Glucocorticoid Receptor Mediates Cortisol Regulation of Glycogen Metabolism in Gills of the Euryhaline Tilapia (Oreochromis mossambicus)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Cortisol and Receptor Antagonist Treatment of Tilapia Larvae
2.3. Total RNA Extraction
2.4. Real-Time PCR
2.5. RNA Probe Synthesis
2.6. In Situ Hybridization
2.7. Immunohistochemistry (IHC) of Gill Sections
2.8. Statistical Analysis
3. Results
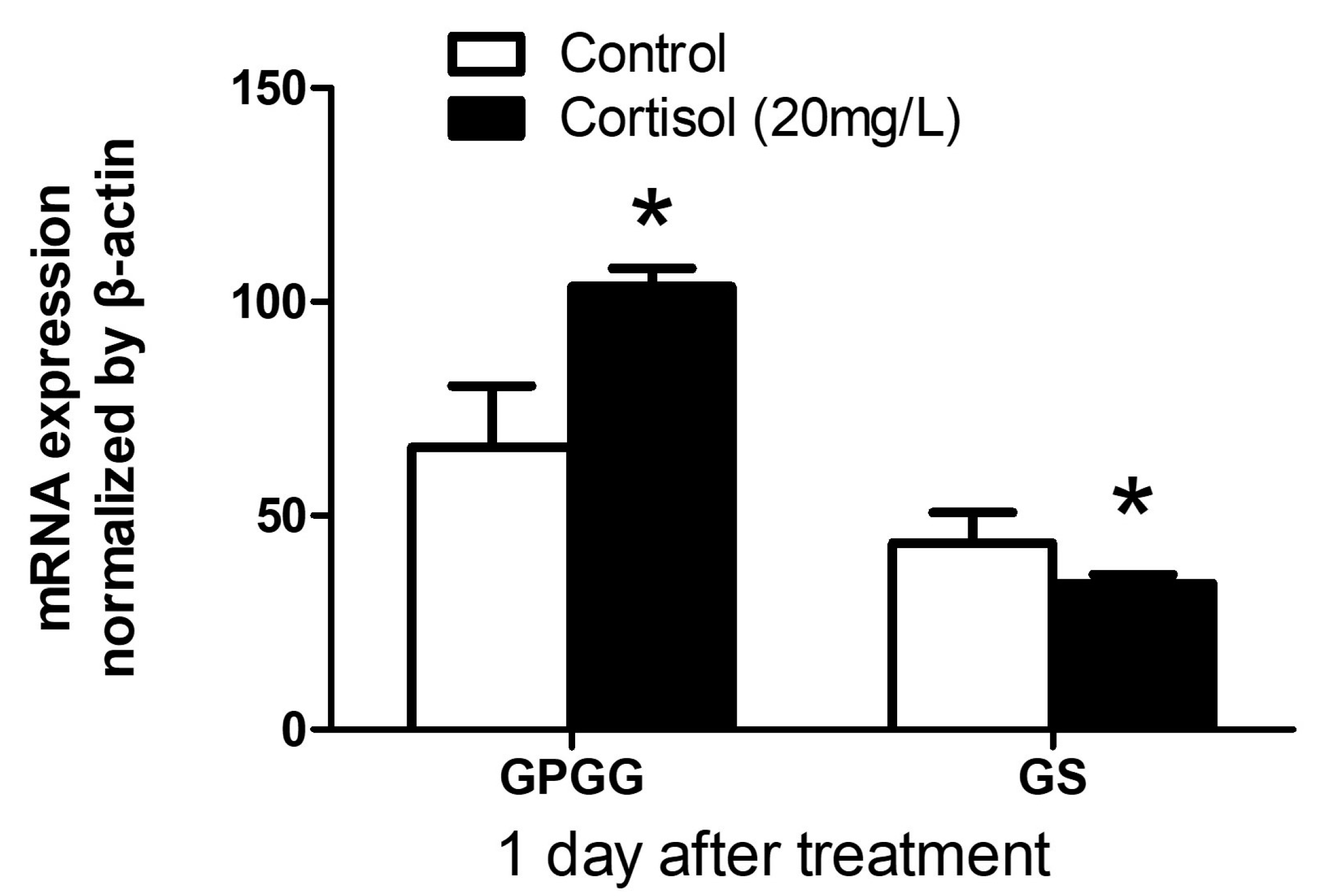
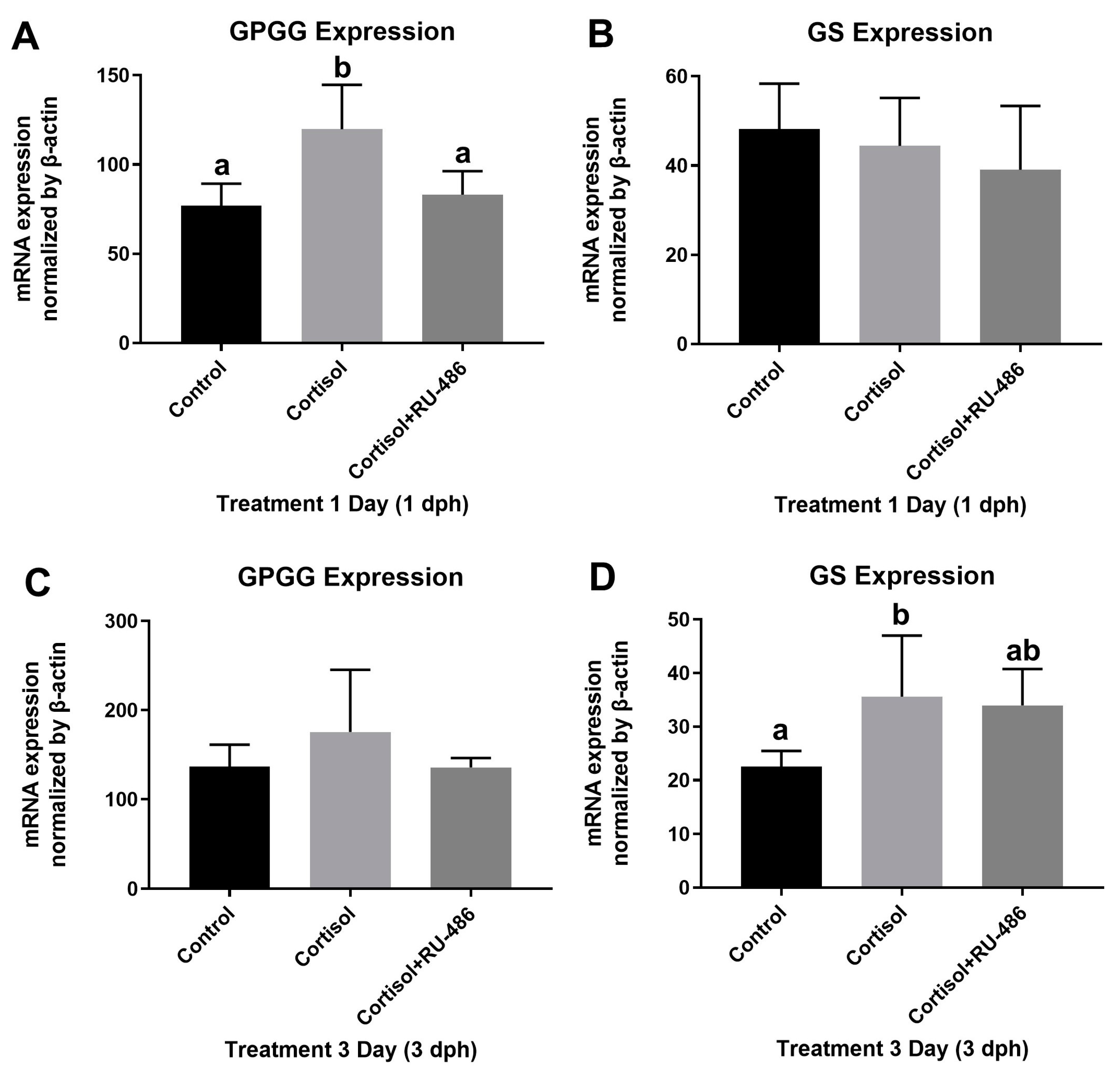
3.1. GPGG and GS mRNA Expression after Exogenous Cortisol Exposure
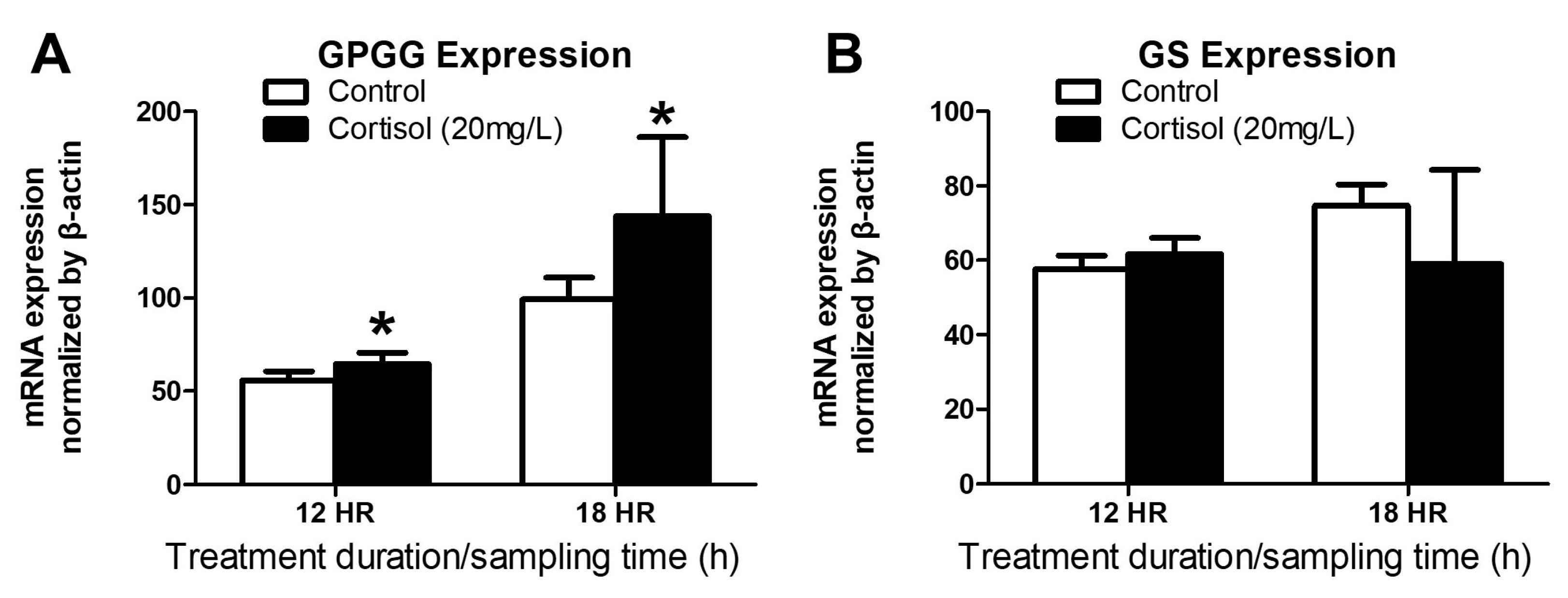
3.2. GPGG and GS mRNA Expression after Exogenous Cortisol Exposure from 12 to 18 h
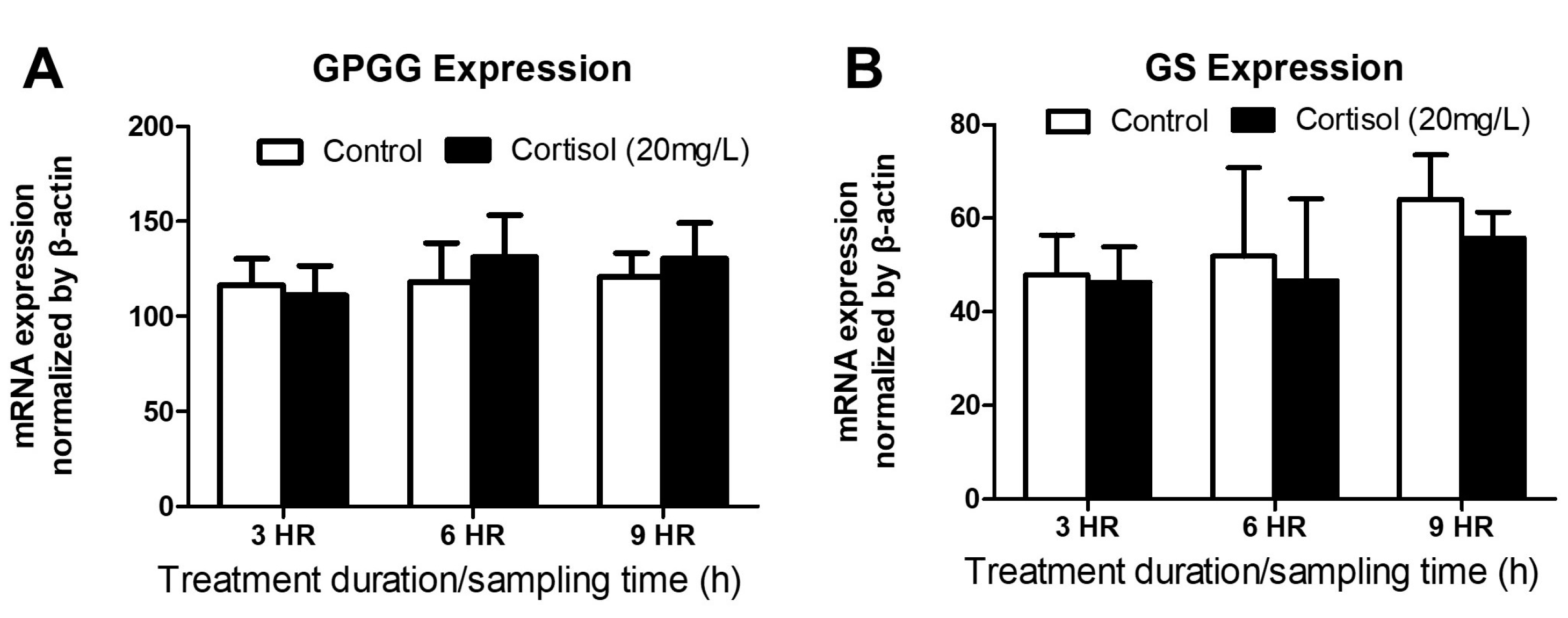
3.3. GPGG and GS mRNA Expression after Exogenous Cortisol Exposure from 3 to 9 h
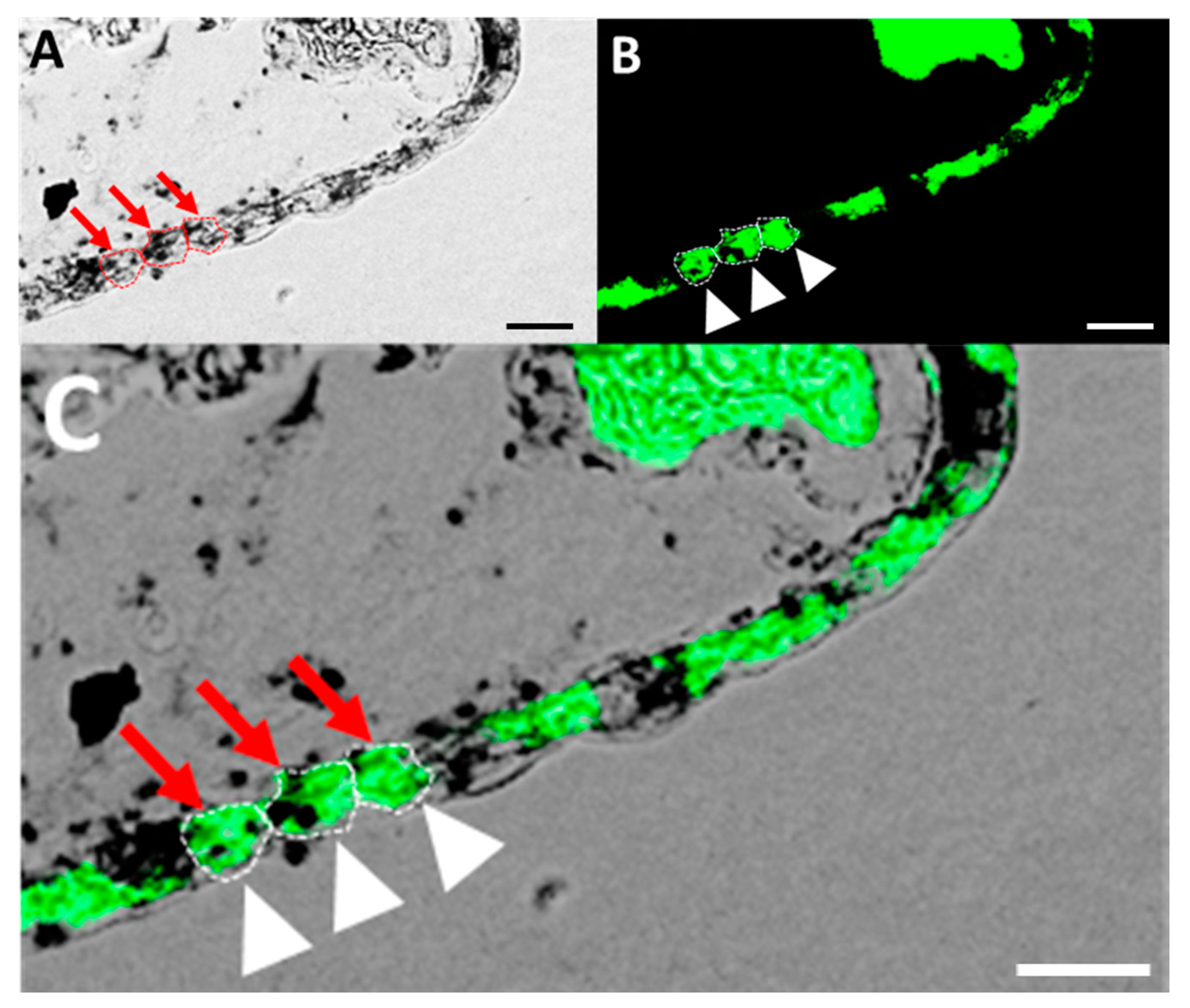
3.4. Localization of GR mRNA and Glycogen-Rich Cells in Adult Tilapia Gill Sections
3.5. GPGG mRNA Expression after GR Antagonist Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roach, P.J.; Cheng, C.; Huang, D.; Lin, A.; Mu, J.; Skurat, A.V.; Wilson, W.; Zhai, L. Novel Aspects of the Regulation of Glycogen Storage. J. Basic Clin. Physiol. Pharmacol. 1998, 9, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Bacca, H.; Huvet, A.; Fabioux, C.; Daniel, J.Y.; Delaporte, M.; Pouvreau, S.; Van Wormhoudt, A.; Moal, J. Molecular cloning and seasonal expression of oyster glycogen phosphorylase and glycogen synthase genes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 140, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J. Some Features of Glycogen Metabolism in Human Skeletal Muscle. Bibl. Nutr. Dieta 1979, 27, 121–125. [Google Scholar]
- Oliveira, G.T.; Rossi, I.C.; Kucharski, L.C.; Da Silva, R.S.M. Hepatopancreas gluconeogenesis and glycogen content during fasting in crabs previously maintained on a high-protein or carbohydrate-rich diet. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2004, 137, 383–390. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Tsai, S.-C.; Lin, H.-C.; Hsiao, C.-D.; Wu, S.M. Changes of glycogen metabolism in the gills and hepatic tissue of tilapia (Oreochromis mossambicus) during short-term Cd exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 296–304. [Google Scholar] [CrossRef]
- Van Der Boon, J.; Van Den Thillart, G.E.E.J.M.; Addink, A.D.F. The effects of cortisol administration on intermediary metabolism in teleost fish. Comp. Biochem. Physiol. Part A Physiol. 1991, 100, 47–53. [Google Scholar] [CrossRef]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the Stress Response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- McLaughlin, F.; Mackintosh, J.; Hayes, B.P.; McLaren, A.; Uings, I.J.; Salmon, P.; Humphreys, J.; Meldrum, E.; Farrow, S.N. Glucocorticoid-induced osteopenia in the mouse as assessed by histomorphometry, microcomputed tomography, and biochemical markers. Bone 2002, 30, 924–930. [Google Scholar] [CrossRef]
- Baker, M.E. Evolution of Glucocorticoid and Mineralocorticoid Responses: Go Fish. Endocrinology 2003, 144, 4223–4225. [Google Scholar] [CrossRef]
- Takahashi, H.; Sakamoto, T. The role of ‘mineralocorticoids’ in teleost fish: Relative importance of glucocorticoid signaling in the osmoregulation and ‘central’ actions of mineralocorticoid receptor. Gen. Comp. Endocrinol. 2013, 181, 223–228. [Google Scholar] [CrossRef]
- Carroll, S.M.; Ortlund, E.A.; Thornton, J.W. Mechanisms for the Evolution of a Derived Function in the Ancestral Glucocorticoid Receptor. PLoS Genet. 2011, 7, e1002117. [Google Scholar] [CrossRef]
- Bridgham, J.T.; Carroll, S.M.; Thornton, J.W. Evolution of Hormone-Receptor Complexity by Molecular Exploitation. Science 2006, 312, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. Chapter 3—The Endocrinology of the Stress Response in Fish: An Adaptation-Physiological View. In Fish Physiology; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 75–111. [Google Scholar]
- Hwang, P.-P.; Lee, T.-H.; Lin, L.-Y. Ion regulation in fish gills: Recent progress in the cellular and molecular mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R28–R47. [Google Scholar] [CrossRef]
- Hwang, P.-P.; Tung, Y.-C.; Chang, M.-H. Effect of environmental calcium levels on calcium uptake in tilapia larvae Oreochromis mossambicus. Fish Physiol. Biochem. 1996, 15, 363–370. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.D.; Bradshaw, D. Hormonal control of salt and water balance in vertebrates. Gen. Comp. Endocrinol. 2006, 147, 3–8. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.D. Endocrine Control of Osmoregulation in Teleost Fish. Am. Zool. 2001, 41, 781–794. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The Multifunctional Fish Gill: Dominant Site of Gas Exchange, Osmoregulation, Acid-Base Regulation, and Excretion of Nitrogenous Waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Redding, J.M.; Schreck, C.B. Influence of Ambient Salinity on Osmoregulation and Cortisol Concentration in Yearling Coho Salmon during Stress. Trans. Am. Fish. Soc. 1983, 112, 800–807. [Google Scholar] [CrossRef]
- Nichols, D.J.; Weisbart, M. Cortisol dynamics during seawater adaptation of Atlantic salmon Salmo salar. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1985, 248, R651–R659. [Google Scholar] [CrossRef]
- Patiño, R.; Redding, J.M.; Schreck, C.B. Interrenal secretion of corticosteroids and plasma cortisol and cortisone concentrations after acute stress and during seawater acclimation in juvenile coho salmon (Oncorhynchus kisutch). Gen. Comp. Endocrinol. 1987, 68, 431–439. [Google Scholar] [CrossRef]
- Sheridan, M.A. Alterations in lipid metabolism accompanying smoltification and seawater adaptation of salmonid fish. Aquaculture 1989, 82, 191–203. [Google Scholar] [CrossRef]
- Franklin, C.E.; Forster, M.E.; Davison, W. Plasma cortisol and osmoregulatory changes in sockeye salmon transferred to sea water: Comparison between successful and unsuccessful adaptation. J. Fish Biol. 1992, 41, 113–122. [Google Scholar] [CrossRef]
- Dean, D.B.; Whitlow, Z.W.; Borski, R.J. Glucocorticoid receptor upregulation during seawater adaptation in a euryhaline teleost, the tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 2003, 132, 112–118. [Google Scholar] [CrossRef]
- Kammerer, B.D.; Cech, J.J.; Kültz, D. Rapid changes in plasma cortisol, osmolality, and respiration in response to salinity stress in tilapia (Oreochromis mossambicus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 157, 260–265. [Google Scholar] [CrossRef]
- Hirose, S.; Kaneko, T.; Naito, N.; Takei, Y. Molecular biology of major components of chloride cells. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136, 593–620. [Google Scholar] [CrossRef]
- Chang, J.C.-H.; Wu, S.-M.; Tseng, Y.-C.; Lee, Y.-C.; Baba, O.; Hwang, P.-P. Regulation of glycogen metabolism in gills and liver of the euryhaline tilapia (Oreochromis mossambicus) during acclimation to seawater. J. Exp. Biol. 2007, 210, 3494–3504. [Google Scholar] [CrossRef]
- Tseng, Y.-C.; Huang, C.-J.; Chang, J.C.-H.; Teng, W.-Y.; Baba, O.; Fann, M.-J.; Hwang, P.-P. Glycogen phosphorylase in glycogen-rich cells is involved in the energy supply for ion regulation in fish gill epithelia. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 293, R482–R491. [Google Scholar] [CrossRef]
- Uchida, K.; Kaneko, T.; Miyazaki, H.; Hasegawa, S.; Hirano, T. Excellent Salinity Tolerance of Mozambique Tilapia: Elevated Chloride Cell Activity in the Branchial and Opercular Epithelia of the Fish Adapted to Concentrated Seawater. Zool. Sci. 2000, 17, 149–160. [Google Scholar] [CrossRef]
- Stickney, R.R. Tilapia Tolerance of Saline Waters: A Review. Progress. Fish-Cult. 1986, 48, 161–167. [Google Scholar] [CrossRef]
- Suresh, A.V.; Lin, C.K. Tilapia culture in saline waters: A review. Aquaculture 1992, 106, 201–226. [Google Scholar] [CrossRef]
- Hiroi, J.; Yasumasu, S.; McCormick, S.D.; Hwang, P.-P.; Kaneko, T. Evidence for an apical Na–Cl cotransporter involved in ion uptake in a teleost fish. J. Exp. Biol. 2008, 211, 2584–2599. [Google Scholar] [CrossRef] [PubMed]
- Cruz, S.A.; Chao, P.-L.; Hwang, P.-P. Cortisol promotes differentiation of epidermal ionocytes through Foxi3 transcription factors in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Hu, H.-J.; Hwang, P.-P. Cortisol regulates sodium homeostasis by stimulating the transcription of sodium-chloride transporter (NCC) in zebrafish (Danio rerio). Mol. Cell. Endocrinol. 2016, 422, 93–102. [Google Scholar] [CrossRef]
- Lin, C.-H.; Shih, T.-H.; Liu, S.-T.; Hsu, H.-H.; Hwang, P.-P. Cortisol Regulates Acid Secretion of H+-ATPase-rich Ionocytes in Zebrafish (Danio rerio) Embryos. Front. Physiol. 2015, 6, 328. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Tsai, I.L.; Su, C.-H.; Tseng, D.-Y.; Hwang, P.-P. Reverse Effect of Mammalian Hypocalcemic Cortisol in Fish: Cortisol Stimulates Ca2+ Uptake via Glucocorticoid Receptor-Mediated Vitamin D3 Metabolism. PLoS ONE 2011, 6, e23689. [Google Scholar] [CrossRef]
- Lin, G.R.; Weng, C.F.; Wang, J.I.; Hwang, P.P. Effects of Cortisol on Ion Regulation in Developing Tilapia (Oreochromis mossambicus) Larvae on Seawater Adaptation. Physiol. Biochem. Zool. 1999, 72, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Kuan, W.-C.; Liao, B.-K.; Deng, A.-N.; Tseng, D.-Y.; Hwang, P.-P. Environmental and cortisol-mediated control of Ca2+ uptake in tilapia (Oreochromis mossambicus). J. Comp. Physiol. B 2016, 186, 323–332. [Google Scholar] [CrossRef]
- Kiilerich, P.; Kristiansen, K.; Madsen, S.S. Cortisol regulation of ion transporter mRNA in Atlantic salmon gill and the effect of salinity on the signaling pathway. J. Endocrinol. 2007, 194, 417–427. [Google Scholar] [CrossRef]
- Kiilerich, P.; Tipsmark, C.K.; Borski, R.J.; Madsen, S.S. Differential effects of cortisol and 11-deoxycorticosterone on ion transport protein mRNA levels in gills of two euryhaline teleosts, Mozambique tilapia (Oreochromis mossambicus) and striped bass (Morone saxatilis). J. Endocrinol. 2011, 209, 115–126. [Google Scholar] [CrossRef]
- Hsiao, C.-D.; You, M.-S.; Guh, Y.-J.; Ma, M.; Jiang, Y.-J.; Hwang, P.-P. A Positive Regulatory Loop between foxi3a and foxi3b Is Essential for Specification and Differentiation of Zebrafish Epidermal Ionocytes. PLoS ONE 2007, 2, e302. [Google Scholar] [CrossRef]
- Han, T.-Y.; Wu, C.-Y.; Tsai, H.-C.; Cheng, Y.-P.; Chen, W.-F.; Lin, T.-C.; Wang, C.-Y.; Lee, J.-R.; Hwang, P.-P.; Lu, F.-I. Comparison of Calcium Balancing Strategies During Hypothermic Acclimation of Tilapia (Oreochromis mossambicus) and Goldfish (Carassius auratus). Front. Physiol. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-P.; Wu, S.-M. Role of Cortisol in Hypoosmoregulation in Larvae of the Tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 1993, 92, 318–324. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Qi, C.; Li, E.; Du, Z.; Qin, J.G.; Chen, L. Metabolic response of Nile tilapia (Oreochromis niloticus) to acute and chronic hypoxia stress. Aquaculture 2018, 495, 187–195. [Google Scholar] [CrossRef]
- Li, S.; Sang, C.; Zhang, J.; Chen, N.; Li, Z.; Jin, P.; Huang, X. Effects of acute hyperglycemia stress on plasma glucose, glycogen content, and expressions of glycogen synthase and phosphorylase in hybrid grouper (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂). Fish Physiol. Biochem. 2018, 44, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Liu, C.-J.; Lu, W.-J.; Wu, L.-Y.; Lai, K.-J.; Lin, Y.-T.; Lee, T.-H. Hypothermal Effects on Energy Supply for Ionocytes in Gills of Freshwater- and Seawater-Acclimated Milkfish, Chanos chanos. Front. Mar. Sci. 2022, 9, 880103. [Google Scholar] [CrossRef]
- Guo, T.; Yang, Y.; Meng, F.; Wang, S.; Xia, S.; Qian, Y.; Li, M.; Wang, R. Effects of low salinity on gill and liver glycogen metabolism of great blue-spotted mudskippers (Boleophthalmus pectinirostris). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 230, 108709. [Google Scholar] [CrossRef] [PubMed]
- Milligan, C.L. A regulatory role for cortisol in muscle glycogen metabolism in rainbow trout Oncorhynchus mykiss Walbaum. J. Exp. Biol. 2003, 206, 3167–3173. [Google Scholar] [CrossRef]
- Hwang, P.-P.; Chou, M.-Y. Zebrafish as an animal model to study ion homeostasis. Pflügers Arch. Eur. J. Physiol. 2013, 465, 1233–1247. [Google Scholar] [CrossRef]
- Takei, Y.; Hwang, P.-P. Chapter 6—Homeostatic Responses to Osmotic Stress. In Fish Physiology; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 207–249. [Google Scholar]
- Guh, Y.-J.; Hwang, P.-P. Insights into molecular and cellular mechanisms of hormonal actions on fish ion regulation derived from the zebrafish model. Gen. Comp. Endocrinol. 2017, 251, 12–20. [Google Scholar] [CrossRef]
- Yan, J.-J.; Hwang, P.-P. Novel discoveries in acid-base regulation and osmoregulation: A review of selected hormonal actions in zebrafish and medaka. Gen. Comp. Endocrinol. 2019, 277, 20–29. [Google Scholar] [CrossRef]
- Perry, S.F.; Laurent, P. Adaptational Responses of Rainbow Trout to Lowered External NaCl Concentration: Contribution of the Branchial Chloride Cell. J. Exp. Biol. 1989, 147, 147–168. [Google Scholar] [CrossRef]
- McCormick, S.D. Cortisol directly stimulates differentiation of chloride cells in tilapia opercular membrane. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1990, 259, R857–R863. [Google Scholar] [CrossRef]
- Uchida, K.; Kaneko, T.; Tagawa, M.; Hirano, T. Localization of Cortisol Receptor in Branchial Chloride Cells in Chum Salmon Fry. Gen. Comp. Endocrinol. 1998, 109, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, J.; Kaneko, T.; Tanaka, M. In vivo sequential changes in chloride cell morphology in the yolk-sac membrane of mozambique tilapia (Oreochromis mossambicus) embryos and larvae during seawater adaptation. J. Exp. Biol. 1999, 202, 3485–3495. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.C.; Chan, D.K.O. Chloride cell subtypes in the gill epithelium of Japanese eel Anguilla japonica. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1999, 277, R517–R522. [Google Scholar] [CrossRef] [PubMed]
- Sloman, K.A.; Desforges, P.R.; Gilmour, K.M. Evidence for a mineralocorticoid-like receptor linked to branchial chloride cell proliferation in freshwater rainbow trout. J. Exp. Biol. 2001, 204, 3953–3961. [Google Scholar] [CrossRef]
- Wong, C.K.; Chan, D.K. Effects of cortisol on chloride cells in the gill epithelium of Japanese eel, Anguilla japonica. J. Endocrinol. 2001, 168, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.R.; Keir, K.R.; Schulte, P.M. Effects of spironolactone and RU486 on gene expression and cell proliferation after freshwater transfer in the euryhaline killifish. J. Comp. Physiol. B 2005, 175, 499–510. [Google Scholar] [CrossRef]
- Shahsavarani, A.; Perry, S.F. Hormonal and environmental regulation of epithelial calcium channel in gill of rainbow trout (Oncorhynchus mykiss). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1490–R1498. [Google Scholar] [CrossRef]
- Cruz, S.A.; Lin, C.-H.; Chao, P.-L.; Hwang, P.-P. Glucocorticoid Receptor, but Not Mineralocorticoid Receptor, Mediates Cortisol Regulation of Epidermal Ionocyte Development and Ion Transport in Zebrafish (Danio rerio). PLoS ONE 2013, 8, e77997. [Google Scholar] [CrossRef] [PubMed]
- Trayer, V.; Hwang, P.-P.; Prunet, P.; Thermes, V. Assessment of the role of cortisol and corticosteroid receptors in epidermal ionocyte development in the medaka (Oryzias latipes) embryos. Gen. Comp. Endocrinol. 2013, 194, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Madsen, S.S. The role of cortisol and growth hormone in seawater adaptation and development of hypoosmoregulatory mechanisms in sea trout parr (Salmo trutta trutta). Gen. Comp. Endocrinol. 1990, 79, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Laurent, P.; Dunel-Erb, S.; Chevalier, C.; Lignon, J. Gill epithelial cells kinetics in a freshwater teleost, Oncorhynchus mykiss during adaptation to ion-poor water and hormonal treatments. Fish Physiol. Biochem. 1994, 13, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.P.; Lee, T.H.; Weng, C.F.; Fang, M.J.; Cho, G.Y. Presence of Na-K-ATPase in Mitochondria-Rich Cells in the Yolk-Sac Epithelium of Larvae of the Teleost Oreochromis mossambicus. Physiol. Biochem. Zool. 1999, 72, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-P.; Perry, S.F. Chapter 8—Ionic and acid–base regulation. In Fish Physiology; Perry, S.F., Ekker, M., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2010; Volume 29, pp. 311–344. [Google Scholar]
- Hwang, P.P. Salinity effects on development of chloride cells in the larvae of ayu (Plecoglossus altivelis). Mar. Biol. 1990, 107, 1–7. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-Y.; Lee, T.-H.; Tseng, D.-Y. Glucocorticoid Receptor Mediates Cortisol Regulation of Glycogen Metabolism in Gills of the Euryhaline Tilapia (Oreochromis mossambicus). Fishes 2023, 8, 267. https://doi.org/10.3390/fishes8050267
Wu C-Y, Lee T-H, Tseng D-Y. Glucocorticoid Receptor Mediates Cortisol Regulation of Glycogen Metabolism in Gills of the Euryhaline Tilapia (Oreochromis mossambicus). Fishes. 2023; 8(5):267. https://doi.org/10.3390/fishes8050267
Chicago/Turabian StyleWu, Chien-Yu, Tsung-Han Lee, and Deng-Yu Tseng. 2023. "Glucocorticoid Receptor Mediates Cortisol Regulation of Glycogen Metabolism in Gills of the Euryhaline Tilapia (Oreochromis mossambicus)" Fishes 8, no. 5: 267. https://doi.org/10.3390/fishes8050267
APA StyleWu, C.-Y., Lee, T.-H., & Tseng, D.-Y. (2023). Glucocorticoid Receptor Mediates Cortisol Regulation of Glycogen Metabolism in Gills of the Euryhaline Tilapia (Oreochromis mossambicus). Fishes, 8(5), 267. https://doi.org/10.3390/fishes8050267