
Regional Variation in Feeding Patterns of Sheepshead (Archosargus probatocephalus) in the Northwest Gulf of Mexico

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
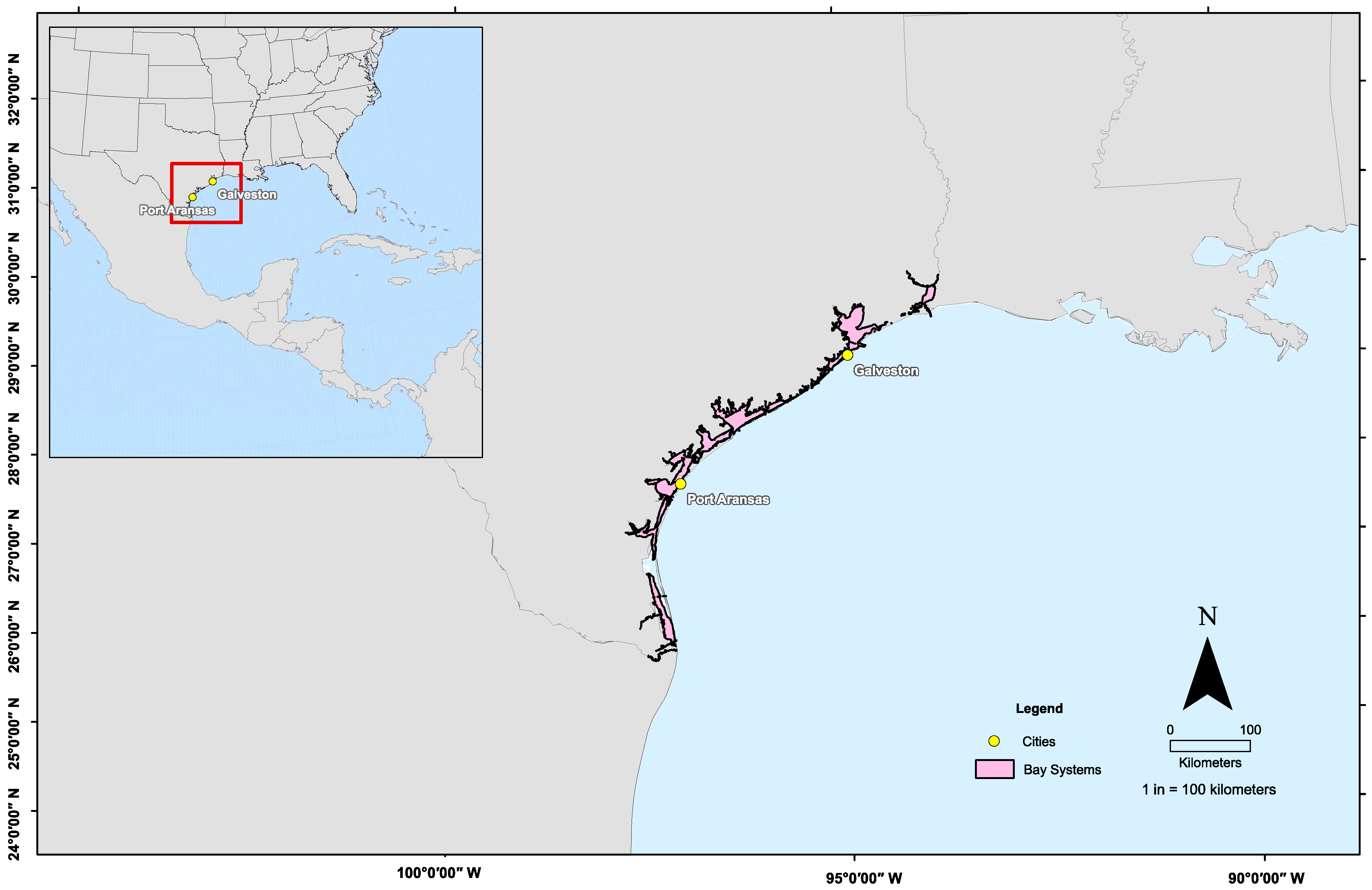
2.1. Sample Collection
2.2. Analysis of Stomach Contents
2.3. Stable Isotope Analysis
2.4. Data Analysis
2.4.1. Analysis of Stomach Contents
2.4.2. Stable Isotope Analysis
3. Results
3.1. Stomach Contents
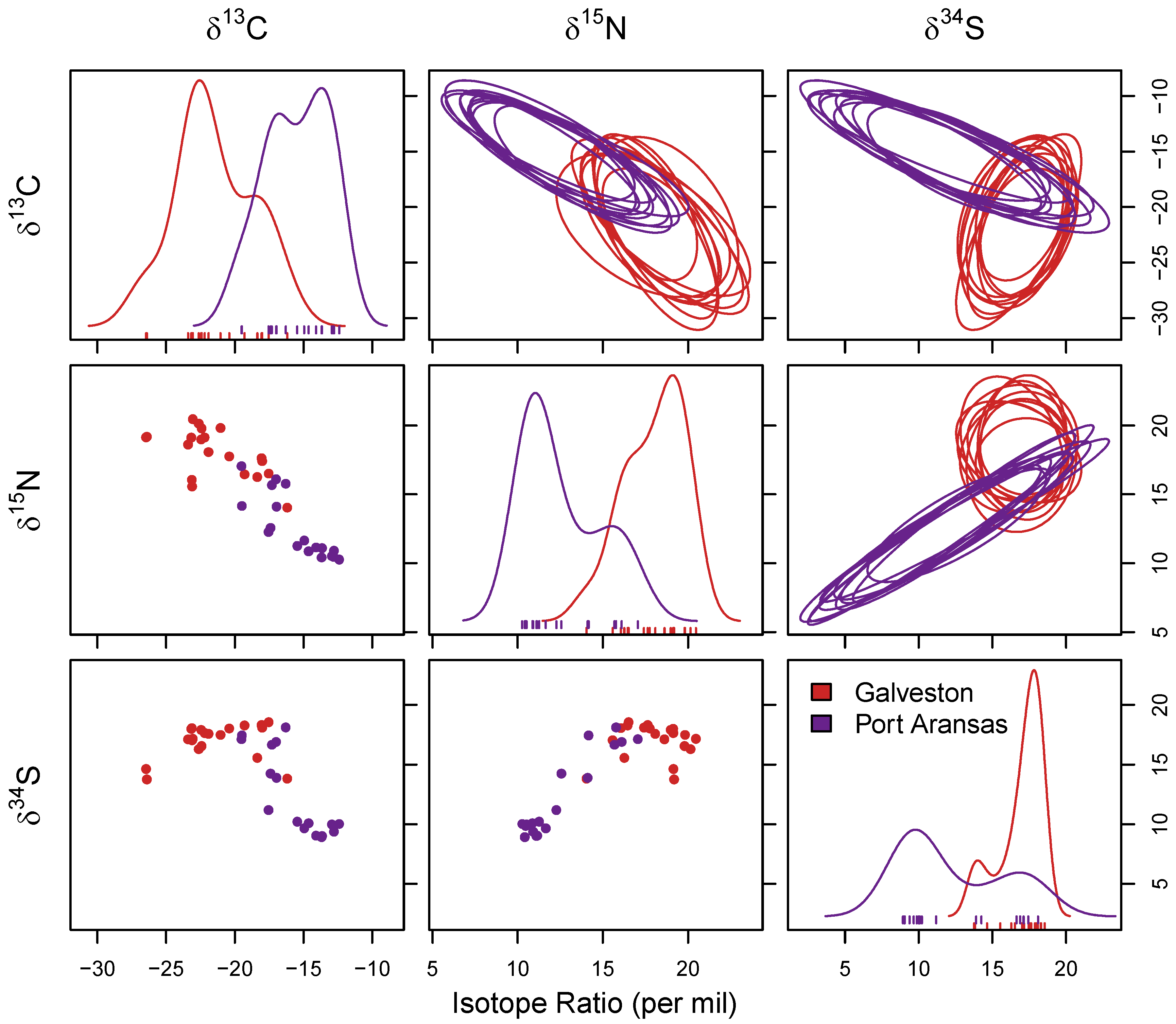
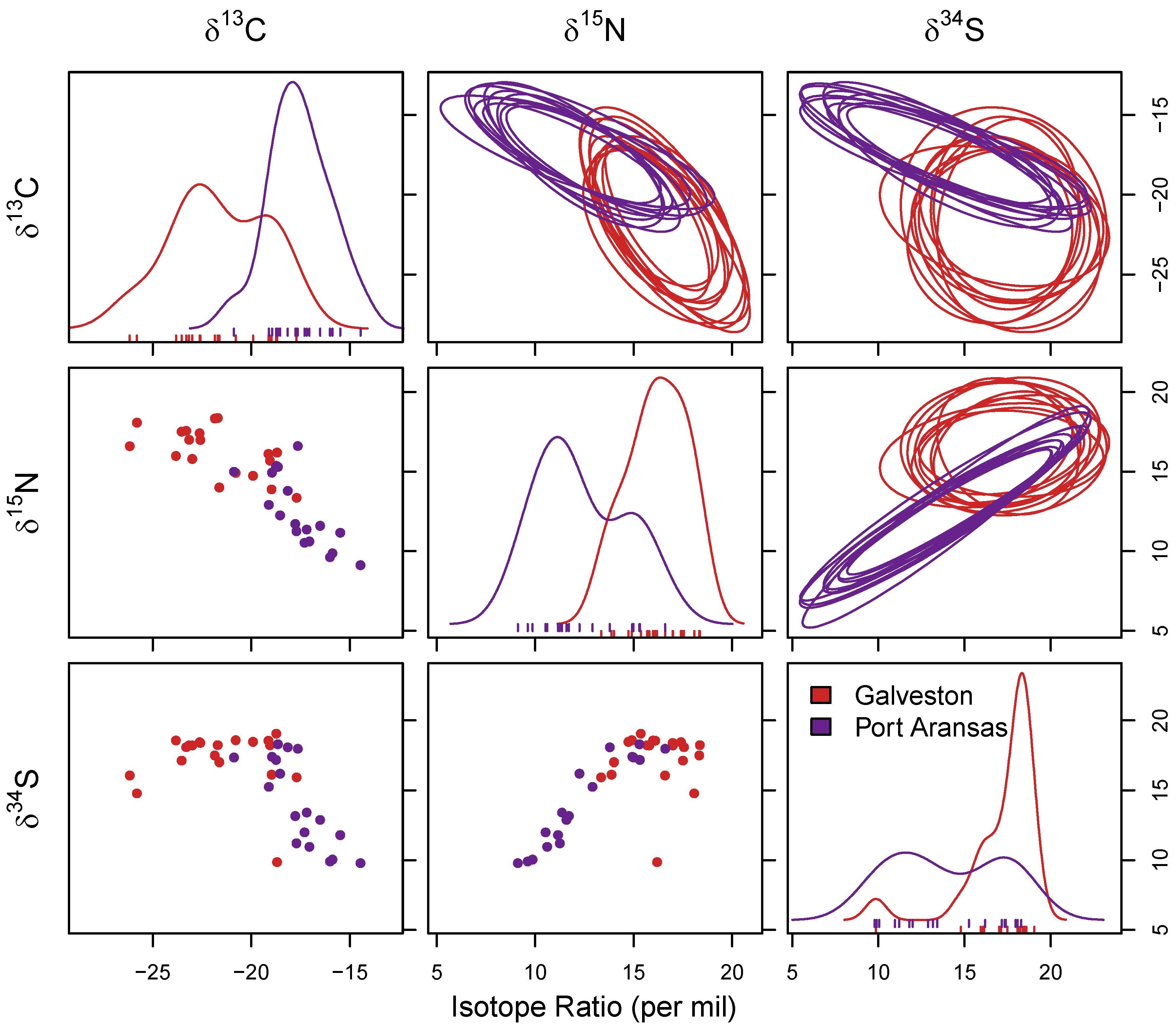
3.2. Stable Isotope Analysis
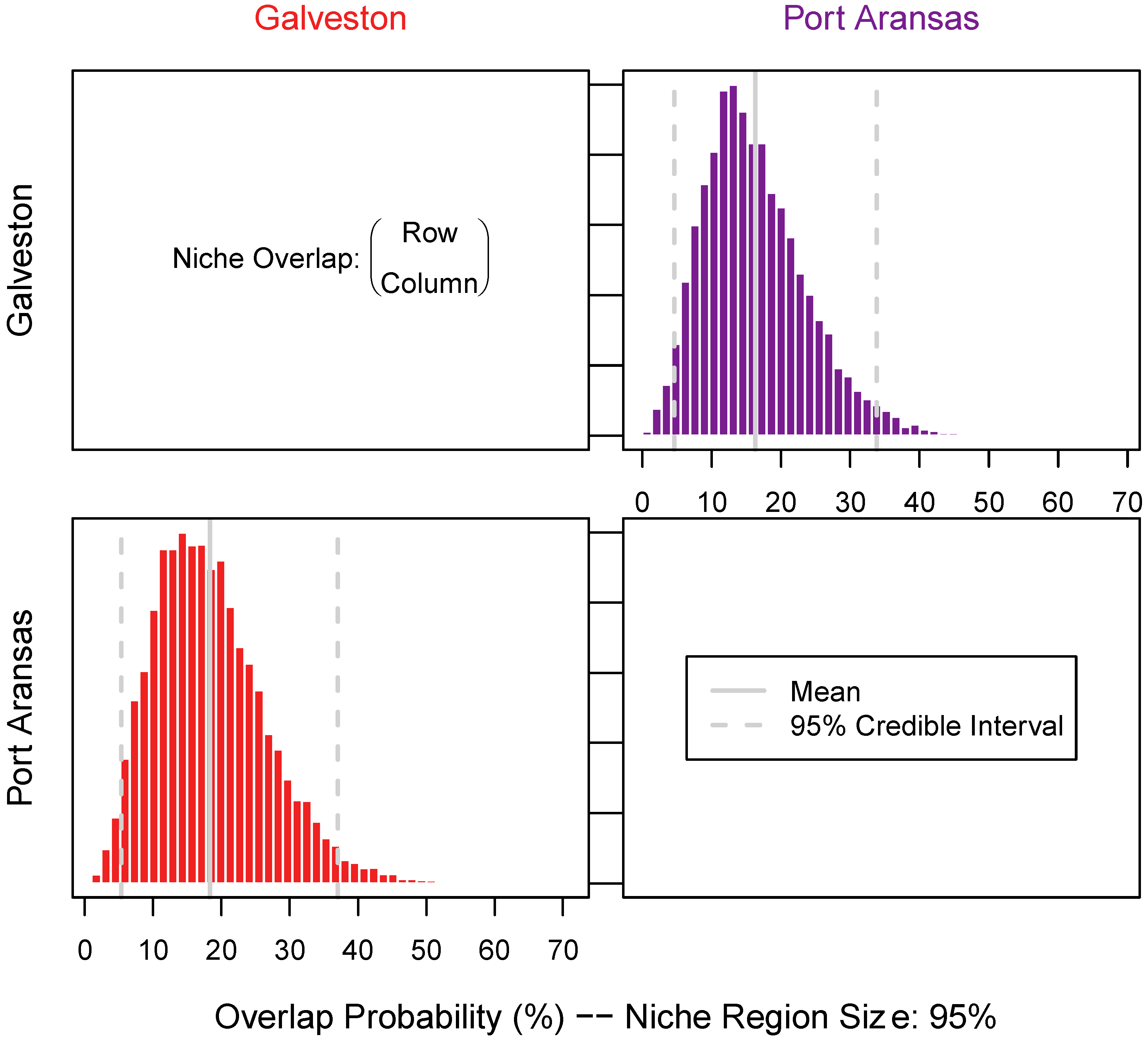
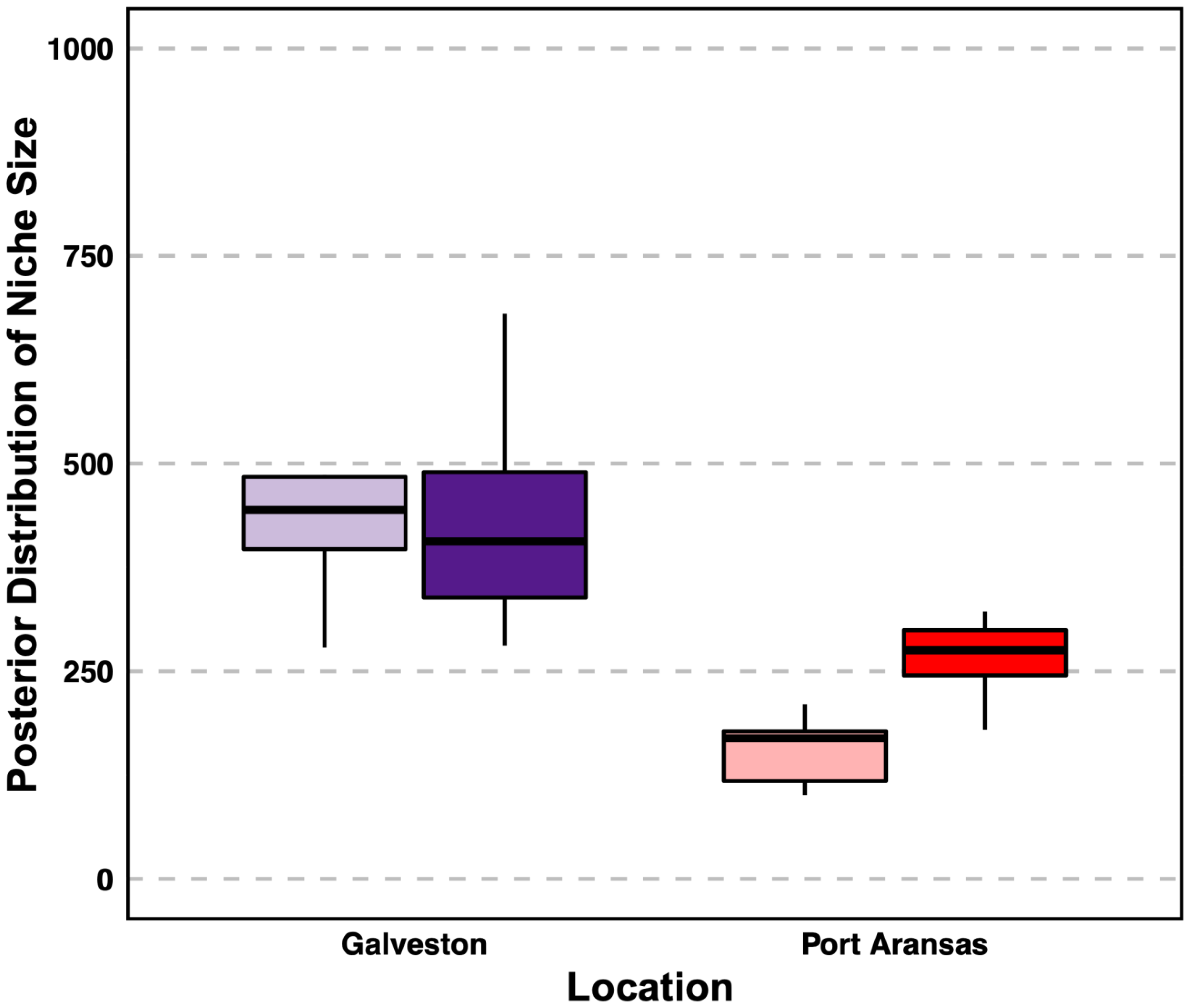
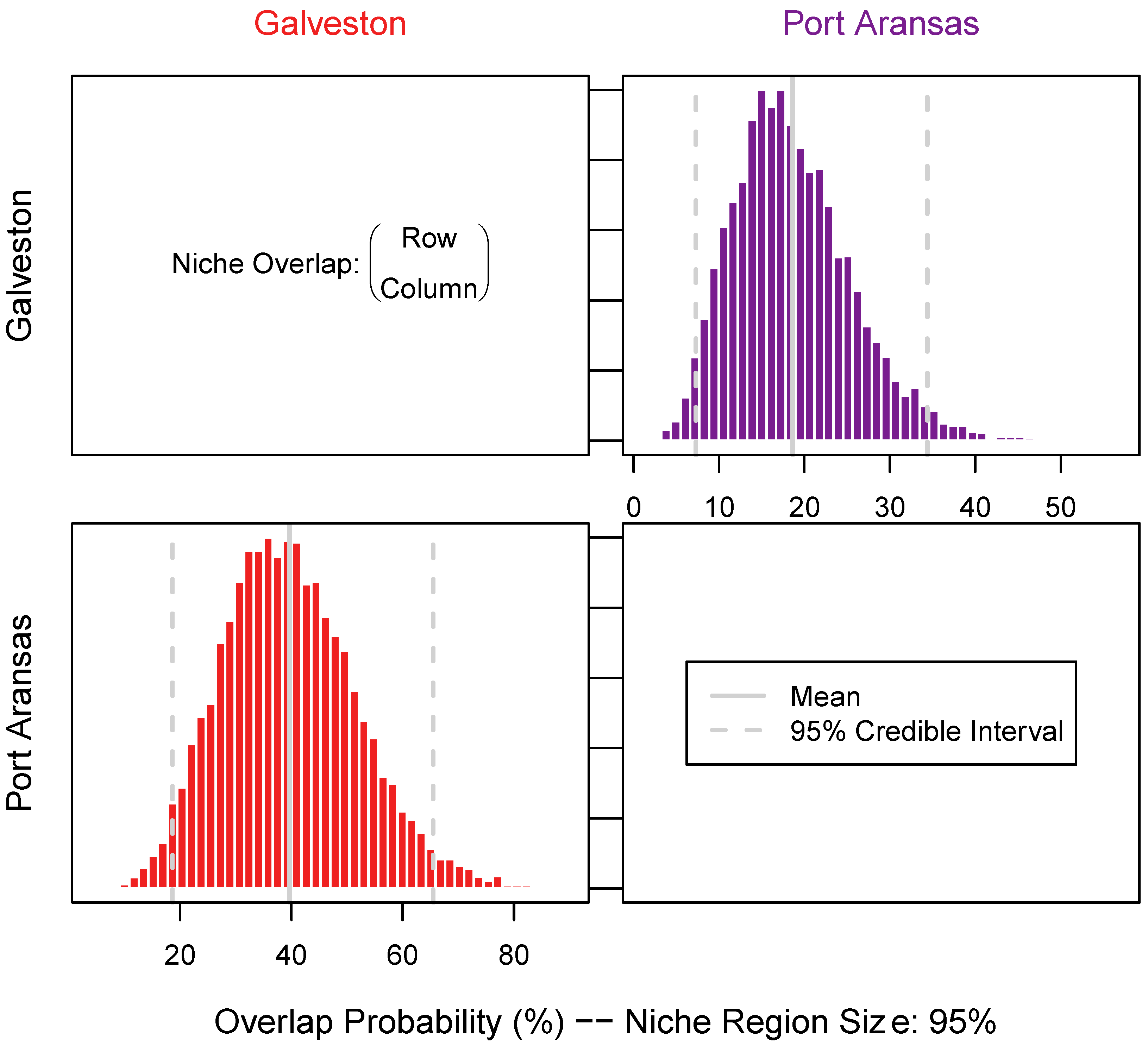
3.3. Niche Size and Overlap
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandez, L.P.H.; Motta, P.J. Trophic consequences of differential performance: Ontogeny of oral jaw-crushing performance in the sheepshead, Archosargus probatocephalus (Teleostei, Sparidae). J. Zool. 1997, 243, 737–756. [Google Scholar] [CrossRef]
- Personal Communication from the National Marine Fisheries Service; NOAA Fisheries, Fisheries Statistics Division: Silver Spring, MD, USA, 2013. Available online: https://www.fisheries.noaa.gov/foss/f?p=215:200:15293546732758:Mail:::: (accessed on 17 December 2022).
- VanderKooy, S.J. (Ed.) The Sheepshead Fishery of the Gulf of Mexico, United States: A Fisheries Profile; Gulf States Marine Fisheries Commission: Ocean Springs, MS, USA, 2006; Publication Number 143; 176p. [Google Scholar]
- Johnson, G.D. Development of Fishes of the Mid-Atlantic Bight: An Atlas of Egg, Larval, and Juvenile Stages. In Carangidae through Ephippidae; U.S. Fish and Wildlife Service: Washington, DC, USA, 1978; Volume 4. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Reeves, D.B.; Chesney, E.J.; Munnelly, R.T.; Baltz, D.M. Sheepshead Foraging Patterns at Oil and Gas Platforms in the Northern Gulf of Mexico. N. Am. J. Fish. Manag. 2018, 38, 1258–1274. [Google Scholar] [CrossRef]
- Sedberry, G.R. Feeding Habits of Sheepshead, Archosargus probatocephalus, in Offshore Reef Habitats of the Southeastern Continental Shelf. Gulf Mex. Sci. 1987, 9, 3. [Google Scholar] [CrossRef]
- Ballenger, J.C. Population Dynamics of Sheepshead (Archosargus probatocephalus; Walbaum 1792) in the Chesapeake Bay Region: A comparison to Other Areas and an Assessment of Their Current Status. Ph.D. Thesis, Old Dominion University, Norfolk, VA, USA, 2011. [Google Scholar] [CrossRef]
- Wenner, C.A.; Archambault, J. Sheepshead: The Natural History and Fishing Techniques in South Carolina; Charleston, SC Marine Resources Research Institute, Marine Resources Division, South Carolina Wildlife and Marine Resources Department: Charleston, SC, USA, 2006; p. 48.
- Dutka-Gianelli, J.; Murie, D. Age and growth of sheepshead, Archosargus probatocephalus (Pisces: Sparidae), from the northwest coast of Florida. Bull. Mar. Sci. 2001, 68, 69–83. [Google Scholar]
- Jennings, C.A. Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Gulf of Mexico): Sheepshead; The Service: Washington, DC, USA, 1985. [Google Scholar]
- Biggs, C.R.; Heyman, W.D.; Farmer, N.A.; Kobara, S.I.; Bolser, D.G.; Robinson, J.; Lowerre-Barbieri, S.K.; Erisman, B.E. The importance of spawning behavior in understanding the vulnerability of exploited marine fishes in the US Gulf of Mexico. PeerJ 2021, 9, e11814. [Google Scholar] [CrossRef]
- Helies, F.C.; Jamison, J.L. Prediction and Verification of Snapper-Grouper Spawning Aggregation Sites on the Offshore Banks of the Northwestern Gulf of Mexico; Final Report for Gulf and Atlantic Fisheries Foundation Inc.: Tampa, FL, USA; Gulf and Atlantic Fisheries Foundation Inc.: Tampa, FL, USA, 2016. [Google Scholar]
- Springer, V.G.; Woodburn, K.D. An Ecological Study of the Fishes of the Tampa Bay Area; Florida State Board of Conservation, Marine Laboratory: St. Petersburg, FL, USA, 1960. [Google Scholar]
- Heil, A.D. Life History, Diet, and Reproductive Dynamics of the Sheepshead (Archosargus probatocephalus) in the Northeastern Gulf of Mexico. Master’s Thesis, Florida State University, Tallahassee, FL, USA, 2017. [Google Scholar]
- Cresson, P.; Ruitton, S.; Harmelin-Vivien, M. Artificial reefs do increase secondary biomass production: Mechanisms evidenced by stable isotopes. Mar. Ecol. Prog. Ser. 2014, 509, 15–26. [Google Scholar] [CrossRef]
- Daigle, S.T.; Fleeger, J.W.; Cowan, J.H., Jr.; Pascal, P.-Y. What Is the Relative Importance of Phytoplankton and Attached Macroalgae and Epiphytes to Food Webs on Offshore Oil Platforms? Mar. Coast. Fish. 2013, 5, 53–64. [Google Scholar] [CrossRef]
- Dance, K.M.; Rooker, J.R.; Shipley, J.B.; Dance, M.A.; Wells, R.J.D. Feeding ecology of fishes associated with artificial reefs in the northwest Gulf of Mexico. PLoS ONE 2018, 13, e0203873. [Google Scholar] [CrossRef]
- Michener, R.; Lajtha, K. Stable Isotopes in Ecology and Environmental Science; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Peterson, B.J.; Fry, B. Stable Isotopes in Ecosystem Studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Plumlee, J.D.; Wells, R.J.D. Feeding ecology of three coastal shark species in the northwest Gulf of Mexico. Mar. Ecol. Prog. Ser. 2016, 550, 163–174. [Google Scholar] [CrossRef]
- Reeves, D.B.; Chesney, E.J.; Munnelly, R.T.; Baltz, D.M.; Maiti, K. Trophic Ecology of Sheepshead and Stone Crabs at Oil and Gas Platforms in the Northern Gulf of Mexico’s Hypoxic Zone. Trans. Am. Fish. Soc. 2019, 148, 324–338. [Google Scholar] [CrossRef]
- Cutwa, M.M.; Turingan, R.G. Intralocality Variation in Feeding Biomechanics and Prey Use in Archosargus probatocephalus (Teleostei, Sparidae), with Implications for the Ecomorphology of Fishes. Environ. Biol. Fishes 2000, 59, 191–198. [Google Scholar] [CrossRef]
- Cortés, E. A critical review of methods of studying fish feeding based on analysis of stomach contents: Application to elasmobranch fishes. Can. J. Fish. Aquat. Sci. 1997, 54, 726–738. [Google Scholar] [CrossRef]
- Pinkas, L.; Oliphant, M.S.; Iverson, I.L. Food Habits of Albacore, Bluefin Tuna and Bonito in California Waters; Fish Bulletin 152; UC San Diego Library: San Diego, CA, USA, 1970; pp. 1–139. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P. The Vegan Package: Community Ecology Package, R Package Version 2.0–2; R Core Team: Vienna, Austria, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Cortés, E. Standardized diet compositions and trophic levels of sharks. ICES J. Mar. Sci. 1999, 56, 707–717. [Google Scholar] [CrossRef]
- Swanson, H.K.; Lysy, M.; Power, M.; Stasko, A.D.; Johnson, J.D.; Reist, J.D. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology 2015, 96, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Biggs, C.; Erisman, B.; Heyman, W.; Kobara, S.; Farmer, N.; Lowerre-Barbieri, S.K.; Karnauskas, M.; Brenner, J. Fish Spawning Aggregations and Fisheries in the Gulf of Mexico: Interactions, Data Gaps and Research Priorities. In Proceedings of the 147th Annual Meeting of the American Fisheries Society, Tampa, FL, USA, 20–24 August 2017. [Google Scholar]
- Odum, W.E.; Heald, E.J. Trophic analyses of an estuarine mangrove community. Bull. Mar. Sci. 1972, 22, 671–738. [Google Scholar]
- Randall, J.E. Food Habits of Reef Fishes of the West Indies; Institute of Marine Sciences, University of Miami: Miami, FL, USA, 1967; Volume 5, pp. 665–847. [Google Scholar]
- TinHan, T.C.; Wells, R.D. Spatial and ontogenetic patterns in the trophic ecology of juvenile Bull Sharks (Carcharhinus leucas) from the Northwest Gulf of Mexico. Front. Mar. Sci. 2021, 8, 664316. [Google Scholar] [CrossRef]
- Egerton, J.P.; Bolser, D.G.; Grüss, A.; Erisman, B.E. Understanding patterns of fish backscatter, size and density around petroleum platforms of the US Gulf of Mexico using hydroacoustic data. Fish. Res. 2021, 233, 105752. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Palardy, J.E.; Witman, J.D. Water flow drives biodiversity by mediating rarity in marine benthic communities. Ecol. Lett. 2011, 14, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.M.; Cowan, J.H.; Lewis, K.A.; Simonsen, K.A. Method for estimating relative abundance and species composition around oil and gas platforms in the northern Gulf of Mexico, USA. Fish. Res. 2018, 201, 44–55. [Google Scholar] [CrossRef]
- Wilson, C.A.; Nieland, D.L. Age and growth of red snapper, Lutjanus campechanus, from the Northern Gulf of Mexico off Louisiana. Fish. Bull. 2001, 99, 653–665. [Google Scholar]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of Biodiversity Loss on Ocean Ecosystem Services. Science 2006, 314, 787. [Google Scholar] [CrossRef]
- Tolan, J.M. El Niño-Southern Oscillation impacts translated to the watershed scale: Estuarine salinity patterns along the Texas Gulf Coast, 1982 to 2004. Estuar. Coast. Shelf Sci. 2007, 72, 247–260. [Google Scholar] [CrossRef]
- Winfield, I.; Escobar-Briones, E.; Morrone, J.J. Updated checklist and identification of areas of endemism of benthic amphipods (Caprellidea and Gammaridea) from offshore habitats in the SW Gulf of Mexico. Sci. Mar. 2006, 70, 99–108. [Google Scholar] [CrossRef]
- Gibb, H.; Cunningham, S.A. Habitat contrasts reveal a shift in the trophic position of ant assemblages. J. Anim. Ecol. 2011, 80, 119–127. [Google Scholar] [CrossRef]
- Lei, J.; Jia, Y.; Wang, Y.; Lei, G.; Lu, C.; Saintilan, N.; Wen, L.I. Behavioural plasticity and trophic niche shift: How wintering geese respond to habitat alteration. Freshw. Biol. 2019, 64, 1183–1195. [Google Scholar] [CrossRef]
- Moosmann, M.; Cuenca-Cambronero, M.; De Lisle, S.; Greenway, R.; Hudson, C.M.; Lürig, M.D.; Matthews, B. On the evolution of trophic position. Ecol. Lett. 2021, 24, 2549–2562. [Google Scholar] [CrossRef] [PubMed]
- Tewfik, A.; Bell, S.S.; McCann, K.S.; Morrow, K. Predator diet and trophic position modified with altered habitat morphology. PLoS ONE 2016, 11, e0147759. [Google Scholar] [CrossRef]
- Van Rijssel, J.C.; Hecky, R.E.; Kishe-Machumu, M.A.; Witte, F. Changing ecology of Lake Victoria cichlids and their environment: Evidence from C13 and N15 analyses. Hydrobiologia 2017, 791, 175–191. [Google Scholar] [CrossRef]
- Reeds, K.A.; Smith, J.A.; Suthers, I.M.; Johnston, E.L. An ecological halo surrounding a large offshore artificial reef: Sediments, infauna, and fish foraging. Mar. Environ. Res. 2018, 141, 30–38. [Google Scholar] [CrossRef]
- Clynick, B.; Chapman, M.G.; Underwood, A. Effects of epibiota on assemblages of fish associated with urban structures. Mar. Ecol. 2007, 332, 201–210. [Google Scholar] [CrossRef]
- Gittman, R.K.; Scyphers, S.B.; Smith, C.S.; Neylan, I.P.; Grabowski, J.H. Ecological Consequences of Shoreline Hardening: A Meta-Analysis. BioScience 2016, 66, 763–773. [Google Scholar] [CrossRef]
- Reeves, D.B. The Influence of River Discharge on Fishes and Invertebrates Associated with Small Oil and Gas Platforms in Nearshore Louisiana. Ph.D. Thesis, Louisiana State University, Baton Rouge, LA, USA, 2018. [Google Scholar]
- Schulze, A.; Erdner, D.L.; Grimes, C.J.; Holstein, D.M.; Miglietta, M.P. Artificial Reefs in the Northern Gulf of Mexico: Community Ecology Amid the “Ocean Sprawl”. Front. Mar. Sci. 2020, 7, 447. [Google Scholar] [CrossRef]
- Kerr, L.A.; Secor, D.H.; Kraus, R.T. Stable isotope (δ13C and δ18O) and Sr/Ca composition of otoliths as proxies for environmental salinity experienced by an estuarine fish. Mar. Ecol. Prog. Ser. 2007, 349, 245–253. [Google Scholar] [CrossRef]
- Rooker, J.R.; Stunz, G.W.; Holt, S.A.; Minello, T.J. Population connectivity of red drum in the northern Gulf of Mexico. Mar. Ecol. Prog. Ser. 2010, 407, 187–196. [Google Scholar] [CrossRef]
- TinHan, T.C.; O’Leary, S.J.; Portnoy, D.S.; Rooker, J.R.; Gelpi, C.G.; Wells, R.J.D. Natural tags identify nursery origin of a coastal elasmobranch Carcharhinus leucas. J. Appl. Ecol. 2020, 57, 1222–1232. [Google Scholar] [CrossRef]
- Oczkowski, A.; Kreakie, B.; McKinney, R.A.; Prezioso, J. Patterns in Stable Isotope Values of Nitrogen and Carbon in Particulate Matter from the Northwest Atlantic Continental Shelf, from the Gulf of Maine to Cape Hatteras. Front. Mar. Sci. 2016, 3, 252. [Google Scholar] [CrossRef]
- Matich, P.; Shipley, O.N.; Weideli, O.C. Quantifying spatial variation in isotopic baselines reveals size-based feeding in a model estuarine predator: Implications for trophic studies in dynamic ecotones. Mar. Biol. 2021, 168, 108. [Google Scholar] [CrossRef]
- Dahlberg, M.D.; Odum, E.P. Annual cycles of species occurrence, abundance, and diversity in Georgia estuarine fish populations. Am. Midl. Nat. 1970, 83, 382–392. [Google Scholar] [CrossRef]
- Merino-Contreras, M.L.; Sánchez-Morales, F.; Jiménez-Badillo, M.L.; Peña-Marín, E.S.; Álvarez-González, C.A. Partial characterization of digestive proteases in sheepshead, Archosargus probatocephalus (Spariformes: Sparidae). Neotrop. Ichthyol. 2018, 16. [Google Scholar] [CrossRef]
- Pattillo, M.E.; Czapla, T.E.; Nelson, D.M.; Monaco, M.E. Distribution and Abundance of Fishes and Invertebrates in Gulf of Mexico Estuaries—Volume II: Species Life History Summaries; ELMR Report No. 11; NOAA/NOS Strategic Environmental Assessments Division: Silver Spring, MD, USA, 1997.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Group | Taxa | %N Number | %W Weight | %O Occurrence | %IRI | ||||
|---|---|---|---|---|---|---|---|---|---|
| Galveston | Port Aransas | Galveston | Port Aransas | Galveston | Port Aransas | Galveston | Port Aransas | ||
| AMPHIPODA | |||||||||
| Bateidae | 0.21 | 1.10 | 0.01 | 0.01 | 2.76 | 2.27 | 0.03 | 0.10 | |
| Caprellidae | 30.92 | 0.98 | 12.16 | 0.01 | 15.86 | 2.27 | 30.68 | 0.09 | |
| Corophiidae | 32.84 | 36.67 | 2.95 | 2.76 | 15.17 | 15.91 | 24.38 | 24.12 | |
| Gammaridae | 9.85 | 5.26 | 0.96 | 0.05 | 8.28 | 4.55 | 4.02 | 0.93 | |
| Unidentified | 2.29 | 9.41 | 0.39 | 0.33 | 2.76 | 2.27 | 0.33 | 0.85 | |
| Total (total %O nonadded) | 76.12 | 53.42 | 16.48 | 3.16 | 44.83 (23.71) | 27.27 (17.95) | 186.3 (61.79) | 26.08 (30.19) | |
| ANNELIDA | 0.01 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | |
| BRYOZOAN | 0.00 | 0.24 | 0.00 | 0.11 | 0.00 | 5.13 | 0.00 | 0.05 | |
| CNIDARIA | 0.02 | 0.00 | 0.03 | 0.00 | 2.06 | 0.00 | 0.00 | 0.00 | |
| DECAPODA | |||||||||
| Portunidae | 0.18 | 0.24 | 0.94 | 0.08 | 3.45 | 4.55 | 0.17 | 0.06 | |
| Unidentified | 14.81 | 32.03 | 1.47 | 2.75 | 6.21 | 2.27 | 4.54 | 3.04 | |
| Total (total %O nonadded) | 15.00 | 32.27 | 2.42 | 2.83 | 9.66 (9.28) | 6.82 (7.69) | 7.55 (4.55) | 3.10 (8.03) | |
| ISOPODA | 2.76 | 1.47 | 0.25 | 0.01 | 16.49 | 7.69 | 1.40 | 0.34 | |
| MOLLUSCA | |||||||||
| Bivalvia | 0.02 | 1.47 | 1.44 | 15.16 | 2.07 | 6.82 | 0.14 | 4.36 | |
| Gastropoda | 0.02 | 0.00 | 0.24 | 0.00 | 1.38 | 0.00 | 0.02 | 0.00 | |
| Total (total %O nonadded) | 0.05 | 1.47 | 1.68 | 15.16 | 3.45 (4.12) | 6.82 (7.69) | 0.27 (0.20) | 4.36 (3.80) | |
| SESSILIA | 1.31 | 5.01 | 8.63 | 52.61 | 11.34 | 23.08 | 3.17 | 39.53 | |
| OTHER | Plantae | 0.08 | 0.49 | 56.54 | 4.71 | 11.34 | 10.26 | 18.07 | 1.58 |
| UNIDENTIFIED | 4.67 | 5.62 | 13.96 | 21.41 | 20.62 | 20.51 | 10.81 | 16.48 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutierrez, E.M.; Plumlee, J.D.; Bolser, D.G.; Erisman, B.E.; Wells, R.J.D. Regional Variation in Feeding Patterns of Sheepshead (Archosargus probatocephalus) in the Northwest Gulf of Mexico. Fishes 2023, 8, 188. https://doi.org/10.3390/fishes8040188
Gutierrez EM, Plumlee JD, Bolser DG, Erisman BE, Wells RJD. Regional Variation in Feeding Patterns of Sheepshead (Archosargus probatocephalus) in the Northwest Gulf of Mexico. Fishes. 2023; 8(4):188. https://doi.org/10.3390/fishes8040188
Chicago/Turabian StyleGutierrez, Elsa M., Jeffrey D. Plumlee, Derek G. Bolser, Brad E. Erisman, and R. J. David Wells. 2023. "Regional Variation in Feeding Patterns of Sheepshead (Archosargus probatocephalus) in the Northwest Gulf of Mexico" Fishes 8, no. 4: 188. https://doi.org/10.3390/fishes8040188
APA StyleGutierrez, E. M., Plumlee, J. D., Bolser, D. G., Erisman, B. E., & Wells, R. J. D. (2023). Regional Variation in Feeding Patterns of Sheepshead (Archosargus probatocephalus) in the Northwest Gulf of Mexico. Fishes, 8(4), 188. https://doi.org/10.3390/fishes8040188