
Chemical Cues Released by Predators’ Consumption of Heterospecific Prey Alter the Embryogenesis of Zebrafish

 and
and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
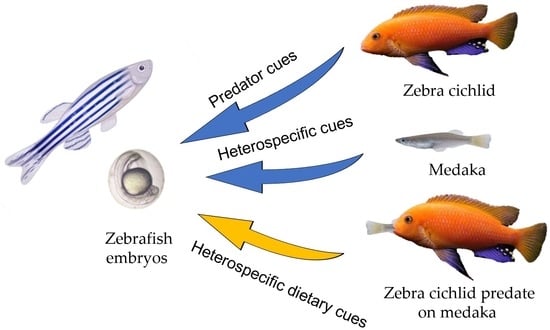
1. Introduction
2. Materials and Methods
2.1. Animals, Housing, and Maintenance
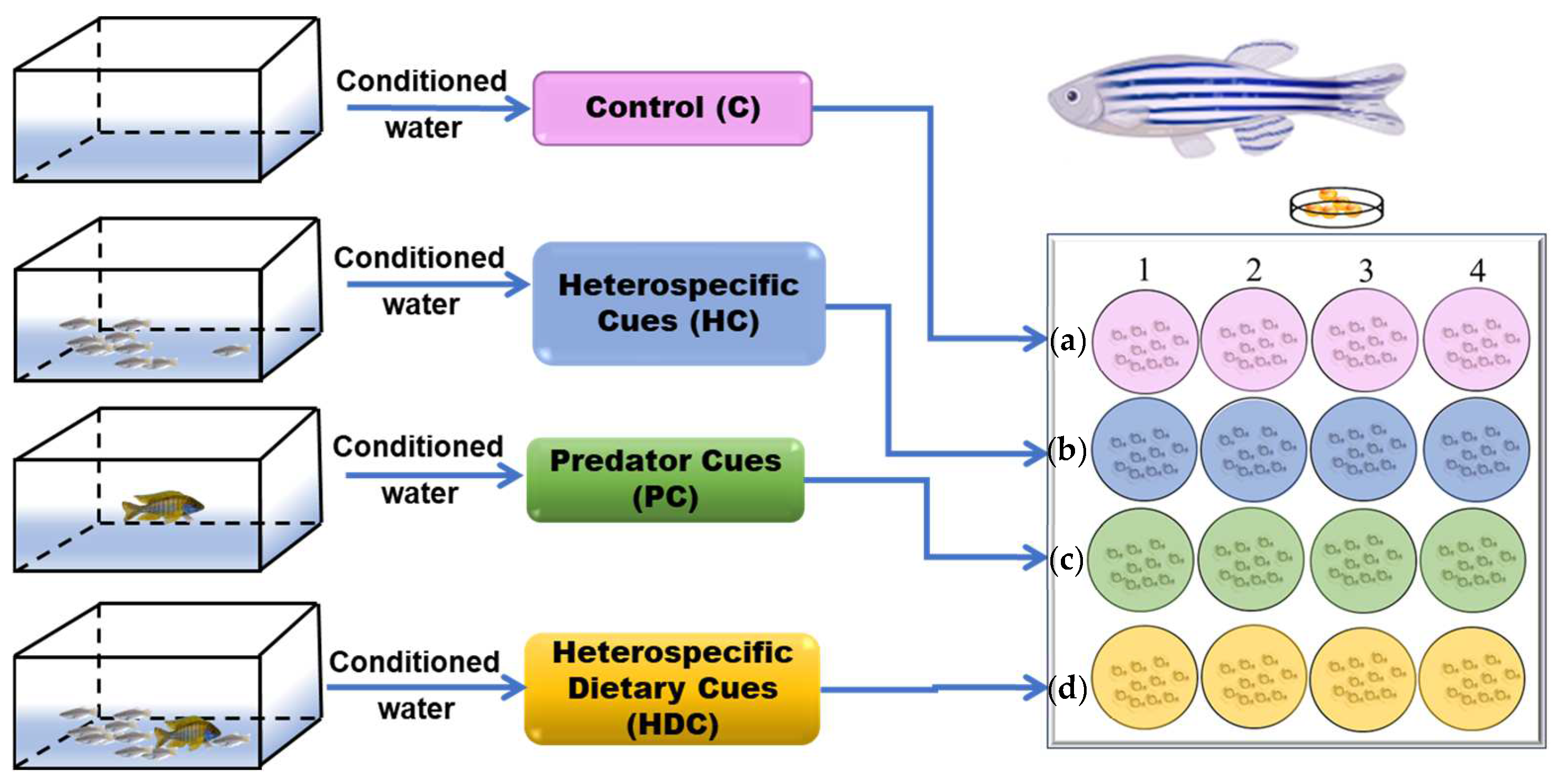
2.2. Preparation of Chemical Stimulus
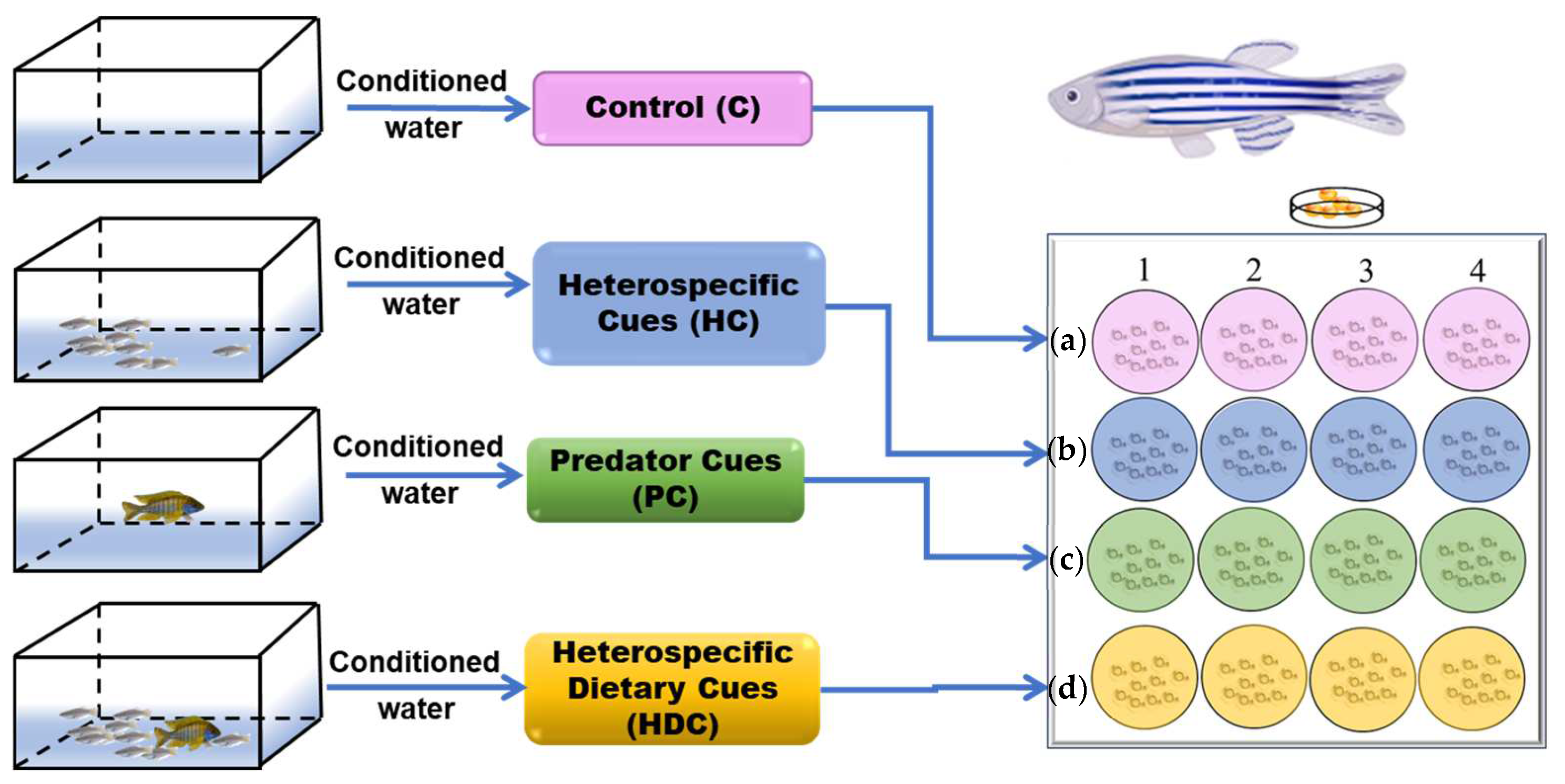
2.3. Experimental Design
2.3.1. Experiment 1—Hatching Time
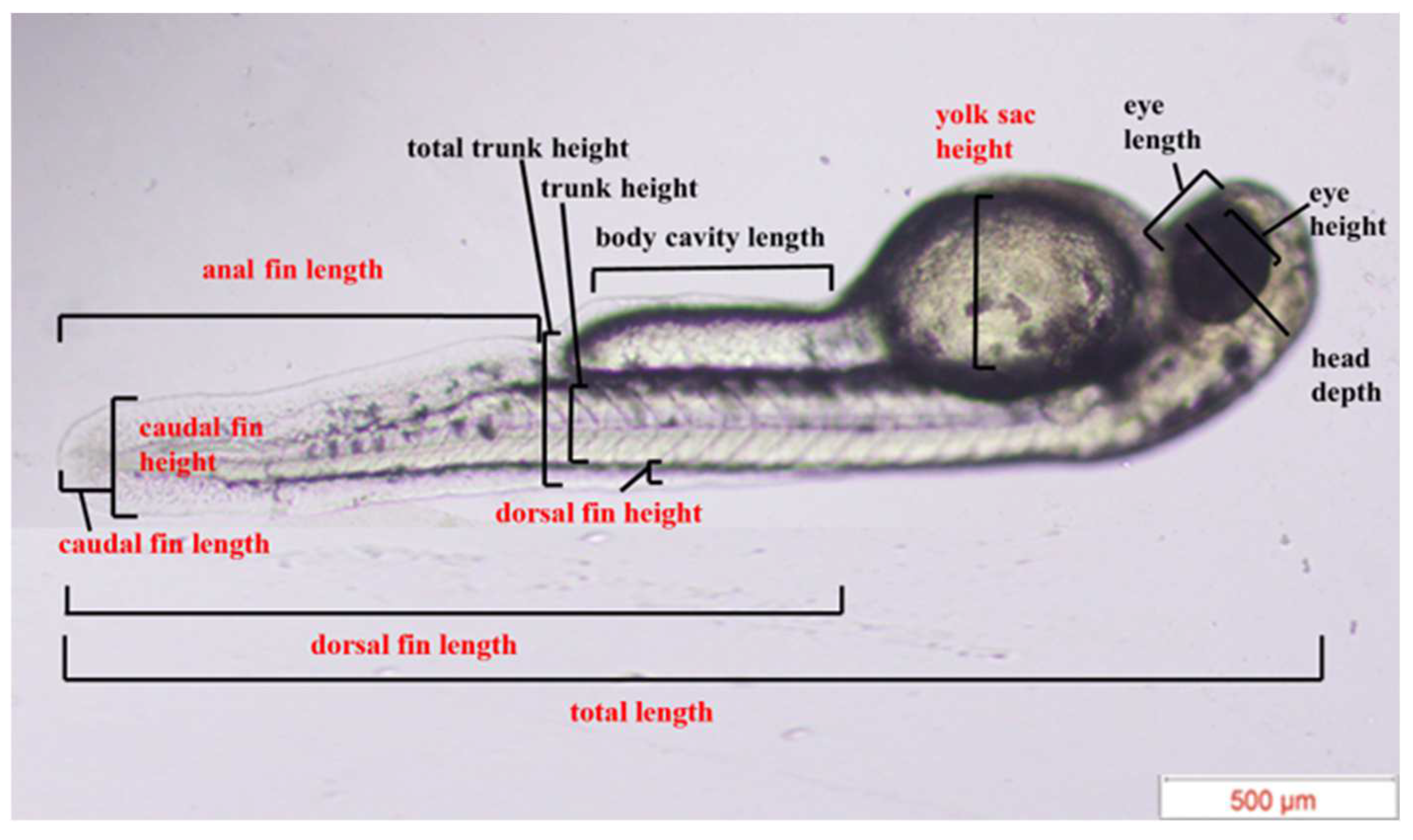
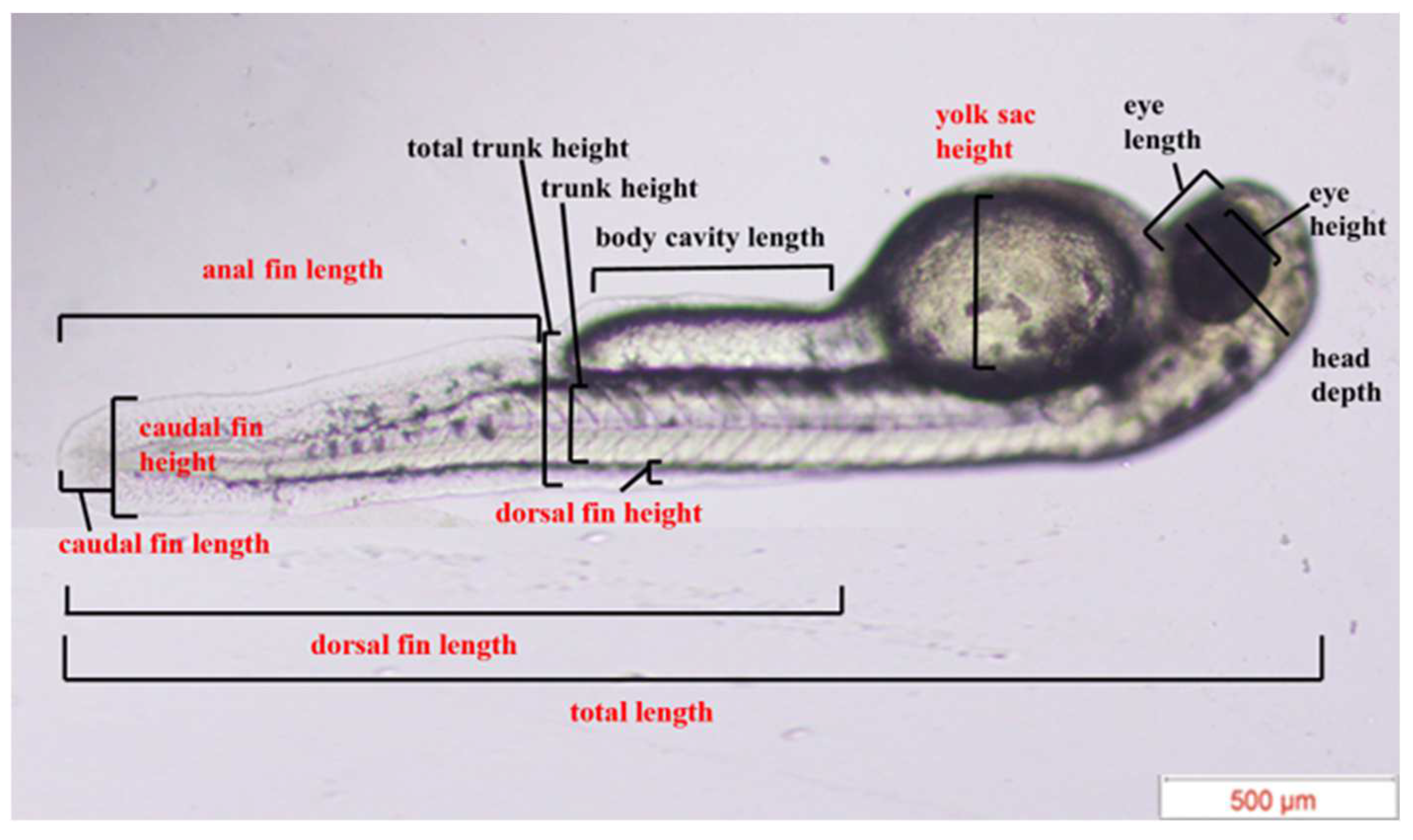
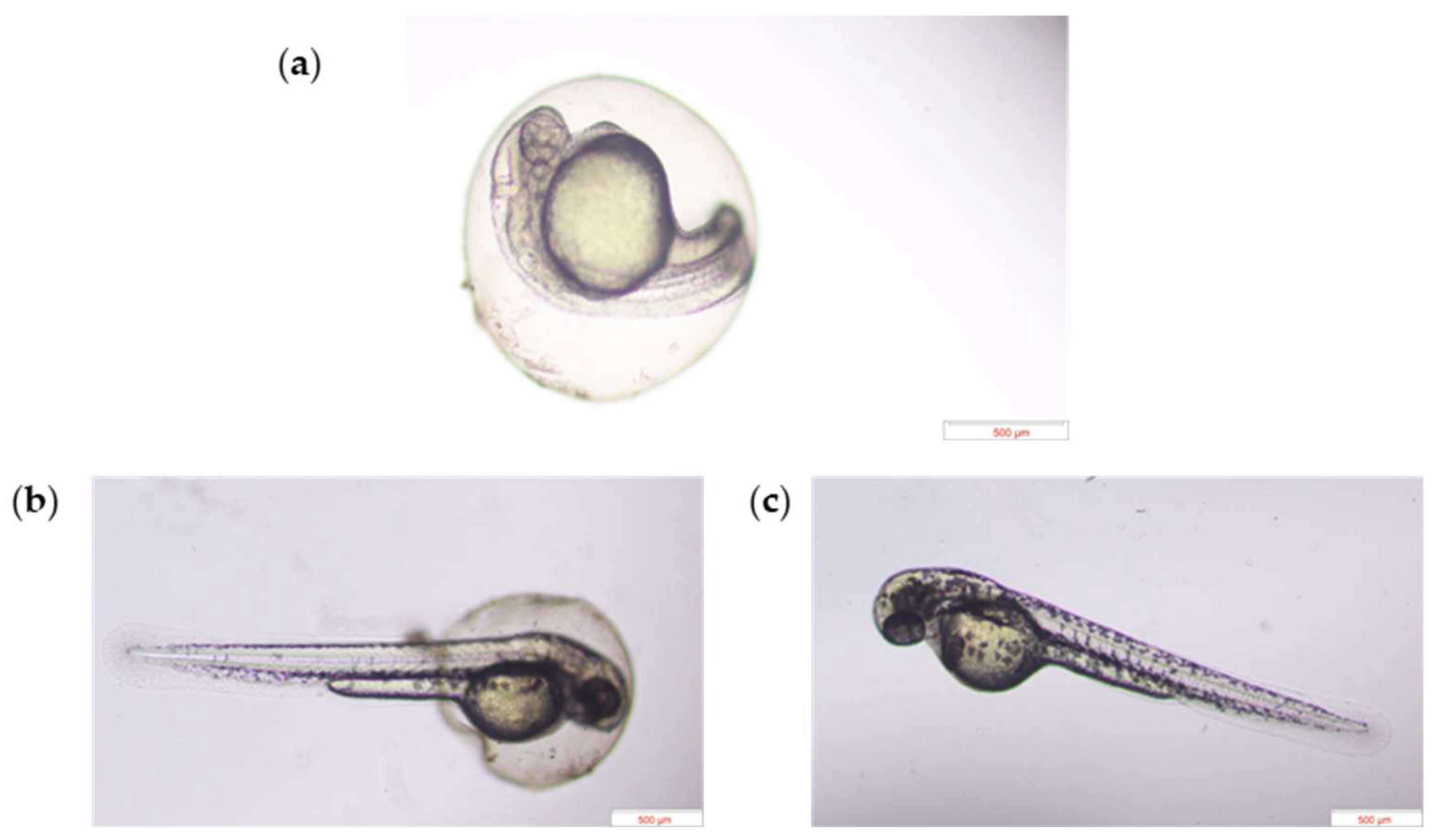
2.3.2. Experiment 2—Morphological Assessment
2.3.3. Experiment 3—Locomotor Behavior Assay
2.4. Statistical Analysis
3. Results
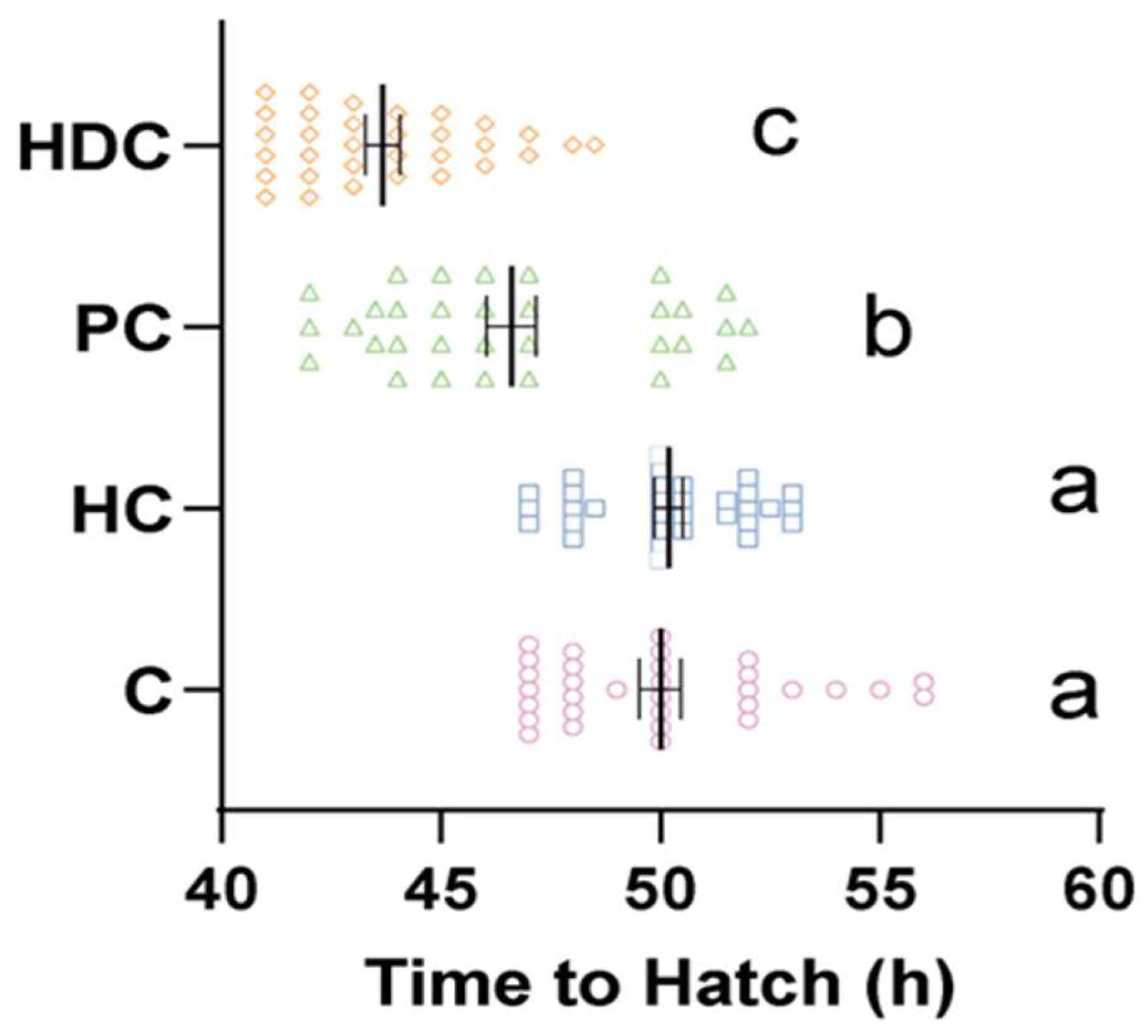
3.1. Heterospecific Cues (HDC) and Predator Cues (PC) Accelerate Hatching Time
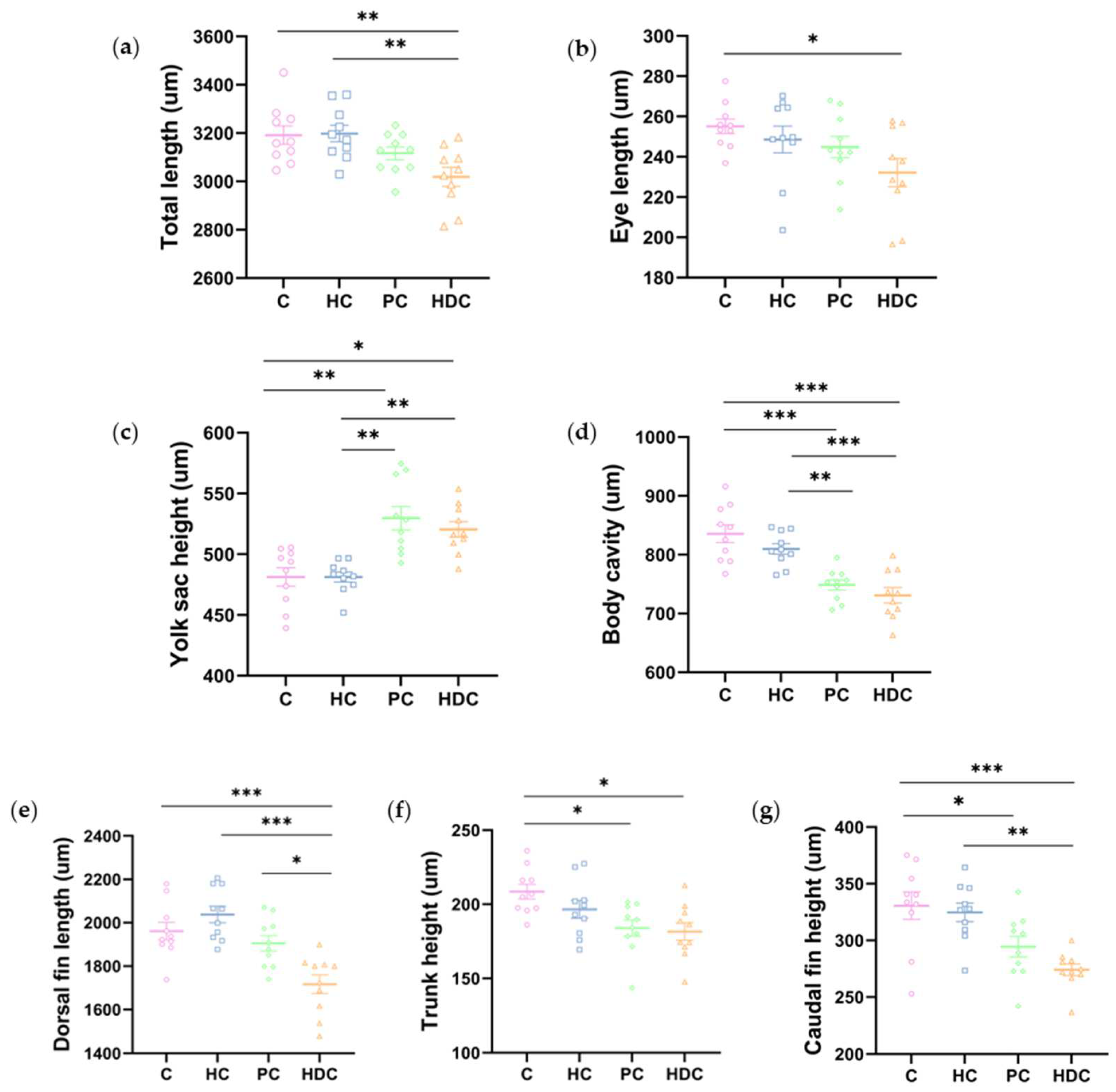
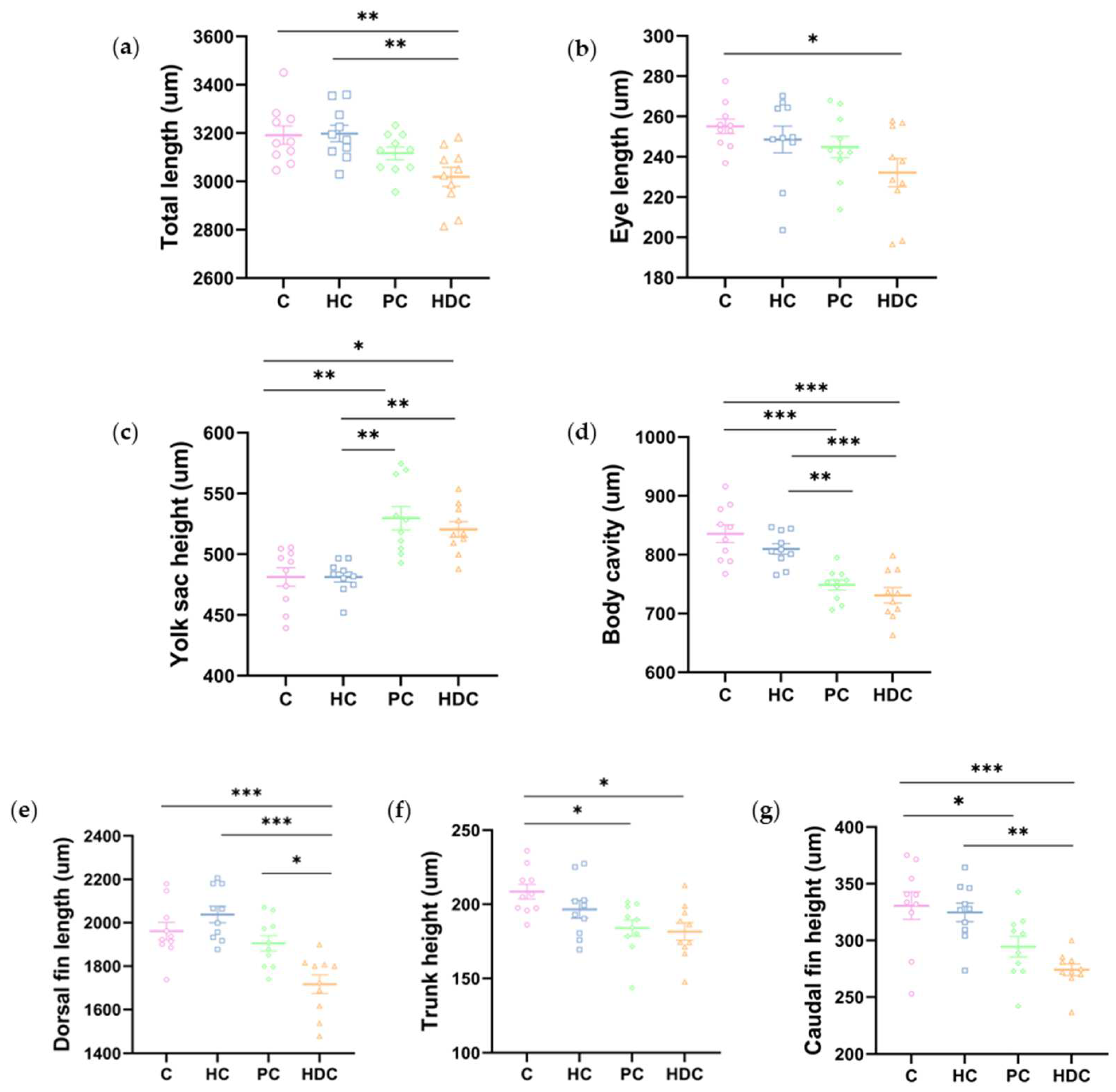
3.2. Heterospecific Dietary Cues (HDCs) and Predator Cues (PCs) Altered Developmental Morphology
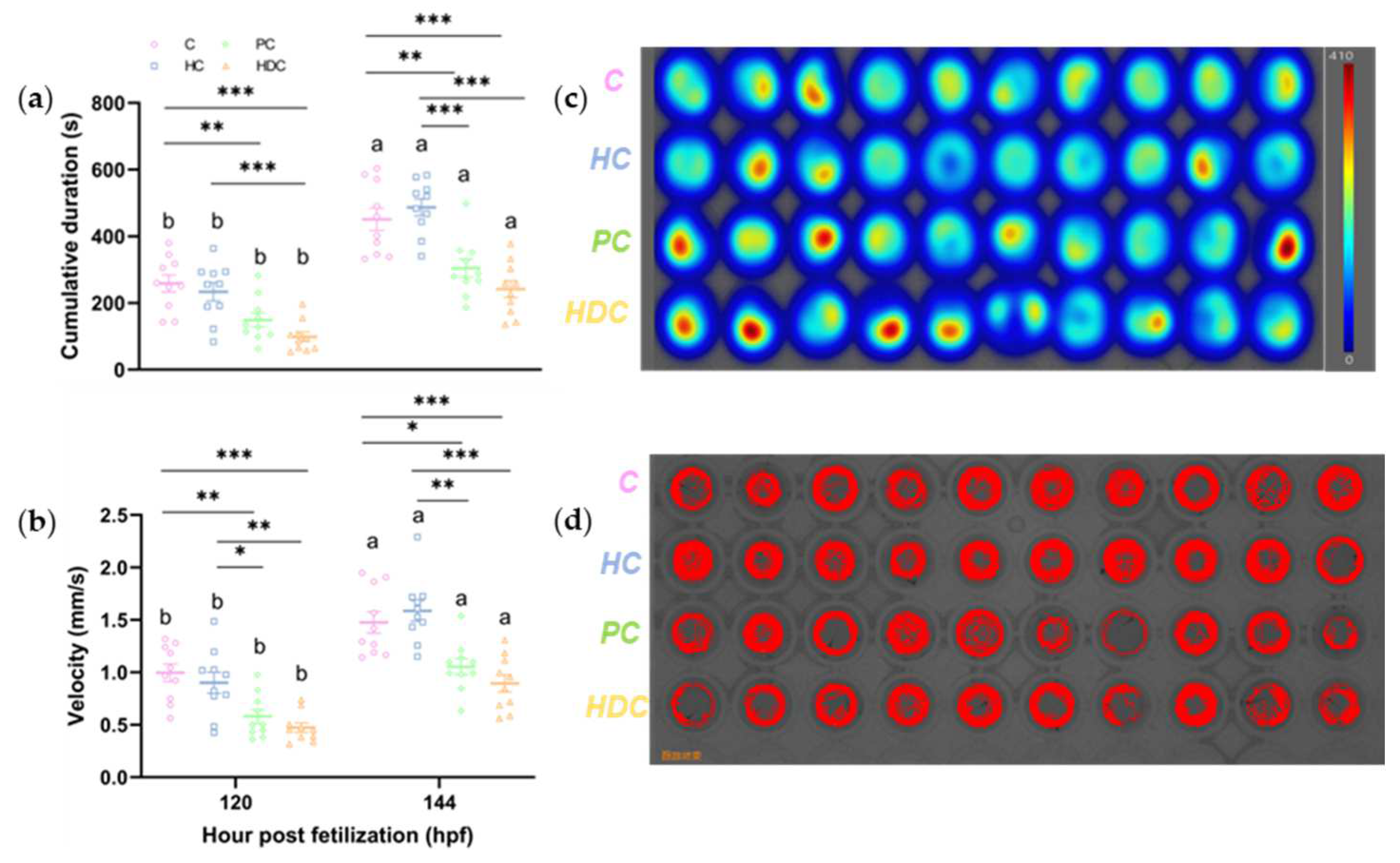
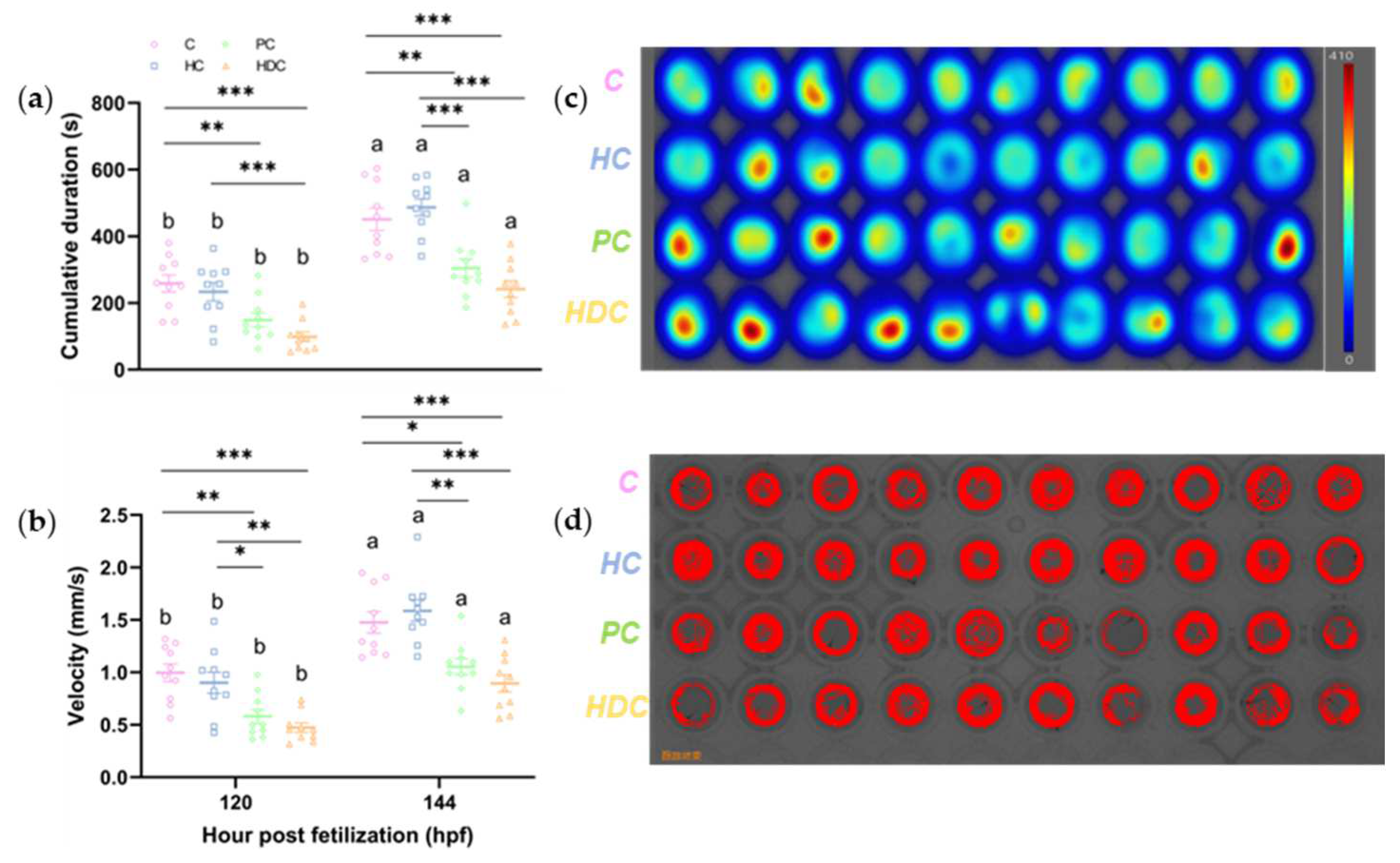
3.3. Heterospecific Dietary Cues (HDCs) and Predator Cues (PCs) Attenuate the Locomotor Ability of Zebrafish Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Zanette, L.Y.; Clinchy, M. Ecology and neurobiology of fear in free-living wildlife. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 297–318. [Google Scholar] [CrossRef]
- Du, W.G.; Shine, R. The behavioural and physiological ecology of embryos: Responding to the challenges of life inside an egg. Biol. Rev. 2022, 97, 1272–1286. [Google Scholar] [CrossRef]
- Hamdoun, A.; Epel, D. Embryo stability and vulnerability in an always changing world. Proc. Natl. Acad. Sci. USA 2007, 104, 1745–1750. [Google Scholar] [CrossRef] [PubMed]
- Ituarte, R.B.; Vázquez, M.G.; Bas, C.C. Chemically induced plasticity in early life history of Palaemon argentinus: Are chemical alarm cues conserved within palaemonid shrimps? J. Exp. Biol. 2019, 222, jeb199984. [Google Scholar]
- Warkentin, K.M. Environmentally cued hatching across taxa: Embryos respond to risk and opportunity. Integr. Comp. Biol. 2011, 51, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Wisenden, B.D. Chemical cues that indicate risk of predation. In Fish Pheromones Related Cues; Sorensen, P.W., Wisenden, B.D., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 131–148. [Google Scholar]
- Gervais, C.R.; Nay, T.; Brown, C. Friend or foe? Development of odour detection, differentiation and antipredator response in an embryonic elasmobranch. Mar. Freshw. Res. 2021, 72, 942–949. [Google Scholar] [CrossRef]
- Oulton, L.J.; Haviland, V.; Brown, C. Predator recognition in rainbowfish, Melanotaenia duboulayi, embryos. PLoS ONE 2013, 8, e76061. [Google Scholar] [CrossRef]
- Crowder, C.; Ward, J. Embryonic antipredator defenses and behavioral carryover effects in the fathead minnow (Pimephales promelas). Behav. Ecol. Sociobiol. 2022, 76, 27. [Google Scholar] [CrossRef]
- Horn, M.E.; Chivers, D.P. Preschool for small frys: Threat-sensitive learning of predators by embryonic fathead minnows. Anim. Behav. 2021, 178, 49–55. [Google Scholar] [CrossRef]
- Ferrari, M.C.O.; Chivers, D.P. The ghost of predation future: Threat-sensitive and temporal assessment of risk by embryonic woodfrogs. Behav. Ecol. Sociobiol. 2010, 64, 549–555. [Google Scholar] [CrossRef]
- Atherton, J.A.; McCormick, M.I. Active in the sac: Damselfish embryos use innate recognition of odours to learn predation risk before hatching. Anim. Behav. 2015, 103, 1–6. [Google Scholar] [CrossRef]
- Mathis, A.; Ferrari, M.C.; Windel, N.; Messier, F.; Chivers, D.P. Learning by embryos and the ghost of predation future. Proc. R. Soc. B 2008, 275, 2603–2607. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.B.; Alemadi, S.D.; Wisenden, B.D. Learned recognition of novel predator odour by convict cichlid embryos. Behav. Ecol. Sociobiol. 2013, 67, 1269–1273. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Paulson, D.C.; Orr, M. Zebrafish embryos hatch early in response to chemical and mechanical indicators of predation risk, resulting in underdeveloped swimming ability of hatchling larvae. Open Biol. 2022, 11, bio059229. [Google Scholar] [CrossRef] [PubMed]
- Mathis, A.; Smith, R.J.F. Chemical labeling of northern pike (Esox lucius) by the alarm pheromone of fathead minnows (Pimephales promelas). J. Chem. Ecol. 1993, 19, 1967–1979. [Google Scholar] [CrossRef]
- Laurila, A.; Kujasalo, J.; Ranta, E. Different antipredator behaviour in two anuran tadpoles: Effects of predator diet. Behav. Ecol. Sociobiol. 1997, 40, 329–336. [Google Scholar] [CrossRef]
- Laurila, A.; Kujasalo, J.; Ranta, E. Predator-induced changes in life history in two anuran tadpoles: Effects of predator diet. Oikos 1998, 83, 307–317. [Google Scholar] [CrossRef]
- Betancur-R, R.; Wiley, E.O.; Arratia, G.; Acero, A.; Bailly, N.; Miya, M.; Lecointre, G.; Ortí, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17, 162. [Google Scholar] [CrossRef]
- Wittbrodt, J.; Shima, A.; Schartl, M. Medaka—A model organism from the far East. Nat. Rev. Genet. 2002, 3, 53–64. [Google Scholar] [CrossRef]
- Santos, M.E.; Lopes, J.F.; Kratochwil, C.F. East African cichlid fishes. EvoDevo 2023, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Larsen, G.D. Uncommon uses for a common fish. Lab. Anim. 2015, 44, 429. [Google Scholar] [CrossRef] [PubMed]
- Mourabit, S.; Rundle, S.; Spicer, J.; Sloman, K. Alarm substance from adult zebrafish alters early embryonic development in offspring. Biol. Lett. 2010, 6, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Li, W. Embryonic substances induce alarm response in adult zebrafish (Danio rerio). J. Fish Biol. 2020, 97, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, E.; Páramo-Delgadillo, S. The culture of cichlids of southeastern Mexico. Aquac. Res. 2008, 39, 777–783. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Padilla, S.; Hunter, D.L.; Padnos, B.; Frady, S.; MacPhail, R.C. Assessing locomotor activity in larval zebrafish: Influence of extrinsic and intrinsic variables. Neurotoxicol. Teratol. 2011, 33, 624–630. [Google Scholar] [CrossRef]
- Murphy, C.A.; Rose, K.A.; del Carmen Alvarez, M.; Fuiman, L.A. Modeling larval fish behavior: Scaling the sublethal effects of methylmercury to population-relevant endpoints. Aquat. Toxicol. 2008, 86, 470–484. [Google Scholar] [CrossRef]
- Ulhaq, M.; Örn, S.; Carlsson, G.; Morrison, D.A.; Norrgren, L. Locomotor behavior in zebrafish (Danio rerio) larvae exposed to perfluoroalkyl acids. Aquat. Toxicol. 2013, 144, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ho, N.Y.; Alshut, R.; Legradi, J.; Weiss, C.; Reischl, M.; Mikut, R.; Liebel, U.; Müller, F.; Strähle, U. Zebrafish embryos as models for embryotoxic and teratological effects of chemicals. Reprod. Toxicol. 2009, 28, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Kohda, M. Individual specialized foraging repertoires in the piscivorous cichlid fish Lepidiolamprologus profundicola. Anim. Behav. 1994, 48, 1123–1131. [Google Scholar] [CrossRef]
- Shima, A.; Mitani, H. Medaka as a research organism: Past, present and future. Mech. Dev. 2004, 121, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Warkentin, K.M. Adaptive plasticity in hatching age: A response to predation risk trade-offs. Proc. Natl. Acad. Sci. USA 1995, 92, 3507–3510. [Google Scholar] [CrossRef] [PubMed]
- Warkentin, K.M. Wasp predation and wasp-induced hatching of red-eyed treefrog eggs. Anim. Behav. 2000, 60, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Serrano-Rojas, S.J.; Warkentin, K.M. Multimodal mechanosensing enables treefrog embryos to escape egg-predators. J. Exp. Biol. 2020, 223, jeb236141. [Google Scholar] [CrossRef]
- Ferrari, M.C.; Wisenden, B.D.; Chivers, D.P. Chemical ecology of predator–prey interactions in aquatic ecosystems: A review and prospectus. Can. J. Zool. 2010, 88, 698–724. [Google Scholar] [CrossRef]
- Touchon, J.C.; Wojdak, J.M. Plastic hatching timing by red-eyed treefrog embryos interacts with larval predator identity and sublethal predation to affect prey morphology but not performance. PLoS ONE 2014, 9, e100623. [Google Scholar] [CrossRef]
- Kostoglou, K.N.; van Dongen, W.F.; Bowe, S.J.; Weston, M.A. Shorebird embryos exhibit anti-predator responses. Ibis 2021, 163, 1425–1436. [Google Scholar] [CrossRef]
- Mezrai, N.; Arduini, L.; Dickel, L.; Chiao, C.-C.; Darmaillacq, A.-S. Awareness of danger inside the egg: Evidence of innate and learned predator recognition in cuttlefish embryos. Learn. Behav. 2020, 48, 401–410. [Google Scholar] [CrossRef]
- Kempster, R.M.; Hart, N.S.; Collin, S.P. Survival of the stillest: Predator avoidance in shark embryos. PLoS ONE 2013, 8, e52551. [Google Scholar] [CrossRef]
- Ball, R.E.; Oliver, M.K.; Gill, A.B. Early life sensory ability—Ventilatory responses of thornback ray embryos (Raja clavata) to predator-type electric fields. Dev. Neurobiol. 2016, 76, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Gazzola, A.; Brandalise, F.; Rubolini, D.; Rossi, P.; Galeotti, P. Fear is the mother of invention: Anuran embryos exposed to predator cues alter life-history traits, post-hatching behaviour and neuronal activity patterns. J. Exp. Biol. 2015, 218, 3919–3930. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, A.; Li, Y.; Yan, Z.; Zhang, H.; Jiang, B.; Chen, J.; Wang, D.; Wang, H.; Li, X.; Lu, Z.; et al. Chemical Cues Released by Predators’ Consumption of Heterospecific Prey Alter the Embryogenesis of Zebrafish. Fishes 2024, 9, 95. https://doi.org/10.3390/fishes9030095
Lin A, Li Y, Yan Z, Zhang H, Jiang B, Chen J, Wang D, Wang H, Li X, Lu Z, et al. Chemical Cues Released by Predators’ Consumption of Heterospecific Prey Alter the Embryogenesis of Zebrafish. Fishes. 2024; 9(3):95. https://doi.org/10.3390/fishes9030095
Chicago/Turabian StyleLin, Ainuo, Yaxi Li, Zhi Yan, Huilin Zhang, Baozhen Jiang, Jingjing Chen, Derui Wang, Huan Wang, Xiaodong Li, Zhen Lu, and et al. 2024. "Chemical Cues Released by Predators’ Consumption of Heterospecific Prey Alter the Embryogenesis of Zebrafish" Fishes 9, no. 3: 95. https://doi.org/10.3390/fishes9030095
APA StyleLin, A., Li, Y., Yan, Z., Zhang, H., Jiang, B., Chen, J., Wang, D., Wang, H., Li, X., Lu, Z., & Li, K. (2024). Chemical Cues Released by Predators’ Consumption of Heterospecific Prey Alter the Embryogenesis of Zebrafish. Fishes, 9(3), 95. https://doi.org/10.3390/fishes9030095