
Assessing the Conservation Value of Artificial and Natural Reefs via Ichthyoplankton Spatio-Temporal Dynamics

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
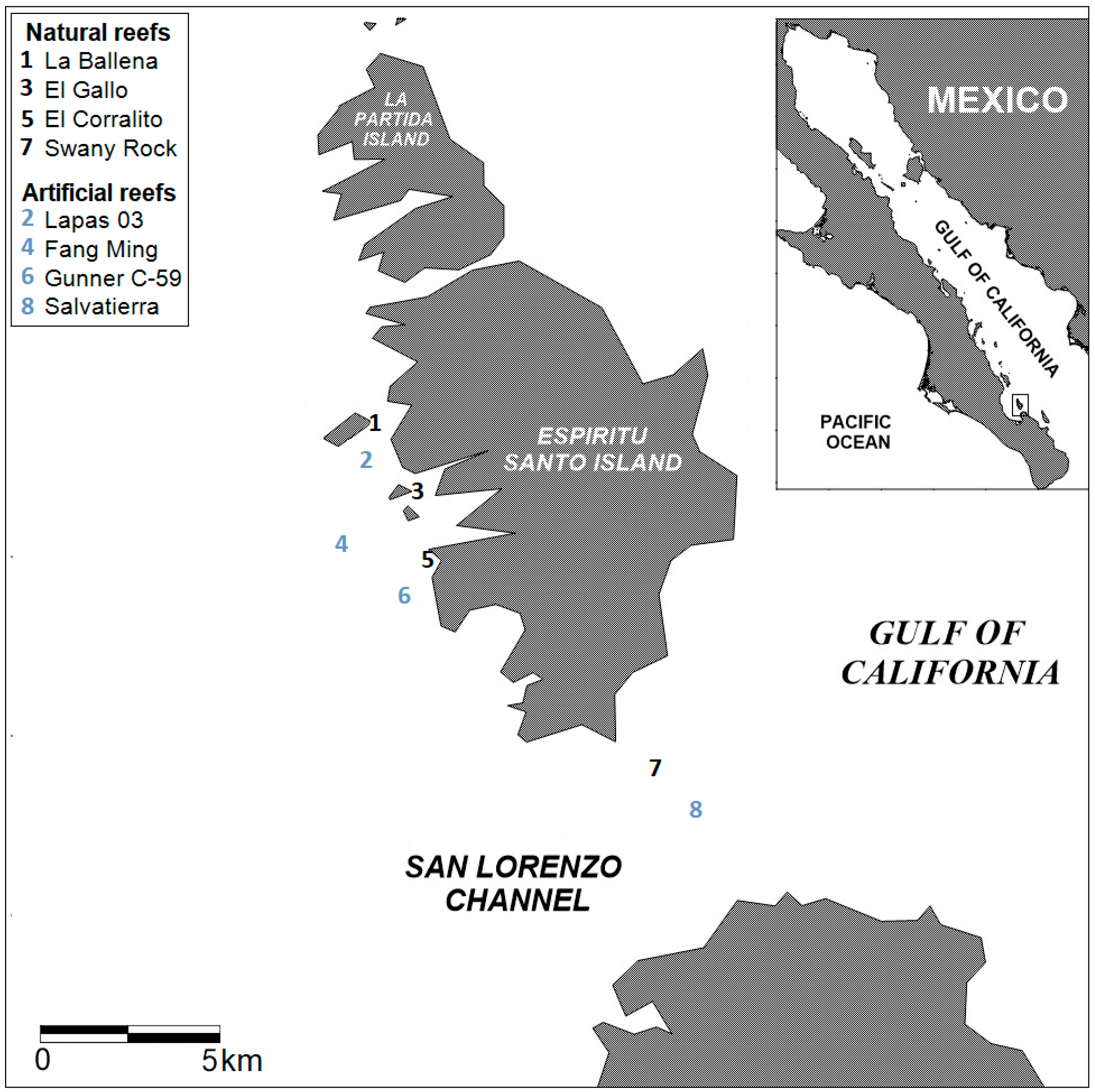
2.1. Study Area
2.2. Sampling Procedure In Situ
2.3. Sampling Procedure in Laboratory
2.4. Breeding Habitat for Reef Fish
2.5. Statistical Analysis
3. Results
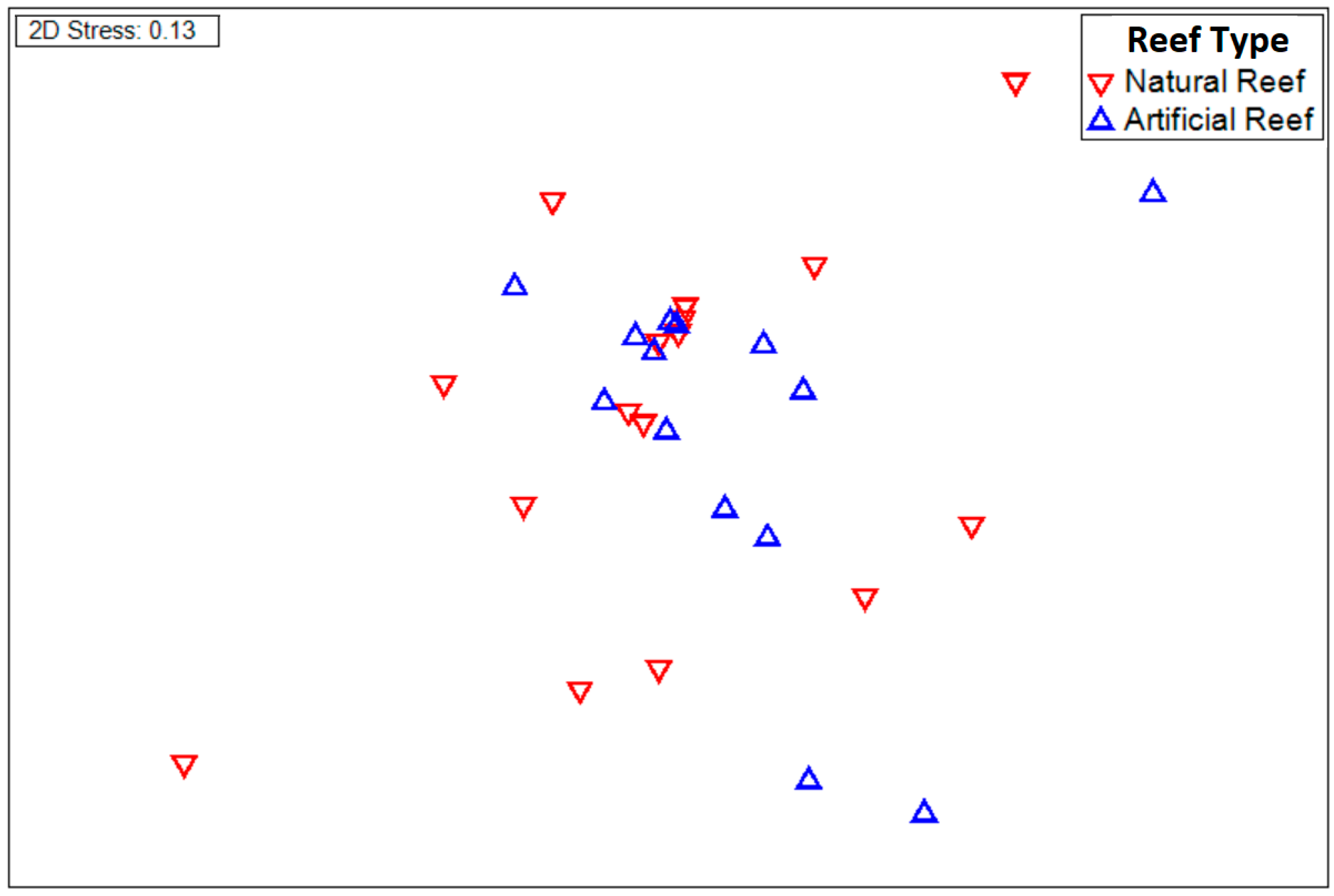
3.1. Egg and Larva Distribution
3.2. Egg Development Stage
3.3. Seasonality (Warm Season vs. Cold Season)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Leis, J.M. What does larval fish biology tell us about the design and efficacy of Marine Protected Areas. In Aquatic Protected Areas: What Works Best and How Do We Know, Proceedings of the World Congress on Aquatic Protected Areas, Cairns, Australia; Australian Society for Fish Biology: Newcastle, Australia, 2003; pp. 170–180. [Google Scholar]
- Félix-Hackradt, F.C.; Hackradt, C.W.; Treviño-Otón, J.; Segovia-Viadero, M.; Pérez-Ruzafa, A.; García-Charton, J.A. Environmental determinants on fish post-larval distribution in coastal areas of south-western Mediterranean Sea. Estuar. Coast. Shelf Sci. 2013, 129, 59–72. [Google Scholar] [CrossRef]
- Green, A.L.; Maypa, A.P.; Almany, G.R.; Rhodes, K.L.; Weeks, R.; Abesamis, R.A.; Gleason, M.G.; Mumby, P.J.; White, A.T. Larval dispersal and movement patterns of coral reef fishes, and implications for marine reserve network design. Biol. Rev. 2015, 90, 1215–1247. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.D.; Possingham, H.P.; Muelbert, J.H. Incorporating early life stages of fishes into estuarine spatial conservation planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 1013–1030. [Google Scholar] [CrossRef]
- Carter, A.B.; Davies, C.R.; Emslie, M.J.; Mapstone, B.D.; Russ, G.R.; Tobin, A.J.; Williams, A.J. Reproductive benefits of no-take marine reserves vary with region for an exploited coral reef fish. Sci. Rep. 2017, 7, 9693. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, B. Life History and Early Development of Fishes. In Biology of Fishery Resources; Springer: Singapore, 2022; pp. 55–69. [Google Scholar] [CrossRef]
- Fortier, L.; Leggett, W.C. Fickian Transport and the Dispersal of Fish Larvae in Estuaries. Can. J. Fish. Aquat. Sci. 1982, 39, 1150–1163. [Google Scholar] [CrossRef]
- Norcross, B.L.; Shaw, R.F. Oceanic and estuarine transport of fish eggs and larvae: A review. Trans. Am. Fish. Soc. 1984, 113, 153–165. [Google Scholar] [CrossRef]
- Pattrick, P.; Weidberg, N.; Goschen, W.S.; Jackson, J.M.; McQuaid, C.D.; Porri, F. Larval fish assemblage structure at coastal fronts and the influence of environmental variability. Front. Ecol. Evol. 2021, 347, 684502. [Google Scholar] [CrossRef]
- Counsell, C.W.W.; Coleman, R.R.; Lal, S.S.; Bowen, B.W.; Franklin, E.C.; Neuheimer, A.B.; Powell, B.; Toonen, R.; Donahue, M.; Hixon, M.; et al. Interdisciplinary analysis of larval dispersal for a coral reef fish: Opening the black box. Mar. Ecol. Prog. Ser. 2022, 684, 117–132. [Google Scholar] [CrossRef]
- Garcés-Rodríguez, Y.; Sánchez-Velasco, L.; Parés-Sierra, A.; Jiménez-Rosenberg, S.P.A.; Tenorio-Fernández, L.; Montes-Aréchiga, J.; Godínez-Sandoval, V. Distribution and transport of Fish larvae at the entrance of the Gulf of California (September, 2016). Deep Sea Res. Part I Oceanogr. Res. Pap. 2022, 193, 103957. [Google Scholar] [CrossRef]
- Armstrong, M.J.; Witthames, P.R. Developments in understanding of fecundity of fish stocks in relation to egg production methods for estimating spawning stock biomass. Fish. Res. 2012, 117, 35–47. [Google Scholar] [CrossRef]
- Stige, L.C.; Yaragina, N.A.; Langangen, Ø.; Bogstad, B.; Stenseth, N.C.; Ottersen, G. Effect of a fish stock’s demographic structure on offspring survival and sensitivity to climate. Proc. Natl. Acad. Sci. USA 2017, 114, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Lowerre-Barbieri, S.K. Reproduction in relation to conservation and exploitation of marine fishes. In Reproductive Biology and Phylogeny of Fishes (Agnathans and Bony Fishes); CRC Press: Boca Raton, FL, USA, 2019; pp. 371–394. [Google Scholar]
- Somarakis, S.; Tsoukali, S.; Giannoulaki, M.; Schismenou, E.; Nikolioudakis, N. Spawning stock, egg production and larval survival in relation to small pelagic fish recruitment. Mar. Ecol. Prog. Ser. 2019, 617, 113–136. [Google Scholar] [CrossRef]
- Aceves-Medina, G.; Emilio, A.; Inda-Díaz, O.; Ubisha-Hernández, A.; Álvarez-Ramírez, I. Review of some aspects of the early life history of marine fish and their implications in the biogeographic distribution. CICIMAR Ocean 2020, 35, 1–18. [Google Scholar]
- Mitterwallner, P.; Shima, J.S. The relative influence of environmental cues on reproductive allocation of a highly iteroparous coral reef fish. Coral Reefs 2022, 41, 1323–1335. [Google Scholar] [CrossRef]
- Colin, P.L. Aggregation Spawning: Biological Aspects of the Early Life History. In Reef Fish Spawning Aggregations: Biology, Research and Management; Sadovy de Mitcheson, Y., Colin, P., Eds.; Fish & Fisheries Series; Springer: Dordrecht, The Netherlands, 2012; Volume 35. [Google Scholar] [CrossRef]
- Hamner, W.M.; Largier, J.L. Oceanography of the planktonic stages of aggregation spawning reef fishes. In Reef Fish Spawning Aggregations: Biology, Research and Management; Springer: Dordrecht, The Netherlands, 2012; pp. 159–190. [Google Scholar]
- Teodosio, M.A.; Paris, C.B.; Wolanski, E.; Morais, P. Biophysical processes leading to the ingress of temperate fish larvae into estuarine nursery areas: A review. Estuar. Coast. Shelf Sci. 2016, 183, 187–202. [Google Scholar] [CrossRef]
- Stephens, J., Jr.; Pondella, D. Larval productivity of a mature artificial reef: The ichthyoplankton of King Harbor, California, 1974–1997. ICES J. Mar. Sci. 2002, 59, S51–S58. [Google Scholar] [CrossRef]
- Nishimoto, M.M.; Simons, R.D.; Love, M.S. Offshore oil production platforms as potential sources of larvae to coastal shelf regions off southern California. Bull. Mar. Sci. 2019, 95, 535–558. [Google Scholar] [CrossRef]
- Alegretti, C.B.; Grande, H.; Namiki, C.A.P.; Loose, R.H.; Brandini, F.P. A Preliminary assessment of larval fish assemblages on artificial reefs in the nearshore Southern Brazil. Ocean Coast. Res. 2021, 69, e21017. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, H.; Yu, D.; Song, J.; Duan, G. The migration of viscous fish eggs in artificial reefs. Ecol. Model. 2022, 469, 109985. [Google Scholar] [CrossRef]
- Kraufvelin, P.; Bergström, L.; Sundqvist, F.; Ulmestrand, M.; Wennhage, H.; Wikström, A.; Bergström, U. Rapid re-establishment of top-down control at a no-take artificial reef. Ambio 2022, 52, 556–570. [Google Scholar] [CrossRef]
- Contreras, T.; Olivar, M.P.; Hulley, P.A.; Fernández de Puelles, M.L. Feeding ecology of early life stages of mesopelagic fishes in the equatorial and tropical Atlantic. ICES J. Mar. Sci. 2019, 76, 673–689. [Google Scholar] [CrossRef]
- Stukel, M.R.; Gerard, T.; Kelly, T.; Knapp, A.N.; Laiz-Carrión, R.; Lamkin, J.; Landry, M.R.; Malca, E.; Selph, K.E.; Shiroza, A.; et al. Plankton food webs of the Gulf of Mexico spawning grounds of Atlantic Bluefin tuna. bioRxiv 2020. [Google Scholar] [CrossRef]
- Shiroza, A.; Malca, E.; Lamkin, J.T.; Trika, G.; Landry, M.R.; Stukel, M.R.; Laiz-Carrión, R.; Swalethorp, R. Active prey selection in developing larvae of Atlantic bluefin tuna (Thunnus thynnus) in spawning grounds of the Gulf of Mexico. J. Plankton Res. 2022, 44, 728–746. [Google Scholar] [CrossRef]
- El-Regal, M.A. Spawning seasons, spawning grounds and nursery grounds of some Red Sea fishes. Glob. J. Fish. Aqua. 2013, 6, 105–125. [Google Scholar]
- Reglero, P.; Ortega, A.; Balbín, R.; Abascal, F.J.; Medina, A.; Blanco, E.; de la Gándara, F.; Alvarez-Berastegui, D.; Hidalgo, M.; Rasmuson, L.; et al. Atlantic bluefin tuna spawn at suboptimal temperatures for their offspring. Proc. R. Soc. B Biol. Sci. 2018, 285, 20171405. [Google Scholar] [CrossRef]
- Granneman, J.E.; Steele, M.A. Fish growth, reproduction, and tissue production on artificial reefs relative to natural reefs. ICES J. Mar. Sci. 2014, 71, 2494–2504. [Google Scholar] [CrossRef]
- Yin, R.; Jiang, R.; Bi, Y.; Zhang, L.; Fan, Z.; Chen, L.; Sui, Y.; Wang, S. Community structure of fish eggs and larvae in artificial reef area of Ma’an Archipelago. J. Fish. China 2019, 43, 1937–1951. [Google Scholar]
- Pickering, H.; Whitmarsh, D. Artificial reefs and fisheries exploitation: A review of the ‘attraction versus production’ debate, the influence of design and its significance for policy. Fish. Res. 1997, 31, 39–59. [Google Scholar] [CrossRef]
- Brickhill, M.J.; Lee, S.Y.; Connolly, R.M. Fishes associated with artificial reefs: Attributing changes to attraction or production using novel approaches. J. Fish Biol. 2005, 67, 53–71. [Google Scholar] [CrossRef]
- Layman, C.A.; Allgeier, J.E. An ecosystem ecology perspective on artificial reef production. J. Appl. Ecol. 2020, 57, 2139–2148. [Google Scholar] [CrossRef]
- Whitmarsh, D.; Santos, M.N.; Ramos, J.; Monteiro, C.C. Marine habitat modification through artificial reefs off the Algarve (southern Portugal): An economic analysis of the fisheries and the prospects for management. Ocean Coast. Manag. 2008, 51, 463–468. [Google Scholar] [CrossRef]
- Kramer, S.H.; Hamilton, C.D.; Spencer, G.C.; Ogston, H.O. Evaluating the Potential for Marine and Hydrokinetic Devices to Act as Artificial Reefs or Fish Aggregating Devices. Based on Analysis of Surrogates in Tropical, Subtropical, and Temperate US West Coast and Hawaiian Coastal Waters; No. DOE-HTH-0006389; HT Harvey & Associates: Honolulu, HI, USA, 2015. [Google Scholar]
- Smith, J.A.; Lowry, M.B.; Champion, C.; Suthers, I.M. A designed artificial reef is among the most productive marine fish habitats: New metrics to address ‘production versus attraction’. Mar. Biol. 2016, 163, 188. [Google Scholar] [CrossRef]
- Abelson, A. Artificial reefs vs coral transplantation as restoration tools for mitigating coral reef deterioration: Benefits, concerns, and proposed guidelines. Bull. Mar. Sci. 2006, 78, 151–159. [Google Scholar]
- Komyakova, V.; Chamberlain, D.; Jones, G.P.; Swearer, S.E. Assessing the performance of artificial reefs as substitute habitat for temperate reef fishes: Implications for reef design and placement. Sci. Total Environ. 2019, 668, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Kramer, S.H.; Hamilton, C.D.; Spencer, G.C.; Ogston, H.O. Collecting and Processing Data on Fish Eggs and Larvae in the California Current Region; NOAA Technical Report NMFS CIRC-370; National Marine Fisheries Service: Silver Spring, MD, USA, 1972; Volume 370, 38p. Available online: https://repository.library.noaa.gov/view/noaa/3248 (accessed on 1 December 2022).
- Jiménez-Rosenberg, S.P.A. Desarrollo Larvario de Eucinostomus cufrani Yáñez-Arancibia, 1978 (Pistes: Gerreidae). Ph.D. Thesis, Universidad Autónoma de Baja California Sur, La Paz, Mexico, 1994. [Google Scholar]
- Jiménez-Rosenberg, S.P.A. Descripción del Desarrollo Larvario de Eucinostomus gracilis y Larvario y Juvenil de Eucinostomus dowii y Diapterus peruvianus. Ph.D. Thesis, Depto. de Plancton y Ecología Marina, CICIMAR, La Paz, Mexico, 1998. [Google Scholar]
- Moser, H.G. Chauliodontidae: Viperfishes. In The Early Stages of Fishes in the California Current Region; Moser, H.G., Ed.; California Cooperative Oceanic Fisheries Investigations (CalCOFI) Atlas No. 33; Allen Press: Lawrence, KS, USA, 1996; pp. 297–299. 1505p. [Google Scholar]
- Richards, W.J. Early Stages of Atlantic Fishes: An Identification Guide for the Western Central North Atlantic; CRC Press: Boca Ratón, FL, USA, 2006. [Google Scholar]
- Ahlstrom, E.H. Influence of the Temperature on the Rate of Development of Pilchard Eggs in Nature. In Studies on the Pacific Pilchard or Sardine (Sardinops caerulaea); Special Scientific Report; U.S. Fish and Wildlife Service: Washington, DC, USA, 1943; Volume 23, pp. 1–26. [Google Scholar]
- Ahlstrom, E.H.; Moser, H.G. Eggs and larvae of fishes and their role in systematic investigatios and in fisheries. Rev. Des Trav. De L’institut Des Pêches Marit. 1976, 40, 379–398. [Google Scholar]
- Kendall, A.W.; Ahlstrom, E.H.; Moser, H.G. Early Life History Stages of Fishes and Their Characters. In Ontogeny and Systematics of Fishes; Moser, H.G., Ed.; American Society of Ichthyologists and Herptetologists: Glen Allen, VA, USA, 1984; Volume 1, pp. 11–22. Available online: https://swfsc-publications.fisheries.noaa.gov/publications/CR/1984/8447.PDF (accessed on 1 December 2021).
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Wasserman, R.J.; Matcher, G.F.; Vink, T.J.; Froneman, P.W. Preliminary evidence for the organisation of a bacterial community by zooplanktivores at the top of an estuarine planktonic food web. Microb. Ecol. 2015, 69, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Grol, M.G.; Rypel, A.L.; Nagelkerken, I. Growth potential and predation risk drive ontogenetic shifts among nursery habitats in a coral reef fish. Mar. Ecol. Prog. Ser. 2014, 502, 229–244. [Google Scholar] [CrossRef]
- Brogan, M.W. Distribution and retention of larval fishes near reefs in the Gulf of California. Mar. Ecol. Prog. Ser. 1994, 115, 1–13. [Google Scholar] [CrossRef]
- Ahern, A.L.M.; Gómez-Gutiérrez, J.; Aburto-Oropeza, O.; Saldierna-Martínez, R.J.; Johnson, A.F.; Harada, A.E.; Sánchez-Uvera, A.R.; Erisman, B.; Arvizú, D.I.C.; Burton, R.S. DNA sequencing of fish eggs and larvae reveals high species diversity and seasonal changes in spawning activity in the southeastern Gulf of California. Mar. Ecol. Prog. Ser. 2018, 592, 159–179. [Google Scholar] [CrossRef]
- Sabatés, A.; Olivar, M.P.; Salat, J.; Palomera, I.; Alemany, F. Physical and biological processes controlling the distribution of fish larvae in the NW Mediterranean. Prog. Oceanogr. 2007, 74, 355–376. [Google Scholar] [CrossRef]
- Le Port, A.; Montgomery, J.C.; Smith, A.N.H.; Croucher, A.E.; McLeod, I.M.; Lavery, S.D. Temperate marine protected area provides recruitment subsidies to local fisheries. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171300. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Hock, K.; Krueck, N.C.; Tyazhelnikov, V.; Mumby, P.J. An MPA design approach to benefit fisheries: Maximising larval export and minimising redundancy. Diversity 2021, 13, 586. [Google Scholar] [CrossRef]
- Lima, J.S.; Sanchez-Jerez, P.; dos Santos, L.N.; Zalmon, I.R. Could artificial reefs increase access to estuarine fishery resources? Insights from a long-term assessment. Estuar. Coast. Shelf Sci. 2020, 242, 106858. [Google Scholar] [CrossRef]
- da Costa, I.D.; da Silva Santos, J.L.; Costa, L.L.; Lima, J.S.; Zalmon, I.R. Reproductive potential and production role of artificial reefs-Southeastern Brazil. Estuar. Coast. Shelf Sci. 2022, 265, 107710. [Google Scholar] [CrossRef]
- Leis, J.M.; Fisher, R. Swimming speed of settlement-stage reef-fish larvae measured in the laboratory and in the field: A comparison of critical speed and in situ speed. In Proceedings of the 10th International Coral Reef Symposium, Okinawa, Japan, 28 June–3 July 2006; pp. 438–445. [Google Scholar]
- Barth, P.; Berenshtein, I.; Besson, M.; Roux, N.; Parmentier, E.; Banaigs, B.; Lecchini, D. From the ocean to a reef habitat: How do the larvae of coral reef fishes find their way home? A state of art on the latest advances. Vie Milieu 2015, 65, 91–100. [Google Scholar]
- Fisher, R. Swimming speeds of larval coral reef fishes: Impacts on self-recruitment and dispersal. Mar. Ecol. Prog. Ser. 2005, 285, 223–232. [Google Scholar] [CrossRef]
- Gerlach, G.; Artema, J. The use of chemical cues in habitat recognition and settlement. In Chemical Ecology in Aquatic Systems; Oxford University Press: Oxford, UK, 2012; pp. 72–89. [Google Scholar]
- Huijbers, C.M.; Nagelkerken, I.; Lössbroek, P.A.; Schulten, I.E.; Siegenthaler, A.; Holderied, M.W.; Simpson, S.D.A. Test of the senses: Fish select novel habitats by responding to multiple cues. Ecology 2012, 93, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Caldarone, E.M.; Chícharo, M.A.; Clemmesen, C.; Faria, A.M.; Faulk, C.; Folkvord, A.; Holt, G.J.; Høie, H.; Kanstinger, P.; et al. On the edge of death: Rates of decline and lower thresholds of biochemical condition in food-deprived fish larvae and juveniles. J. Mar. Syst. 2012, 93, 11–24. [Google Scholar] [CrossRef]
- China, V.; Holzman, R. Hydrodynamic starvation in first-feeding larval fishes. Proc. Natl. Acad. Sci. USA 2014, 111, 8083–8088. [Google Scholar] [CrossRef]
- Thomson, D.A.; Findley, L.T.; Kerstitch, A.N. Reef Fishes of the Sea of Cortez: The Rocky-Shore Fishes of the Gulf of California, Revised ed.; University of Texas Press: Austin, TX, USA, 2000. [Google Scholar]
- Farach, E.B.; López, J.; García, R.; Nevárez, M.O.; Lluch, D.B.; Ortega, S. Temporal Variability of Oceanic Mesoscale Events in the Gulf of California. Remote Sens. 2021, 13, 1774. [Google Scholar] [CrossRef]
- Pantoja, D.A.; Marinone, S.G.; Parés-Sierra, A.; Gómez-Valdivia, F. Numerical modeling of seasonal and mesoscale hydrography and circulation in the Mexican Central Pacific. Cienc. Mar. 2012, 38, 363–379. [Google Scholar] [CrossRef]
- López-Martínez, J.; Farach Espinoza, E.B.; Herrera-Cervantes, H.; García Morales, R. Long-Term Variability in Sea Surface Temperature and Chlorophyll a Concentration in the Gulf of California. Remote Sens. 2023, 15, 4088. [Google Scholar] [CrossRef]
- Aceves-Medina, G.; Emilio, A.; Inda-Díaz, O.; Ubisha-Hernández, A.; Álvarez-Ramírez, I. Fish larvae assemblages in the Gulf of California. J. Fish Biol. 2004, 65, 832–847. [Google Scholar] [CrossRef]
- González-Armas, R.; Funes-Rodríguez, R.; Amador-Buenrostro, A. Larval fish community structure in a seamount of the Gulf of California. Hidrobiológica 2008, 18, 77–88. [Google Scholar]
- Sánchez-Velasco, L.; Jimenéz-Rosenberg, S.P.; Shirasago, B.; Obeso-Nieblas, M. Distribution and abundance of fish larvae in the Bahia de La Paz (Gulf of Califórnia) and their relation to hidrographic variability during Summer (1997–1998). Deep Sea Res. 2004, 51, 723–737. [Google Scholar] [CrossRef]
- Contreras-Catala, F.; Beier, E.J.; Sánchez-Velasco, L.; Godínez, V.M.; Sánchez-Pérez, E.D.; Barton, E.D. Water masses and larval fish habitats in the Pacific tropical-subtropical convergence off Mexico. Cont. Shelf Res. 2021, 230, 10457. [Google Scholar] [CrossRef]
- Lavín, M.F.; Castro, R.; Beier, E.; Godínez, V.M. Mesoscale eddies in the southern Gulf of California during summer: Characteristics and interaction with the wind stress. J. Geophys. Res. Ocean. 2013, 118, 1367–1381. [Google Scholar] [CrossRef]
- Herrera-Cervantes, H. Sea surface temperature, ocean color and wind forcing patterns in the Bay of La Paz, Gulf of California: Seasonal variability. Atmósfera 2019, 32, 25–38. [Google Scholar] [CrossRef]
- Higgins, E.; Metaxas, A.; Scheibling, R.E. A systematic review of artificial reefs as platforms for coral reef research and conservation. PLoS ONE 2022, 17, e0261964. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Species | Average Similarity | Average Dissimilarity | ||||
|---|---|---|---|---|---|---|
| Natural Reefs | Artificial Reefs | Warm Season | Cold Season | Natural vs. Artificial Reefs | Warm vs. Cold Seasons | |
| Thalassoma lucasanum | 18.17 | 15.9 | 16.39 | 19.06 | 7.97 | 8.01 |
| Prionotus stephanophrys | 11.36 | 5.52 | 7.37 | 12.02 | 4.96 | 4.27 |
| Vinciguerria lucetia | 5.17 | - | - | 12.69 | 4.4 | 1.88 |
| Fistularia corneta | - | - | - | 9.56 | 3.45 | 1.68 |
| Synodus sp. 3 | - | 6.8 | - | 6.25 | 3.13 | 1.73 |
| Synodus sp. 2 | 4.35 | - | - | - | 2.93 | 1.59 |
| Synodus sp. 1 | 4.19 | 5.28 | 5 | - | 2.78 | 2.46 |
| Scarus sp. 4 | - | - | - | - | 2.76 | 1.45 |
| Scomber japonicus | - | - | - | - | 2.69 | - |
| Scarus sp. 3 | - | 4.76 | - | - | 2.31 | 4.03 |
| Diodon holocanthus | - | - | - | 5.75 | 2.18 | 1.3 |
| Benthosema panamense | - | - | - | - | 2.07 | 1.37 |
| Synodus sp. 4 | - | 3.67 | - | - | 2.02 | 1.93 |
| Auxis rochei | - | - | 5 | - | 1.98 | 4.15 |
| Anisotremus davidsonni | 6.66 | - | 4.8 | - | 1.85 | 2.71 |
| Synodus sp. 5 | - | - | 0 | - | 1.66 | 1.23 |
| Caranx caballus | - | - | 0 | - | 1.59 | 1.69 |
| Harengula thrissina | - | - | 10.3 | - | 1.57 | - |
| Pseudopeneus grandiscuamis | - | - | - | - | 1.53 | 2.2 |
| Antennarius sp. 1 | - | 6.77 | - | - | 1.52 | - |
| Auxis thazard | - | - | - | - | 1.51 | - |
| Achirus mazatlanus | - | - | - | - | 1.45 | - |
| Hypsoblennius gentilis | - | - | - | - | 1.34 | - |
| Fistularia comersonii | 8.26 | 13.41 | 14.57 | - | 1.29 | 6.19 |
| Lutjanus sp. 1 | - | - | - | - | 1.28 | - |
| Scarus sp. 1 | - | - | - | - | 1.12 | - |
| Harengula thrissina | 5.71 | 9.64 | - | - | - | 5.11 |
| Antennarius sp. 1 | - | - | 6.5 | - | - | 4.02 |
| Abudefduf troschelii | - | - | - | - | - | 1.74 |
| Mugil curema | - | - | - | - | - | 1.68 |
| Coryphaena hippurus | - | - | - | - | - | 1.23 |
| Diapterus peruvianus | - | - | - | - | - | 1.21 |
| Total value of similarity/dissimilarity | 22.90 | 21.57 | 31.77 | 12.19 | 77.80 | 88.91 |
| MVDISP Value | 0.933 | 1.05 | 1.314 | 0.676 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Caballero, C.A.; Borges-Souza, J.M.; Saldierna-Martínez, R.J.; Abelson, A. Assessing the Conservation Value of Artificial and Natural Reefs via Ichthyoplankton Spatio-Temporal Dynamics. Fishes 2024, 9, 166. https://doi.org/10.3390/fishes9050166
Sánchez-Caballero CA, Borges-Souza JM, Saldierna-Martínez RJ, Abelson A. Assessing the Conservation Value of Artificial and Natural Reefs via Ichthyoplankton Spatio-Temporal Dynamics. Fishes. 2024; 9(5):166. https://doi.org/10.3390/fishes9050166
Chicago/Turabian StyleSánchez-Caballero, Carlos Adrián, José Manuel Borges-Souza, Ricardo Javier Saldierna-Martínez, and Avigdor Abelson. 2024. "Assessing the Conservation Value of Artificial and Natural Reefs via Ichthyoplankton Spatio-Temporal Dynamics" Fishes 9, no. 5: 166. https://doi.org/10.3390/fishes9050166
APA StyleSánchez-Caballero, C. A., Borges-Souza, J. M., Saldierna-Martínez, R. J., & Abelson, A. (2024). Assessing the Conservation Value of Artificial and Natural Reefs via Ichthyoplankton Spatio-Temporal Dynamics. Fishes, 9(5), 166. https://doi.org/10.3390/fishes9050166