
Use of Environmental DNA to Evaluate the Spatial Distribution of False Kelpfish (Sebastiscus marmoratus) in Nearshore Areas of Gouqi Island

Abstract
1. Introduction
2. Materials and Methods
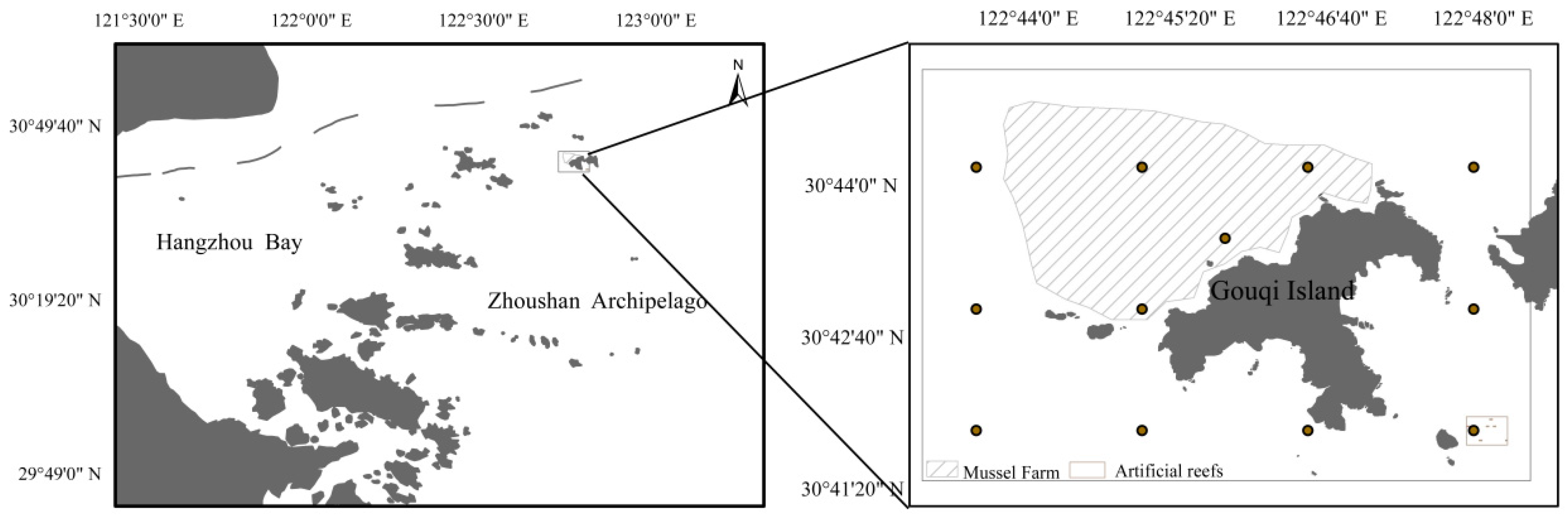
2.1. Sample Collection
2.2. DNA Extraction
2.3. QPCR
2.4. Acquisition of Environmental Factor Data
2.5. Data Analysis Methods
- y is eDNA concentration;
- Β0 is the intercept;
- the sampling seasonal variable is a;
- the sampling layer variable is b;
- the sampling area variable is c;
- (1|Site) indicates the sampling point as a random effect.
3. Results
3.1. Plasmid Construction
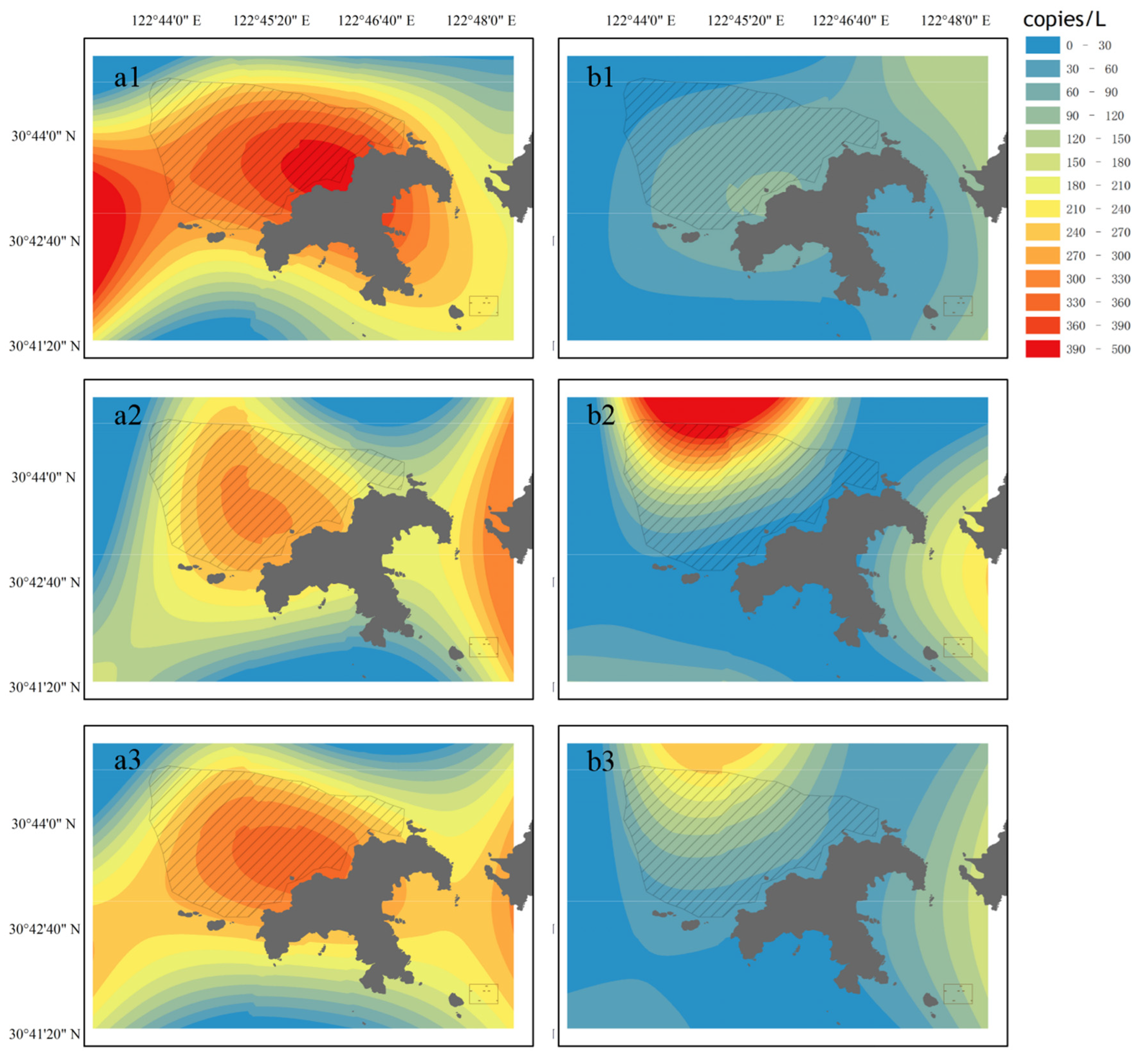
3.2. The Concentration of False Kelpfish (Sebastiscus marmoratus) eDNA
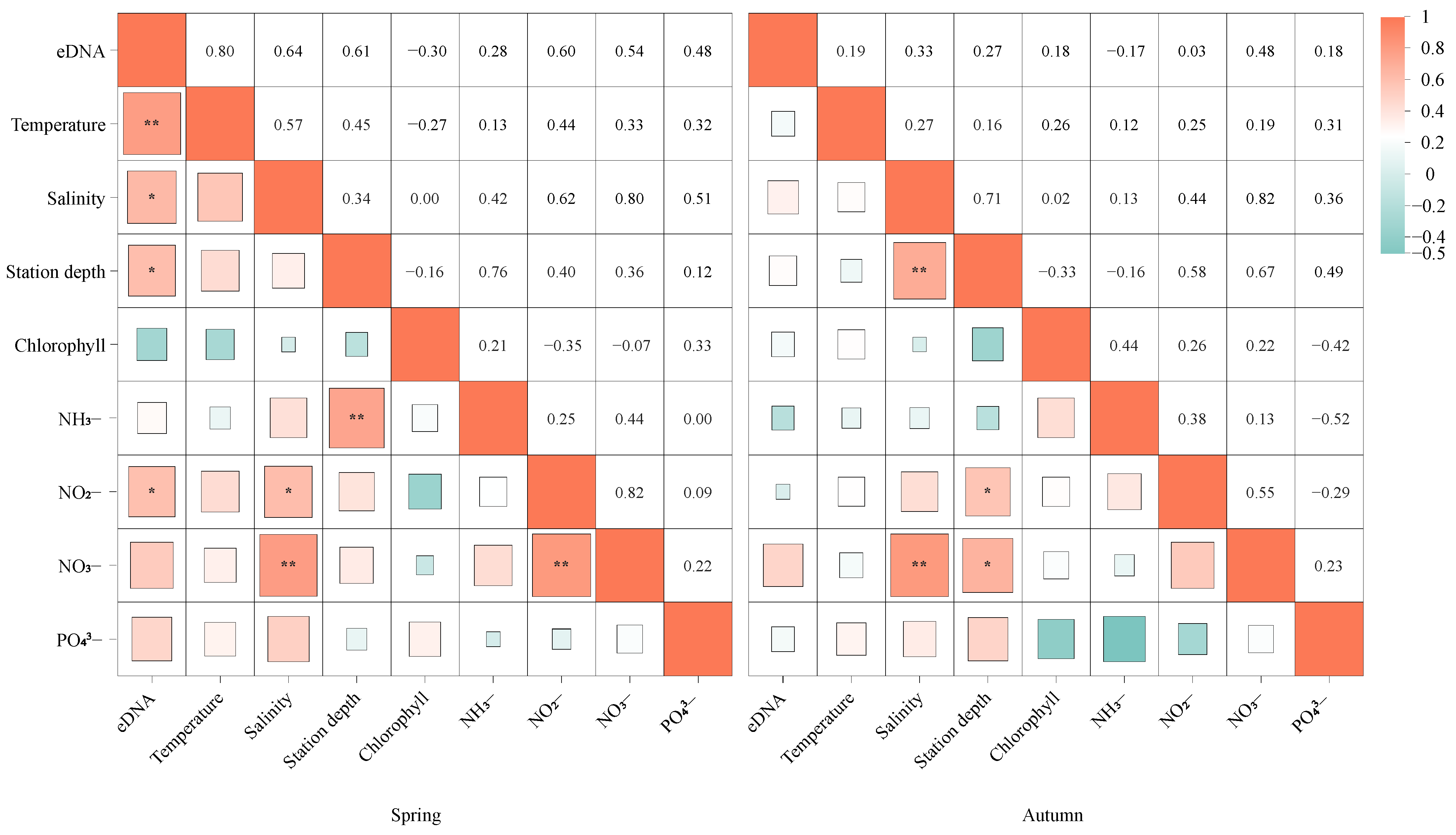
3.3. The Relationship Between False Kelpfish (S. marmoratus) eDNA Concentration and Environmental Factors
4. Discussion
4.1. Distribution Characteristics of S. marmoratus
4.2. Application of eDNA Surveys in Marine Fish: Potential, Limitations, and Prospects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valdez, J.W.; Callaghan, C.T.; Junker, J.; Purvis, A.; Hill, S.L.L.; Pereira, H.M. The undetectability of global biodiversity trends using local species richness. Ecography 2023, 2023, e06604. [Google Scholar] [CrossRef]
- Rullens, V.; Stephenson, F.; Lohrer, A.M.; Townsend, M.; Pilditch, C.A. Combined species occurrence and density predictions to improve marine spatial management. Ocean Coast. Manag. 2021, 209, 105697. [Google Scholar] [CrossRef]
- Itakura, H.; Wakiya, R.; Yamamoto, S.; Kaifu, K.; Sato, T.; Minamoto, T. Environmental DNA analysis reveals the spatialdistribution, abundance, and biomass of Japaneseeels at the river-basin scale. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 331–520. [Google Scholar] [CrossRef]
- Yuan, X.; Jiang, Y.; Zhang, H.; Jin, Y.; Ling, J. Quantitative assessment of fish assemblages on artificial reefs using acoustic and conventional netting methods, in Xiangshan Bay, Zhejiang Province, China. J. Appl. Ichthyol. 2021, 37, 389–399. [Google Scholar] [CrossRef]
- Yamamoto, S.; Minami, K.; Fukaya, K.; Takahashi, K.; Sawada, H.; Murakami, H.; Tsuji, S.; Hashizume, H.; Kubonaga, S.; Horiuchi, T.; et al. Environmental DNA as a ‘snapshot’ of fish distribution: A case study of Japanese jack mackerel in Maizuru Bay, Sea of Japan. PLoS ONE 2016, 11, e0149786. [Google Scholar]
- Doi, H.; Inui, R.; Akamatsu, Y.; Kanno, K.; Yamanaka, H.; Takahara, T.; Minamoto, T. Environmental DNA analysis for estimating the abundance and biomass of stream fish. Freshw. Biol. 2016, 62, 30–39. [Google Scholar] [CrossRef]
- Dougherty, M.M.; Larson, E.R.; Renshaw, M.A.; Gantz, C.A.; Egan, S.P.; Erickson, D.M.; Lodge, D.M. Environmental DNA (eDNA) detects the invasive rusty crayfish Orconectes rusticus at low abundances. J. Appl. Ecol. 2016, 53, 722–732. [Google Scholar] [CrossRef]
- Minamoto, T.; Fukuda, M.; Katsuhara, K.R.; Fujiwara, A. Environmental DNA reflects spatial and temporal jellyfish distribution. PLoS ONE 2017, 12, e0173073. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Estimating occupancy and abundance of stream amphibians using environmental DNA from filtered water samples. Can. J. Fish. Aquat. Sci. 2013, 70, 1123–1130. [Google Scholar] [CrossRef]
- Buxton, A.S.; Groombridge, J.J.; Zakaria, N.B.; Griffiths, R.A. Seasonal variation in environmental DNA in relation to population size and environmental factors. Sci. Rep 2017, 7, 46294. [Google Scholar] [CrossRef]
- Darling, J.A.; Mahon, A.R. From molecules to management: Adopting DNA-based methods for monitoring biological invasions in aquatic environments. Environ. Res. 2011, 111, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Jerde, C.L.; Mahon, A.R.; Chadderton, W.L.; Lodge, D.M. “Sight-unseen” detection of rare aquatic species using environmental DNA. Conserv Lett 2017, 4, 150–157. [Google Scholar] [CrossRef]
- Feist, S.M.; Jones, R.L.; Copley, J.L.; Pearson, L.S.; Berry, G.A.; Qualls, C.P. Development and validation of an environmental DNA method for detection of alligator snapping turtle (Macrochelys temminckii). Chelonian Conserv. Biol. 2018, 17, 271–279. [Google Scholar] [CrossRef]
- Xu, N.; Xiong, M.H.; Shao, K.; Que, Y.F.; Li, J.Y. Preliminary study on environmental DNA metabarcoding for detecting biodiversity in the middle and lower reaches of the Yangtze River. Res. Environ. Sci. 2020, 33, 1187–1196. [Google Scholar]
- Xu, M.Z.; Yang, Y.; Zhang, J.H.; Fu, X.D. Application Challenges of Environmental DNA Technology in Monitoring the Invasion of Bivalves. Acta Ecol. Sin. 2023, 43, 4423–4433. [Google Scholar]
- Wang, X.Y.; Zhang, H.B.; Lu, G.Q.; Gao, T.X. Detection of an invasive species through an environmental DNA approach: The example of the red drum Sciaenops ocellatus in the East China Sea. Sci. Total Environ. 2022, 815, 152865. [Google Scholar] [CrossRef]
- Gao, T.; Chen, Z.; Wang, X. A New Method for Surveying Fish Diversity in Coastal Areas: Environmental DNA Analysis Technology. J. Zhejiang Ocean Univ. (Nat. Sci. Ed.) 2018, 37, 1–7. [Google Scholar]
- Zhao, S.L.; Xu, H.X.; Zhong, J.S. Fauna of Marine Fishes in Zhejiang; Zhejiang Science and Technology Press: Hangzhou, China, 2016. [Google Scholar]
- Gao, T.-X.; Chen, Z.; Wang, X.-Y.; Zhang, H.-B.; Shi, H.-L. Discussion on the Correlation between Density of Sebastes schlegelii Cultivation and its eDNA. J. Hydrobiol. 2022, 46, 1007–1015. [Google Scholar]
- Pei, W.H.; Hu, C.Y.; Zhang, X.M.; Chen, Y.Y.; Liu, Y.D. Study on the Community Structure of Macrobenthos in the Mussel Breeding Area of Gouqi Island. Oceanol. Et Limnol. Sin. 2023, 54, 526–536. [Google Scholar]
- Li, F.; Xu, M.; Liu, Q.; Wang, Z.F.; Xu, W.J. Ecological restoration zoning for a marine protected area: A case study of Haizhouwan National Marine Park, China. Sci. Total Environ. 2014, 98, 158–166. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps. 2019. Available online: https://cran.r-project.org/web/packages/pheatmap/pheatmap.pdf (accessed on 10 April 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Erickson, R.A.; Rees, C.B.; Coulter, A.A.; Merkes, C.M.; McCalla, S.G.; Touzinsky, K.F.; Walleser, L.; Goforth, R.R.; Amberg, J.J. Detecting the Movement and Spawning Activity of Bigheaded Carps with Environmental DNA. Mol. Ecol. Resour. 2016, 16, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, S.; Wang, Z.; Lin, J.; Wang, K. Habitat Suitability Assessment for Sebasticus marmoratus in the Saddle Islands. Acta Ecol. Sin. 2016, 36, 3765–3774. [Google Scholar]
- Hayer, C.-A.; Bayless, M.F.; George, A.E.; Thompson, N.; Richter, C.A.; Chapman, D. Use of Environmental DNA to Detect Grass Carp Spawning Events. Fishes 2020, 5, 27. [Google Scholar] [CrossRef]
- Tillotson, M.D.; Kelly, R.P.; Duda, J.J.; Hoy, M.; Kralj, J.; Quinn, T.P. Concentrations of environmental DNA (eDNA) reflect spawning salmon abundance at fine spatial and temporal scales. Biol. Conserv. 2018, 220, 1–11. [Google Scholar] [CrossRef]
- Wu, C.W. Biological Research on Sebastiscus marmoratus in the Nearshore Waters of Zhoushan, Zhejiang Province. J. Zhejiang Ocean. Univ. (Nat. Sci. Ed.) 1999, 3, 185–190+226. [Google Scholar]
- Mukherjee, S.; Chaudhuri, A.; Kundu, N.; Mitra, S.; Homechaudhuri, S. Comprehensive analysis of fish assemblages in relation to seasonal environmental variables in an estuarine river of Indian sundarbans. Estuaries Coasts 2012, 36, 192–202. [Google Scholar] [CrossRef]
- Yin, M.L. Fish Ecology; China Agricultural Press: Beijing, China, 1995. [Google Scholar]
- Li, X.D. The Relationship between Seawater Temperature and Fishing Grounds. Acta Oceanol. Sin. 1982, 1, 103–113. (In Chinese) [Google Scholar]
- Belkin, I.M.; O’Reilly, J.E. An algorithm for oceanic front detection in chlorophyll and SST satellite imagery. J. Mar. Syst. 2009, 78, 319–326. [Google Scholar] [CrossRef]
- Andersson, A.; Haecky, P.; Hagström, A. Effect of temperature and light on the growth of micro-nano-and pico-plankton: Impact on algal succession. Mar. Biol. 1994, 120, 511–520. [Google Scholar] [CrossRef]
- Mazumder, A.; Taylor, W.D.; McQueen, D.J.; Lean, D.R. Effects of fish and plankton on lake temperature and mixing depth. Science 1990, 247, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.Q. Approaches to Mechanisms and Control of Eutrophication of Shallow Lakes i n the Middle and Lower Reaches of the Yangze River. J. Lake Sci. 2002, 14, 193–202. [Google Scholar]
- Ji, Y.; Ashton, L.; Pedley, S.M.; Edwards, D.P.; Tang, Y.; Nakamura, A.; Kitching, R.; Dolman, P.M.; Woodcock, P.; Edwards, F.A.; et al. Reliable, verifiable and efficient monitoring of biodiversity via meta barcoding. Ecol. Lett. 2013, 16, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.P.; O’donnell, J.L.; Lowell, N.C.; Shelton, A.O.; Samhouri, J.F.; Hennessey, S.M.; Feist, B.E.; Williams, G.D. Genetic signatures of ecological diversity along an urbanization gradient. PeerJ 2016, 4, e2444. [Google Scholar] [CrossRef]
- Barnes, M.A.; Turner, C.R. The ecology of environmental DNA and implications for conservation genetics. Conserv Genet 2016, 17, 1–17. [Google Scholar] [CrossRef]
- Murakami, H.; Yoon, S.; Kasai, A.; Minamoto, T.; Yamamoto, S.; Sakata, M.K.; Horiuchi, T.; Sawada, H.; Kondoh, M.; Yamashita, Y.; et al. Correction to: Dispersion and degradation of environmental DNA from caged fish in a marine environment. Fish Sci. 2019, 85, 1109. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Estimate | Standard Error | T Value |
|---|---|---|---|
| (Intercept) | 6.21 | 0.80 | 7.74 |
| type | 0.53 | 0.31 | 1.73 |
| layer | −0.49 | 0.28 | −1.75 |
| season | −1.35 | 0.28 | −4.85 |
| MLR Model | R2 | RMSE |
|---|---|---|
| Y = 460.16 − 19.10 × Tem | 0.39 | 83.00 |
| Y = 39.60 − 19.18 × Tem + 6.25 × Dep | 0.52 | 73.36 |
| Y = 314.88 − 18.52 × Tem + 5.39 × Dep + 2314.32 × PO43− | 0.55 | 71.56 |
| Y = 324.86 − 19.19 × Tem + 4.32 × Dep + 2760.17 × NO2− + 2928.13 × PO43− | 0.55 | 70.85 |
| Y = 476.70 − 28.26 × Tem − 6.25 × Chl + 279.08 × NH3− + 341.35 × NO3− + 4142.67 × PO43− | 0.58 | 68.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, R.; Hao, H.; Yin, R.; Zhao, P.; Chen, F.; Zhou, Y.; Chai, X. Use of Environmental DNA to Evaluate the Spatial Distribution of False Kelpfish (Sebastiscus marmoratus) in Nearshore Areas of Gouqi Island. Fishes 2024, 9, 418. https://doi.org/10.3390/fishes9100418
Jiang R, Hao H, Yin R, Zhao P, Chen F, Zhou Y, Chai X. Use of Environmental DNA to Evaluate the Spatial Distribution of False Kelpfish (Sebastiscus marmoratus) in Nearshore Areas of Gouqi Island. Fishes. 2024; 9(10):418. https://doi.org/10.3390/fishes9100418
Chicago/Turabian StyleJiang, Rijin, Huibo Hao, Rui Yin, Peng Zhao, Feng Chen, Yongdong Zhou, and Xuejun Chai. 2024. "Use of Environmental DNA to Evaluate the Spatial Distribution of False Kelpfish (Sebastiscus marmoratus) in Nearshore Areas of Gouqi Island" Fishes 9, no. 10: 418. https://doi.org/10.3390/fishes9100418
APA StyleJiang, R., Hao, H., Yin, R., Zhao, P., Chen, F., Zhou, Y., & Chai, X. (2024). Use of Environmental DNA to Evaluate the Spatial Distribution of False Kelpfish (Sebastiscus marmoratus) in Nearshore Areas of Gouqi Island. Fishes, 9(10), 418. https://doi.org/10.3390/fishes9100418