
Sex Differences in Fatty Acid Composition of Chinese Tongue Sole (Cynoglossus semilaevis) Tissues


1
State Key Laboratory of Mariculture Biobreeding and Sustainable Goods, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
2
College of Fisheries and Life Science, Dalian Ocean University, Dalian 116023, China
*
Author to whom correspondence should be addressed.
Fishes 2023, 8(8), 421; https://doi.org/10.3390/fishes8080421
Submission received: 13 July 2023
/
Revised: 5 August 2023
/
Accepted: 16 August 2023
/
Published: 17 August 2023
(This article belongs to the Section Physiology and Biochemistry)
Abstract
:The fatty acid accumulation in fish tissues are closely related to fish sex, yet the sex differences in fish fatty acids have not been fully known. This experiment investigated the sex differences in fatty acid composition of Chinese tongue sole (Cynoglossus semilaevis), which have typical sexual dimorphism and reversal. Three sexes, namely females, males and pseudomales, were involved in this study. Five batches of fish samples were collected from different fish farms in Shandong Province, China. These fish were identified to be at the gonadal development stage of III–IV. The disparity of fatty acid between sexes (female vs male and pseudomale vs. male) was evaluated by the distance coefficient (Dsex) and multi-variate similarity of percentages analysis (SIMPER). The highest Dsex values between females and males were found in gonad, followed by liver and eye. The fatty acids that differed between females and males were mainly DHA, 16:1n-7, 18:1n-9, and 18:0. The sex difference in DHA and 18:1n-9 contributed significantly to the overall sex difference in fatty acid of brain, eye, and gonad, whereas that in 16:1n-7 contributed significantly in heart, gill, liver, and muscle. The sex difference in 18:0 contributed significantly to the overall sex difference in fatty acid of heart, gill, gut, and skin. The sex differences in fatty acid between pseudomales and males were generally insignificant. In conclusion, at this gonadal development stage (III–IV), the sex differences in fatty acid between female and male Chinese tongue sole have emerged, mainly in the gonad. The testes had higher DHA levels but lower levels of 16:1n-7 and 18:1n-9 than ovaries. In addition, for pseudomales, the phenotypic sex more substantially determined the fatty acid composition than the genetic sex.
Key Contribution: This study revealed the sex differences in fatty acids between different sexes, namely, females, males and pseudomales, of a flatfish species, which have typical sexual dimorphism and reversal.
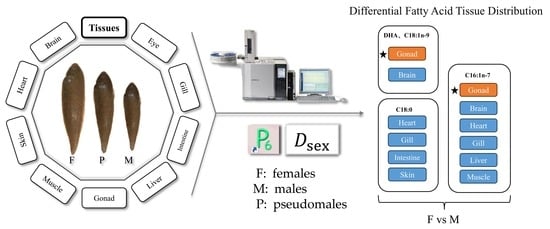
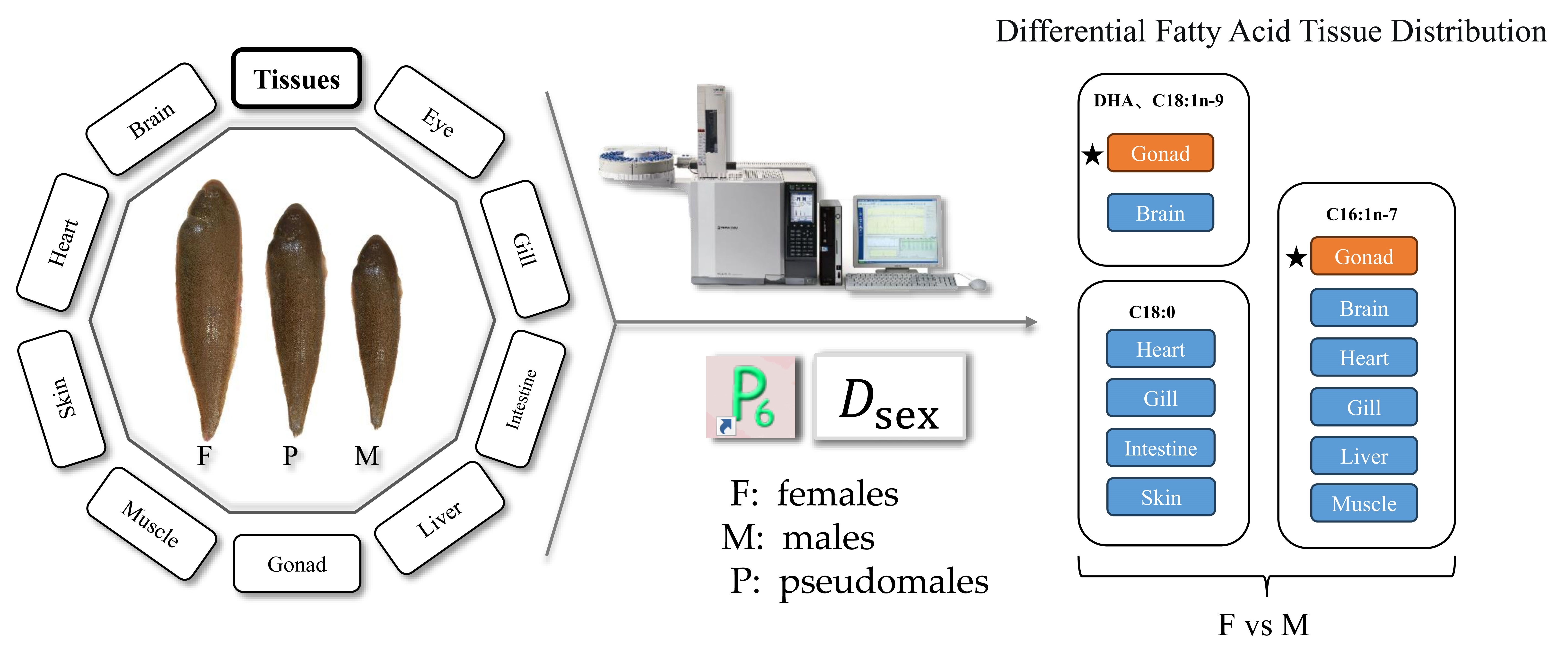
1. Introduction
Fatty acids are essential nutrients for aquatic animals and play important roles in energy supply and other physiological processes such as maintenance of cell brane fluidity, immune response and reproduction [1,2,3,4,5,6]. During fish development, fatty acids are deposited selectively in different tissues according to their function, especially in the gonads during the reproductive periods [7,8,9]. Considering the different characteristics between ovaries and testes regarding cell structure, energy requirement, and gametogenesis, it is reasonable to assume that the fatty acid deposition probably largely differs between sexes.
Indeed, several studies have shown the selective deposition of fatty acids depending on sex, season and reproductive cycle [10,11,12,13,14]. The selective deposition of fatty acids in fish of specific sexes may reflect the specific fatty acid preference and utilization. The relevant mechanisms involved have not been well-known. The study on the selective fatty acids deposition in fish of different sexes may not only enrich our knowledge in this area, but also provide useful information for management of sex-specific feeds for fish broodstocks.
Chinese tongue sole (Cynoglossus semilaevis) is an economically important aquaculture species in Northern China. This species has a typical sexual dimorphism. Males grow slowly, whereas females grow about four times faster than males [15,16]. Also, females can be sexually transformed into pseudomales during the juvenile stage, breaking the normal male/female ratio and leading to a reduction in the number of females [17]. These characteristics make Chinese tongue sole a good model species for studies on sex difference in fatty acid. The present study aims to comprehensively investigate the differences in fatty acid among different sexes of Chinese tongue sole, namely, female, male, and pseudomale. The results may enhance our knowledge about the sex difference in fish fatty acids.
2. Materials and Methods
2.1. Experiment Fish
Five batches of experiment fish were collected from five different fish farms in Hongdao (HD), Rizhao (RZ), and Weifang (three batches: WF1, WF2 and WF3), Shandong Province. Each batch had 30 healthy fish. All the fish were nearly 11 months old. The average weight of the female fish, male fish, and pseudomale fish of all batches was 182.48 g, 124.94 g, and 138.89 g, respectively. Fish were transported to the laboratory in bags inflated with oxygen and water since purchased. After being anesthetized with MS-222, the body weight, body length, and liver weight were measured to calculate the hepatosomatic index (HSI) and condition factor (CF). The fish were then dissected on ice and a total of 10 tissues (fin, brain, eye, gill, heart, liver, intestine, gonad, skin, and muscle) were collected. The samples were stored at −20 °C before used. All sampling protocols, as well as all fish handling processes, in this study were reviewed and approved by the Animal Care and Use Committee of Yellow Sea Fisheries Research Institute.
2.2. Sex Identification
The fin DNA was extracted and the genetic sex was identified by the Marine Animal Tissue Genome DNA Extraction kit (TIANGEN, Beijing, China). A total of 5 μL DNA template was obtained for the subsequent PCR study. Sex-specific markers (F: CCTAAATGATGGATGTAGATTCTGTC, R: GATCCAGAGAAAATAAACCCAGG) were synthesized by Tsingke Biotechnology Co., Ltd. (Qingdao, China). The system of Polymerase Chain Reaction (PCR) is shown in Table 1. The amplified fragment length was detected with 2% agarose gel at 180 v and 30 min. At the end of electrophoresis, the bands were checked under ultraviolet light. The genetic males had one band (169 bp) and the genetic females had two bands (169 bp/134 bp) (see examples in Figure 1). The biological sex of each fish was determined by observational check. The testes were grayish white in appearance, and a small amount of white sperm can be seen. Under the microscope, the spermatic sac cannot be seen and the spermatic lobule was filled with white sperm. The testes development stage was around IV. The ovaries were translucent and light red, and small blood vessels can be seen on the surface. The eggs cannot be clearly seen by naked eye. The ovaries development stage was III–IV. For pseudomale fish, the genetic sex was female and the biological sex was male. Based on the check of fish biological and genetic sex, the sex identification results are shown in Table 2.
2.3. Analysis of Total Lipid and Fatty Acid Compositions
Three randomly selected fish from a same sex of each group were used for the analysis of total lipid content and fatty acid composition. The total lipid of the liver, muscle and gonad samples was extracted and analyzed with the chloroform: methanol (2:1 v/v) method [18]. The fatty acid compositions were analyzed with gas chromatograph (GC-2010 pro, Shimadzu, Japan). Briefly, fatty acids in wet samples (about 0.5 g) (the fatty acids in the liver, muscle, and gonads are derived from lipid (about 50 μL) extracted by the chloroform methanol method) were esterified first with KOH-methanol and then with BF3-methanol on 75 °C water bath. Fatty acid methyl esters were extracted with hexane and then subjected to the gas chromatography equipped with a fused silica capillary column (SH-RT-2560, 100 m × 0.25 mm × 0.20 μm) and a flame ionization detector. The column temperature increase was programmed: from 100 °C up to 190 °C at a rate of 10 °C min−1; then from 190 °C to 200 °C at a rate of 0.3 °C min−1; and then from 200 °C to 230 °C at a rate of 4 °C min−1. Both injector and detector temperature were 230 °C. Temperature of flame-ionization detector, injector, and column was 300 °C, 250 °C, and 100 °C, respectively. The flow rate of the carrier gas helium was 3 mL/min. For each sample, a total of 1 μL was injected into the machine. The split ratio was 50%. A mixture of 37 standard reference fatty acids (Solarbio, Beijing, China) was used to identify the fatty acid peaks. The results were expressed as % total fatty acids (%TFA, % peak areas).
2.4. Calculations and Statistical Methods
Formula for calculating the hepatosomatic index: HSI (%) = wet liver weight/fish body weight × 100
Formula for calculating the condition factor: CF = body weight/(body length3) × 100
The inter-sex distance coefficient (Dsex) is used to indicate the difference in fatty acid composition between different sexes [19]. The Dsex is calculated according to the following equation [12]:
where Dsex is the distance coefficient between the female sample f and the male sample m; Pif and Pim are contents of fatty acid i in the sample f and m, respectively. For each fish batch, an average content was used for each fatty acid in each tissue. One Dsex value was returned for each tissue. In general, the Dsex represents the geometrical distance, or disparity, between the fatty acid composition of females and males. The Dsex between males and pseudomales was calculated in similar way.
Besides, the multi-variate similarity of percentages analysis (SIMPER) was also used to analyze the fatty acid dissimilarities between sexes [12,20]. The fatty acid compositions were pretreated with Prime-E (Version 6.1.10, Plymouth Routines in Multivariate Ecological Research, Ivybridge, UK). Additionally, the relative contribution of each individual fatty acid’s sex difference to the total sex difference in fatty acid was also evaluated by SIMPER.
For other data, one-way ANOVA in SPSS 16.0 for Windows were used. Tukey’s multiple range test was used to detect the significant difference between the means. There was significant difference when p ˂ 0.05. The data of female and male fish in the WF3 group were analyzed using an independent sample t-test. The results are presented as means ± standard error.
3. Results
3.1. Somatic Indicators and Total Lipid Content
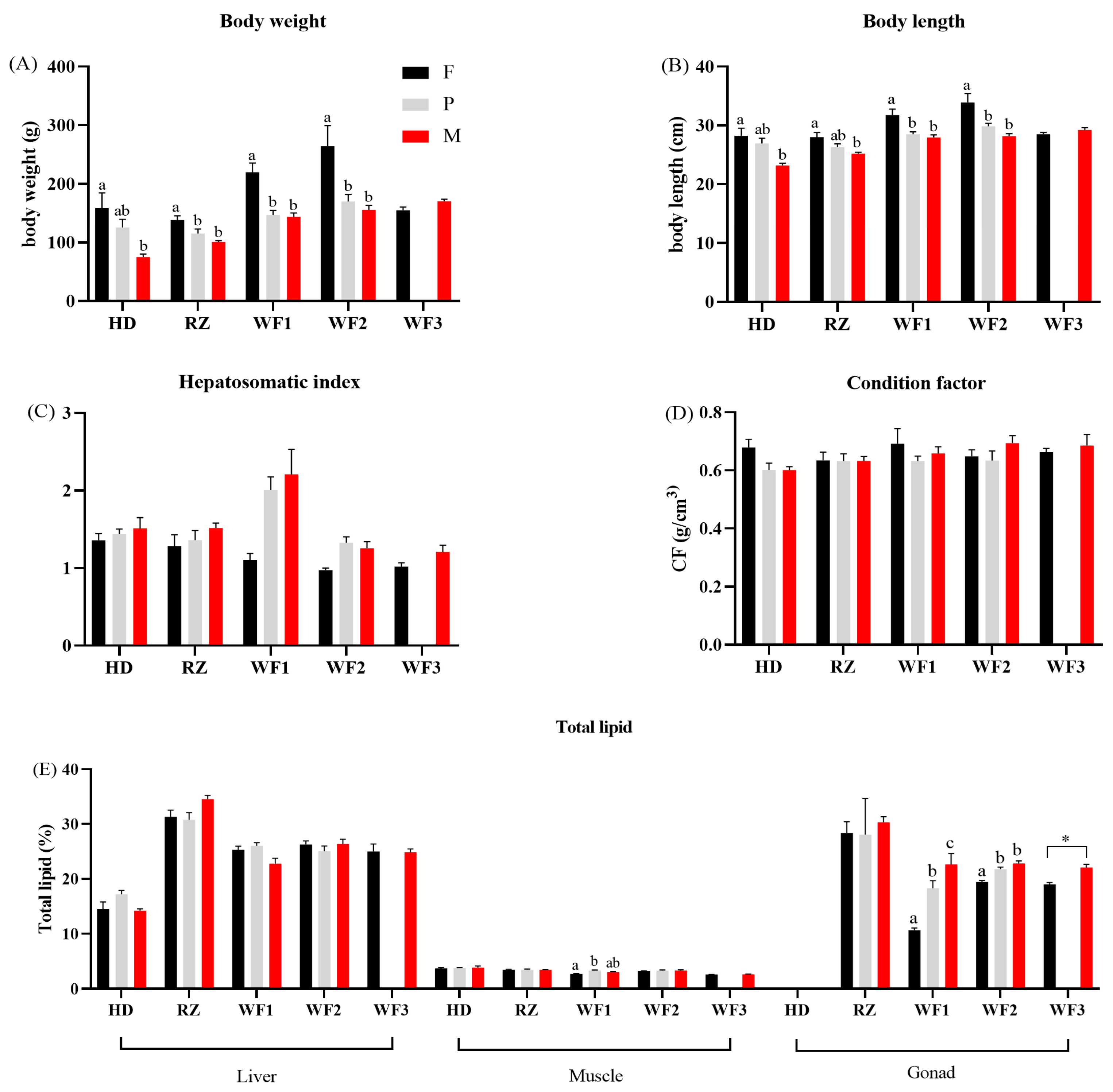
In four batch groups (HD, RZ, WF1 and WF2), female fish had the highest body weight, followed by pseudomales, and males had the lowest body weight (Figure 2, Supplementary Table S1). In general, females had significantly (p < 0.05) higher body weight than males. The fish body length followed similar trends to the body weight. However, the hepatosomatic index (HSI) had an opposite trend. The males had the highest HSI values, whereas the females had the lowest ones. In the WF1 and WF2 groups, significant differences were observed between and females and pseudomales/males (p < 0.05). There was no significant difference (p > 0.05) in condition factor among sexes in all fish batches. The differences in total lipid content among sexes were observed mostly in gonads. It was unexpected that testes had significantly (p < 0.05) higher lipid content than ovaries, with pseudomales having intermediate values.
3.2. Dsex Values
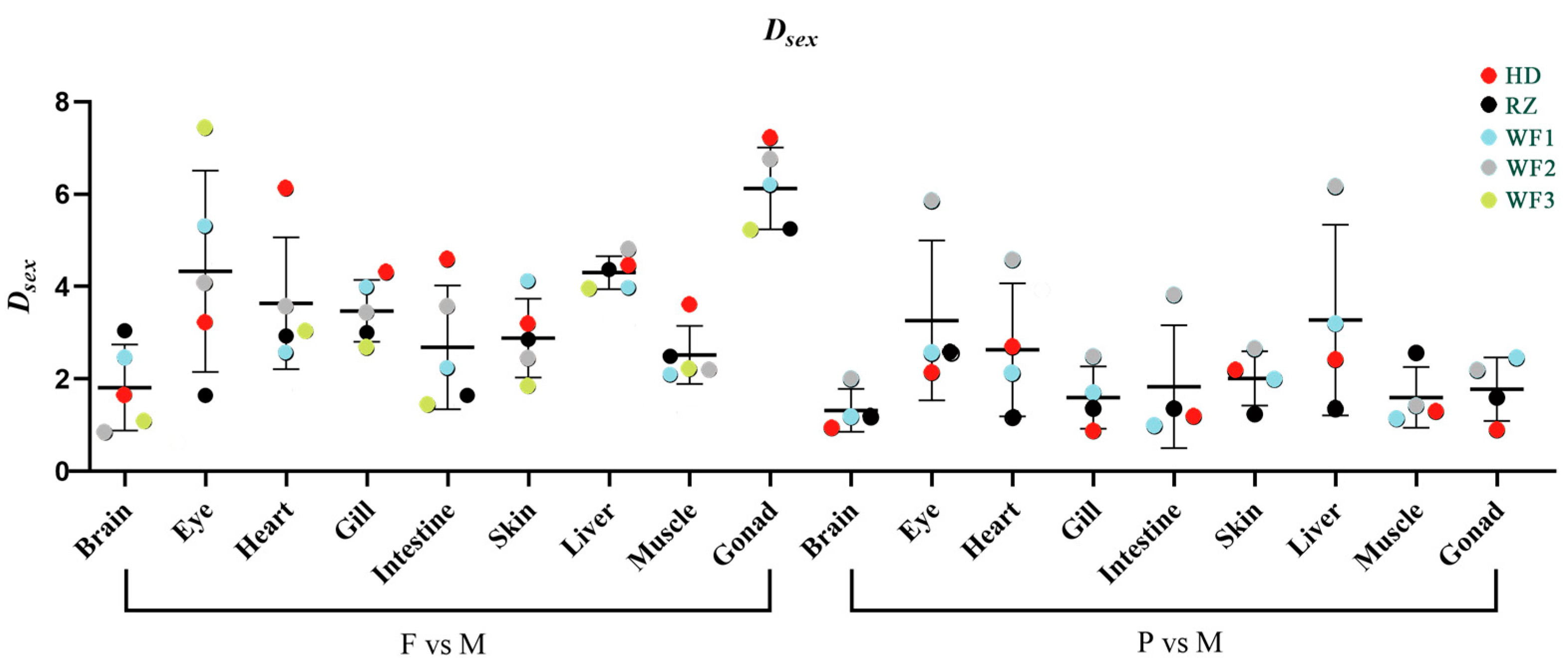
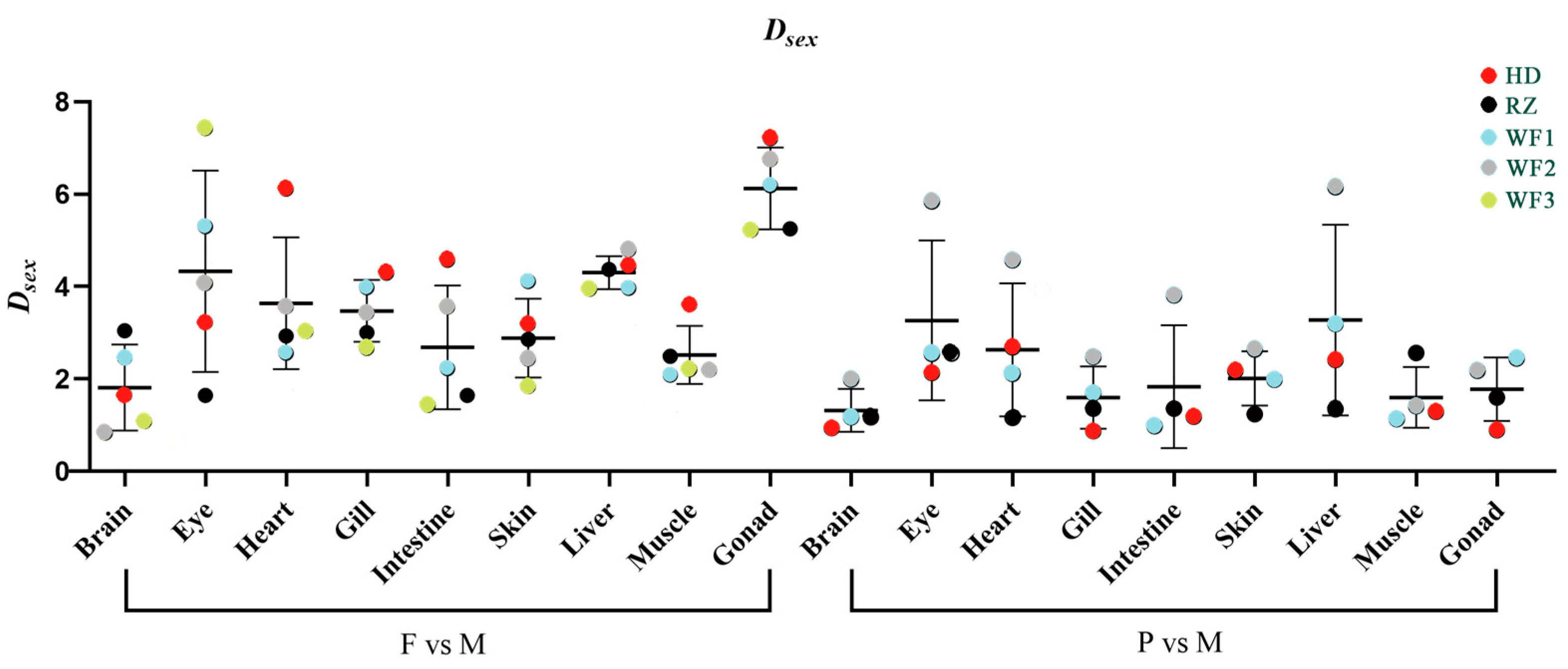
The detailed fatty acid compositions of brain, eye, heart, gill, intestine, skin, liver, muscle and gonad are presented in Supplementary Tables S2–S10, respectively. For the comparison between females and males, gonad had the highest average Dsex value, followed by liver and eye, whereas the brain had the lowest average Dsex value (Figure 3). The average Dsex values between pseudomales and males were generally lower than those between females and males. For the comparison between pseudomales and males, liver and eye had the highest average Dsex values, and the brain also had the lowest value.
3.3. SIMPER Analysis
The TOP 5 differentially abundant fatty acids between females and males (DFAsex) were analyzed by SIMPER (Table 3). Of all the TOP 5 DFAsex, docosahexaenoic acid (DHA, 22:6n-3) emerged most frequently (34/225), followed by 18:0 (33/225), 16:1n-7 (32/225), 18:1n-9 (20/225), and 18:2n-6 (19/225) (Table 3). Even only the 1st DFAsex was considered, DHA emerged most frequently (11/45), followed by 18:0 (9/45), 16:1n-7 (7/45), and 18:1n-9 (6/45) (Table 3). However, the 1st DFAsex, was largely different among different fish batches and different tissues of a certain fish batch. For the HD fish, the 1st DFAsex was DHA (brain, eye, heart, and intestine) and 16:1n-7 (gill, skin, liver, muscle, and gonad). Male fish had higher DHA content and females had higher 16:1n-7 content. Other batches have largely different 1st DFAsex in different tissues. In general, female fish had higher contents of mono-unsaturated fatty acid (MUFA) such as 16:1n-7 and 18:1n-9, and saturated fatty acid (SFA) such as 18:0 and 16:0, especially in gonads.
The TOP 5 DFAsex between pseudomale and male is presented in Table 4. Of all the TOP 5 DFAsex, 18:0 emerged most frequently (28/180), followed by DHA (24/180), 16:1n-7 (23/180), 18:1n-9 (19/180), and 18:2n-6 (18/180) (Table 4). If only the 1st DFAsex was considered, DHA emerged most frequently (10/36), followed by 16:1n-7 (5/36), 18:0 (5/36), and 18:2n-6 (4/36) (Table 4). Similarly, compared to pseudomales, males tended to have higher DHA content, but lower contents of MUFA and SFA, in particular in the gonads. The sex difference in fatty acid between pseudomales and males was batch- and tissue-specific too (Supplementary Tables S1–S10). The Dsex between female (F) and male (M) used the mean of all the five groups, whereas the Dsex between pseudomale (P) and M used the mean of all groups except WF3.
4. Discussion
Chinese tongue sole have typical sexual dimorphism. The females are much larger in size than males. In this study, the weight and length of females were higher than those of pseudomales and males, indicating that at this development stage (around 150 g), significant differences in growth rate among females, pseudomales and males had emerged. This was in line with our knowledge that female Chinese tongue sole grow at a higher rate than pseudomales and males [15]. The lack of sex difference in fish body weight of the WF3 group may be due to the unbalanced fish number of females and males (too few males collected), which may enlarge the occasionality. In addition, in most fish groups, females had lower gonadal lipid content than both pseudomales and males. This result was unexpected. Usually, ovaries have higher lipid contents than testes. The present result could be due to the different degrees of gonadal development in the three sexes. Females generally reach gonadal maturity at age 2–3, whereas males reach sexual maturity little earlier than females, with the gonads maturing after only one year [21]. The rapid deposition of lipid in the gonads of pseudomales and males at this stage helps to provide energy for gonad development, sperm formation and maintenance of sperm viability [22,23].
The Distance Coefficient (Dsex) between sexes can reflect the level of disparity in fatty acids between sexes. It was easy to understand the present result which showed that the gonads had higher Dsex than other tissues. This was consistent with our previous findings in other fish species [12]. Gonads are the main sites where the energy for gametogenesis is prepared. The different fatty acid composition between ovaries and testes indicates that different fatty acids are required during gonadal development and gametogenesis between ovaries and testes [24,25]. The analysis of the fatty acid dissimilarity between ovaries and testes of this five fish batches clearly showed that more 18:1n-9 and 16:1n-7 were deposited in ovaries but more DHA was deposited in testes. Both 18:1n-9 and 16:1n-7 are preferential substrates of β-oxidation for energy purpose, which was consistent to the energy storage requirement of ovaries [26,27]. It has been demonstrated that DHA plays important roles in the maintenance of sperm viability [28,29].
The liver and eye had moderate Dsex levels and other tissues have much lower Dsex levels. The liver is a center of fatty acid metabolism. During fish reproduction, fatty acids from the liver can be selectively transferred to the gonads to support the energy storage in the gonads [5,30,31]. The Dsex value in the eye had a large variation among different fish batches. The relatively high average Dsex value in the eye may be associated with the fact that fish eyes have high levels of DHA, which was a primary differing fatty acid between sexes. In human studies, it has also been shown that DHA is an important fatty acid for maintaining visual function and is enriched in the cell membranes of the outer segments of the optic rod cells [32,33,34]. Other tissues had relatively lower Dsex values, indicating that they are less involved in the reproduction-related fatty acid transportation across tissues.
In general, the Dsex values between pseudomales and males were much lower compared to those between females and males. This indicates that the fatty acid profile is much more similar between pseudomales and males than between females and males. Although pseudomales contain the same sex chromosome ZW as females, the gonads are physiologically developed as testes and produce sperm in the same way as males. This further indicates that the phenotypic sex more substantially determined the fatty acid composition than the genetic sex. The fatty acids deposition and mobilization may be similar between pseudomales and males at the stages near testicular maturity.
The fatty acid dissimilarity analysis by SIMPER showed that DHA, 16:1n-7, 18:0 and 18:1n-9 are the main differentially abundant fatty acids between sexes. This was similar to what observed in our previous studies [35]. As mentioned above, testes may require more DHA than ovaries at this stage. Previous studies on Chinese tongue sole [5], European sea bass (Dicentrarchus labrax) [36], and European eel (Anguilla anguilla) [37] have shown that DHA is an important regulator of testes maturation and sex steroid hormones synthesis. However, the specific mechanisms involved in the regulation of androgen synthesis and testes development by DHA remain unclear and need to be elucidated by future studies. Different from our previous studies, which showed that arachidonic acid (ARA, 20:4n-6) was also largely different in concentration between female and male tongue sole [5], this study did not reveal a similar result. The discrepancy could be mainly related to the gonadal development stage. The roles of ARA in fish reproduction and many other physiological processes have been reported to be stage-specific [6]. In contrast, the sex difference in eicosapentaenoic acid (EPA, 20:5n-3) was low, indicating that EPA might not be very important to fish reproduction [5].
Also as mentioned above, 16:1n-7 and 18:1n-9 are main differentially abundant fatty acids between sexes too. They were more abundant in ovaries than in testes. In general, these two fatty acids play major roles in energy supply. Although the sex difference in 16:1n-7 and 18:1n-9 were not consistent across fish batches and tissues, their transportation across tissues could be active during gonadal development. Currently, 16:1n-7 has not been well-studied in aquatic animals. This fatty acid has been reported to be closely associated with insulin sensitivity and regulation of triglyceride and glucose metabolism in white adipose tissue [38,39,40]. Besides the energy roles, 18:1n-9 also has regulatory effects on cholesterol and insulin and can reduce the production of steroid hormones [41,42]. The present study showed that the testes contain lower levels of 16:1n-7 and 18:1n-9 than the ovaries, which was easy to understand from the energy storage aspect.
Palmitic acid (16:0) is also an important substrate for energy supply. However, for several tissues, such as testis of HD and WF3 fish, intestine and muscle of RZ fish, as well as heart, gill and liver of WF2 fish, male fish had higher 16:0 levels than females. At this stage, male fish may have different preferential fatty acids for energy supply. The higher 16:0 content in the testes than in the ovaries was consistent with our previous studies [12]. It was speculated that 16:0 not only has an energetic role for the testis, but may also has an unknown physiological function in reproductive processes of male fish. Further studies are needed to explain the current findings. Stearic acid (18:0) was another important differentially abundant fatty acid between sexes. However, whether males or females had a higher 18:0 content was fish batch- and tissue-specific. Specific to gonads, in general testes had higher 18:0 levels than ovaries. Similar results have also been observed in our previous studies [12] as well as studies on Chinese sturgeon (Acipenser sinensis) [43] and sweet smelt (Plecoglossus altivelis) [44] by other researchers.
Considering that the differences between the fatty acids of pseudomale and male fish were not so significant, these difference were not to be discussed in detail. Nevertheless, it was worth mentioning that males still had higher DHA contents than pseudomales, similar to the comparison between males and females, indicating the gradual change of fatty acid composition during sex reversal. Also, it was obvious and important that the phenotypic sex more substantially determined the fatty acid composition than the genetic sex.
5. Conclusions
In conclusion, at the age of 11 months, Chinese tongue sole, which have typical sexual dimorphism and reversal, have had a large sex difference in tissue fatty acid compositions. The highest sex difference in fatty acid was observed in gonads, followed by liver and eye. The fatty acids that differed between sexes were mainly DHA, 16:1n-7, 18:1n-9, and 18:0. The testes tended to have higher DHA levels but lower levels of 16:1n-7 and 18:1n-9 than ovaries. The differences between the fatty acids of pseudomale and male fish were not significant, indicating a more fatty acid-determining role of phenotypic sex than genetic sex, but an obvious difference in DHA can still be observed between pseudomales and males. Future studies are needed to investigate the sex difference in fish fatty acids at different gonadal development stages.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fishes8080421/s1, Table S1: Physical indices and total lipid content of liver, muscle and gonad in five batches of Chinese tongue sole; Table S2: The brain fatty acid composition of experimental fish; Table S3: The eye fatty acid composition of experimental fish; Table S4: The heart fatty acid composition of experimental fish; Table S5: The gill fatty acid composition of experimental fish; Table S6: The intestine fatty acid composition of experimental fish; Table S7: The skin fatty acid composition of experimental fish; Table S8: The liver fatty acid composition of experimental fish; Table S9: The muscle fatty acid composition of experimental fish; Table S10: The gonad fatty acid composition of experimental fish.
Author Contributions
Conceptualization, J.L. and H.X.; methodology, F.Z.; software, Y.W.; validation, Q.M.; formal analysis, J.L. and F.Z.; investigation, J.L.; resources, M.L.; data curation, Y.W.; writing—original draft preparation, J.L.; writing—review and editing, H.X.; visualization, J.L.; supervision, H.X.; project administration, M.L. and H.X.; funding acquisition, M.L. and H.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Foundation of Shandong Province Outstanding Youth Foundation (grant number ZR2021YQ24), Central Public-Interest Scientific Institution Basal Research Fund, CAFS (2020TD48), and China Agriculture Research System (grant number CARS-47).
Institutional Review Board Statement
The animal-study protocol was approved by the Animal Care and Use Committee of Yellow Sea Fisheries Research Institute (protocol code ACUC202302122514; date of approval, 12 February 2023).
Data Availability Statement
The data that support the findings of this study are included in the text and in the tables. Raw data for all figures are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Xishuai Cui for his help in fish collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martins, D.A.; Engrola, S.; Morais, S.; Bandarra, N.; Coutinho, J.; Yúfera, M.; Conceição, L.E.C. Cortisol response to air exposure in Solea senegalensis post-larvae is affected by dietary arachidonic acid-to-eicosapentaenoic acid ratio. Fish Physiol. Biochem. 2011, 37, 733–743. [Google Scholar] [CrossRef]
- Martins, D.A.; Rocha, F.; Castanheira, F.; Mendes, A.; Pousão-Ferreira, P.; Bandarra, N.; Coutinho, J.; Morais, S.; Yúfera, M.; Conceição, L.E.C.; et al. Effects of dietary arachidonic acid on cortisol production and gene expression in stress response in Senegalese sole (Solea senegalensis) post-larvae. Fish Physiol. Biochem. 2013, 39, 1223–1238. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Almatar, S.M.; James, C.M. Effects of varying dietary docosahexaenoic acid levels on growth, proximate composition and tissue fatty acid profile of juvenile silver pomfrets, Pampus argenteus (Euphrasen, 1788). Aquac. Res. 2012, 43, 1599–1610. [Google Scholar] [CrossRef]
- Cardona, E.; Segret, E.; Cachelou, Y. Effect of micro-algae Schizochytrium sp. supplementation in plant diet on reproduction of female rainbow trout (Oncorhynchus mykiss): Maternal programming impact of progeny. J. Anim. Sci. Biotechnol. 2022, 13, 33. [Google Scholar] [CrossRef]
- Xu, H.; Cao, L.; Zhang, Y.; Johnson, R.B.; Wei, Y.; Zheng, K.; Liang, M. Dietary arachidonic acid differentially regulates the gonadal steroidogenesis in the marine teleost, tongue sole (Cynoglossus semilaevis), depending on fish gender and maturation stage. Aquaculture 2017, 468, 378–385. [Google Scholar] [CrossRef]
- Xu, H.; Meng, X.; Wei, Y.; Ma, Q.; Liang, M.; Turchini, G.M. Arachidonic acid matters. Rev. Aquac. 2022, 14, 1912–1944. [Google Scholar] [CrossRef]
- Manor, M.L.; Weber, G.M.; Cleveland, B.M.; Kenney, P.B. Effects of feeding level and sexual maturation on fatty acid composition of energy stores in diploid and triploid rainbow trout (Oncorhynchus mykiss). Aquaculture 2014, 418, 17–25. [Google Scholar] [CrossRef]
- Ding, L.; Liu, Y.; Kang, M.; Wei, X.; Geng, C.; Liu, W.; Han, L.; Yuan, F.; Wang, P.; Wang, B.; et al. UPLC-QTOF/MS metabolomics and biochemical assays reveal changes in hepatic nutrition and energy metabolism during sexual maturation in female rainbow trout (Oncorhynchus mykiss). Biology 2022, 11, 1679. [Google Scholar] [CrossRef]
- Bhat, R.A.; Saini, S.; Saoca, C.; Maricchiolo, G.; Fazio, F. Analysis of fatty acids and sex steroid hormones in rainbow trout testes (Oncorhynchus mykiss) during the reproductive process. Aquac. Res. 2022, 53, 4426–4436. [Google Scholar] [CrossRef]
- Luzia, L.A.; Sampaio, G.R.; Castellucci, C.M.N.; Torres, E.A.F.S. The influence of season on the lipid profiles of five commercially important species of Brazilian fish. Food Chem. 2003, 83, 93–97. [Google Scholar] [CrossRef]
- Nogueira, N.; Fernandes, I.; Fernandes, T.; Cordeiro, N. A comparative analysis of lipid content and fatty acid composition in muscle, liver and gonads of Seriola fasciata Bloch 1793 based on gender and maturation stage. J. Food Compost. Anal. 2017, 59, 68–73. [Google Scholar] [CrossRef]
- Zhang, F.; Liu, J.; Bi, Q.; Ma, Q.; Wei, Y.; Liang, M.; Xu, H. Sex difference in fatty acid composition of six marine teleosts. J. Fish Biol. 2022, 101, 1606–1610. [Google Scholar] [CrossRef] [PubMed]
- Luczynska, J.; Tonska, E.; Krejszeff, S.; Zarski, D. Comparison of Fatty Acids in the Muscles and Liver of Pond-Cultured and Wild Perch, Perca fluviatilis (L.), in Poland. Turk. J. Fish Aquat. Sci. 2016, 16, 19–27. [Google Scholar] [CrossRef]
- Zarski, D.; Palinska-Zarska, K.; Luczynska, J.; Krejszeff, S. The type of spawning agent affects the egg composition during out-of season spawning but not during in-season spawning in Eurasian perch, Perca fluviatilis. Gen. Comp. Endocrinol. 2017, 245, 19–29. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Q.; Hu, Y.; Xu, W.; Yang, Y.; Chen, S.; Wang, N. Identification of lncRNA-miRNA-mRNA network involved in sexual size dimorphism of Chinese Tongue Sole (Cynoglossus semilaevis). Front. Mar. Sci. 2022, 9, 795525. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, M.; Liu, J.; Liu, Y.; Lu, J.; Sun, X. Sexually dimorphic gene expression associated with growth and reproduction of tongue sole (Cynoglossus semilaevis) revealed by brain transcriptome analysis. Int. J. Mol. Sci. 2016, 17, 1402. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Wang, J.; Yang, Y.; Chen, Z.; Wang, Q.; Wang, J.; Zhang, T.; Xu, W.; Chen, S. Identification and expression pattern of cyp26b1 gene in gonad of the Chinese tongue sole (Cynoglossus semilaevis). Animals 2022, 12, 2652. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Turchini, G.M.; Francis, D.S.; De Silva, S.S. Finishing diets stimulate compensatory growth: Results of a study on Murray cod, Maccullochella peelii peelii. Aquacult. Nutr. 2007, 13, 351–360. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C. Substitution of fish oil with camelina oil and inclusion of camelina meal in diets fed to Atlantic cod (Gadus morhua) and their effects on growth, tissue lipid classes, and fatty acids. J. Anim. Sci. 2014, 92, 1055–1067. [Google Scholar] [CrossRef]
- Ji, X.; Liu, H.; Chen, S.; Jiang, Y.; Tian, Y. Growth differences and dimorphic expression of growth hormone (GH) in female and male Cynoglossus semilaevis after male sexual maturation. Mar. Genom. 2011, 4, 9–16. [Google Scholar] [CrossRef]
- Martín, M.V.; Rodríguez, C.; Cejas, J.R.; Pérez, M.J.; Jerez, S.; Lorenzo, A. Body lipid and fatty acid composition in male gilthead seabream broodstock at different stages of the reproductive cycle: Effects of a diet lacking n-3 and n-6 HUFA. Aquacult. Nutr. 2009, 15, 60–72. [Google Scholar] [CrossRef]
- Bogevik, A.S.; Hayman, E.S.; Bjerke, M.T.; Dessen, J.E.; Rørvik, K.A.; Luckenbach, J.A. Phospholipid and LC-PUFA metabolism in Atlantic salmon (Salmo salar) testes during sexual maturation. PLoS ONE 2020, 15, e0233322. [Google Scholar] [CrossRef]
- Esmaeili, V.; Shahverdi, A.H.; Moghadasian, M.H.; Alizadeh, A.R. Dietary fatty acids affect semen quality: A review. Andrology 2015, 3, 450–461. [Google Scholar] [CrossRef]
- Hauville, M.R.; Rhody, N.R.; Resley, M.J.; Bell, J.G.; Main, K.L.; Migaud, H. Comparative study of lipids and fatty acids in the liver, muscle, and eggs of wild and captive common snook broodstock. Aquaculture 2015, 446, 227–235. [Google Scholar] [CrossRef]
- Nie, L.; Ren, Y.; Schulz, H. Identification and characterization of escherichia coli thioesterase III that functions in fatty acid beta-oxidation. Biochemistry 2008, 47, 7744–7751. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, R.; Feng, C.; Liu, R.; Zheng, Y.; Hoque, S.A.M.; Wu, D.; Lu, H.; Zhang, T.; Zeng, W. Exogenous oleic acid and palmitic acid improve boar sperm motility via enhancing mitochondrial β-oxidation for ATP generation. Animals 2020, 10, 591. [Google Scholar] [CrossRef]
- Gholami, H.; Chamani, M.; Towhidi, A.; Fazeli, M.H. Effect of feeding a docosahexaenoic acid-enriched nutriceutical on the quality of fresh and frozen-thawed semen in Holstein bulls. Theriogenology 2010, 74, 1548–1558. [Google Scholar] [CrossRef]
- Kaeoket, K.; Sang-Urai, P.; Thamniyom, A.; Chanapiwat, P.; Techakumphu, M. Effect of docosahexaenoic acid on quality of cryopreserved boar semen in different breeds. Reprod. Domest. Anim. 2010, 45, 458–463. [Google Scholar] [CrossRef]
- Wassef, E.A.; Wahbi, O.M.; Shalaby, S.H. Effects of dietary vegetable oils on liver and gonad fatty acid metabolism and gonad maturation in gilthead seabream (Sparus aurata) males and females. Aquacult. Int. 2012, 20, 255–281. [Google Scholar] [CrossRef]
- Rudchenko, A.E.; Yablokov, N.O. Composition and content of fatty acids in the tissues of males and females of Eurasian perch Perca fluviatilis at the late stages of reproductive cycle. Contemp. Probl. Ecol. 2018, 11, 309–319. [Google Scholar] [CrossRef]
- Anderson, R.E. Lipids of ocular tissues. IV: A comparison of the phospholipids from the retina of six mammalian species. Exp. Eye Res. 1970, 10, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Docosahexaenoic Acid. Ann. Nutr. Metab. 2016, 69, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA effects in brain development and function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef]
- Zhang, F.; Li, L.; Bi, Q.; Wei, Y.; Liang, M.; Xu, H. Lipid distribution and fatty acid profile of five benthic marine fish species in the yellow sea. Prog. Fish. Sci. 2023, 44, 97–110, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Asturiano, J.F.; Sorbera, L.A.; Zanuy, S.; Carrillo, M. Effects of polyunsaturated fatty acids and gonadotropin on prostaglandin series E production in a primary testis cell culture system for the European sea bass. J. Fish Biol. 2000, 57, 1563–1574. [Google Scholar] [CrossRef]
- Baeza, R.; Peñaranda, D.S.; Vílchez, M.C.; Tveiten, H.; Perez, L.; Asturiano, J.F. Exploring correlations between sex steroids and fatty acids and their potential roles in the induced maturation of the male European eel. Aquaculture 2015, 435, 328–335. [Google Scholar] [CrossRef]
- Burhans, M.S.; Flowers, M.T.; Harrington, K.R.; Bond, L.M.; Guo, C.A.; Anderson, R.M.; Ntambi, J.M. Hepatic oleate regulates adipose tissue lipogenesis and fatty acid oxidation. J. Lipid Res. 2015, 56, 304–318. [Google Scholar] [CrossRef]
- Burns, T.A.; Kadegowda, A.K.G.; Duckett, S.K.; Pratt, S.L.; Jenkins, T.C. Palmitoleic (16:1 cis-9) and cis-vaccenic (18:1 cis-11) acid alter lipogenesis in bovine adipocyte cultures. Lipids 2012, 47, 1143–1153. [Google Scholar] [CrossRef]
- Bu, S.Y.; Mashek, D.G. Hepatic long-chain acyl-CoA synthetase 5 mediates fatty acid channeling between anabolic and catabolic pathways. J. Lipid Res. 2010, 51, 3270–3280. [Google Scholar] [CrossRef]
- Ducheix, S.; Montagner, A.; Polizzi, A.; Lasserre, F.; Regnier, M.; Marmugi, A.; Benhamed, F.; Bertrand-Michel, J.; Mselli-Lakhal, L.; Postic, C.; et al. Dietary oleic acid regulates hepatic lipogenesis through a liver X receptor-dependent signaling. PLoS ONE 2017, 12, e0181393. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Mo, Z.; Li, Y.; Huang, L.; Yu, S.; Ge, L.; Hu, Y.; Shi, S.; Zhang, L.; Wang, L.; et al. Oleic acid reduces steroidogenesis by changing the lipid type stored in lipid droplets of ovarian granulosa cells. J. Anim. Sci. Biotechnol. 2022, 13, 27. [Google Scholar] [CrossRef]
- Zhou, H.; Leng, X.; Tan, Q.; Du, H.; Wu, J.; Liang, X.; Wei, Q. Identification of key nutrients for gonadal development by comparative analysis of proximate composition and fatty/amino acid profile in tissues and eggs of Chinese sturgeon (Acipenser sinensis Gray, 1835). J. Appl. Ichthyol. 2017, 33, 885–891. [Google Scholar] [CrossRef]
- Jeong, B.Y.; Jeong, W.G.; Moon, S.K.; Ohshima, T. Preferential accumulation of fatty acids in the testis and ovary of cultured and wild sweet smelt Plecoglossus altivelis. Comp. Biochem. Phys. B 2002, 131, 251–259. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Examples of genetic sex identification by PCR study. Observation of two bands represents the genetic females Chinese tongue sole Cynoglossus semilaevis and a single band represents the genetic males Chinese tongue sole Cynoglossus semilaevis.
Figure 1.
Examples of genetic sex identification by PCR study. Observation of two bands represents the genetic females Chinese tongue sole Cynoglossus semilaevis and a single band represents the genetic males Chinese tongue sole Cynoglossus semilaevis.

Figure 2.
Body weight (A), body length (B), hepatosomatic index (C), condition factor (D), and total lipid content (E) for five batches of Chinese tongue sole Cynoglossus semilaevis. For each group, the data from all replicate fish were used to create this graph. The small number of pseudomale fish (only one fish) in batch WF3 failed to meet the requirement of statistical analysis, resulting in a data gap in this group. In addition, the gonad mass of the HD group was small and the intra-group error was too large, making the statistics less reliable. Values are means with their standard errors represented by vertical bars. a–c Mean values within a fish batch not sharing a same letter are significantly different (p < 0.05), and “*” means significantly different values tested by t-test. F: female; P: pseudomale; M: male.
Figure 2.
Body weight (A), body length (B), hepatosomatic index (C), condition factor (D), and total lipid content (E) for five batches of Chinese tongue sole Cynoglossus semilaevis. For each group, the data from all replicate fish were used to create this graph. The small number of pseudomale fish (only one fish) in batch WF3 failed to meet the requirement of statistical analysis, resulting in a data gap in this group. In addition, the gonad mass of the HD group was small and the intra-group error was too large, making the statistics less reliable. Values are means with their standard errors represented by vertical bars. a–c Mean values within a fish batch not sharing a same letter are significantly different (p < 0.05), and “*” means significantly different values tested by t-test. F: female; P: pseudomale; M: male.
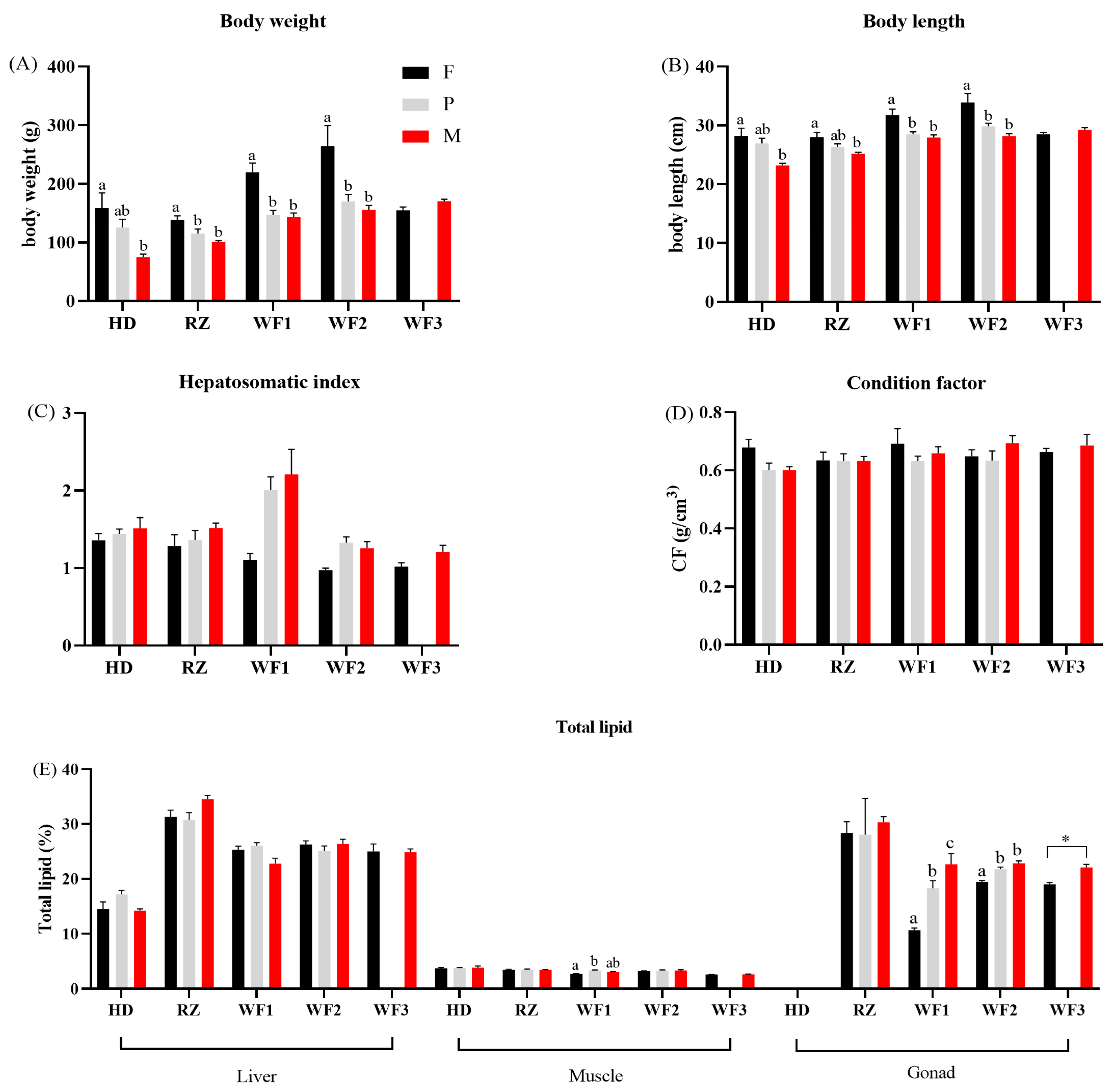
Figure 3.
Average Dsex values of different tissues across groups of Chinese tongue sole Cynoglossus semilaevis. The Dsex between females (F) and males (M) used the mean of all the five groups, whereas the Dsex between pseudomales (P) and M used the mean of all groups except WF3. The small number of pseudomale fish (only one fish) in batch WF3 failed to meet the requirement of statistical analysis, resulting in a data gap in this group.
Figure 3.
Average Dsex values of different tissues across groups of Chinese tongue sole Cynoglossus semilaevis. The Dsex between females (F) and males (M) used the mean of all the five groups, whereas the Dsex between pseudomales (P) and M used the mean of all groups except WF3. The small number of pseudomale fish (only one fish) in batch WF3 failed to meet the requirement of statistical analysis, resulting in a data gap in this group.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
PCR reaction system used for sex identification.
| Reagent | Dosage | Temperature | Time | |
|---|---|---|---|---|
| 2 × A8 PCR MasterMix 1 | 12.5 μL | 95 °C | 3 min (pre-degeneration) | |
| Forward primer | 1 μL | 95 °C | 10 s | 35 cycles |
| Reverse primer | 1 μL | 55 °C | 15 s | |
| DNA | 2.5 μL | 72 °C | 1 min | |
| ddH2O | 8 μL | 72 °C | 5 min (Radical extension) |
1 2 × A8 PCR MasterMix from Aidlab Biotechnologies Co., Ltd. (Beijing, China).
Table 2.
Sex distribution (number) of the Chinese tongue sole Cynoglossus semilaevis collected from different farms.
Table 2.
Sex distribution (number) of the Chinese tongue sole Cynoglossus semilaevis collected from different farms.
| Sex | HD | RZ | WF1 | WF2 | WF3 |
|---|---|---|---|---|---|
| Female | 5 | 7 | 8 | 10 | 25 |
| Male | 7 | 17 | 9 | 11 | 3 |
| Pseudomale | 18 | 6 | 13 | 9 | 1 1 |
1 The small number of pseudomale fish (only one fish) in batch WF3 failed to meet the requirements of statistical analysis. Therefore, this fish was discarded in following analysis.
Table 3.
TOP 5 differentially abundant FA between female and male Chinese tongue sole Cynoglossus semilaevis (DFAsex) analyzed with SIMPER.
Table 3.
TOP 5 differentially abundant FA between female and male Chinese tongue sole Cynoglossus semilaevis (DFAsex) analyzed with SIMPER.
| Group | Tissue | 1st DFAsex 1 | 2nd DFAsex | 3rd DFAsex | 4th DFAsex | 5th DFAsex | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FA | Contrib% 2 | FA | Contrib% | FA | Contrib% | FA | Contrib% | FA | Contrib% | ||
| HD | brain | 22:6n-3 3 | 16.7 | 16:0 | 10.8 | 18:1n-9 | 10.6 | 16:1n-7 | 9.7 | 18:0 | 9.6 |
| eye | 22:6n-3 | 22.2 | 16:1n-7 | 16.5 | 20:5n-3 | 14.3 | 14:0 | 13.9 | 18:0 | 8.0 | |
| heart | 22:6n-3 | 21.7 | 20:5n-3 | 17.0 | 16:1n-7 | 16.8 | 18:0 | 10.3 | 14:0 | 8.4 | |
| gill | 16:1n-7 | 16.6 | 22:6n-3 | 13.9 | 20:5n-3 | 13.1 | 14:0 | 10.5 | 18:0 | 9.0 | |
| intestine | 22:6n-3 | 12.6 | 18:0 | 9.4 | 20:5n-3 | 9.2 | 16:1n-7 | 8.4 | 22:5n-3 | 8.1 | |
| skin | 16:1n-7 | 13.8 | 20:5n-3 | 11.7 | 18:2n-6 | 10.1 | 16:0 | 8.5 | 14:0 | 8.0 | |
| liver | 16:1n-7 | 23.4 | 18:0 | 14.5 | 22:6n-3 | 12.9 | 18:1n-9 | 11.0 | 22:5n-3 | 7.3 | |
| muscle | 16:1n-7 | 20.4 | 18:0 | 18.9 | 22:6n-3 | 10.1 | 14:0 | 10.0 | 16:0 | 8.1 | |
| gonad | 16:1n-7 | 16.2 | 20:5n-3 | 15.7 | 16:0 | 15.4 | 18:0 | 11.8 | 18:2n-6 | 10.3 | |
| RZ | brain | 18:2n-6 | 20.2 | 22:6n-3 | 13.0 | 20:5n-3 | 12.7 | 16:1n-7 | 9.9 | 18:1n-9 | 9.5 |
| eye | 18:2n-6 | 18.4 | 22:5n-3 | 14.2 | 14:0 | 11.2 | 22:6n-3 | 11.0 | 16:1n-7 | 11.0 | |
| heart | 18:0 | 14.1 | 16:1n-7 | 13.5 | 22:6n-3 | 12.5 | 20:5n-3 | 11.5 | 18:1n-9 | 10.0 | |
| gill | 20:3n-3 | 12.9 | 16:1n-7 | 11.8 | 18:0 | 10.9 | 16:0 | 10.9 | 20:5n-3 | 9.0 | |
| intestine | 16:0 | 10.4 | 20:5n-3 | 10.3 | 18:0 | 10.0 | 20:2n-6 | 8.4 | 20:3n-6 | 8.1 | |
| skin | 18:0 | 13.5 | 14:0 | 13.5 | 16:0 | 11.5 | 20:2n-6 | 11.1 | 16:1n-7 | 8.8 | |
| liver | 16:1n-7 | 16.5 | 22:6n-3 | 15.2 | 18:0 | 12.5 | 20:5n-3 | 9.6 | 16:0 | 9.3 | |
| muscle | 16:1n-7 | 15.1 | 16:0 | 14.3 | 14:0 | 13.1 | 18:0 | 11.2 | 20:5n-3 | 8.6 | |
| gonad | 18:1n-9 | 16.9 | 16:1n-7 | 16.4 | 18:0 | 13.0 | 22:6n-3 | 10.4 | 14:0 | 8.3 | |
| WF1 | brain | 18:1n-9 | 18.1 | 18:0 | 11.8 | 22:6n-3 | 10.9 | 18:2n-6 | 9.4 | 16:1n-7 | 9.2 |
| eye | 22:6n-3 | 17.1 | 18:1n-9 | 15.6 | 18:0 | 13.3 | 16:1n-7 | 9.0 | 20:3n-3 | 8.9 | |
| heart | 18:1n-9 | 18.4 | 16:1n-7 | 15.5 | 18:0 | 10.9 | 20:5n-3 | 9.9 | 20:3n-3 | 8.6 | |
| gill | 18:0 | 15.8 | 14:0 | 12.5 | 16:1n-7 | 11.8 | 17:1n-7 | 9.5 | 20:3n-3 | 9.2 | |
| intestine | 18:0 | 13.9 | 16:1n-7 | 10.8 | 18:2n-6 | 9.8 | 18:1n-9 | 8.2 | 22:5n-3 | 8.1 | |
| skin | 18:0 | 20.7 | 16:1n-7 | 12.1 | 22:6n-3 | 9.4 | 18:3n-3 | 9.2 | 20:3n-3 | 9.1 | |
| liver | 18:0 | 21.2 | 22:6n-3 | 10.6 | 18:1n-9 | 9.9 | 16:1n-7 | 8.8 | 18:2n-6 | 7.4 | |
| muscle | 18:1n-9 | 16.9 | 14:0 | 15.6 | 22:5n-3 | 10.5 | 16:0 | 9.2 | 22:6n-3 | 8.6 | |
| gonad | 22:6n-3 | 22.3 | 22:5n-3 | 15.0 | 16:1n-7 | 9.8 | 18:1n-9 | 7.9 | 18:3n-3 | 7.2 | |
| WF2 | brain | 22:6n-3 | 15.8 | 18:1n-9 | 13.6 | 16:1n-7 | 12.9 | 22:5n-3 | 12.2 | 16:0 | 8.9 |
| eye | 22:6n-3 | 17.3 | 18:1n-9 | 14.5 | 20:5n-3 | 13.6 | 16:0 | 13.3 | 18:2n-6 | 12.7 | |
| heart | 16:0 | 13.1 | 18:2n-6 | 12.7 | 16:1n-7 | 11.1 | 18:0 | 10.8 | 22:6n-3 | 10.0 | |
| gill | 16:0 | 17.0 | 20:5n-3 | 12.2 | 22:6n-3 | 9.4 | 20:3n-3 | 8.5 | 18:0 | 8.4 | |
| intestine | 20:2n-6 | 15.4 | 18:2n-6 | 11.0 | 22:6n-3 | 10.9 | 22:5n-3 | 8.8 | 20:5n-3 | 8.0 | |
| skin | 18:0 | 11.0 | 18:2n-6 | 9.8 | 20:3n-3 | 8.6 | 16:0 | 8.3 | 14:0 | 7.5 | |
| liver | 16:0 | 17.9 | 18:0 | 15.6 | 18:2n-6 | 11.7 | 22:6n-3 | 9.3 | 18:1n-9 | 8.0 | |
| muscle | 18:2n-6 | 22.3 | 18:3n-3 | 13.0 | 22:5n-3 | 12.2 | 22:6n-3 | 11.6 | 18:0 | 8.0 | |
| gonad | 18:1n-9 | 19.4 | 16:1n-7 | 17.0 | 22:6n-3 | 16.8 | 14:0 | 7.9 | 20:5n-3 | 5.9 | |
| WF3 | brain | 22:6n-3 | 16.6 | 18:1n-9 | 14.3 | 16:1n-7 | 11.2 | 16:0 | 9.6 | 18:0 | 8.4 |
| eye | 22:6n-3 | 25.9 | 18:1n-9 | 16.9 | 16:0 | 12.1 | 18:2n-6 | 11.4 | 14:0 | 8.5 | |
| heart | 18:0 | 23.2 | 20:3n-3 | 12.3 | 18:2n-6 | 10.7 | 22:6n-3 | 10.2 | 18:1n-9 | 8.2 | |
| gill | 18:2n-6 | 12.9 | 18:0 | 10.4 | 22:6n-3 | 10.1 | 20:3n-3 | 9.5 | 16:0 | 8.5 | |
| intestine | 22:5n-3 | 18.0 | 20:5n-3 | 13.4 | 16:0 | 10.1 | 18:0 | 8.2 | 22:6n-3 | 5.8 | |
| skin | 18:0 | 17.3 | 18:2n-6 | 9.3 | 22:5n-3 | 8.8 | 16:1n-7 | 7.6 | 17:1n-7 | 7.1 | |
| liver | 22:6n-3 | 15.6 | 18:2n-6 | 13.9 | 16:0 | 11.0 | 18:0 | 10.7 | 20:3n-3 | 7.8 | |
| muscle | 18:2n-6 | 16.4 | 22:5n-3 | 10.8 | 16:1n-7 | 10.0 | 22:6n-3 | 9.1 | 17:1n-7 | 9.1 | |
| gonad | 18:1n-9 | 18.1 | 16:1n-7 | 12.4 | 22:6n-3 | 11.0 | 16:0 | 10.2 | 20:4n-6 | 8.7 | |
1 1st, 2nd, 3rd, 4th, and 5th DFAsex is the fatty acid contributing the most, the second, the third, the fourth, and the fifth largely to the total fatty acid difference between sexes, respectively. 2 Contrib%—contribution % of a certain fatty acid difference to the total fatty acid difference between sexes. 3 Underlined fatty acids had higher content in males compared to females.
Table 4.
TOP 5 differentially abundant fatty acid between pseudomale and male Chinese tongue sole Cynoglossus semilaevis (DFAsex) analyzed with SIMPER.
Table 4.
TOP 5 differentially abundant fatty acid between pseudomale and male Chinese tongue sole Cynoglossus semilaevis (DFAsex) analyzed with SIMPER.
| Group | Tissue | 1st DFAsex 1 | 2nd DFAsex | 3rd DFAsex | 4th DFAsex | 5th DFAsex | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FA | Contrib% 2 | FA | Contrib% | FA | Contrib% | FA | Contrib% | FA | Contrib% | ||
| HD | brain | 22:6n-3 3 | 16.8 | 16:1n-7 | 11.7 | 18:0 | 10.9 | 20:5n-3 | 9.5 | 22:5n-3 | 9.0 |
| eye | 22:6n-3 | 25.9 | 16:1n-7 | 17.3 | 14:0 | 12.0 | 18:1n-9 | 9.5 | 18:0 | 7.8 | |
| heart | 22:6n-3 | 19.5 | 16:1n-7 | 17.5 | 20:5n-3 | 11.7 | 18:0 | 10.3 | 14:0 | 9.3 | |
| gill | 22:6n-3 | 17.2 | 16:1n-7 | 15.6 | 14:0 | 9.2 | 18:0 | 8.9 | 20:5n-3 | 7.0 | |
| intestine | 16:1n-7 | 12.8 | 22:6n-3 | 12.6 | 14:0 | 10.2 | 18:0 | 8.8 | 20:5n-3 | 8.1 | |
| skin | 18:2n-6 | 10.8 | 22:6n-3 | 10.7 | 16:1n-7 | 9.9 | 20:5n-3 | 8.1 | 14:0 | 8.0 | |
| liver | 16:1n-7 | 22.1 | 22:6n-3 | 16.1 | 18:0 | 12.4 | 18:1n-9 | 12.4 | 20:3n-3 | 6.8 | |
| muscle | 18:0 | 20.9 | 16:1n-7 | 17.5 | 18:0 | 10.3 | 18:3n-3 | 7.2 | 16:0 | 7.0 | |
| gonad | 16:1n-7 | 13.5 | 18:2n-6 | 10.6 | 20:5n-3 | 10.3 | 16:0 | 9.4 | 18:0 | 9.1 | |
| RZ | brain | 18:2n-6 | 21.6 | 22:6n-3 | 15.3 | 18:1n-9 | 11.4 | 16:1n-7 | 10.7 | 18:0 | 9.6 |
| eye | 22:6n-3 | 18.4 | 18:2n-6 | 12.2 | 14:0 | 11.4 | 20:5n-3 | 11.3 | 18:0 | 10.9 | |
| heart | 22:6n-3 | 14.6 | 16:1n-7 | 14.3 | 20:5n-3 | 11.7 | 18:1n-9 | 11.4 | 18:0 | 10.7 | |
| gill | 20:5n-3 | 11.6 | 18:3n-3 | 10.6 | 18:2n-6 | 9.7 | 18:0 | 9.7 | 16:1n-7 | 8.7 | |
| intestine | 20:5n-3 | 10.4 | 18:0 | 10.0 | 16:0 | 8.6 | 20:2n-6 | 8.3 | 20:3n-6 | 7.8 | |
| skin | 18:0 | 11.5 | 18:3n-3 | 10.2 | 17:1n-7 | 9.6 | 20:2n-6 | 8.0 | 20:3n-3 | 7.4 | |
| liver | 16:1n-7 | 13.7 | 18:0 | 12.4 | 18:2n-6 | 11.0 | 16:0 | 9.5 | 18:1n-9 | 9.4 | |
| muscle | 16:1n-7 | 21.6 | 18:0 | 15.2 | 22:6n-3 | 12.6 | 18:1n-9 | 9.0 | 14:0 | 8.2 | |
| gonad | 18:0 | 13.0 | 20:5n-3 | 12.7 | 18:2n-6 | 10.9 | 18:1n-9 | 10.2 | 16:1n-7 | 10.1 | |
| WF1 | brain | 22:5n-3 | 14.3 | 18:1n-9 | 11.3 | 22:6n-3 | 10.0 | 18:0 | 9.2 | 14:0 | 8.1 |
| eye | 18:2n-6 | 14.2 | 16:0 | 12.3 | 22:6n-3 | 11.9 | 16:1n-7 | 11.0 | 18:0 | 9.9 | |
| heart | 18:1n-9 | 18.7 | 22:6n-3 | 15.6 | 20:5n-3 | 12.3 | 16:1n-7 | 8.9 | 20:3n-3 | 7.9 | |
| gill | 18:0 | 15.9 | 16:1n-7 | 12.1 | 20:3n-3 | 11.8 | 14:0 | 10.3 | 18:1n-9 | 9.1 | |
| intestine | 16:0 | 11.7 | 16:1n-7 | 10.6 | 18:1n-9 | 9.9 | 18:2n-6 | 9.7 | 18:0 | 9.6 | |
| skin | 18:0 | 16.7 | 16:1n-7 | 10.2 | 20:3n-3 | 8.6 | 20:2n-6 | 7.7 | 14:0 | 7.5 | |
| liver | 14:0 | 17.9 | 16:0 | 12.2 | 18:1n-9 | 8.5 | 22:6n-3 | 8.0 | 18:0 | 7.6 | |
| muscle | 16:0 | 12.7 | 22:5n-3 | 11.1 | 18:0 | 11.0 | 18:2n-6 | 10.0 | 17:1n-7 | 9.1 | |
| gonad | 22:5n-3 | 20.2 | 22:6n-3 | 11.6 | 18:1n-9 | 8.0 | 14:0 | 7.8 | 16:1n-7 | 7.7 | |
| WF2 4 | brain | 22:6n-3 | 19.5 | 18:1n-9 | 14.1 | 18:0 | 11.6 | 18:2n-6 | 11.4 | 16:0 | 9.0 |
| eye | 22:6n-3 | 23.2 | 20:5n-3 | 16.0 | 18:2n-6 | 15.7 | 18:1n-9 | 13.2 | 16:1n-7 | 7.8 | |
| heart | 22:6n-3 | 20.9 | 18:2n-6 | 18.0 | 18:1n-9 | 9.8 | 16:1n-7 | 9.8 | 16:0 | 9.7 | |
| gill | 20:4n-6 | 12.6 | 20:5n-3 | 10.7 | 16:0 | 9.2 | 18:2n-6 | 9.0 | 18:1n-9 | 8.8 | |
| intestine | 20:2n-6 | 13.2 | 22:6n-3 | 12.4 | 20:5n-3 | 11.3 | 18:0 | 8.9 | 18:2n-6 | 8.7 | |
| skin | 16:0 | 13.0 | 22:6n-3 | 10.4 | 18:3n-3 | 9.2 | 18:2n-6 | 7.9 | 18:1n-9 | 7.8 | |
| liver | 22:6n-3 | 24.1 | 18:2n-6 | 15.8 | 20:5n-3 | 10.9 | 18:3n-3 | 10.2 | 16:0 | 9.0 | |
| muscle | 18:2n-6 | 19.4 | 18:0 | 15.3 | 16:0 | 10.0 | 22:6n-3 | 8.7 | 18:3n-3 | 8.5 | |
| gonad | 18:1n-9 | 17.9 | 22:6n-3 | 10.1 | 14:0 | 9.9 | 20:5n-3 | 9.5 | 20:3n-3 | 8.9 | |
1 1st, 2nd, 3rd, 4th, and 5th DFAsex is the fatty acid contributing the most, the second, the third, the fourth, and the fifth largely to the total fatty acid difference between sexes, respectively. 2 Contrib%—contribution % of a certain fatty acid difference to the total fatty acid difference between sexes. 3 Underlined fatty acids had higher content in males compared to pseudomales. 4 The WF3 group did not have enough pseudomales for this analysis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, J.; Zhang, F.; Ma, Q.; Wei, Y.; Liang, M.; Xu, H. Sex Differences in Fatty Acid Composition of Chinese Tongue Sole (Cynoglossus semilaevis) Tissues. Fishes 2023, 8, 421. https://doi.org/10.3390/fishes8080421
AMA Style
Liu J, Zhang F, Ma Q, Wei Y, Liang M, Xu H. Sex Differences in Fatty Acid Composition of Chinese Tongue Sole (Cynoglossus semilaevis) Tissues. Fishes. 2023; 8(8):421. https://doi.org/10.3390/fishes8080421
Chicago/Turabian StyleLiu, Jiahao, Feiran Zhang, Qiang Ma, Yuliang Wei, Mengqing Liang, and Houguo Xu. 2023. "Sex Differences in Fatty Acid Composition of Chinese Tongue Sole (Cynoglossus semilaevis) Tissues" Fishes 8, no. 8: 421. https://doi.org/10.3390/fishes8080421