
Thermal Tolerance of Spotted Sea Bass (Lateolabrax maculatus) and Pearl Gentian Grouper (Epinephelus fuscoguttatus Female × E. lanceolatus Male) in Different Environmental Temperatures


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Management
2.2. Experiment Design
2.3. Experiment Management
2.4. Data Statistics and Analysis
3. Results
3.1. CTM Experiment in Spring
3.2. CTM Experiment in Summer
3.3. CTM Experiment in Autumn
3.4. CTM Experiment in Winter
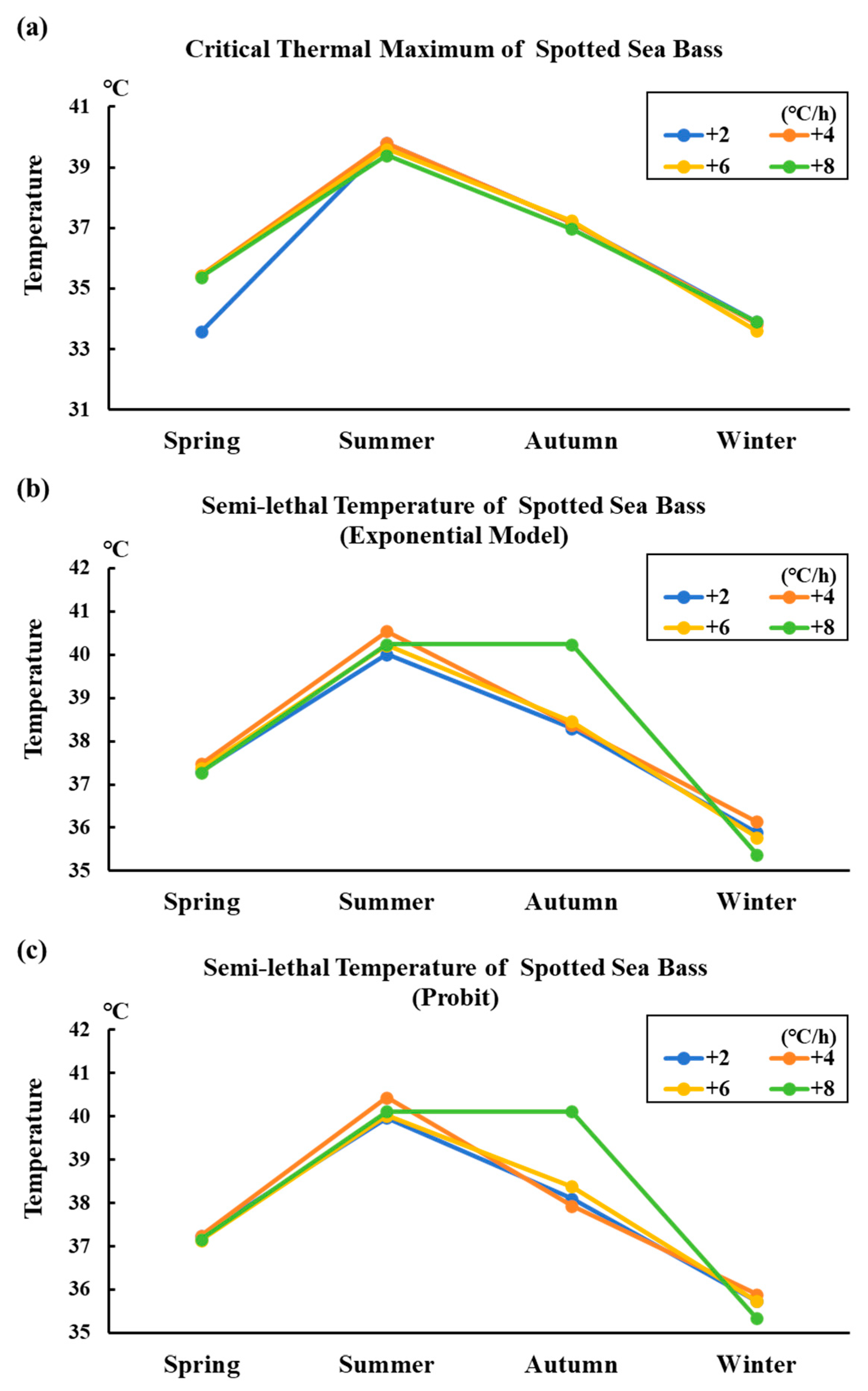
3.5. Variation Trend of Thermal Tolerance in Different Seasonal Conditions of Spotted Sea Bass
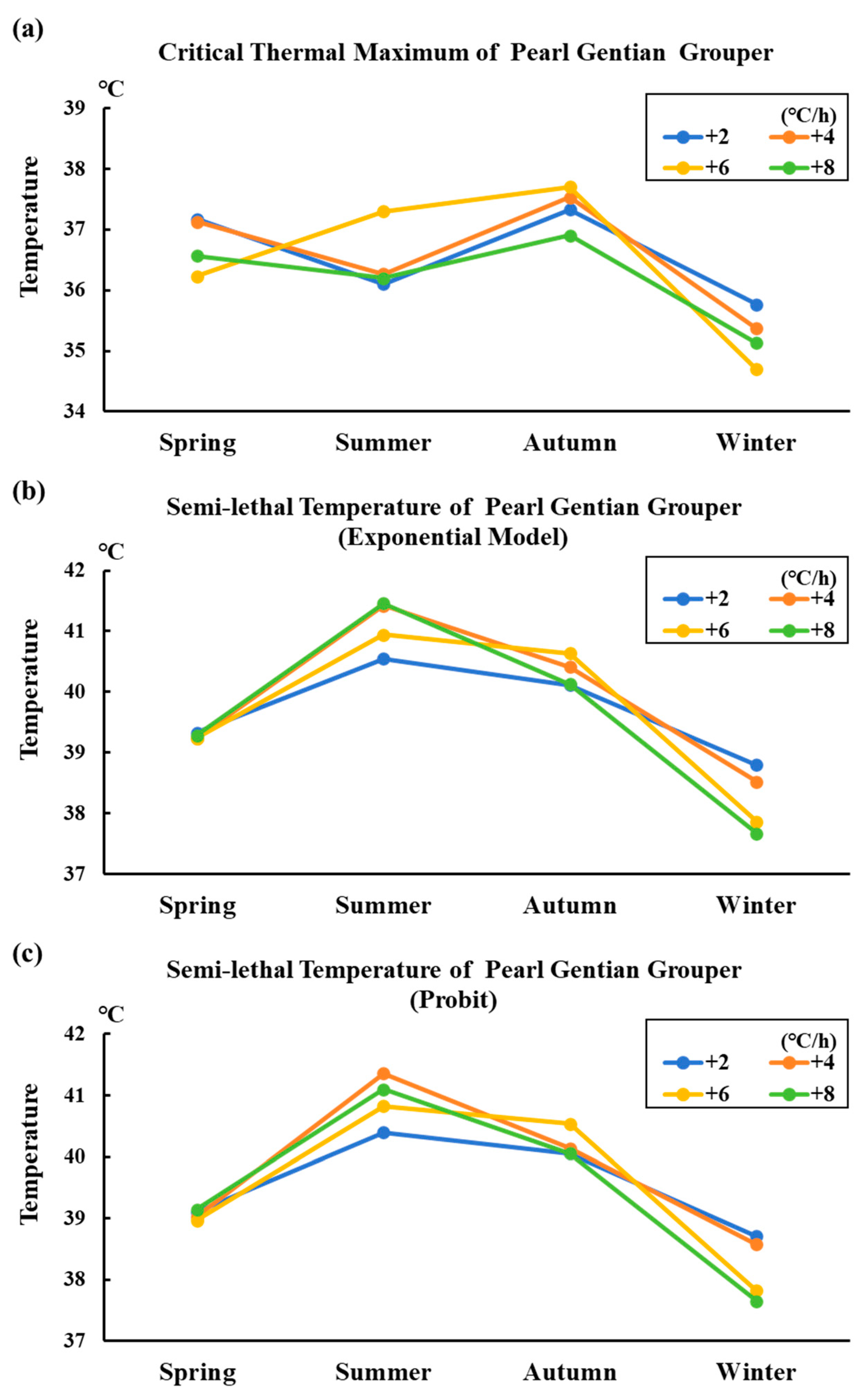
3.6. Variation Trend of Thermal Tolerance in Different Seasonal Conditions of Grouper
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Long, Z.Y.; Qin, H.H.; Huang, Z.F.; Xu, A.L.; Ye, Y.L.; Li, Z.B. Effects of heat stress on physiological parameters, biochemical parameters and expression of heat stress protein gene in Lateolabrax maculatus. J. Therm. Biol. 2023, 115, 103606. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chao, Y.C.; Jiang, T.L.; Chen, Z.S. Facilitating developments of solar thermal power and nuclear power generations for carbon neutral: A study based on evolutionary game theoretic method. Sci. Total Environ. 2022, 814, 151927. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.Y.; Guo, Z.Y.; Lin, Y.; Wei, C.; Ye, S.F. Application of satellite remote sensing data for monitoring thermal discharge pollution from Tianwan nuclear power plant in eastern China. In Proceedings of the 2012 5th International Congress on Image and Signal Processing (CISP), Chongqing, China, 16–18 October 2012; Chen, Q., Cheng, Q., Li, Y., Zhang, T., Wang, L., Eds.; IEEE: Piscataway, NJ, USA, 2012; pp. 1019–1023. [Google Scholar]
- Torres, M.; Bevia, F.R. Chlorine use reduction in nuclear or conventional power plants: A combined cooling-and-stripping tower for coastal power plants. J. Clean. Prod. 2012, 26, 1–8. [Google Scholar] [CrossRef]
- Zhang, J.J.; Wang, Y.B.; Ottmann, D.; Cao, P.; Yang, J.S.; Yu, J.B.; Lv, Z.B. Seasonal variability of phytoplankton community response to thermal discharge from nuclear power plant in temperate coastal area. Environ. Pollut. 2023, 318, 120898. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Y. Discussion on utilization of surplus heat of circulating cooling water of fossil fuel nuclear power plants. J. China Inst. Water Resour. Hydropower Res. 2004, 2, 81–86. (In Chinese) [Google Scholar] [CrossRef]
- Dolan, T.E.; Lynch, P.D.; Karazsia, J.L.; Serafy, J.E. Statistical power to detect change in a mangrove shoreline fish community adjacent to a nuclear power plant. Environ. Monit. Assess. 2016, 188, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.J.; Kristan, W.B.; Eme, J. Thermal tolerance and routine oxygen consumption of convict cichlid, Archocentrus nigrofasciatus, acclimated to constant temperatures (20 degrees C and 30 degrees C) and a daily temperature cycle (20 degrees C -> 30 degrees C). J. Comp. Physiol. B 2021, 191, 479–491. [Google Scholar] [CrossRef]
- Ortega, E.; Reyes, J.; Rodriguez, M.H. Growth, thermal preference and critical thermal maximum for Totoaba macdonaldi: Effect of acclimation temperature and inclusion of soybean meal in the diet. Lat. Am. J. Aquat. Res. 2021, 49, 258–271. [Google Scholar] [CrossRef]
- Penny, F.M.; Pavey, S.A. Increased acute thermal tolerance and little change to hematology following acclimation to warm water in juvenile striped bass, Morone saxatilis. Environ. Biol. Fish. 2021, 104, 489–500. [Google Scholar] [CrossRef]
- Bilyk, K.T.; Sformo, T.L. Varying heat tolerance among Arctic nearshore fishes. Polar Biol. 2021, 44, 607–612. [Google Scholar] [CrossRef]
- Kir, M.; Sunar, M.C.; Topuz, M.; Sariipek, M. Thermal acclimation capacity and standard metabolism of the Pacific white shrimp Litopenaeus vannamei (Boone, 1931) at different temperature and salinity combinations. J. Therm. Biol. 2023, 112, 103429. [Google Scholar] [CrossRef] [PubMed]
- Radtke, G.; Wolnicki, J.; Kapusta, A.; Przybylski, M.; Kaczkowski, Z. Critical thermal maxima of three small-bodied fish species (Cypriniformes) of different origin and protection status. Eur. Zool. J. 2022, 89, 1351–1361. [Google Scholar] [CrossRef]
- Cereja, R. Critical thermal maxima in aquatic ectotherms. Ecol. Indic. 2020, 119, 106856. [Google Scholar] [CrossRef]
- Rajaguru, S. Critical thermal maximum of seven estuarine fishes. J. Therm. Biol. 2002, 27, 125–128. [Google Scholar] [CrossRef]
- Qin, H.H.; Long, Z.Y.; Huang, Z.F.; Ma, J.R.; Kong, L.M.; Lin, Y.; Lin, H.; Zhou, S.S.; Li, Z.B. A comparison of the physiological responses to heat stress of two sizes of juvenile spotted seabass (Lateolabrax maculatus). Fishes 2023, 8, 340. [Google Scholar] [CrossRef]
- Messmer, V.; Pratchett, M.S.; Hoey, A.S.; Tobin, A.J.; Coker, D.J.; Cooke, S.J.; Clark, T.D. Global warming may disproportionately affect larger adults in a predatory coral reef fish. Global Change Biol. 2017, 23, 2230–2240. [Google Scholar] [CrossRef]
- Stavrakidis-Zachou, O.; Lika, K.; Pavlidis, M.; Asaad, M.H.; Papandroulakis, N. Metabolic scope, performance and tolerance of juvenile European sea bass Dicentrarchus labrax upon acclimation to high temperatures. PLoS ONE 2022, 17, e0272510. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Chen, C.S.; Chen, J.C. Salinity and temperature tolerance of brown-marbled grouper Epinephelus fuscoguttatus. Fish Physiol. Biochem. 2013, 39, 277–286. [Google Scholar] [CrossRef]
- Wu, Y.D.; Yu, X.K.; Suo, N.; Bai, H.Q.; Ke, Q.Z.; Chen, J.; Pan, Y.; Zheng, W.Q.; Xu, P. Thermal tolerance, safety margins and acclimation capacity assessments reveal the climate vulnerability of large yellow croaker aquaculture. Aquaculture 2022, 561, 738665. [Google Scholar] [CrossRef]
- Chen, X.M.; Li, J.K.; Wang, Z.Y.; Cai, M.Y.; Liu, X.D. Thermal tolerance evaluation and related microsatellite marker screening and identification in the large yellow croaker Larimichthys crocea. China J. Oceanol. Limn. 2017, 35, 566–571. [Google Scholar] [CrossRef]
- White, D.P.; Wahl, D.H. Growth and physiological responses in largemouth bass populations to environmental warming: Effects of inhabiting chronically heated environments. J. Therm. Biol. 2020, 88, 102467. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Zhou, Z.X.; Shi, Y.; Gong, J.; Li, C.Y.; Zhou, T.; Li, Y.; Zhang, D.C.; Xu, P. Genome-wide evolutionary signatures of climate adaptation in spotted sea bass inhabiting different latitudinal regions. Evol. Appl. 2023, 16, 1029–1043. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Season | Environmental Water Temperature (°C) | Average Weight of Fish (g) |
|---|---|---|
| Spotted sea bass | ||
| Spring | 21.6 | 3.10 ± 0.22 |
| Summer | 26.5 | 3.80 ± 0.80 |
| Autumn | 25.0 | 52.66 ± 11.49 |
| Winter | 14.5 | 104.25 ± 19.54 |
| Grouper | ||
| Spring | 23.0 | 48.05 ± 4.75 |
| Summer | 25.9 | 32.21 ± 3.01 |
| Autumn | 25.9 | 59.91 ± 6.99 |
| Winter | 16.6 | 24.15 ± 3.40 |
| Parameter | Temperature Rise Rate (°C/h) | |||
|---|---|---|---|---|
| +2 | +4 | +6 | +8 | |
| Spotted sea bass | ||||
| Critical thermal maximum (°C) | 35.57 ± 0.25 | 35.43 ± 0.31 | 35.40 ± 0.17 | 35.37 ± 0.21 |
| Initial death temperature (°C) | 37.23 ± 0.32 | 36.87 ± 0.15 | 36.80 ± 0.10 | 36.87 ± 0.12 |
| Absolute lethal temperature (°C) | 37.67 ± 0.12 | 37.97 ± 0.12 | 37.80 ± 0.10 | 37.47 ± 0.21 |
| Semi-lethal temperature (°C) a | 37.30 | 37.48 | 37.37 | 37.28 |
| Fit coefficient (R2) a | 0.768 | 0.787 | 0.762 | 0.948 |
| Semi-lethal temperature (°C) b | 37.24 | 37.24 | 37.14 | 37.16 |
| Grouper | ||||
| Critical thermal maximum (°C) | 37.17 ± 0.06 | 37.13 ± 0.34 | 36.23 ± 0.06 | 36.57 ± 0.32 |
| Initial death temperature (°C) | 38.70 ± 0.10 | 38.73 ± 0.12 | 38.67 ± 0.31 | 38.43 ± 0.40 |
| Absolute lethal temperature (°C) | 39.83 ± 0.12 | 39.53 ± 0.23 | 39.83 ± 0.55 | 39.47 ± 0.31 |
| Semi-lethal temperature (°C) a | 39.33 | 39.24 | 39.24 | 39.28 |
| Fit coefficient (R2) a | 0.781 | 0.745 | 0.587 | 0.923 |
| Semi-lethal temperature (°C) b | 39.10 | 39.03 | 38.97 | 39.15 |
| Parameter | Temperature Rise Rate (°C/h) | |||
|---|---|---|---|---|
| +2 | +4 | +6 | +8 | |
| Spotted sea bass | ||||
| Critical thermal maximum (°C) | 39.80 ± 0.20 | 39.80 ± 0.20 | 39.60 ± 0.20 | 39.40 ± 0.20 |
| Initial death temperature (°C) | 39.83 ± 0.21 | 39.97 ± 0.06 | 39.83 ± 0.15 | 39.50 ± 0.26 |
| Absolute lethal temperature (°C) | 40.17 ± 0.32 | 40.93 ± 0.32 | 40.50 ± 0.17 | 40.73 ± 0.12 |
| Semi-lethal temperature (°C) | 40.01 | 40.54 | 40.22 | 40.24 |
| Fit coefficient (R2) | 0.515 | 0.924 | 0.622 | 0.965 |
| Semi-lethal temperature (°C) | 39.97 | 40.44 | 40.02 | 40.11 |
| Grouper | ||||
| Critical thermal maximum (°C) | 36.10 ± 0.20 | 36.27 ± 0.45 | 37.30 ± 0.17 | 36.20 ± 0.10 |
| Initial death temperature (°C) | 40.13 ± 0.15 | 40.77 ± 0.32 | 40.63 ± 0.06 | 40.40 ± 0.35 |
| Absolute lethal temperature (°C) | 40.93 ± 0.12 | 42.10 ± 0.17 | 41.40 ± 0.20 | 42.13 ± 0.38 |
| Semi-lethal temperature (°C) | 40.55 | 41.42 | 40.94 | 41.46 |
| Fit coefficient (R2) | 0.746 | 0.895 | 0.680 | 0.837 |
| Semi-lethal temperature (°C) | 40.40 | 41.36 | 40.83 | 41.10 |
| Parameter | Temperature Rise Rate (°C/h) | |||
|---|---|---|---|---|
| +2 | +4 | +6 | +8 | |
| Spotted sea bass | ||||
| Critical thermal maximum (°C) | 37.17 ± 0.15 | 37.17 ± 0.15 | 37.23 ± 0.12 | 36.97 ± 0.42 |
| Initial death temperature (°C) | 37.53 ± 0.15 | 37.43 ± 0.21 | 37.67 ± 0.06 | 37.57 ± 0.42 |
| Absolute lethal temperature (°C) | 39.20 ± 0.36 | 39.03 ± 0.15 | 39.23 ± 0.15 | 39.27 ± 0.06 |
| Semi-lethal temperature (°C) | 38.30 | 38.37 | 38.46 | 40.24 |
| Fit coefficient (R2) | 0.805 | 0.767 | 0.917 | 0.965 |
| Semi-lethal temperature (°C) | 38.10 | 37.93 | 38.38 | 40.11 |
| Grouper | ||||
| Critical thermal maximum (°C) | 37.33 ± 0.25 | 37.53 ± 0.21 | 37.70 ± 0.26 | 36.90 ± 0.50 |
| Initial death temperature (°C) | 39.40 ± 0.72 | 39.70 ± 0.00 | 39.53 ± 0.55 | 39.47 ± 0.58 |
| Absolute lethal temperature (°C) | 40.40 ± 0.10 | 40.67 ± 0.06 | 41.00 ± 0.10 | 40.40 ± 0.26 |
| Semi-lethal temperature (°C) | 40.11 | 40.41 | 40.64 | 40.13 |
| Fit coefficient (R2) | 0.682 | 0.854 | 0.901 | 0.914 |
| Semi-lethal temperature (°C) | 40.06 | 40.14 | 40.54 | 40.06 |
| Parameter | Temperature Rise Rate (°C/h) | |||
|---|---|---|---|---|
| +2 | +4 | +6 | +8 | |
| Spotted sea bass | ||||
| Critical thermal maximum (°C) | 33.90 ± 0.20 | 33.83 ± 0.06 | 33.60 ± 0.44 | 33.90 ± 0.10 |
| Initial death temperature (°C) | 35.27 ± 0.15 | 35.23 ± 0.06 | 35.33 ± 0.06 | 34.93 ± 0.12 |
| Absolute lethal temperature (°C) | 36.10 ± 0.00 | 35.90 ± 0.52 | 36.23 ± 0.42 | 35.63 ± 0.06 |
| Semi-lethal temperature (°C) | 35.89 | 36.14 | 35.78 | 35.38 |
| Fit coefficient (R2) | 0.822 | 0.913 | 0.838 | 0.856 |
| Semi-lethal temperature (°C) | 35.73 | 35.89 | 35.73 | 35.33 |
| Grouper | ||||
| Critical thermal maximum (°C) | 35.77 ± 0.25 | 35.37 ± 0.45 | 34.70 ± 0.66 | 35.13 ± 0.45 |
| Initial death temperature (°C) | 38.20 ± 0.36 | 38.30 ± 0.52 | 36.93 ± 0.50 | 37.20 ± 0.10 |
| Absolute lethal temperature (°C) | 39.23 ± 0.32 | 38.87 ± 0.06 | 38.07 ± 0.06 | 38.03 ± 0.21 |
| Semi-lethal temperature (°C) | 38.80 | 38.52 | 37.86 | 37.67 |
| Fit coefficient (R2) | 0.816 | 0.671 | 0.853 | 0.895 |
| Semi-lethal temperature (°C) | 38.71 | 38.58 | 37.82 | 37.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.; Huang, Y.; Zhang, K.; Bo, J.; Zheng, R.; Ye, Y.; Liu, K. Thermal Tolerance of Spotted Sea Bass (Lateolabrax maculatus) and Pearl Gentian Grouper (Epinephelus fuscoguttatus Female × E. lanceolatus Male) in Different Environmental Temperatures. Fishes 2023, 8, 576. https://doi.org/10.3390/fishes8120576
Lin H, Huang Y, Zhang K, Bo J, Zheng R, Ye Y, Liu K. Thermal Tolerance of Spotted Sea Bass (Lateolabrax maculatus) and Pearl Gentian Grouper (Epinephelus fuscoguttatus Female × E. lanceolatus Male) in Different Environmental Temperatures. Fishes. 2023; 8(12):576. https://doi.org/10.3390/fishes8120576
Chicago/Turabian StyleLin, Heshan, Yaqin Huang, Kun Zhang, Jun Bo, Ronghui Zheng, Youling Ye, and Kun Liu. 2023. "Thermal Tolerance of Spotted Sea Bass (Lateolabrax maculatus) and Pearl Gentian Grouper (Epinephelus fuscoguttatus Female × E. lanceolatus Male) in Different Environmental Temperatures" Fishes 8, no. 12: 576. https://doi.org/10.3390/fishes8120576
APA StyleLin, H., Huang, Y., Zhang, K., Bo, J., Zheng, R., Ye, Y., & Liu, K. (2023). Thermal Tolerance of Spotted Sea Bass (Lateolabrax maculatus) and Pearl Gentian Grouper (Epinephelus fuscoguttatus Female × E. lanceolatus Male) in Different Environmental Temperatures. Fishes, 8(12), 576. https://doi.org/10.3390/fishes8120576