
Swimming under Pressure: The Sub-Lethal Effects of a Pesticide on the Behaviour of Native and Non-Native Cypriniformes Fish

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pyrethroid Insecticide
2.2. Target Species and Life Stages
2.3. Fish Sampling and Holding Facilities
2.4. Mesocosm Facility
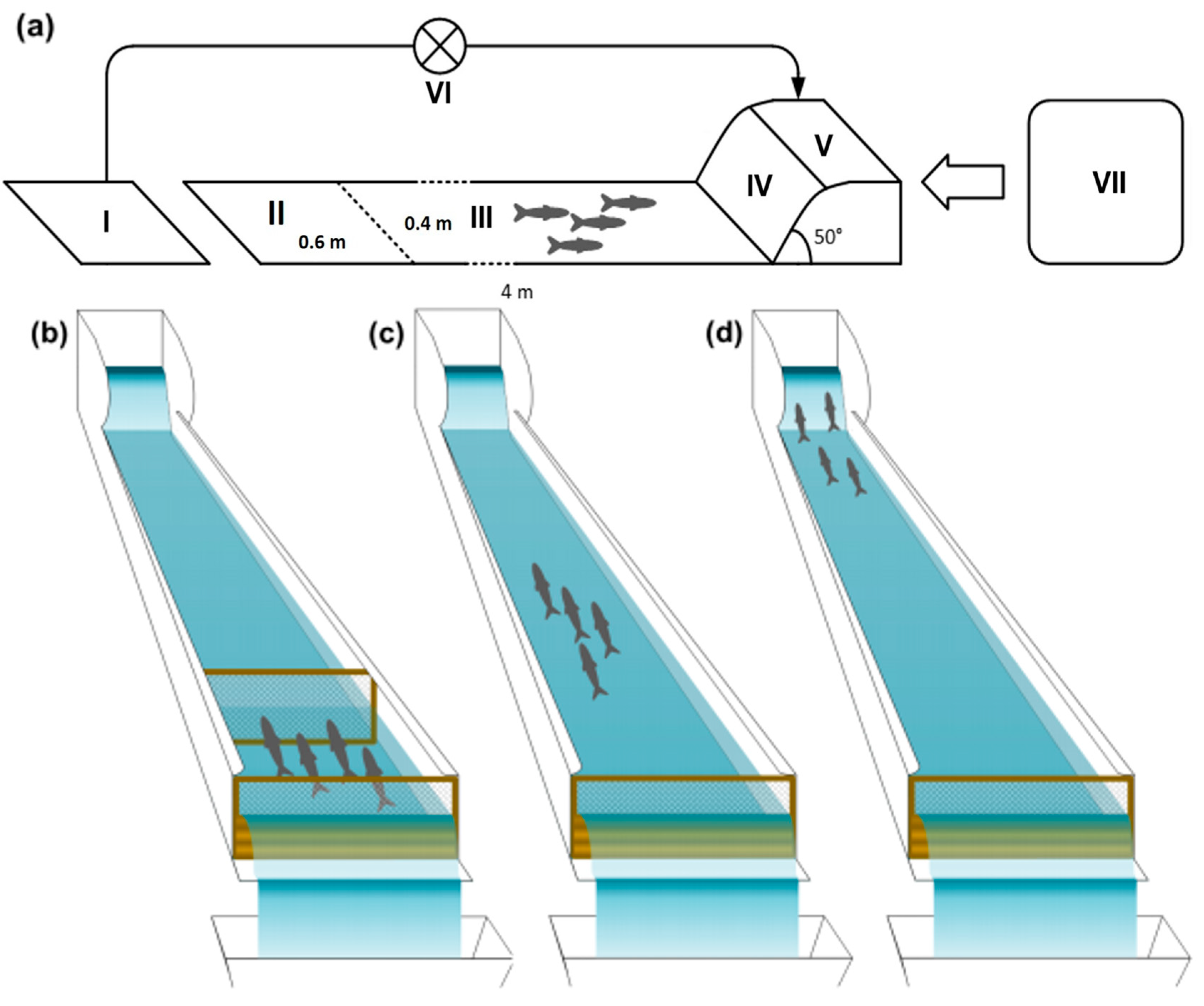
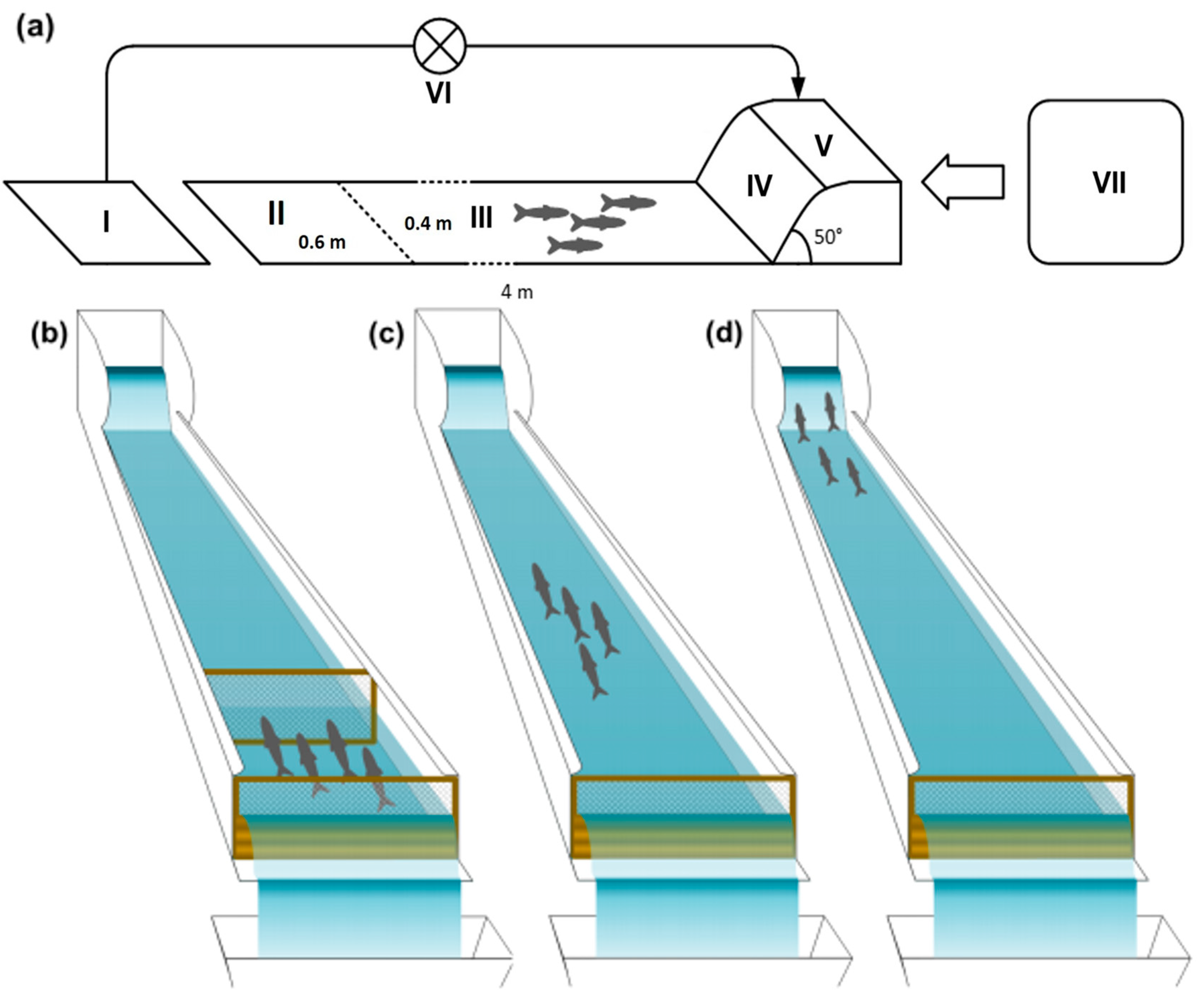
2.5. Experimental Set-Up
2.6. Data Analyses
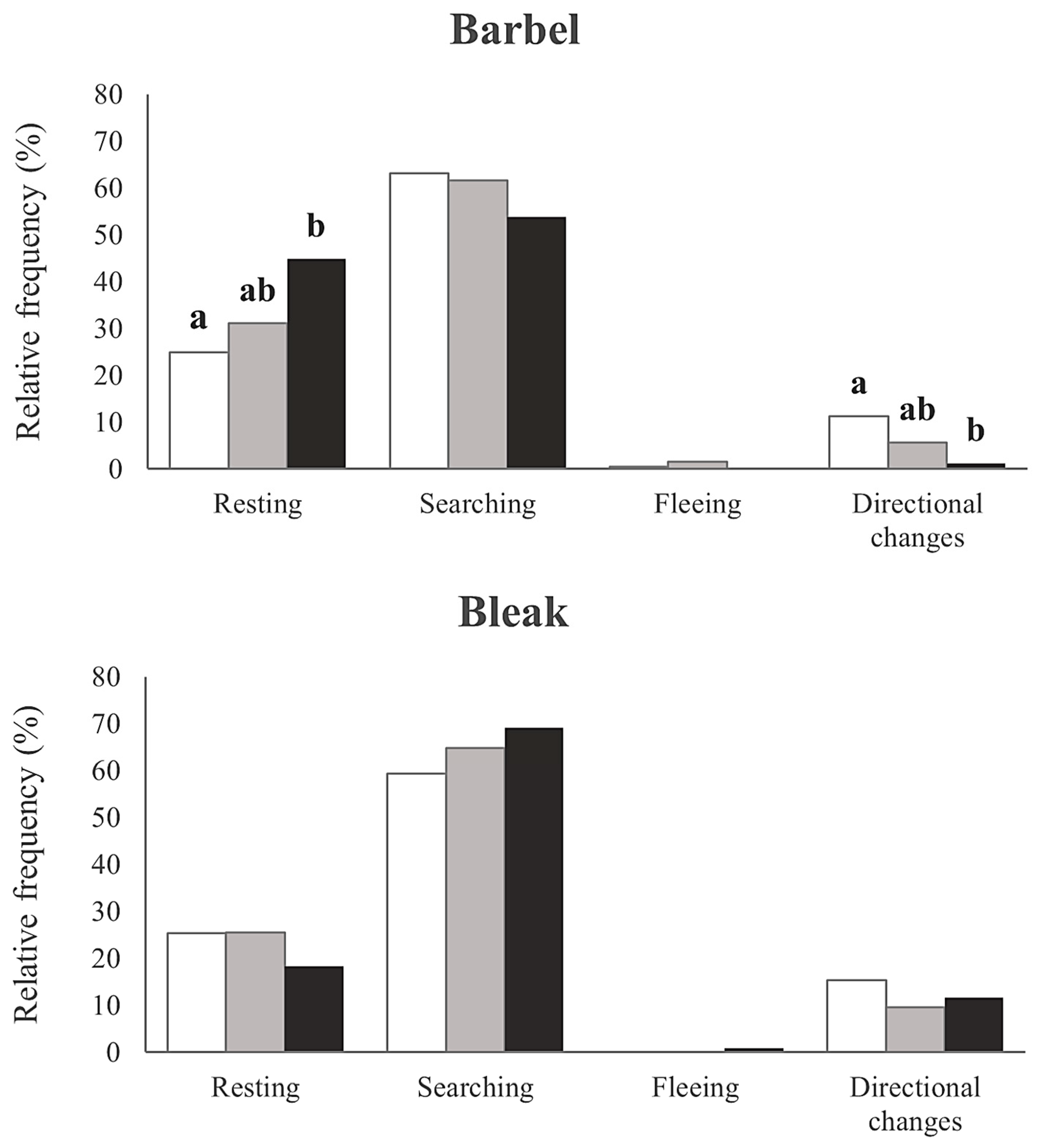
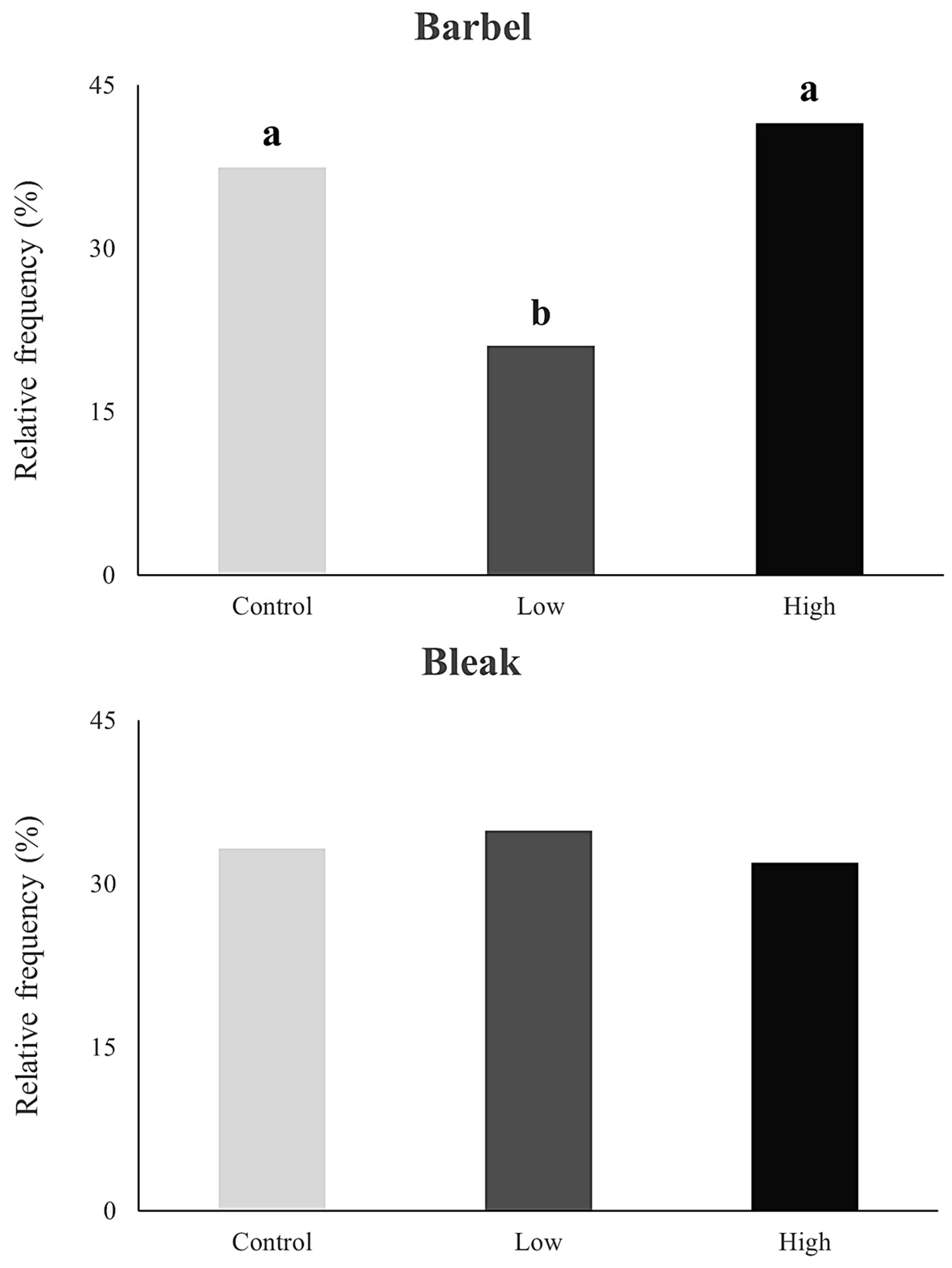
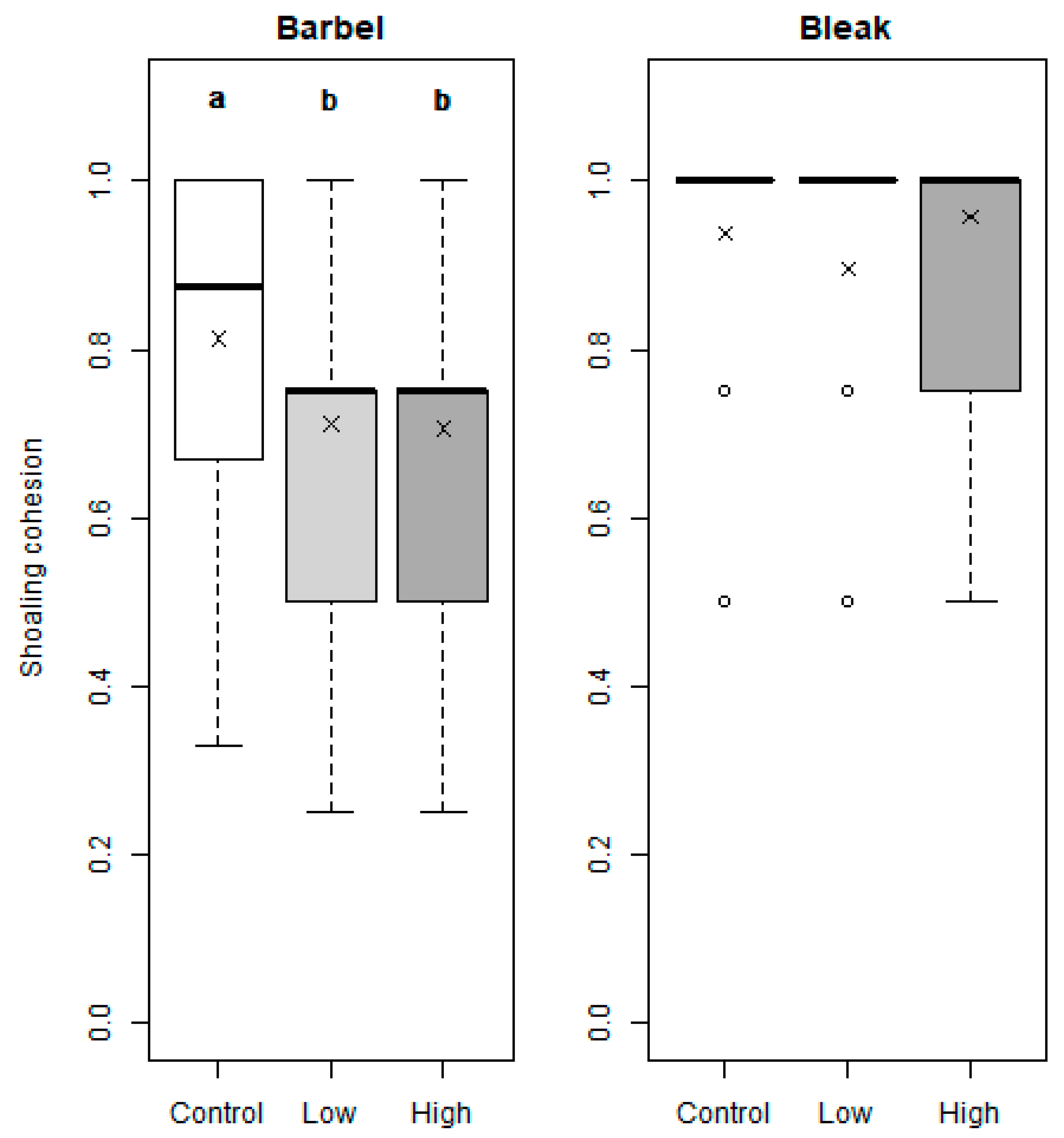
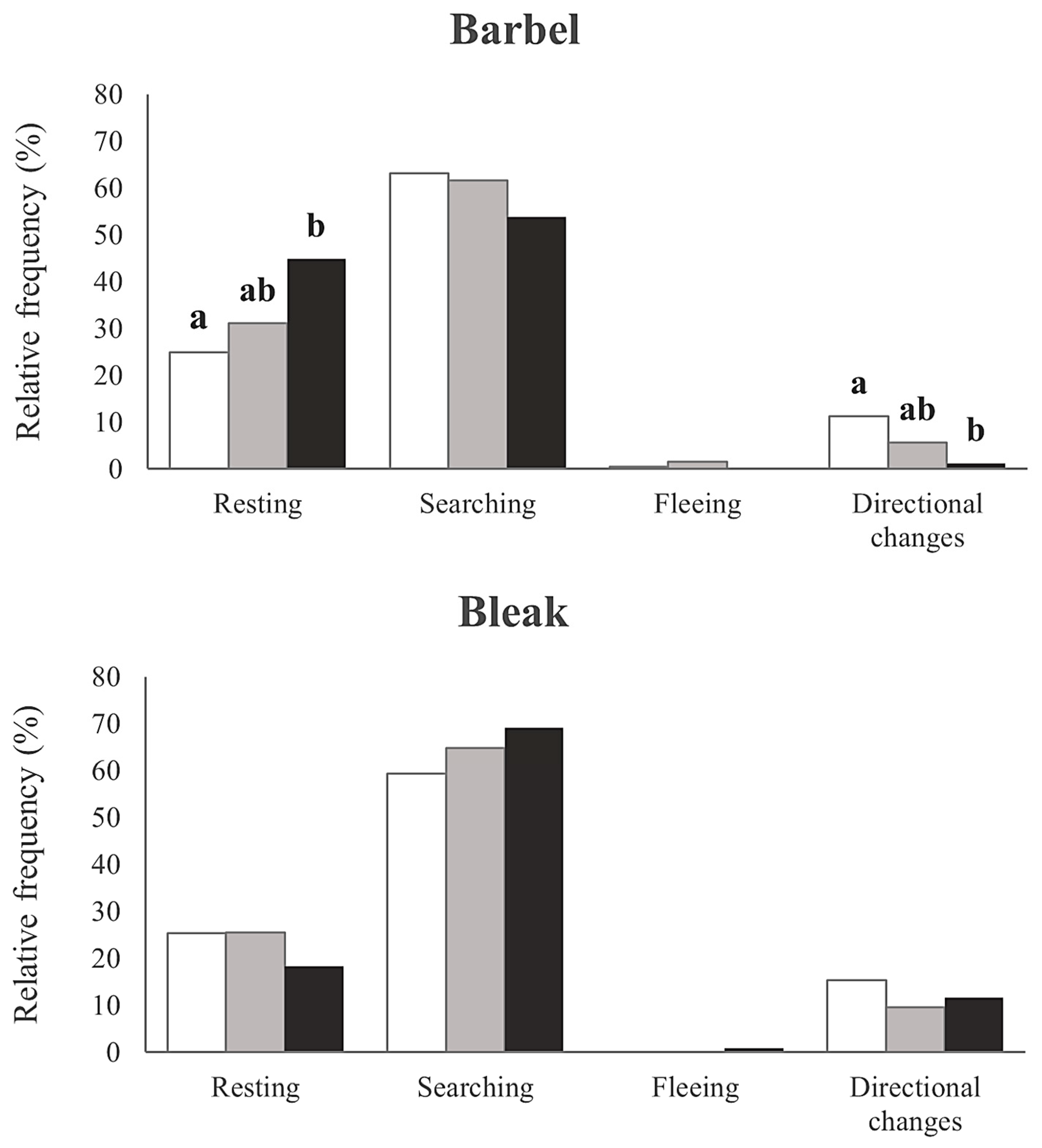
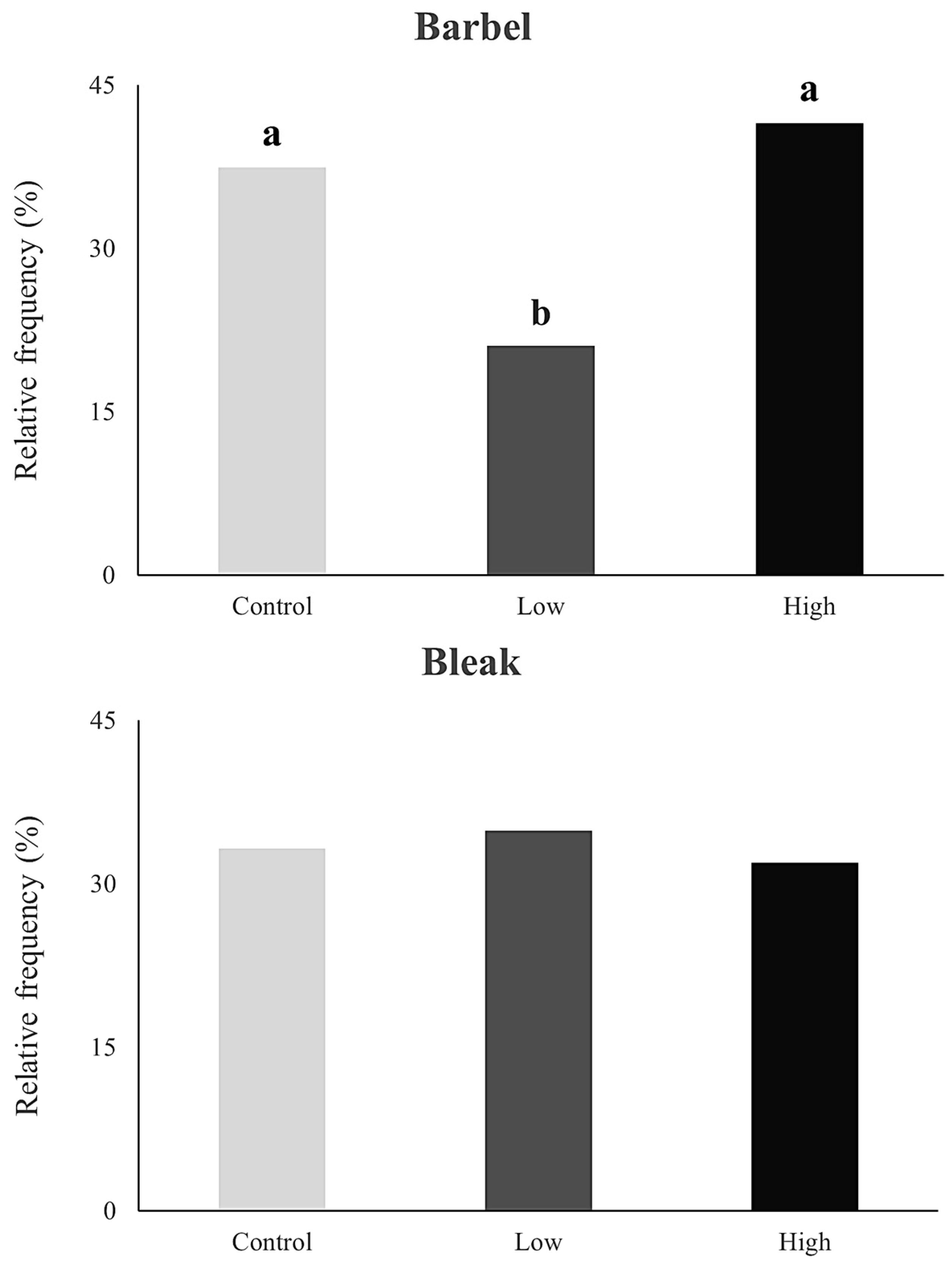
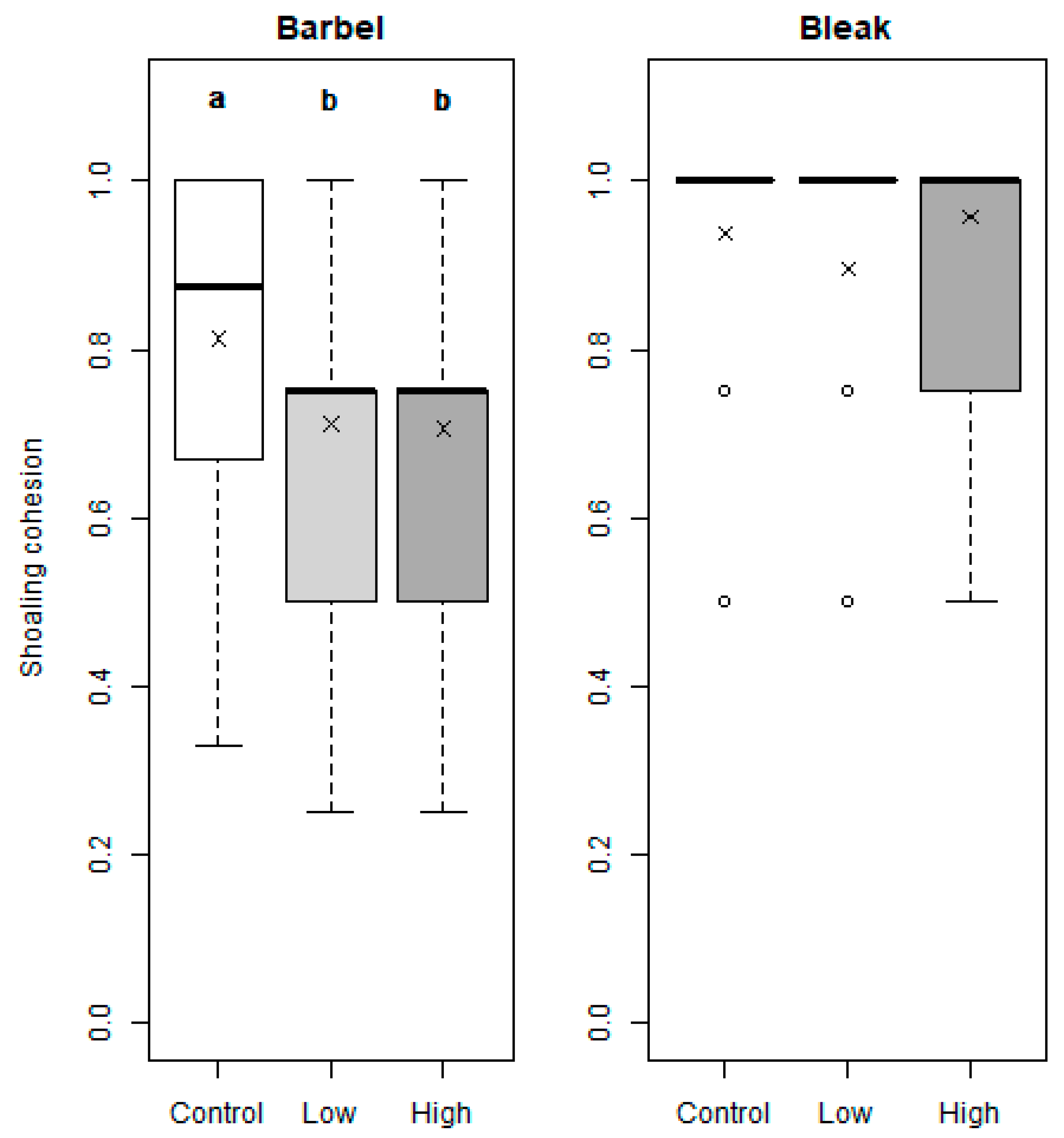
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Naiman, R.J.; Knowler, D.J.; Le, C. Freshwater Biodiversity: Importance, Threats, Status and Conservation Challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.G.; Kottelat, M.; Smith, G.R.; Stiassny, M.L.J.; Gill, A.C. So Many Fishes, So Little Time: An Overview of Recent Ichthyological Discovery in Continental Waters. Ann. Mo. Bot. Gard. 2000, 87, 26. [Google Scholar] [CrossRef]
- Valle, R.F.; Varandas, S.G.P.; Pacheco, F.A.L.; Pereira, V.R.; Santos, C.F.; Cortes, R.M.V.; Sanches, L.F. Land Use Policy Impacts of Land Use Conflicts on Riverine Ecosystems. Land Use Policy 2015, 43, 48–62. [Google Scholar] [CrossRef]
- Costa, M.J.; Duarte, G.; Segurado, P.; Branco, P. Major Threats to European Freshwater Fish Species. Sci. Total Environ. 2021, 797, 149105. [Google Scholar] [CrossRef] [PubMed]
- IUCN. IUCN Red List of Threatened Species 2020. Available online: https://www.iucnredlist.org/ (accessed on 20 June 2023).
- Xie, W.; Zhao, J.; Zhu, X.; Chen, S.; Yang, X. Pyrethroid Bioaccumulation in Wild Fish Linked to Geographic Distribution and Feeding Habit. J. Hazard. Mater. 2022, 430, 128470. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lim, W.; Song, G. Mediation of Oxidative Stress Toxicity Induced by Pyrethroid Pesticides in Fish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 234, 108758. [Google Scholar] [CrossRef]
- Brander, S.M.; Gabler, M.K.; Fowler, N.L.; Connon, R.E.; Schlenk, D. Pyrethroid Pesticides as Endocrine Disruptors: Molecular Mechanisms in Vertebrates with a Focus on Fishes. Environ. Sci. Technol. 2016, 50, 8977–8992. [Google Scholar] [CrossRef]
- Werner, I.; Geist, J.; Okihiro, M.; Rosenkranz, P.; Hinton, D.E. Effects of Dietary Exposure to the Pyrethroid Pesticide Esfenvalerate on Medaka (Oryzias latipes). Mar. Environ. Res. 2002, 54, 609–614. [Google Scholar] [CrossRef]
- Barry, M.J.; Logan, D.C.; Ahokas, J.T.; Holdway, D.A. Effects of Esfenvalerate Pulse-exposure on the Survival and Growth of Larval Australian Crimson-spotted Rainbow Fish (Melanotaenia fluviatilis). Environ. Toxicol. Water Qual. 1995, 10, 267–274. [Google Scholar] [CrossRef]
- Stehle, S.; Schulz, R. Agricultural Insecticides Threaten Surface Waters at the Global Scale. Proc. Natl. Acad. Sci. USA 2015, 112, 5750–5755. [Google Scholar] [CrossRef]
- Rosa, R.; Bordalo, M.D.; Soares, A.M.V.M.; Pestana, J.L.T. Effects of the Pyrethroid Esfenvalerate on the Oligochaete, Lumbriculus variegatus. Bull. Environ. Contam. Toxicol. 2016, 96, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Brady, J.A.; Wallender, W.W.; Werner, I.; Fard, B.M.; Zalom, F.G.; Oliver, M.N.; Wilson, B.W.; Mata, M.M.; Henderson, J.D.; Deanovic, L.A.; et al. Pesticide Runoff from Orchard Floors in Davis, California, USA: A Comparative Analysis of Diazinon and Esfenvalerate. Agric. Ecosyst. Environ. 2006, 115, 56–68. [Google Scholar] [CrossRef]
- Forbes, V.E.; Cold, A. Effects of the Pyrethroid Esfenvalerate on Life-Cycle Traits and Population Dynamics of Chironomus riparius—Importance of Exposure Scenario. Environ. Toxicol. Chem. 2005, 24, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.; Smith, S.; And, J.R.; Moore, M. Surface Water, Ground Water and Sediment Quality in Three Oxbow Lake Watersheds in the Mississippi Delta Agricultural Region: Pesticides. Int. J. Ecol. Environ. Sci. 2003, 29, 17–18. [Google Scholar]
- Affum, A.O.; Acquaah, S.O.; Osae, S.D.; Kwaansa-Ansah, E.E. Distribution and Risk Assessment of Banned and Other Current-Use Pesticides in Surface and Groundwaters Consumed in an Agricultural Catchment Dominated by Cocoa Crops in the Ankobra Basin, Ghana. Sci. Total Environ. 2018, 633, 630–640. [Google Scholar] [CrossRef]
- Ullah, S.; Li, Z.; Zuberi, A.; Arifeen, M.Z.U.; Baig, M.M.F.A. Biomarkers of Pyrethroid Toxicity in Fish. Environ. Chem. Lett. 2019, 17, 945–973. [Google Scholar] [CrossRef]
- DeMicco, A.; Cooper, K.R.; Richardson, J.R.; White, L.A. Developmental Neurotoxicity of Pyrethroid Insecticides in Zebrafish Embryos. Toxicol. Sci. 2009, 113, 177–186. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of Pyrethroid Neurotoxicity: Implications for Cumulative Risk Assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Connon, R.E.; Geist, J.; Pfeiff, J.; Loguinov, A.V.; D’Abronzo, L.S.; Wintz, H.; Vulpe, C.D.; Werner, I. Linking Mechanistic and Behavioral Responses to Sublethal Esfenvalerate Exposure in the Endangered Delta Smelt; Hypomesus transpacificus (Fam. Osmeridae). BMC Genom. 2009, 10, 608. [Google Scholar] [CrossRef]
- Baldwin, D.H.; Spromberg, J.A.; Collier, T.K.; Scholz, N.L. A Fish of Many Scales: Extrapolating Sublethal Pesticide Exposures to the Productivity of Wild Salmon Populations. Ecol. Appl. 2009, 19, 2004–2015. [Google Scholar] [CrossRef]
- Rehage, J.S.; Sih, A. Dispersal Behavior, Boldness, and the Link to Invasiveness: A Comparison of Four Gambusia Species. Biol. Invasions 2004, 6, 379–391. [Google Scholar] [CrossRef]
- Magliozzi, C.; Tsiamis, K.; Vigiak, O.; Deriu, I.; Gervasini, E.; Cardoso, A.C. Assessing Invasive Alien Species in European Catchments: Distribution and Impacts. Sci. Total Environ. 2020, 732, 138677. [Google Scholar] [CrossRef] [PubMed]
- Gurevitch, J.; Padilla, D.K. Are Invasive Species a Major Cause of Extinctions? Trends Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Mooney, H.A.; Cleland, E.E. The Evolutionary Impact of Invasive Species. Proc. Natl. Acad. Sci. USA 2001, 98, 5446–5451. [Google Scholar] [CrossRef] [PubMed]
- Yalçın Özdilek, Ş.; Partal, N.; Jones, R.I. An Invasive Species, Carassius gibelio, Alters the Native Fish Community through Trophic Niche Competition. Aquat. Sci. 2019, 81, 29. [Google Scholar] [CrossRef]
- Watts, G.; Battarbee, R.W.; Bloomfield, J.P.; Crossman, J.; Daccache, A.; Durance, I.; Elliott, J.A.; Garner, G.; Hannaford, J.; Hannah, D.M.; et al. Climate Change and Water in the UK—Past Changes and Future Prospects. Prog. Phys. Geogr. 2015, 39, 6–28. [Google Scholar] [CrossRef]
- Bond, N.; Thomson, J.; Reich, P.; Stein, J. Using Species Distribution Models to Infer Potential Climate Change-Induced Range Shifts of Freshwater Fish in South-Eastern Australia. Mar. Freshw. Res. 2011, 62, 1043–1061. [Google Scholar] [CrossRef]
- Markovic, D.; Carrizo, S.; Freyhof, J.; Cid, N.; Lengyel, S.; Scholz, M.; Kasperdius, H.; Darwall, W. Europe’s Freshwater Biodiversity under Climate Change: Distribution Shifts and Conservation Needs. Divers. Distrib. 2014, 20, 1097–1107. [Google Scholar] [CrossRef]
- Almeida, C.; Branco, P.; Segurado, P.; Ramos, T.B.; Ferreira, T.; Neves, R.; Oliveira, R.P. De Evaluation of the Trophic Status in a Mediterranean Reservoir under Climate Change: An Integrated Modelling Approach. J. Water Clim. Chang. 2021, 12, 817–832. [Google Scholar] [CrossRef]
- Jarić, I.; Lennox, R.J.; Kalinkat, G.; Cvijanović, G.; Radinger, J. Susceptibility of European Freshwater Fish to Climate Change: Species Profiling Based on Life-History and Environmental Characteristics. Glob. Chang. Biol. 2019, 25, 448–458. [Google Scholar] [CrossRef]
- Renick, V.C.; Anderson, T.W.; Morgan, S.G.; Cherr, G.N. Interactive Effects of Pesticide Exposure and Habitat Structure on Behavior and Predation of a Marine Larval Fish. Ecotoxicology 2015, 24, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Laubenstein, T.D.; Rummer, J.L.; Nicol, S.; Parsons, D.M.; Pether, S.M.J.; Pope, S.; Smith, N.; Munday, P.L. Correlated Effects of Ocean Acidification and Warming on Behavioral and Metabolic Traits of a Large Pelagic Fish. Diversity 2018, 10, 35. [Google Scholar] [CrossRef]
- Brownscombe, J.W.; Nowell, L.; Samson, E.; Danylchuk, A.J.; Steven, J. Fishing-Related Stressors Inhibit Refuge-Seeking Behavior in Released Subadult Great Barracuda. Trans. Am. Fish Soc. 2014, 143, 613–617. [Google Scholar] [CrossRef]
- Dugatkin, L.A.; Alfieri, M.S. Boldness, Behavioral Inhibition and Learning. Ethol. Ecol. Evol. 2003, 15, 43–49. [Google Scholar] [CrossRef]
- Wilson, D.S.; Coleman, K.; Clark, A.B.; Biederman, L. Shy-Bold Continuum in Pumpkinseed Sunfish (Lepomis gibbosus): An Ecological Study of a Psychological Trait. J. Comp. Psychol. 1993, 107, 250–260. [Google Scholar] [CrossRef]
- Hoover, Z.; Ferrari, M.C.O.; Chivers, D.P. The Effects of Sub-Lethal Salinity Concentrations on the Anti-Predator Responses of Fathead Minnows. Chemosphere 2013, 90, 1047–1052. [Google Scholar] [CrossRef]
- Lagesson, A.; Saaristo, M.; Brodin, T.; Fick, J.; Klaminder, J.; Martin, J.M.; Wong, B.B.M. Fish on Steroids: Temperature-Dependent Effects of 17Β-Trenbolone on Predator Escape, Boldness, and Exploratory Behaviors. Environ. Pollut. 2019, 245, 243–252. [Google Scholar] [CrossRef]
- Fraser, D.F.; Gilliam, J.F.; Daley, M.J.; Le, A.N.; Skalski, G.T. Explaining Leptokurtic Movement Distributions: Intrapopulation Variation in Boldness and Exploration. Am. Nat. 2001, 158, 124–135. [Google Scholar] [CrossRef]
- López, P.; Hawlena, D.; Polo, V.; Amo, L.; Martín, J. Sources of Individual Shy-Bold Variations in Antipredator Behaviour of Male Iberian Rock Lizards. Anim. Behav. 2005, 69, 1–9. [Google Scholar] [CrossRef]
- Magnhagen, C.; Staffan, F. Is Boldness Affected by Group Composition in Young-of-the-Year Perch (Perca fluviatilis)? Behav. Ecol. Sociobiol. 2005, 57, 295–303. [Google Scholar] [CrossRef]
- Alfonso, S.; Sadoul, B.; Gesto, M.; Joassard, L.; Chatain, B.; Geffroy, B.; Bégout, M.L. Coping Styles in European Sea Bass: The Link between Boldness, Stress Response and Neurogenesis. Physiol. Behav. 2019, 207, 76–85. [Google Scholar] [CrossRef]
- Coates, W.D.; Hale, R.; Morrongiello, J.R. Dispersal Decisions and Personality in a Freshwater Fish. Anim. Behav. 2019, 157, 209–218. [Google Scholar] [CrossRef]
- Budaev, S.V. Personality in the Guppy (Poecilia reticulata): A Correlational Study of Exploratory Behavior and Social Tendency. J. Comp. Psychol. 1997, 111, 399–411. [Google Scholar] [CrossRef]
- Budaev, S.V.; Zhuikov, A.Y. Avoidance Learning and “Personality” in the Guppy (Poecilia reticulata). J. Comp. Psychol. 1998, 112, 92–94. [Google Scholar] [CrossRef]
- Herde, A.; Eccard, J.A. Consistency in Boldness, Activity and Exploration at Different Stages of Life. BMC Ecol. 2013, 13, 49. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Donelson, M.; Fox, R.J.; Booth, D.J.; Donelson, J.M. ‘Stick with Your Own Kind, or Hang with the Locals?’ Implications of Shoaling Strategy for Tropical Reef Fish on a Range-Expansion Frontline. Glob. Chang. Biol. 2018, 24, 1663–1672. [Google Scholar] [CrossRef]
- Lemasson, B.; Tanner, C.; Woodley, C.; Threadgill, T.; Qarqish, S.; Smith, D. Motion Cues Tune Social Influence in Shoaling Fish. Sci. Rep. 2018, 8, 9785. [Google Scholar] [CrossRef]
- Chamberlain, A.C.; Ioannou, C.C. Turbidity Increases Risk Perception but Constrains Collective Behaviour during Foraging by Fish Shoals. Anim. Behav. 2019, 156, 129–138. [Google Scholar] [CrossRef]
- Romenskyy, M.; Herbert-Read, J.E.; Ioannou, C.C.; Szorkovszky, A.; Ward, A.J.W.; Sumpter, D.J.T. Quantifying the Structure and Dynamics of Fish Shoals under Predation Threat in Three Dimensions. Behav. Ecol. 2020, 31, 311–321. [Google Scholar] [CrossRef]
- Ward, A.J.W.; Hart, P.J.B. Foraging Benefits of Shoaling with Familiars May Be Exploited by Outsiders. Anim. Behav. 2005, 69, 329–335. [Google Scholar] [CrossRef]
- Camacho-Cervantes, M.; Palomera-Hernadez, V.; García, C.M. Foraging Behaviour of a Native Topminnow When Shoaling with Invaders. Aquat. Invasions 2019, 14, 490–501. [Google Scholar] [CrossRef]
- Mukherjee, I.; Bhat, A. What Drives Mixed-Species Shoaling among Wild Zebrafish? The Roles of Predators, Food Access, Abundance of Conspecifics and Familiarity. Biol. Open 2023, 12, bio059529. [Google Scholar] [CrossRef] [PubMed]
- Paijmans, K.C.; Booth, D.J.; Wong, M.Y.L. Towards an Ultimate Explanation for Mixed-Species Shoaling. Fish Fish. 2019, 20, 921–933. [Google Scholar] [CrossRef]
- Santos, R.G.; Pinheiro, H.T.; Martins, A.S.; Riul, P.; Bruno, S.C.; Janzen, F.J.; Ioannou, C.C. The Anti-Predator Role of within-Nest Emergence Synchrony in Sea Turtle Hatchlings. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160697. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.; Gerlai, R. Quantification of Shoaling Behaviour in Zebrafish (Danio rerio). Behav. Brain Res. 2007, 184, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Archard, G.A.; Braithwaite, V.A. Increased Exposure to Predators Increases Both Exploration and Activity Level in Brachyrhaphis episcopi. J. Fish Biol. 2011, 78, 593–601. [Google Scholar] [CrossRef]
- Baldwin, D.S.; Rees, G.N.; Mitchell, A.M.; Watson, G.; Williams, J. The Short-Term Effects of Salinization on Anaerobic Nutrient Cycling and Microbial Community Structure in Sediment from a Freshwater Wetland. Wetlands 2006, 26, 455–464. [Google Scholar] [CrossRef]
- Geist, J.; Werner, I.; Eder, K.J.; Leutenegger, C.M. Comparisons of Tissue-Specific Transcription of Stress Response Genes with Whole Animal Endpoints of Adverse Effect in Striped Bass (Morone saxatilis) Following Treatment with Copper and Esfenvalerate. Aquat. Toxicol. 2007, 85, 28–39. [Google Scholar] [CrossRef]
- Epstein, L.; Bassein, S.; Zalom, F.G. Almond and Stone Fruit Growers Reduce OP, Increase Pyrethroid Use in Dormant Sprays. Calif. Agric. 2000, 54, 14–19. [Google Scholar] [CrossRef]
- Palmquist, K.; Salatas, J.; Fairbrother, A. Pyrethroid Insecticides: Use, Environmental Fate, and Ecotoxicology. In Insecticides-Advances in Integrated Pest Management; Perveen, F., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 251–278. ISBN 978-953-307-780-2. [Google Scholar]
- Floyd, E.Y.; Geist, J.P.; Werner, I. Acute, Sublethal Exposure to a Pyrethoid Insecticide Alter Behavior, Growth, and Predation Risk in Larvae of the Fathead Minnow (Pimephales promelas). Environ. Toxicol. Chem. 2008, 27, 1780–1787. [Google Scholar] [CrossRef]
- Collares-Pereira, M.J.; Alves, M.J.; Ribeiro, F.; Domingos, I.; Almeida, P.R.; da Costa, L.; Gante, H.; Filipe, A.F.; Aboim, M.A.; Rodrigues, P.M.; et al. Guia Dos Peixes de Água Doce e Migradores de Portugal Continental; Edições Afrontamento: Porto, Portugal, 2021. [Google Scholar]
- Schönhuth, S.; Vukić, J.; Šanda, R.; Yang, L.; Mayden, R.L. Phylogenetic Relationships and Classification of the Holarctic Family Leuciscidae (Cypriniformes: Cyprinoidei). Mol. Phylogenet. Evol. 2018, 127, 781–799. [Google Scholar] [CrossRef] [PubMed]
- Latorre, D.; Masó, G.; Cano-Barbacil, C.; Zamora-Marin, J.M.; Almeida, D.; Vilizzi, L.; Britton, J.R.; Cruz, A.; Fernández-Delgado, C.; González-Rojas, A.G.; et al. A Review and Meta-Analysis of the Environmental Biology of Bleak Alburnus alburnus in Its Native and Introduced Ranges, with Reflections on Its Invasiveness. Rev. Fish Biol. Fish. 2023. [Google Scholar] [CrossRef]
- Miranda, R.; Oscoz, J.; Leunda, P.M.; Escala, M.C. Weight-Length Relationships of Cyprinid Fishes of the Iberian Peninsula. J. Appl. Ichtyol. 2006, 22, 297–298. [Google Scholar] [CrossRef]
- Masó, G.; Latorre, D.; Tarkan, A.S.; Vila-Gispert, A.; Almeida, D. Inter-Population Plasticity in Growth and Reproduction of Invasive Bleak, Alburnus alburnus (Cyprinidae, Actinopterygii), in Northeastern Iberian Peninsula. Folia Zool. 2016, 65, 10–14. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Ferreira, A.P.; Ferreira, M.T. Intrabasin Variations in Age and Growth of Barbus bocagei Populations. J. Appl. Ichthyol. 2002, 18, 134–139. [Google Scholar] [CrossRef]
- Gonino, G.; Branco, P.; Benedito, E.; Teresa, M.; Santos, J.M. Short-Term Effects of Wild Fire Ash Exposure on Behaviour and Hepatosomatic Condition of a Potamodromous Cyprinid Fish, the Iberian Barbel Luciobarbus bocagei ( Steindachner, 1864). Sci. Total Environ. 2019, 665, 226–234. [Google Scholar] [CrossRef]
- Leite, T.; Santos, J.M.; Ferreira, M.T.; Canhoto, C.; Branco, P. Does Short-Term Salinization of Freshwater Alter the Behaviour of the Iberian Barbel (Luciobarbus bocagei, Steindachner 1864)? Sci. Total Environ. 2019, 651, 648–655. [Google Scholar] [CrossRef]
- Mameri, D.; Branco, P.; Ferreira, M.T.; Santos, J.M. Heatwave Effects on the Swimming Behaviour of a Mediterranean Freshwater Fish, the Iberian Barbel Luciobarbus bocagei. Sci. Total Environ. 2020, 730, 139152. [Google Scholar] [CrossRef]
- Stammler, K.L.; Corkum, L.D. Assessment of Fish Size on Shelter Choice and Intraspecific Interactions by Round Gobies Neogobius melanostomus. Environ. Biol. Fishes 2005, 73, 117–123. [Google Scholar] [CrossRef]
- Leite, T.; Branco, P.; Ferreira, M.T.; Santos, J.M. Activity, Boldness and Schooling in Freshwater Fish Are Affected by River Salinization. Sci. Total Environ. 2022, 819, 153046. [Google Scholar] [CrossRef]
- Manek, A.K.; Ferrari, M.C.O.; Niyogi, S.; Chivers, D.P. The Interactive Effects of Multiple Stressors on Physiological Stress Responses and Club Cell Investment in Fathead Minnows. Sci. Total Environ. 2014, 476–477, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging Threats and Persistent Conservation Challenges for Freshwater Biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.J.; Deng, D.; Werner, I.; Teh, F.; Hung, S.S.O. Sublethal Toxicity of Orchard Stormwater Runoff in Sacramento Splittail (Pogonichthys macrolepidotus) Larvae. Mar. Environ. Res. 2005, 59, 203–216. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, D.J.; Shingles, A.; Taylor, E.W. Sub-Lethal Plasma Ammonia Accumulation and the Exercise Performance of Salmonids. In Comparative Biochemistry and Physiology—A Molecular and Integrative Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2003; Volume 135, pp. 515–526. [Google Scholar]
- Werner, I.; Schneeweiss, A.; Segner, H.; Junghans, M. Environmental Risk of Pesticides for Fish in Small-and Medium-Sized Streams of Switzerland. Toxics 2012, 9, 79. [Google Scholar] [CrossRef]
- Markovic, D.; Carrizo, S.F.; Kärcher, O.; Walz, A.; David, J.N.W. Vulnerability of European Freshwater Catchments to Climate Change. Glob. Chang. Biol. 2017, 23, 3567–3580. [Google Scholar] [CrossRef]
- Bevelhimer, M.; Bennett, W. Assessing Cumulative Thermal Stress in Fish during Chronic Intermittent Exposure to High Temperatures. Environ. Sci. Policy 2000, 3, 211–216. [Google Scholar] [CrossRef]
- Colchen, T.; Teletchea, F.; Fontaine, P.; Pasquet, A. Temperature Modifies Activity, Inter-Individual Relationships and Group Structure in a Fish. Curr. Zool. 2017, 63, 175–183. [Google Scholar] [CrossRef]
- Le Pichon, C.; Lestel, L.; Courson, E.; Merg, M.L.; Tales, E.; Belliard, J. Historical Changes in the Ecological Connectivity of the Seine River for Fish: A Focus on Physical and Chemical Barriers since the Mid-19th Century. Water 2020, 12, 1352. [Google Scholar] [CrossRef]
- Lan, T.T.P.; Hien, T.T.T.; Le Cam Tu, T.; Van Khanh, N.; Haga, Y.; Phu, T.M. Salinization Intensifies the Effects of Elevated Temperatures on Channa striata, a Common Tropical Freshwater Aquaculture Fish in the Mekong Delta, Vietnam. Fish. Sci. 2020, 86, 1029–1036. [Google Scholar] [CrossRef]
- Searle, C.L.; Shaw, C.L.; Hunsberger, K.K.; Prado, M.; Duffy, M.A. Salinization Decreases Population Densities of the Freshwater Crustacean, Daphnia dentifera. Hydrobiologia 2016, 770, 165–172. [Google Scholar] [CrossRef]
- Cañedo-Argüelles, M. A Review of Recent Advances and Future Challenges in Freshwater Salinization. Limnetica 2020, 39, 185–211. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Clim. Chang. 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Liebergesell, M.; Reu, B.; Stahl, U.; Freiberg, M.; Welk, E.; Kattge, J.; Cornelissen, J.H.C.; Peãuelas, J.; Wirth, C. Functional Resilience against Climate-Driven Extinctions—Comparing the Functional Diversity of European and North American Tree Floras. PLoS ONE 2016, 11, e0148607. [Google Scholar] [CrossRef] [PubMed]
- Domenici, P.; Seebacher, F. The Impacts of Climate Change on the Biomechanics of Animals. Conserv. Physiol. 2020, 8, coz102. [Google Scholar] [CrossRef] [PubMed]
- Little, E.E.; Dwyer, F.J.; Fairchild, J.F.; Delonay, A.J.; Zajicek, J.L. Survival of Bluegilll and Their Behavioural Responses during Continuous and Pulsed Exposures to Esfenvalerate, a Pyrethroid Insecticide. Environ. Toxicol. Chem. 1993, 12, 871–878. [Google Scholar] [CrossRef]
- Sih, A.; Mathot, K.J.; Moirón, M.; Montiglio, P.O.; Wolf, M.; Dingemanse, N.J. Animal Personality and State-Behaviour Feedbacks: A Review and Guide for Empiricists. Trends Ecol. Evol. 2015, 30, 50–60. [Google Scholar] [CrossRef]
- Harcourt, J.L.; Sweetman, G.; Johnstone, R.A.; Manica, A. Personality Counts: The Effect of Boldness on Shoal Choice in Three-Spined Sticklebacks. Anim. Behav. 2009, 77, 1501–1505. [Google Scholar] [CrossRef]
- Sih, A.; Cote, J.; Evans, M.; Fogarty, S.; Pruitt, J. Ecological Implications of Behavioural Syndromes. Ecol Lett 2012, 15, 278–289. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Réale, D. Natural Selection and Animal Personality. Behaviour 2005, 142, 1159–1184. [Google Scholar] [CrossRef]
- Réale, D.; Garant, D.; Humphries, M.M.; Bergeron, P.; Careau, V.; Montiglio, P.O. Personality and the Emergence of the Pace-of-Life Syndrome Concept at the Population Level. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 4051–4063. [Google Scholar] [CrossRef]
- Budaev, S.V. Alternative Styles in the European Wrasse, Symphodus ocellatus: Boldness-Related Schooling Tendency. Environ. Biol. Fishes 1997, 49, 71–78. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Wright, J.; Kazem, A.J.N.; Thomas, D.K.; Hickling, R.; Dawnay, N. Behavioural Syndromes Differ Predictably between 12 Populations of Three-Spined Stickleback. J. Anim. Ecol. 2007, 76, 1128–1138. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Van der Plas, F.; Wright, J.; Réale, D.; Schrama, M.; Roff, D.A.; Van der Zee, E.; Barber, I. Individual Experience and Evolutionary History of Predation Affect Expression of Heritable Variation in Fish Personality and Morphology. Proc. R. Soc. B Biol. Sci. 2009, 276, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.; Matono, P.; Barata, E.N.; Bernardo, J.M.; Costa, A.M.; Ilhéu, M. Behavioural Interactions between the Endangered Native Fish Saramugo, Anaecypris hispanica, and the Invasive Bleak, Alburnus alburnus. Limnetica 2019, 38, 517–533. [Google Scholar] [CrossRef]
- Matono, P.; da Silva, J.; Ilhéu, M. How Does an Invasive Cyprinid Benefit from the Hydrological Disturbance of Mediterranean Temporary Streams? Diversity 2018, 10, 47. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides Reduce Regional Biodiversity of Stream Invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef]
- Fleeger, J.W.; Carman, K.R.; Nisbet, R.M. Indirect Effects of Contaminants in Aquatic Ecosystems. Sci. Total Environ. 2003, 317, 207–233. [Google Scholar] [CrossRef]
- Macneale, K.H.; Kiffney, P.M.; Scholz, N.L. Pesticides, Aquatic Food Webs, and the Conservation of Pacific Salmon. Front. Ecol. Environ. 2010, 8, 475–482. [Google Scholar] [CrossRef]
- Cote, J.; Clobert, J. Social Personalities Influence Natal Dispersal in a Lizard. Proc. R. Soc. B Biol. Sci. 2007, 274, 383–390. [Google Scholar] [CrossRef]
- Sih, A.; Ferrari, M.C.O.; Harris, D.J. Evolution and Behavioural Responses to Human-Induced Rapid Environmental Change. Evol. Appl. 2011, 4, 367–387. [Google Scholar] [CrossRef]
- Smith, B.R.; Blumstein, D.T. Fitness Consequences of Personality: A Meta-Analysis. Behav. Ecol. 2008, 19, 448–455. [Google Scholar] [CrossRef]
- Hooke, J.M. Human Impacts on Fluvial Systems in the Mediterranean Region. Geomorphology 2006, 79, 311–335. [Google Scholar] [CrossRef]
- Boix, D.; García-Berthou, E.; Gascón, S.; Benejam, L.; Tornés, E.; Sala, J.; Benito, J.; Munné, A.; Solà, C.; Sabater, S. Response of Community Structure to Sustained Drought in Mediterranean Rivers. J. Hydrol. 2010, 383, 135–146. [Google Scholar] [CrossRef]
- Grzybowski, M.; Glińska-Lewczuk, K. Principal Threats to the Conservation of Freshwater Habitats in the Continental Biogeographical Region of Central Europe; Springer: Dordrecht, The Netherlands, 2019; Volume 28, ISBN 0123456789. [Google Scholar]
- Segurado, P.; Ferreira, T.; Branco, P. Assessing the Effects of Multiple Stressors on Aquatic Systems across Temporal and Spatial Scales: From Measurement to Management. Water 2021, 13, 3549. [Google Scholar] [CrossRef]
- Segurado, P.; Santos, J.M.; Pont, D.; Melcher, A.H.; Jalon, D.G.; Hughes, R.M.; Ferreira, M.T. Estimating Species Tolerance to Human Perturbation: Expert Judgment versus Empirical Approaches. Ecol. Indic. 2011, 11, 1623–1635. [Google Scholar] [CrossRef]
- Aguiar, T.R.; Bortolozo, F.R.; Hansel, F.A.; Rasera, K.; Ferreira, M.T. Riparian Buffer Zones as Pesticide Filters of No-till Crops. Environ. Sci. Pollut. Res. 2015, 22, 10618–10626. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control (0.0 μg/L) | Low (1.2 μg/L) | High (2.0 μg/L) |
|---|---|---|---|
| Barbel (Luciobarbus bocagei) | |||
| Fish length (TL, mm) | 145.3 ± 17.1 | 138.8 ± 21.3 | 146.0 ± 14.8 |
| Fish body mass (g) | 29.81 ±11.51 | 28.33 ±9.36 | 30.75 ± 9.22 |
| Water temperature (°C) | 17.37 ± 0.21 | 17.37 ± 0.06 | 17.57 ± 0.06 |
| pH | 7.92 ± 0.11 | 7.79 ± 0.33 | 7.96 ± 0.06 |
| Conductivity (mS/cm) | 0.86 ± 0.10 | 0.86 ± 0.08 | 0.80 ± 0.02 |
| Bleak (Alburnus alburnus) | |||
| Fish length (TL, mm) | 132.3 ± 11.6 | 126.3 ± 10.2 | 128.6 ± 12.8 |
| Fish body mass (g) | 15.00 ± 3.95 | 14.08 ± 3.42 | 14.58 ± 3.85 |
| Water temperature (°C) | 18.43 ± 0.72 | 18.43 ± 0.67 | 18.63 ± 0.85 |
| pH | 7.93 ± 0.03 | 7.93 ± 0.03 | 7.93 ± 0.02 |
| Conductivity (mS/cm) | 0.81 ± 0.01 | 0.81 ± 0.00 | 0.82 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, T.; Mameri, D.; Branco, P.; Vieira, I.; Oliveira, M.; Santos, J.M. Swimming under Pressure: The Sub-Lethal Effects of a Pesticide on the Behaviour of Native and Non-Native Cypriniformes Fish. Fishes 2023, 8, 462. https://doi.org/10.3390/fishes8090462
Leite T, Mameri D, Branco P, Vieira I, Oliveira M, Santos JM. Swimming under Pressure: The Sub-Lethal Effects of a Pesticide on the Behaviour of Native and Non-Native Cypriniformes Fish. Fishes. 2023; 8(9):462. https://doi.org/10.3390/fishes8090462
Chicago/Turabian StyleLeite, Tamara, Daniel Mameri, Paulo Branco, Inês Vieira, Margarida Oliveira, and José Maria Santos. 2023. "Swimming under Pressure: The Sub-Lethal Effects of a Pesticide on the Behaviour of Native and Non-Native Cypriniformes Fish" Fishes 8, no. 9: 462. https://doi.org/10.3390/fishes8090462
APA StyleLeite, T., Mameri, D., Branco, P., Vieira, I., Oliveira, M., & Santos, J. M. (2023). Swimming under Pressure: The Sub-Lethal Effects of a Pesticide on the Behaviour of Native and Non-Native Cypriniformes Fish. Fishes, 8(9), 462. https://doi.org/10.3390/fishes8090462