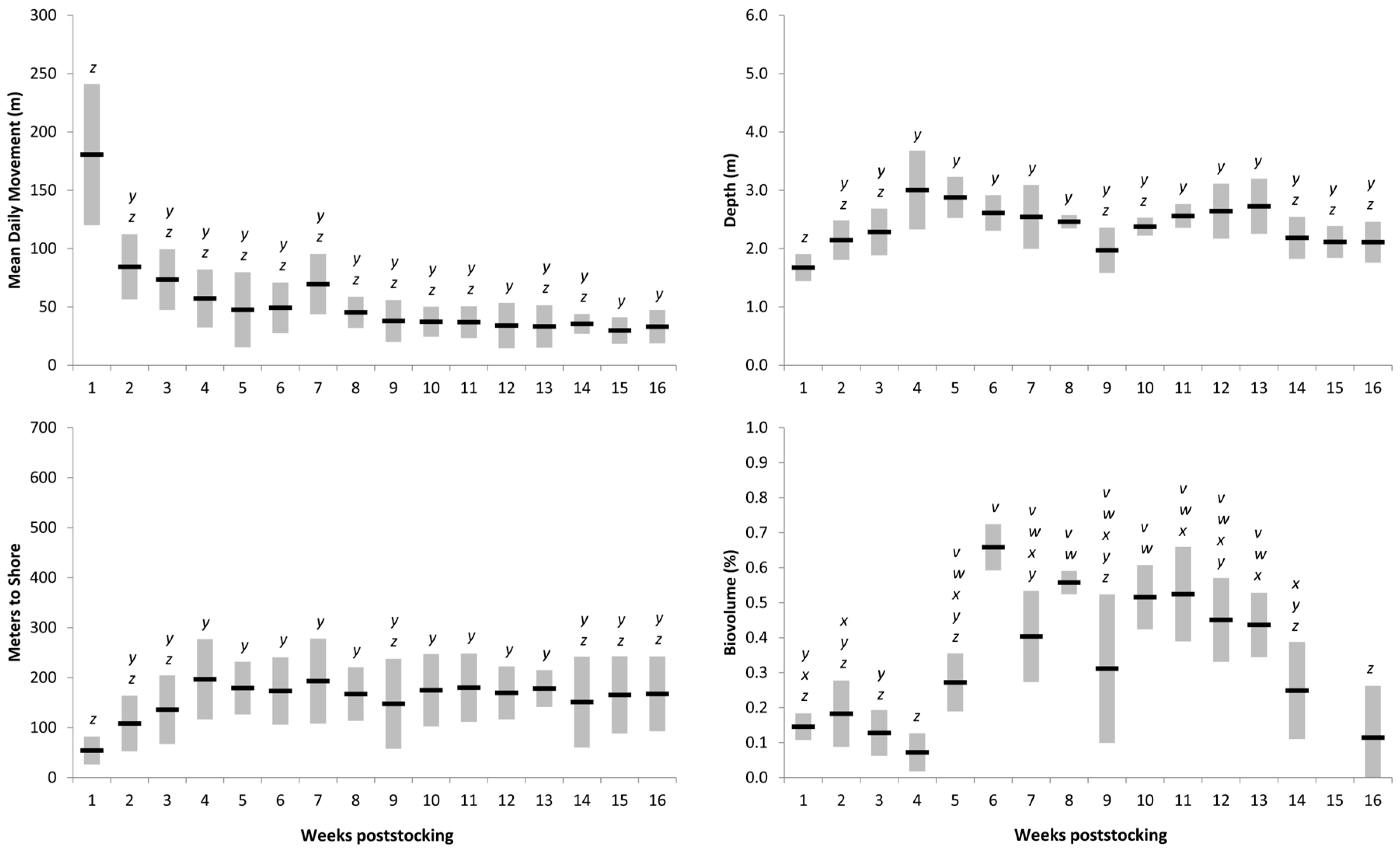
Table 2), indicating that Muskellunge movement behavior was similar among lakes through time. Mean daily movement was highest in both lakes within the first week poststocking, after which rates generally declined (
Figure 3 and
Figure 4).
Muskellunge proximity to shore, depth, and aquatic vegetation usage—Muskellunge were located in nearshore (<25 m from shore) habitats during 27 of 402 (6.7%) locations and frequently preferred offshore habitats in both lakes (mean ± SE = 226 ± 8.7 m). Muskellunge proximity to shore in Spirit Lake (mean ± SE = 273 ± 12.2 m) nearly doubled compared to that in Lake Cornelia (mean ± SE = 151 ± 8.5 m); however, the effect of lake and interaction between lake–weeks poststocking was not significant (
p > 0.05;
Table 2). Muskellunge proximity to shore differed among weeks poststocking (
p < 0.001;
Table 2). At Lake Cornelia, Muskellunge were closer to shore during the first week poststocking (mean ± SE = 54 ± 14 m) and then moved further from shore between weeks 1 and 4 (mean ± SE = 197 ± 41 m;
Figure 4). Muskellunge proximity to shore was similar between weeks 5 and 16. A similar pattern was observed at Spirit Lake where fish moved offshore between weeks 1 and 5 but then moved closer to shore between weeks 6 and 16 (
Figure 3). The depth of location of Muskellunge ranged between 0.15 and 6.3 m (mean ± SE = 3.17 ± 0.10 m) and was deeper for Muskellunge in Spirit Lake (mean ± SE = 3.69 ± 0.10 m) than in Lake Cornelia (mean ± SE = 2.34 ± 0.10 m). Muskellunge depth differed among weeks and lakes (
p < 0.02), but the lake × week interaction was insignificant (
p = 0.091;
Table 2). Muskellunge depth usage followed a similar pattern to proximity to shore as fish depth usage generally increased during weeks 1 to 5 and then either slightly decreased or was similar throughout the study (
Figure 3 and
Figure 4). Muskellunge in both systems used SAV throughout the study (337 of 366 locations had SAV), and biovolume locations ranged from 0% to 100% of the water column (mean ± standard deviation [SD] = 0.42 ± 0.25; no biovolume comparisons were conducted for week 15 due to equipment failure at Lake Cornelia). Biovolume of location differed among lakes and weeks, and the interaction effect was significant (
p < 0.001;
Table 2). Biovolume was higher in Spirit Lake (mean ± SD = 0.48 ± 0.25) than in Lake Cornelia (mean ± SD = 0.32 ± 0.23), but for both lakes, biovolume of location was less during the first 4 weeks poststocking than those weeks that followed. For both lakes, Muskellunge biovolume of location generally followed a pattern of increasing SAV usage between stocking and weeks 8–10 and then steadily declined throughout the remainder of the study.
4. Discussion
Agency stocking programs and adult Muskellunge populations would benefit from a better understanding of Muskellunge poststocking behavior and the survival of hatchery products. Telemetry studies may provide critical information regarding initial stocking success and factors that may be contributing to recruitment bottlenecks. This study documented that poststocking survival of large yearling Muskellunge in two natural lakes of differing biotic and abiotic characteristics was exceptionally high (86.7–90.0%). While others have documented substantial mortality resulting from terrestrial and fish predators [
5,
10], only three fish died in this study. The large size of yearling Muskellunge evaluated here likely contributed to high poststocking survival rates as fish size alone can reduce predation risks from other fish predators [
3,
4,
28,
29,
31]. These findings also were considerably different from previous studies conducted in Iowa as mortality resulting from avian and terrestrial predation alone accounted for 46.7% and 32.3% of known mortalities in Spirit Lake and Big Creek Reservoir, respectively [
6,
31]. Besides the Muskellunge size being substantially larger, Muskellunge predation via avian and other terrestrial predators in Spirit Lake may have been minimized in this study through stocking fish offshore, as previously observed during a short-term poststocking survival evaluation of smaller Muskellunge (325–386 mm; [
29]). For example, in this study, extensive curly-leaf pondweed beds (followed by abundant native submersed aquatic vegetation) were immediately available to yearling Muskellunge stocked offshore in deep (≥3 m) water habitats, and fish displayed high affinity (179 of 244 fish detections) to these areas throughout the telemetry experiment. Others have documented that young Muskellunge (wild or stocked) often prefer shallow vegetated habitats [
2,
5,
35,
36], but these environments can also have negative consequences since Muskellunge may become more susceptible to terrestrial predation [
6,
31], especially in clear water. In Spirit Lake, aquatic vegetation was abundant in both shallow and deep water habitats throughout the telemetry investigation, yet 96.3% of fish detections were in depths ≥1 m and ≥45 m from the shoreline, thus substantially limiting impacts of nearshore predator–prey interactions. Conversely, at Lake Cornelia, yearling Muskellunge stocked at a boat ramp had high survival rates, regardless of depth usage. However, we did not expect high predation at Lake Cornelia due to the recent fish renovation and fewer terrestrial predation risks there (Eric Rector, Wright County Conservation, personal communication). Nevertheless, this study did document that Muskellunge size as well as stocking practices may help reduce most short-term predation risks in both simple and complex natural lakes, and managers may need to consider both tools when developing stocking plans for managed Muskellunge fisheries.
The high survival of radio-tagged yearling Muskellunge during this study provided multiple observations of dispersal, movement patterns, and habitat usages at two very distinct natural lakes managed for Muskellunge for a period up to 112 DPS. Although several studies have examined poststocking Muskellunge movement and habitat usage via telemetry in rivers and impoundments across the managed range of this species [
5,
6,
10,
34,
37], little information exists for young Muskellunge poststocking behavior in natural lakes stocked in either the fall [
2] or spring [
29,
31]. Muskellunge stocked in the fall in two northern Wisconsin lakes dispersed rapidly after a short recovery period of 1 DPS and selected nearshore, shallow (<3.1 m), vegetated areas up to 34 DPS. Those stocked in the spring, however, varied substantially in their dispersal and movement patterns within the first 10 DPS, with fish stocked in May exhibiting lower dispersal rates than those stocked in late June [
29]. Dispersal of May-stocked fish, however, increased substantially after 11 DPS. It was hypothesized that dispersal and movement rates observed for May-stocked Muskellunge may be influenced by the availability of emergent and submersed aquatic vegetation at the time of stocking [
29]. In this study and for evaluations conducted in Spirit Lake for June-stocked yearling Muskellunge [
29], submersed aquatic vegetation at the time of stocking was extensive, and fish dispersal from stocking locations and movement rates were rapid. Those stocked in May, however, were associated more with emergent vegetation that was common near the stocking location, and observed dispersal rates were much lower [
29]. At Lake Cornelia, nearshore stocking of Muskellunge in habitats that were void of emergent vegetation had rapid dispersal and movement patterns. These observations may suggest that increased fish size at stocking may facilitate increased dispersal and movement patterns compared to smaller stocked counterparts. However, linear models relating fish size to mean first-week dispersal rates were inconclusive for radio-tagged Muskellunge in this study (
p > 0.28; r
2 < 0.20). In addition, Muskellunge stocked in May and June during 2020 that had large differences in 10 DPS dispersal rates were of similar size (mean of 336 and 343 mm [
29]). Thus, the propensity for stocked Muskellunge to disperse and exhibit large movement patterns seems to be more related to the timing of stocking and the abundance and type of aquatic vegetation residing at each stocking location. Correspondingly, these movement patterns substantially influence the location, depth, and aquatic plant biovolume usage frequency of Muskellunge throughout the experiment. In this study, it was well documented that Muskellunge were associated with deeper, vegetated habitats soon after stocking and most remained exclusively in these habitats throughout the entire telemetry experiment.
The results of this study indicate that Muskellunge yearling survival can be improved in natural lakes by stocking larger fish and validate previous models relating increased Muskellunge size at stocking to increased survival in the wild for fish ≥ 375 mm TL [
29]. These relationships as reported here and elsewhere have prompted increased agency interest in culture techniques that improve stocked fish size across the Midwest US. However, the ability to consistently produce hatchery-reared Muskellunge yearlings at sizes beyond those traditionally reared can be difficult. For example, some agencies have experimented with overwintering fingerling Muskellunge in earthen ponds but observed minimal overwinter growth (21.7 mm; mean total length [TL] of 267 mm) and suggested that the expense of live forage may become cost-prohibitive [
38]. The culture method used during this evaluation incorporated a hybrid cultivation technique that has not been used extensively in Iowa previously. However, consistent production of quality large yearling Muskellunge may require changes in cultivation techniques, duration of cultivation, and increased direct and indirect expenses that may inhibit cost-effective solutions. Thus, culture techniques that consistently produce large hatchery-produced Muskellunge continue to be explored in Iowa and elsewhere. Nonetheless, the results from this study provide fundamental information regarding stocked Muskellunge survival, behavior, and habitat usage that can be used as a framework in future evaluations of hatchery-reared Muskellunge in natural lakes.



{kind=link}
{kind=link}
{kind=link}
{kind=link}