Abstract
Intelectins (ITLNs) are a family of calcium-dependent lectins with carbohydrate-binding capacity, are distributed across various vertebrates, and play an important role in the innate immune response against pathogen infection. The yellow catfish Pelteobagrus fulvidraco (Siluriformes: Bagridae) is an economically important fish in China. The aim of this study was to quantify the gene expression of ITLN in response to pathogen-associated molecular patterns (PAMPs) stimulation. Here, the ITLN gene of P. fulvidraco was characterized and named PfITLN. The full-length cDNA of PfITLN was 1132 bp, including a 5’-untranslated region (UTR) of 140 bp, a 3’-UTR of 110 bp, and an open reading frame (ORF) of 882 bp encoding a polypeptide of 293 amino acids, which contains a signal peptide and two fibrinogen-related domains (FReDs). PfITLN had a molecular weight of 32.39 kDa with a theoretical pI of 5.03. The deduced PfITLN amino acid sequence had 81%, 64%, and 55% homology with Ictalurus furcatus, Danio rerio, and Homo sapiens, respectively. Moreover, the predicted tertiary protein structure of PfITLN was highly similar to that of other animals, and phylogenetic analysis showed that the PfITLN protein was close to those of other Teleostei. Real-time quantitative reverse transcription-PCR (qRT-PCR) analysis showed PfITLN expression in all examined tissues, with the highest abundance seen in the liver, followed by the head kidney, spleen, trunk kidney, and muscle. After PAMP infection with lipopolysaccharide (LPS) and polyriboinosinic polyribocytidylic acid (poly I:C), the expression levels of PfITLN were significantly upregulated at different time points. These results suggested that PfITLN might be involved in innate immunity.
Key Contribution:
An intelectin (ITLN) gene was cloned and detected in the liver of the yellow catfish Pelteobagrus fulvidraco. The expression of P. fulvidraco ITLN mRNA levels was quantified in a wide range of tissues and was upregulated when challenged with poly I:C or LPS.
1. Introduction
The immune system of fish expresses both innate and acquired immunity [1]. Innate immunity is an important line of defense against bacteria, viruses, fungi, and parasites, which is mediated by pattern recognition receptors (PRRs) and can activate the adaptive immune response. Fish have less diverse adaptive immunity compared to mammals, thereby making innate immunity more vital for fish. Scientists have only discovered several immunoglobulin isotypes and confirmed that fish do not possess a major histocompatibility complex [2]. Groups of related microorganisms have unique microbial molecules, also known as pathogen-associated molecular patterns (PAMPs), which can be recognized by host PRRs. The primary components of microbes’ cell walls are often carbohydrate chains such as LPS (lipopolysaccharides), PGN (peptidoglycan), LTA (lipoteichoic acids), and β-glucan; these carbohydrate chains are commonly found in microbial molecules [3]. LPS, a major cell wall component of the outer membrane of Gram-negative bacteria, plays a key role in host–pathogen interactions with the innate immune system. Polyinosinic-polycytidylic acid (poly I:C) functions as a synthetic counterpart of double-stranded RNA (dsRNA), initiating the activation of innate immunity against infections by triggering specific PRRs such as Toll-like receptor 3 (TLR 3) and retinoic acid-inducible gene I (RIG-I)-like receptors, which consist of RIG-I and melanoma differentiation-associated gene 5. Various PRRs include PGRPs (peptidoglycan recognition proteins), TEPs (thioester-containing proteins), GNBPs (Gram-negative bacteria-binding proteins), SCRs (scavenger receptors), lectins, GALEs (galectins), TLRs (Toll-like receptors), RIG-like receptors (retinoic acid-inducible gene-like receptors), hemoglobin, and NOD-like receptors [4]. Lectins are an important component of the innate immune system for PRRs and include one or more carbohydrate recognition domains (CRDs) that can recognize and bind to a carbohydrate on the surface of bacteria, fungi, and viruses [5]. Based on their structure, binding specificity, and calcium dependency, lectins are divided into seven different families, including C-type lectins [6], F-type lectins [7], galectins [8], intelectins (ITLNs) [9], rhamnose-binding lectins [10], I-type lectins [11], and Lily-type lectins [12].
ITLNs are a newly recognized type of glycan-binding lectin involved in many physiological and pathological processes, including polyspermy [13], immune defense induced by pathogen infections [14], immune responses induced by parasites [15], asthma [16], iron metabolism [17], cancer, and the regulation of bone density [18]. ITLN was first isolated and cloned in Xenopus laevis oocytes and named X-lectin, and discovered a closely-related gene in the small intestinal tract of mice and gave it the name “intelectin” [19]. Subsequent ITLN homologs have been identified in many species, including mammals [20], amphibians [21], and fish [22,23], while avian genomes lack homology. Although the sequences of ITLNs remain consistent across various species, their expression patterns, quaternary structures, and functions vary significantly both within and between species [24]. The amino acid sequences of ITLNs in vertebrates exhibit a high level of conservation, encompassing the N-terminal fibrinogen-related domain (FReD) and the C-terminal intelectin domain [25]. As vertebrates, fish act as a bridge between innate and adaptive immunity and are considered an important model in comparative immunology studies [1]. Thus far, ITLNs have been identified and studied in many fish species, such as blunt snout bream (Megalobrama amblycephala) [26], channel catfish (Ictalurus punctatus) [27], rainbow trout (Oncorhynchus mykiss) [28], common carp (Cyprinus carpio) [29], and grass carp (Ctenopharyngodon idella) [30]. The crystal structure of human ITLN1 (hITLN1) and Xenopus embryonic epidermal lectin (XEEL) were reported, which provided novel insights into understanding their carbohydrate binding capacity and role in the innate immune response. Meanwhile, both XEEL and hITLN1 exhibit a high degree of similarity in both their natural forms and the way they bind ligands [24,25]. Several studies have confirmed that ITLNs play an important role in the immune response against pathogen infection and that expression levels are upregulated after pathogen challenge [31]. While the structure, expression pattern, and function of ITLNs are known in mammals and amphibians, information on the expression regulation and structure of ITLNs in P. fulvidraco is yet to be reported.
The yellow catfish Pelteobagrus fulvidraco is an important commercial freshwater aquaculture species in Asian countries, with a high market value in China. Recently, yellow catfish have been used as an experimental model for fish breeding [32], development [33], lipid metabolism [34], genomics [35], and toxicology [36]. Although infectious disease outbreaks associated with pathogenic microorganisms have caused high mortality rates and led to catastrophic economic losses in farmed P. fulvidraco, little is known about the immune system of this species. We previously screened a cDNA library of P. fulvidraco upon immune challenge [37,38,39,40] and identified an ITLN homolog that showed high similarity with other fish ITLNs. The full-length cDNA was obtained via rapid amplification of cDNA ends (RACE)-PCR for the investigation of expression patterns in different tissues and of the immune responses. The ITLNs protein sequences from various animals were comparatively analyzed and used to align and reconstruct their phylogenetic relationship. These data provide insights into the role of ITLN in yellow catfish.
2. Materials and Methods
2.1. Sample Collection
In May 2020, yellow catfish weighing 50 ± 10 g were collected from Yancheng, Jiangsu province, China, and were kept at 24 °C prior to experimentation. Thirteen tissues, including blood, brain, gill, head kidney, heart, intestine, liver, muscle, ovary, skin, spleen, testis, and trunk kidney were dissected to measure the expression pattern of PfITLN. Thirty fish were allocated across five PVC containers and kept at 24 °C. Five fish were randomly chosen and injected with either 100 μL of phosphate-buffered saline as control or lipopolysaccharides (LPS, L-2654, Sigma, St. Louis, MO, USA) or polyribose acid (poly I:C, P9582, Sigma St. Louis, MO, USA). After treatment, head kidney, blood, liver, and spleen tissues were quickly collected after 3, 6, 12, 24, 36, and 48 h and stored at −80 °C.
2.2. RNA Extraction and cDNA Synthesis
Total RNA was extracted using TRIzol reagent (Sangon, Shanghai, China) based on the manufacturer’s instructions. RNase-Free DNase I was used to remove contaminant genomic DNA (Promega, Madison, WI, USA). The purity and amount of extracted RNA were quantitatively measured using a NanoDrop 2000c spectrophotometer at OD260/OD280 (NanoDrop, Wilmington, DE, USA). Using the TRUEscript cDNA Synthesis Kit (Aidlab, Beijing, China) following the manufacturer’s instructions, the overall RNA concentration obtained from each sample was adjusted to 1 μg, and the first-strand cDNA was generated and stored at −20 °C for later use. The SMART™ RACE cDNA amplification kit (Clontech, Terra Bella, CA, USA) was used to incorporate single-stranded cDNAs into RACE-PCR.
2.3. Cloning PfITLN cDNA
Expressed sequence tags (EST) encoding an ITLN homolog were isolated from P. fulvidraco by random sequencing and transcriptome analysis [34,35,36,37]. Oligonucleotide primers were designed using the Primer 5.0 software (www.premierbiosoft.com/primerdesign/ (accessed on 15 October 2020)) (Table 1). Primers RC3 and RC5 were used for RACE-PCR to obtain full-length cDNA with 5 min at 95 °C, followed by 5 cycles at 95 °C for 60 s, 120 s at 65 °C, and then 35 cycles at 95 °C for 30 s, 30 s at 56 °C, and 45 s at 72 °C. PCR products were examined using 1% agarose gel. After purification, PCR products were linked to the T vector (Sangon, China) and sequenced.

Table 1.
Primers used.
2.4. Sequence Analysis of PfITLN
A BLAST search of Genbank (http://blast.ncbi.nlm.nih.gov/blast.cgi (accessed on 5 January 2021)) was conducted using the DNASTAR Lasergene 11 (Madison, WI, USA) to distinguish the open reading frame (ORF) of PfITLN and the amino acid sequence. The Expert Protein Analysis System (ExPAsy) was used to predict the isoelectric point (pI) and molecular weight (MW) of the derived amino acid sequence using the pI/MW tool (http://web.expasy.org/compute_pi/ (accessed on 7 March 2021)). To predict the protein signal peptide, an amino acid sequence was deduced using online SignalP server tools (http://www.cbs.dtu.dk/services/SignalP/ (accessed on 7 March 2021)). The SMART software (http://smart.embl-heidelberg.de/ (accessed on 7 March 2021)) was used to predict the functional domains. The topological structure of the transmembrane protein was investigated using the TMHMM online tool (http://www.cbs.dtu.dk/services/TMHMM (accessed on 7 March 2021)). The derived PfITLN amino acid sequence was submitted to the Swiss model protein folding server (https://swissmodel.expasy.org/ (accessed on 20 March 2021)) for automatic protein structure homology modeling.
2.5. Homologous Alignment and Phylogenetic Analysis
ITLN amino acid sequences from diverse species were downloaded from the GenBank database (www.ncbi.nlm.nih.gov/ (accessed on 12 July 2021)) for phylogenetic analysis. These ITLN sequences included Bubalus bubalis (XP_006060368), Bos taurus (CAO77313), Ovis aries (ABR23345), Homo sapiens (AAI17226), Pan paniscus (XP_008962500), Microcebus murinus (XP_012618482), Mus musculus (AAI50796), Cricetulus griseus (XP_007618315), Cavia porcellus (XP_013004555), Xenopus tropicalis (AAH61445), Salmo salar (XP_014001611), Astyanax mexicanus (XP_007238284), Danio rerio (XP_001335910), Silurus asotus (BAL14267), Ictalurus furcatus (ABW07848), Ictalurus punctatus (ABW07846), and P. fulvidraco. Sequence alignments were performed utilizing the Clustal X software (https://evomics.org/resources/software/bioinformatics-software/clustal-x/ (accessed on 16 August 2021)) [41]. The amino acid and nucleotide sequence alignment were performed, and amino acid sequences of the ITLN gene were used to reconstruct the phylogenetic relationships based on the neighbor-joining method. The phylogenetic tree was constructed using the molecular evolutionary genetics analysis software MEGA 6.0 [42]. The data were analyzed using the Poisson modification, and gaps were eliminated by absolute deletion. The topological stability of adjacent trees was estimated using 1000 bootstrap replicates.
2.6. Quantitative Analysis of PfITLN
qRT-PCR was utilized to investigate the level of mRNA expression of the PfITLN gene after immune stimulation in multiple tissues and PAMPs. The expression level of the actin gene was utilized as a housekeeping control. The primers used for qRT-PCR are shown in Table 1. qRT-PCR was performed using the Mastercycler Ep Realplex thermocycler (Eppendorf, Hamburg, Germany), utilizing the SYBR Green qPCR Mix kit (Aidlab, Beijing, China). The reaction mixture (20 μL) included 10 μL of 2 × SYBR Green qPCR Mix, 1 μL of forward and reverse primers, 1 μL of cDNA, and 7 μL of RNase-free H2O. The PCR protocol included 40 cycles of 50 s at 95 °C, 15 s at 55 °C, and 30 s at 72 °C. The melting point was identified as between 60 °C and 95 °C. Each individual experiment was performed three times, and the relative gene expression was measured using the methods of Livak and Schmittgen [43].
2.7. Data Analysis
Data are shown as mean ± standard error of the mean (SEM). One-way ANOVA tests and a p-value < 0.05 were used to identify significant differences.
3. Results and Discussion
3.1. Sequence Analysis of the PfITLN Gene
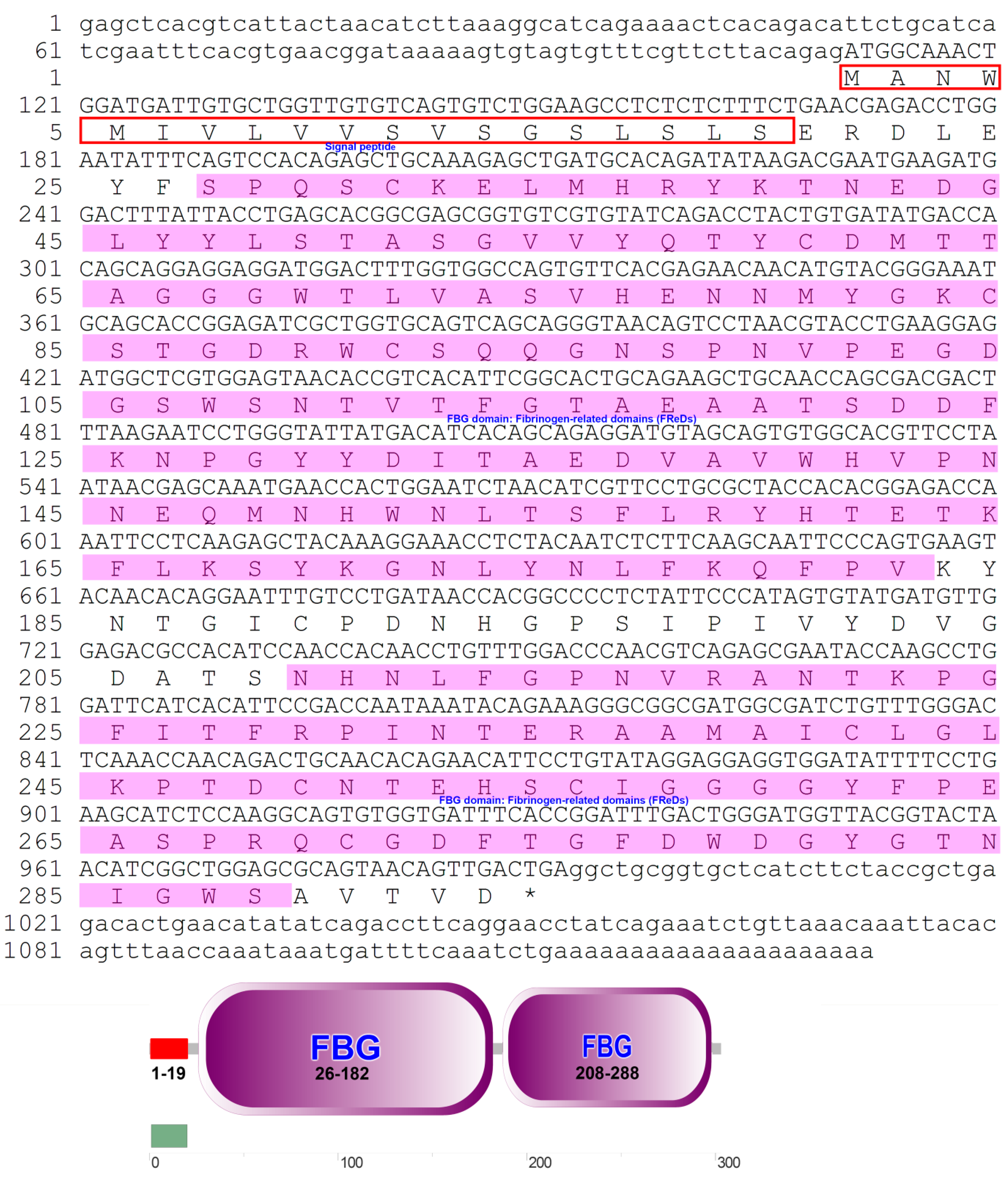
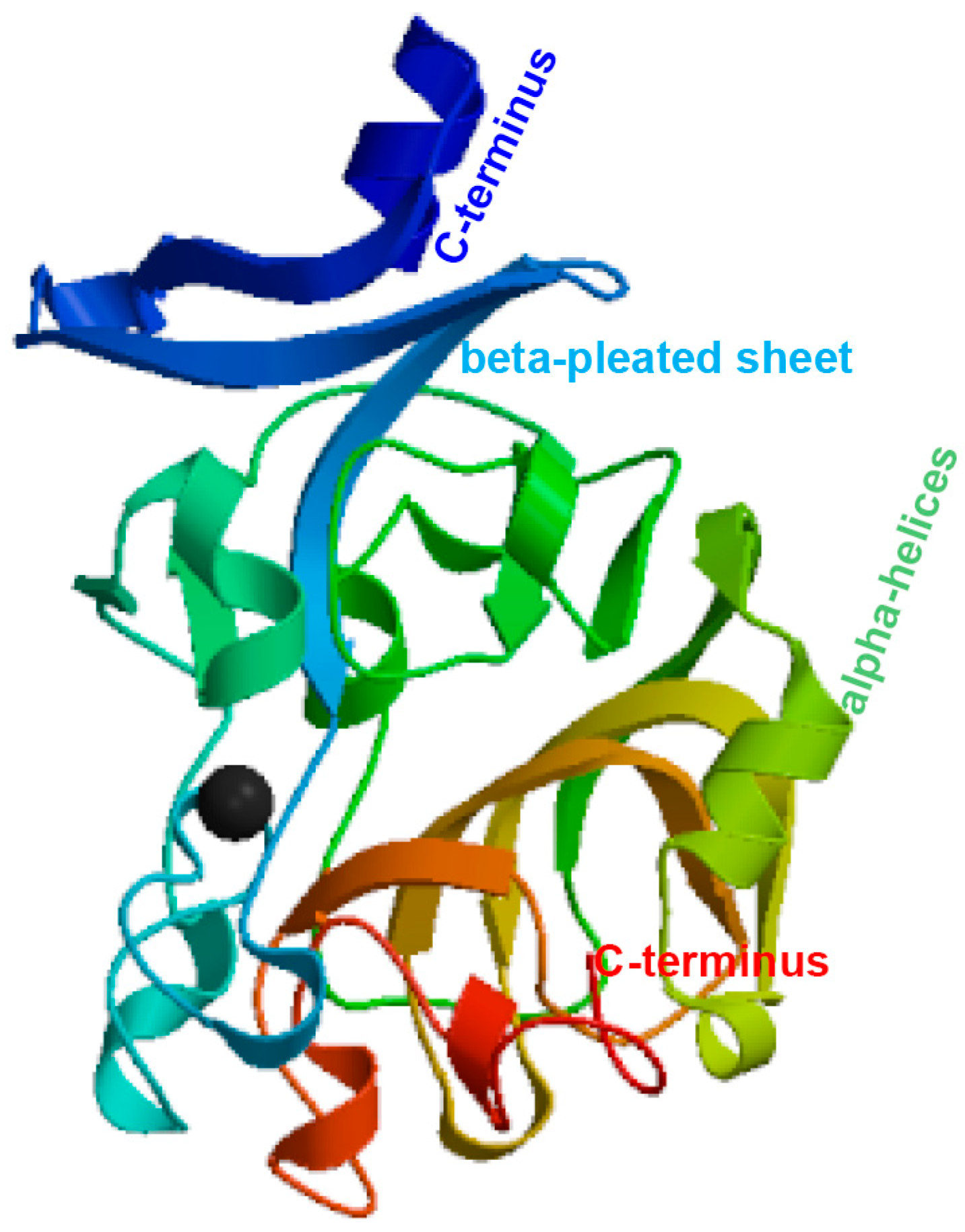
The ITLN gene was identified using RNA extracted from the liver of P. fulvidraco. Full-length cDNA from PfITLN was obtained via RT–PCR and RACE–PCR. The resulting 1132-bp cDNA sequence included a 140-bp 5′-untranslated sequence, a putative ORF of 882-bp encoding a polypeptide of 293 amino acids with two typical structural features, fibrinogen-related domains, and a 110-bp 3′ untranslated area with a 21-bp poly (A) tail. The nucleotide and amino acid sequences of PfITLN are displayed in Figure 1. Based on the amino acid sequence, the molecular weight of PfITLN was predicted to be 32.39 kDa, and the equipotential point was 5.03. A protein signal peptide forecast protocol within the SMART software (Heidelberg, Germany) identified a signaling peptide, suggesting that this is a secreted protein. Motif-scan results showed that the PfITLN protein contained a N-glycosylation site, nine casein kinase II phosphorylation sites, six protein kinase C phosphorylation sites, seven N-myristoylation sites, and a tyrosine kinase phosphorylation site. As displayed in Figure 2, the forecasted tertiary structure of PfITLN protein by the Swiss model protein folding server included alpha helices, a beta-sheet, a C-terminus, and a N-terminus, indicating that ITLN has a conserved region and similar function as in human [44]. By utilizing the conventional area forecast of the SMART software, PfITLN contains two fibrinogen-related domains, which play vital roles in blood clotting, platelet aggregation, and regulation of immune activity [25,45]. This result also revealed that this protein belonged to the lectin superfamily, a diverse family of proteins involved in activity towards N-acetylglucosamine via their fibrogen-like domains that are needed in a variety of cellular processes, including blood clotting, immune response, and the regulation of neurogenesis.
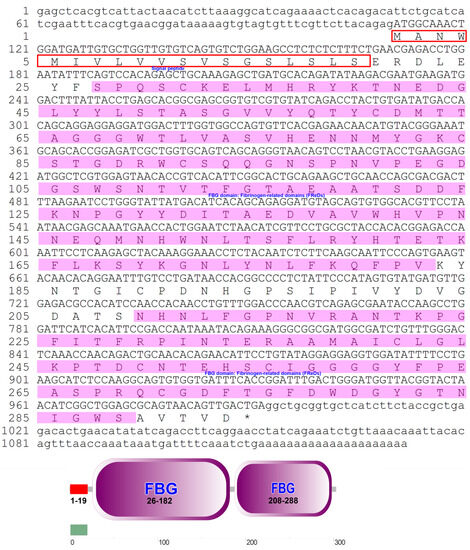
Figure 1.
Complete nucleotide and derived amino acid sequences of the ITLN gene of P. fulvidraco. Amino acid residues are represented as letters. The open reading frame (ORF) from the start codon ATG to the stop codon TAA is capitalized. Red boxes represent the signal peptide, and pink boxes represent the fibrinogen-related domains.
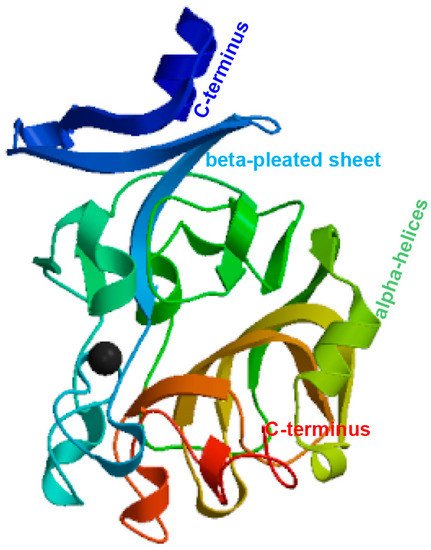
Figure 2.
Predicted tertiary structure of the PfITLN protein.
3.2. Homologous Sequence Alignment and Phylogenetic Analysis
As shown in Figure 3, nine ITLN protein sequences were compared using the Clustal X software (Ballwin, MO, USA) to evaluate the evolutionary affinity of ITLN. The inferred PfITLN protein sequence was strongly consistent with the evolutionary position of the organism, with 81%, 64%, and 55% homology with Ictalurus furcatus, Danio rerio, and Homo sapiens, respectively. The amino acid sequence homology was over 50%, indicating that ITLN is strongly conserved during evolution [42]. Additionally, sequence alignments and functional domain predictions showed that amino acid sequences for the conserved regions of ITLN were highly similar to those in animal ITLN.

Figure 3.
Sequence alignment of the PfITLN protein with its homologs. The ITLN sequences from H. sapiens (AAI17226), M. musculus (AAI50796), Cavia porcellus (XP_013004555), X. tropicalis (AAH61445), A. mexicanus (XP_007238284), D. rerio (XP_001335910), S. asotus (BAL14267), I. furcatus (ABW07848), I. punctatus (ABW07846), and P. fulvidraco are aligned by Clustal X software. Conserved amino acids are highlighted in black, and similar amino acids in grey. Purple and red boxes represent the fibrinogen-related domains, and green boxes represent the signal peptide.
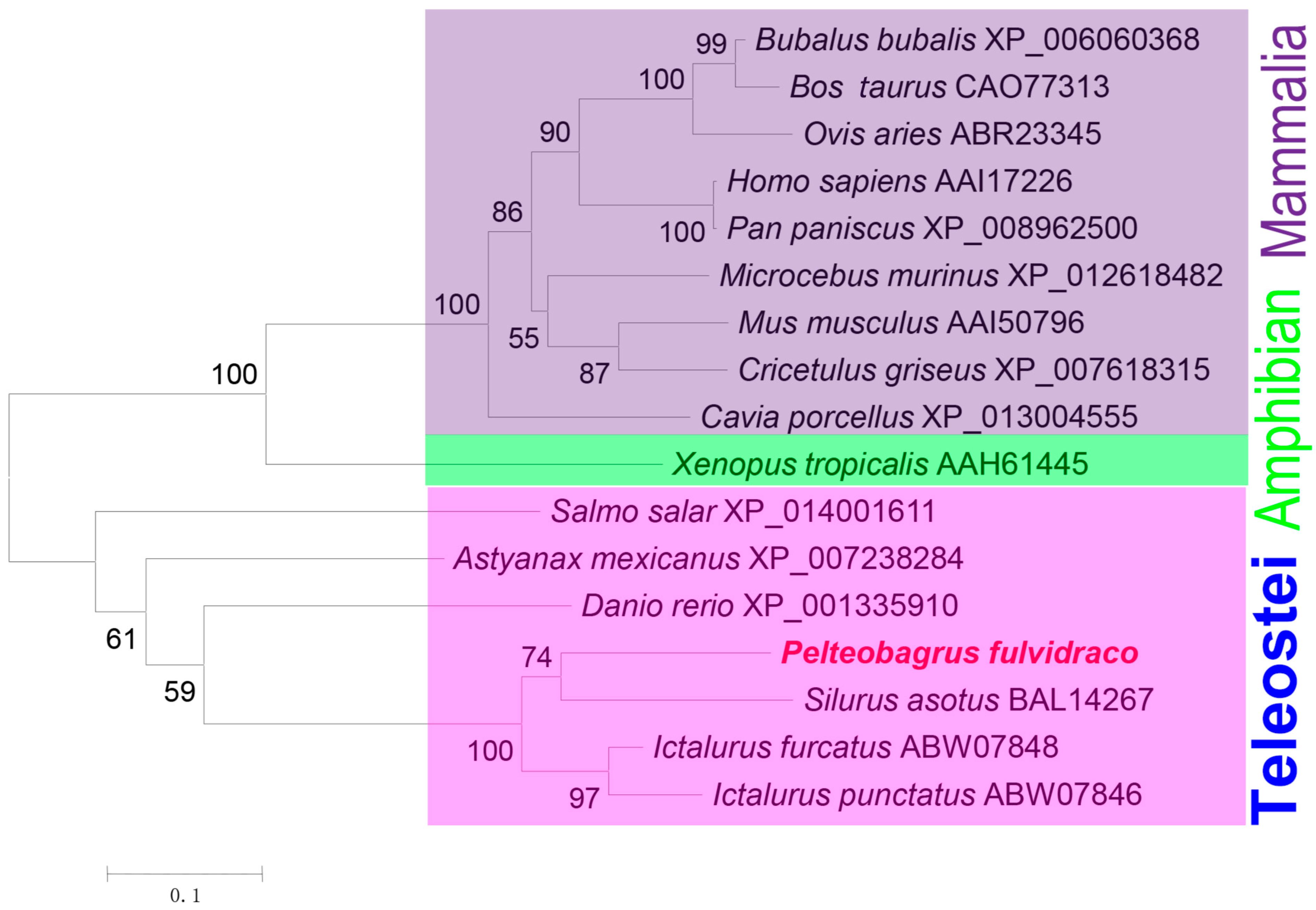
We subsequently aimed to identify the correlation between configurational elements and the immune activity of ITLN. To categorize and investigate the molecular evolution of ITLN, phylogenetic relationships of 17 typical ITLN sequences were reconstructed using a neighbor-joining (NJ) approach based on the amino acid sequences. As shown in Figure 4, a total of 30 representative ITLN amino acid sequences from different organisms including P. fulvidraco were used to reconstruct the phylogenetic relationship. Results showed that the sequences analyzed could be grouped into four distinct categories, including mammalian, amphibian, and bony fish clades. All bony fish ITLNs were closely related within one major clade, while PfITLN was most closely related to that of Silurus asotus. In general, the phylogenetic tree analysis corresponded with their phylogenetic affiliations and supported the recognition of the PfITLN protein, as it has substantial homology with other phylogenetic clusters [46].

Figure 4.
MEGA phylogenetic tree established via the NJ algorithm. Public database accession numbers of ITLN proteins are shown following the names of organisms. The bootstrap values of each branch (1000 repeats) are indicated, and the number on each node corresponds to the value.
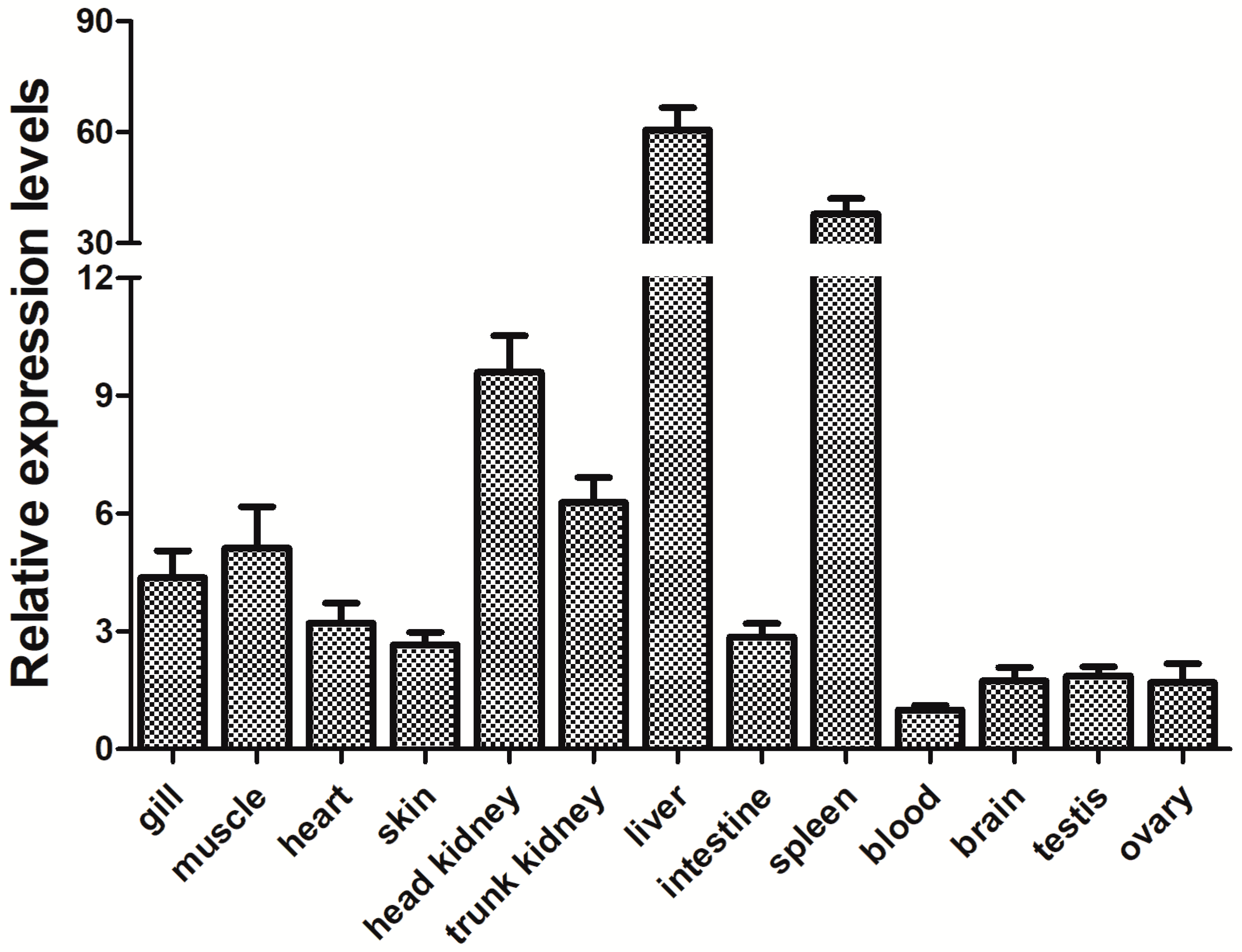
3.3. Expression of the PfITLN Gene in Yellow Catfish
The relative level of expression of PfITLN across tissues was measured using qRT-PCR. The level of expression for each tissue was standardized to that of actin. The qRT-PCR results are shown in Figure 5 and indicate that the PfITLN is expressed in all investigated tissues, suggesting that the ITLN gene may play an important role in the development of yellow catfish. Namely, the lowest expression of PfITLN was detected in the blood, while the highest expression was in the liver, spleen, head kidney, trunk kidney, and muscle. The pattern of tissue expression of ITLN was consistent with that observed in previous studies, which also reported high expression levels in the liver, spleen, head, kidney, and intestine. Transcript levels of ITLN from rainbow trout were found in the sputum, liver, intestine, and skin [28]. The common carp ITLN gene was detected in all tissues and at high levels in the hindgut, midgut, and spleen [29]. ITLNs from Ctenopharyngodon idella were primarily detected in the head, kidney, and spleen, as well as the intestine and gill [30]. The ITLN expression of Megalobrama amblycephala was primarily detected in the liver and spleen but also in the heart, kidney, muscle, blood, brain, intestine, and gills [47]. ITLN2 expression was generally detected in zebrafish, with the highest level detected in the intestine and base levels in the liver [48]. In the channel and blue catfishes, the highest expression of ITLN2 was observed in the liver, and lower levels were seen in other tissues [49]. ITLN was mainly expressed in the spleen, liver, and kidney of Carassius auratus gibelio [50]. The ITLN gene was found to be expressed in various tissues, including the blood, intestines, kidney, heart, gill, liver, adipose tissue, and gonads of the lamprey, Lampetra japonica [51]. AmphiITLN71469 showed a high level of expression in the digestive tract and skin in amphioxus [52]. AmphiITLN239631 is expressed in the muscle, epidermis, sputum, hepatic caecum, intestinal tract, and testis, with the hepatic cecum exhibiting the highest expression and the muscle displaying the lowest expression [52]. Human ITLN1 showed a wider range of expression, including heart, small intestine, colon, kidney collecting tubule cells, bladder umbrella cells, some mesothelial cells, and follicular cells, present in both the small intestine and colon, particularly in goblet cells, with no changes in its expression observed in Crohn’s disease (CD); human ITLN2 was selectively expressed in Paneth cells of the small intestine [53,54]. Six ITLNs from the 129S7 mouse strain were identified with site-specific expressions in the gastrointestinal tract [55]. Taken together, ITLN was primarily observed in immunologically imported tissues, including the spleen, head, kidney, and liver, which suggests that ITLN may play an essential role in preventing microbial infection in aquatic conditions [25].
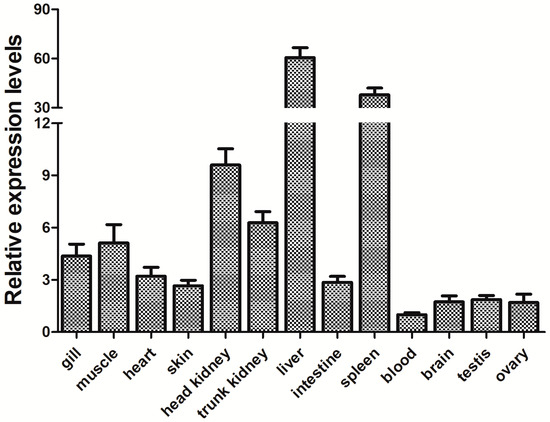
Figure 5.
Relative expression of the PfITLN gene in different tissues. The actin gene was utilized as an internal control. The lowest gene expression level was set as 1.0. Data are shown as average fold-change relative to the untreated control (mean ± SE, n = 3).
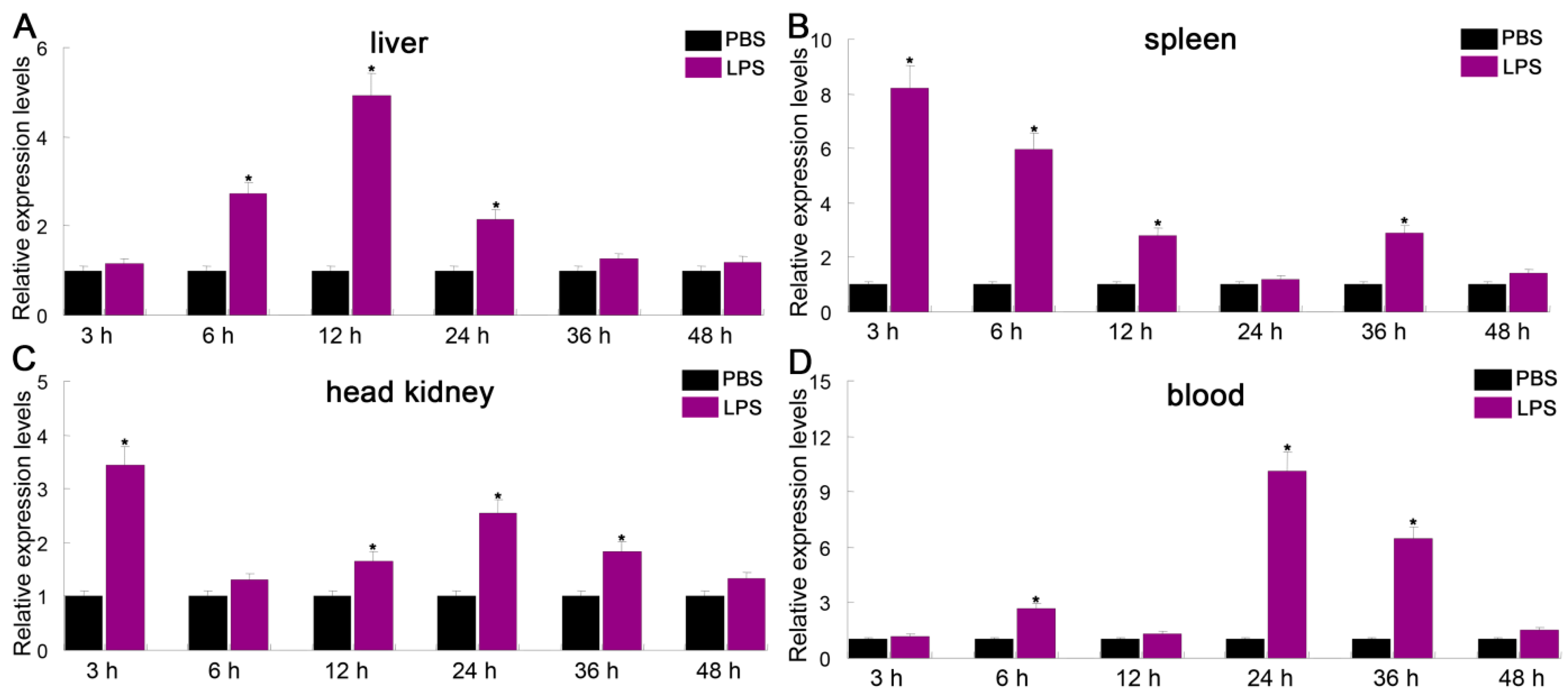
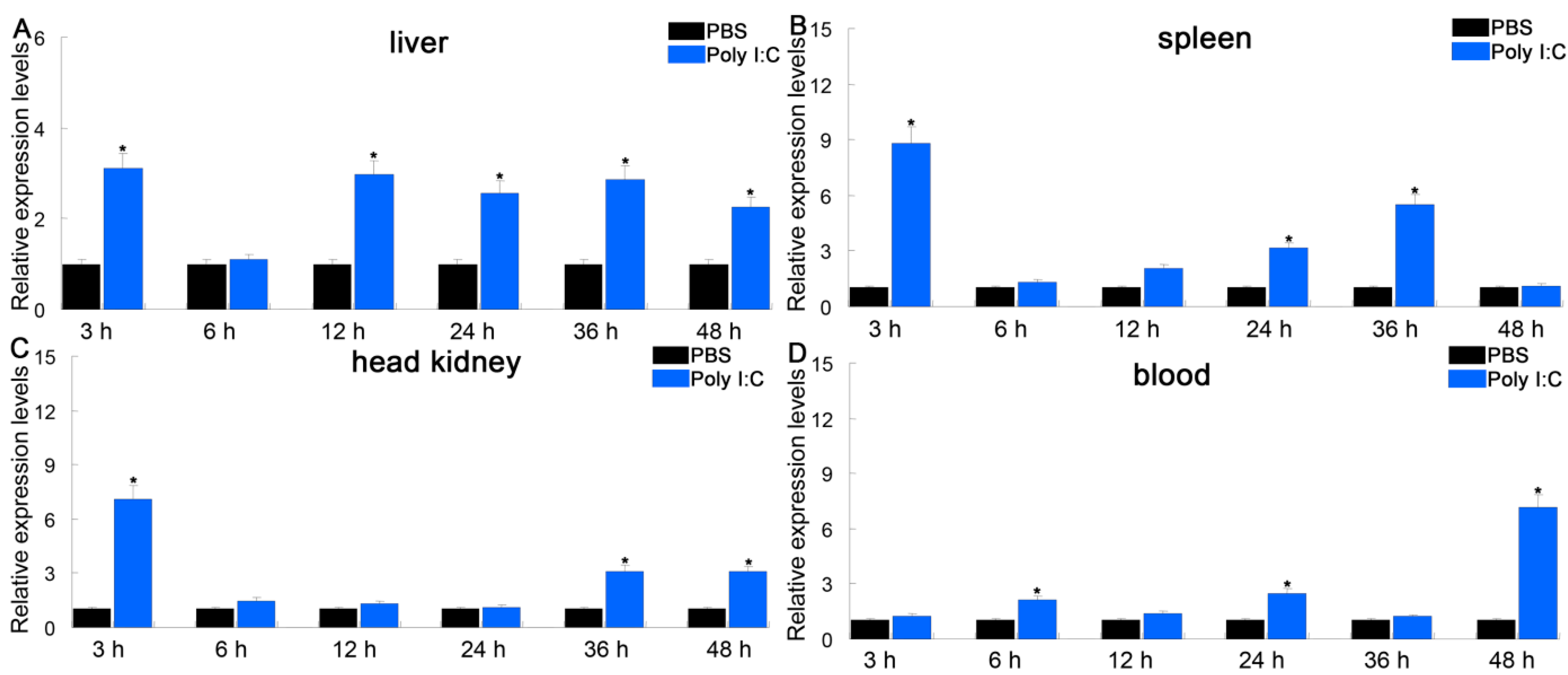
3.4. Expression Profiles of PfITLN Challenged by LPS or Poly I:C
Intelectin is a lectin with the capacity to recognize and bind to carbohydrates [25]. ITLN genes play a critical role in innate immune responses to microbial infections in fish. To investigate the immune responses of PfITLN, the expression of PfITLN was investigated in immunological tissues upon antigen exposure utilizing the qRT-PCR method. Tissues were collected at the critical time points for the presentation and attachment of PAMPs (LPS and Poly I:C). As shown in Figure 6 and Figure 7, the expression of PfITLN significantly upregulated in the spleen and head kidney after 3 h using LPS. Following the LPS challenge, the expression of PfITLN peaked at 3 h in the head kidney, at 12 h in the liver, and at 24 h in the blood, respectively. Following the poly I:C challenge, the expression of PfITLN was significantly upregulated after 3 h in the liver, spleen, and head kidney and peaked at 48 h in the blood. PfITLN expression was upregulated in other fish following LPS challenge or infection with bacteria. The spleen enables host cells to destroy invasive pathogens [56]. The expression of ITLN in grass carp Ctenopharyngodon idella was increased after LPS injection [30]. The mRNA transcript and protein levels of ITLN in M. amblycephala were dramatically upregulated in immune-related tissues at 24 h in response to the Aeromonas hydrophila challenge [47]. In Danio rerio, ITLN2 caused bacterial agglutination and bound to the LPS or PGN of bacteria [48]. The ITLN expression levels of channel catfish were induced in macrophage-rich tissues injected with Edwardsiella ictaluri [49]. In Branchiostoma japonicum, ITLN expression was upregulated in the intestine 8 h after the Staphylococcus aureus challenge [57]. The ITLN mRNA transcript levels of rainbow trout were upregulated in response to Listonella anguillarum [58]. After infection with Trichuris muris, the small intestine of Mus musculus showed an increase in intelectin-2 expression [59]. The expression of ITLN3 was significantly increased in all tissues of common carp after being infected with S. aureus or A. hydrophila [60]. Human ITLN-1 was a host defense lectin that assisted phagocytic clearance of microorganisms and possessed the ability to bind exocyclic 1,2-diols found in the surface glycans of human pathogens such as Streptococcus pneumoniae, Vibrio cholerae, Mycobacterium bovis and Helicobacter pylori in its secreted glycoprotein form [61,62]. The co-purification of sheep intelectin-2 with mucin Muc5ac from gastric mucus suggests that intelectin might have a part in altering the consistency of mucus [63]. The expression of ITLN3 in X. laevis was upregulated in the intestinal and rectal [64]. These results suggest that ITLN is an essential protein that plays a role in the immune response of fish.
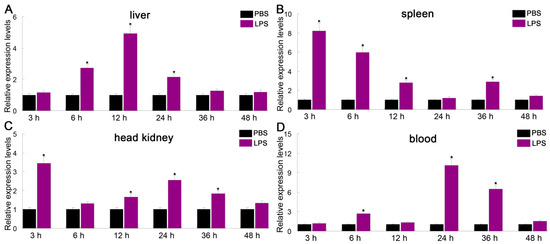
Figure 6.
Relative expression of PfITLN in response to LPS in different tissues. The actin gene was utilized as an internal control. Data are shown as average fold change relative to the untreated control (mean ± SE, n = 3). It was considered statistically significant when p-values were less than 0.05 and indicated by an asterisk (*).
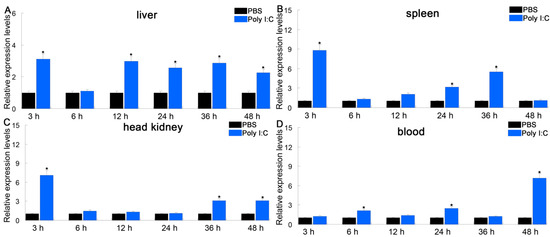
Figure 7.
Relative expression of PfITLN in response to poly I:C in different tissues. The actin gene was utilized as an internal control. Data are shown as average fold-change relative to the untreated control (mean ± SE, n = 3). It was considered statistically significant when p-values were less than 0.05 and indicated by an asterisk (*).
4. Conclusions
Innate immunity is the primary defense mechanism in fish. ITLN, which act as pattern recognition receptors, plays a key role in the initial defense against pathogens. The ITLN gene from P. fulvidraco was identified and characterized, and the levels of expression were examined under the PAMPs challenge. Our results suggest the PfITLN may play a key role in mediating an innate immune response to PAMP exposure. Gene expression data can offer insights into the innate immune defense mechanisms and disease management and aid the development of molecular markers for disease resistance. However, further functional research should be conducted to better describe the effectiveness of using ITLNs to improve disease resistance.
Author Contributions
Conceptualization, Q.L. and B.T.; Data curation, Q.L.; Formal analysis, S.J., Y.L. (Yuting Lei) and R.M.; Funding acquisition, S.J., Q.L. and B.T.; Investigation, S.J., Y.L. (Yuting Lei), W.S., Q.L. and B.T.; Methodology, S.J. and Q.L.; Project administration, Q.L. and B.T.; Resources, Q.L.; Software, Q.L.; Supervision, S.J., Q.L. and B.T.; Validation, Y.L. (Yanxuan Li), W.S. and T.W.; Writing—original draft, S.J. and Q.L.; Writing—review and editing, Q.L. and B.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Research General Program of Jiangsu Provincial Higher Education Institutions (22KJA240002 and 21KJA240003); the Fishery High-Quality Development Research Project of Yancheng, China (YCSCYJ2021022); the Transverse Projects of Xiangshui Haichen Agriculture Development Company Limited (2022320907000829), the Science and Technology Vice-President Project of Jiangsu Province (FZ20230549); the National Natural Science Foundation of China (31440089, 32370556, and 32270487); the Natural Science Foundation of Jiangsu Province (BE2020673); and the National Key R&D Program of China (2019YFD0900404-05). The study was sponsored by the Qing Lan Project of Jiangsu Province and the “Outstanding Young Talents” of Yancheng Teachers University.
Institutional Review Board Statement
The Committee of the Yancheng Teachers University approved the animal protocols, and all experiments were performed under the applicable standards, with access No. YCTU-2020005.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhu, L.; Nie, L.; Zhu, G.; Xiang, L.X.; Shao, J.Z. Advances in research of fish immune-relevant genes: A comparative overview of innate and adaptive immunity in teleosts. Dev. Comp. Immunol. 2013, 39, 39–62. [Google Scholar] [CrossRef]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]
- Yuan, S.; Sun, Z.; Gao, Q.; Li, Z.; Qi, Z.; Zheng, S.; Liu, D. Molecular characterization and expression analysis of NLRC3-like, ASC, and caspase1 in spotted sea bass (Lateolabrax maculatus). Fishes 2023, 8, 378. [Google Scholar] [CrossRef]
- Guan, R.; Mariuzza, R.A. Peptidoglycan recognition proteins of the innate immune system. Trends Microbiol. 2007, 15, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Fai Cheung, R.C.; Wing Ng, C.C.; Fang, E.F.; Wong, J.H. A review of fish lectins. Curr. Protein Pept. Sci. 2015, 16, 337–351. [Google Scholar] [CrossRef]
- Tang, X.H.; Ye, X.T.; Wu, Q.C.; Tang, Y.Y.; Zhang, D.Z.; Liu, Q.N.; Tang, B.P.; Zhu, H.R. Molecular characterization and expression analysis of a novel C-type lectin (CTL) gene in yellow catfish Pelteobagrus fulvidraco. Aquacult. Rep. 2021, 20, 100640. [Google Scholar] [CrossRef]
- Vasta, G.R.; Amzel, L.M.; Bianchet, M.A.; Cammarata, M.; Feng, C.; Saito, K. F-type lectins: A highly diversified family of fucose-binding proteins with a unique sequence motif and structural fold, involved in self/non-self-recognition. Front. Immunol. 2017, 8, 1648. [Google Scholar] [CrossRef]
- Niu, J.; Huang, Y.; Liu, X.; Wu, F.; Tang, J.; Wang, B.; Lu, Y.; Cai, J.; Jian, J. Fish galectin8-like exerts positive regulation on immune response against bacterial infection. Front. Immunol. 2020, 11, 1140. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, Y.; Li, H.; Wang, X.; Zhang, M.; Shen, X.; Cheng, H.; Xu, J.; Wang, X.; Liu, H.; et al. Novel insights into the immune regulatory effects of Megalobrama amblycephala intelectin on the phagocytosis and killing activity of macrophages. Mol. Immunol. 2021, 137, 145–154. [Google Scholar] [CrossRef]
- Sugawara, S.; Tatsuta, T.; Hosono, M. Bacterial Expression of Rhamnose-Binding Lectin from Catfish Eggs. Methods Mol. Biol. 2020, 2132, 359–367. [Google Scholar]
- Angata, T.; Brinkman-Van der Linden, E. I-type lectins. Biochim. Biophys. Acta 2002, 1572, 294–316. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Wang, Z.; Zhang, Q. Immunological characterization and function analysis of L-type lectin from spotted knifejaw, Oplegnathus punctatus. Front. Immunol. 2022, 13, 993777. [Google Scholar] [CrossRef] [PubMed]
- Wyrick, R.E.; Nishihara, T.; Hedrick, J.L. Agglutination of jelly coat and cortical granule components and the block to polyspermy in the amphibian Xenopus laevis. Proc. Natl. Acad. Sci. USA 1974, 71, 2067–2071. [Google Scholar] [CrossRef] [PubMed]
- Blanco Vázquez, C.; Balseiro, A.; Alonso-Hearn, M.; Juste, R.A.; Iglesias, N.; Canive, M.; Casais, R. Bovine intelectin 2 expression as a biomarker of paratuberculosis disease progression. Animals (Basel) 2021, 11, 1370. [Google Scholar] [CrossRef]
- French, A.T.; Bethune, J.A.; Knight, P.A.; McNeilly, T.N.; Wattegedera, S.; Rhind, S.; Miller, H.R.P.; Pemberton, A.D. The expression of intelectin in sheep goblet cells and upregulation by interleukin-4. Vet. Immunol. Immunopathol. 2007, 120, 41–46. [Google Scholar] [CrossRef]
- Kuperman, D.; Lewis, C.; Woodruff, P.; Rodriguez, M.W.; Yang, Y.H.; Dolganov, G.M.; Fahy, J.V.; Erle, D.J. Dissecting asthma using focused transgenic modelling and functional genomics. J. Allergy Clin. Immunol. 2005, 116, 305–311. [Google Scholar] [CrossRef]
- Akiyama, Y.; Oshima, K.; Kuhara, T.; Shin, K.; Abe, F.; Iwatsuki, K.; Nadano, D.; Matsuda, T. A lactoferrin-receptor, intelectin 1, affects uptake, sub-cellular localization and release of immunochemically detectable lactoferrin by intestinal epithelial Caco-2 cells. J. Biochem. 2013, 154, 437–448. [Google Scholar] [CrossRef]
- Washimi, K.; Yokose, T.; Yamashita, M.; Kageyama, T.; Suzuki, K.; Yoshihara, M.; Miyagi, Y.; Hayashi, H.; Tsuji, S. Specific expression of human intelectin-1 in malignant pleural mesothelioma and gastrointestinal goblet cells. PLoS ONE 2012, 7, e39889. [Google Scholar] [CrossRef]
- Lee, J.K.; Buckhaults, P.; Wilkes, C.; Teilhet, M.; King, M.L.; Moremen, K.W.; Pierce, M. Cloning and expression of a Xenopus laevis oocyte lectin and characterization of its mRNA levels during early development. Glycobiology 1997, 7, 367–372. [Google Scholar] [CrossRef]
- Tsuji, S.; Uehori, J.; Matsumoto, M.; Suzuki, Y.; Matsuhisa, A.; Toyoshima, K.; Seya, T. Human intelectin is a novel soluble lectin that recognizes galactofuranose in carbohydrate chains of bacterial cell walls. J. Biol. Chem. 2001, 276, 23456–23463. [Google Scholar] [CrossRef]
- Nagata, S. Xenopus laevis macrophage-like cells produce XCL-1, an intelectin family serum lectin that recognizes bacteria. Immunol. Cell Biol. 2018, 96, 872–878. [Google Scholar] [CrossRef]
- Cui, H.; Liu, Y.; Zheng, Y.; Li, H.; Zhang, M.; Wang, X.; Zhao, X.; Cheng, H.; Xu, J.; Chen, X.; et al. Intelectin enhances the phagocytosis of macrophages via CDC42-WASF2-ARPC2 signalling axis in Megalobrama amblycephala. Int. J. Biol. Macromol. 2023, 236, 124027. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Yang, M.; Jiang, H.; Li, T.; Duan, G.; Ling, J.; Gao, Q. Effect of astragalus membranaceus on transcriptome and survival of hybrid yellow catfish (Pseudobagrus vachellii ♂ × Tachysurus fulvidraco ♀) in response to Aeromonas hydrophila challenge. Fishes 2023, 8, 454. [Google Scholar] [CrossRef]
- Wesener, D.A.; Dugan, A.; Kiessling, L.L. Recognition of microbial glycans by soluble human lectins. Curr. Opin. Struct. Biol. 2017, 44, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, J.; Yang, G. A comparative review of interactions. Scand J. Immunol. 2020, 92, e12882. [Google Scholar] [CrossRef]
- Ding, Z.; Zhao, X.; Wang, J.; Zhang, F.; Wang, W.; Liu, H. Intelectin mediated phagocytosis and killing activity of macrophages in blunt snout bream (Megalobrama amblycephala). Fish Shellfish Immunol. 2019, 87, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Singrang, N.; Sitthiyotha, T.; Chomanee, N.; Watthanasak, C.; Chunsrivirot, S.; Wangkanont, K. Molecular properties and ligand specificity of zebrafish intelectin-2. Fish Shellfish Immunol. 2022, 123, 528–536. [Google Scholar] [CrossRef]
- Russell, S.; Hayes, M.A.; Lumsden, J.S. Immunohistochemical localization of rainbow trout ladder lectin and intelectin in healthy and infected rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2009, 26, 154–163. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.; Gu, W.; Xu, F.; Li, H.; Shan, S.; Sun, X.; Yin, M.; Yang, G.; Chen, L. Characterization of a common carp intelectin gene with bacterial binding and agglutination activity. Fish Shellfish Immunol. 2021, 108, 32–41. [Google Scholar] [CrossRef]
- Chang, M.X.; Nie, P. Intelectin gene from the grass carp Ctenopharyngodon idella: cDNA cloning, tissue expression, and immunohistochemical localization. Fish Shellfish Immunol. 2007, 23, 128–140. [Google Scholar] [CrossRef]
- Meiers, J.; Siebs, E.; Zahorska, E.; Titz, A. Lectin antagonists in infection, immunity, and inflammation. Curr. Opin. Chem. Biol. 2019, 53, 51–67. [Google Scholar] [CrossRef]
- Liu, H.; Guan, B.; Xu, J.; Hou, C.; Tian, H.; Chen, H. Genetic manipulation of sex ratio for the large-scale breeding of YY super-male and XY all-male yellow catfish (Pelteobagrus fulvidraco (Richardson)). Mar. Biotechnol. 2013, 15, 321–328. [Google Scholar] [CrossRef]
- Gao, D.; Huang, J.; Lin, G.; Lu, J. Time-course transcriptome analysis of gonads from yellow catfish (Pelteobagrus fulvidraco) reveals genes associated with gonad development. BMC Genom. 2022, 23, 409. [Google Scholar] [CrossRef]
- Liu, G.H.; Zhang, D.G.; Lei, X.J.; Tan, X.Y.; Song, C.C.; Zheng, H.; Luo, Z. Effects of dietary selenium and oxidized fish oils on intestinal lipid metabolism and antioxidant responses of yellow catfish Pelteobagrus fulvidraco. Antioxidants 2022, 11, 1904. [Google Scholar] [CrossRef]
- Gong, G.; Dan, C.; Xiao, S.; Guo, W.; Huang, P.; Xiong, Y.; Wu, J.; He, Y.; Zhang, J.; Li, X.; et al. Chromosomal-level assembly of yellow catfish genome using third-generation DNA sequencing and Hi-C analysis. Gigascience 2018, 7, giy120. [Google Scholar] [CrossRef]
- Chen, Q.L.; Gong, Y.; Luo, Z.; Zheng, J.L.; Zhu, Q.L. Differential effect of waterborne cadmium exposure on lipid metabolism in liver and muscle of yellow catfish Pelteobagrus fulvidraco. Aquat. Toxicol. 2013, 142–143, 380–386. [Google Scholar] [CrossRef]
- Liu, Y.; Xin, Z.Z.; Zhang, D.Z.; Wang, Z.F.; Zhu, X.Y.; Tang, B.P.; Jiang, S.-H.; Zhang, H.-B.; Zhou, C.-L.; Chai, X.-Y.; et al. Transcriptome analysis of yellow catfish (Pelteobagrus fulvidraco) liver challenged with polyriboinosinic polyribocytidylic acid (poly I:C). Fish Shellfish Immunol. 2017, 68, 395–403. [Google Scholar] [CrossRef]
- Liu, Q.N.; Xin, Z.Z.; Liu, Y.; Zhang, D.Z.; Jiang, S.H.; Chai, X.Y.; Wang, Z.F.; Zhang, H.-B.; Bian, X.-G.; Zhou, C.-L.; et al. De novo transcriptome assembly and analysis of differential gene expression following lipopolysaccharide challenge in Pelteobagrus fulvidraco. Fish Shellfish Immunol. 2018, 73, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.H.; Wu, L.X.; Cai, Y.T.; Ma, R.T.; Zhang, H.B.; Zhang, D.Z.; Tang, B.P.; Liu, Q.N.; Dai, L.S. Differentially expressed genes in head kidney of Pelteobagrus fulvidraco following Vibrio cholerae challenge. Front. Immunol. 2023, 13, 1039956. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.N.; Tang, Y.Y.; Zhou, M.J.; Luo, S.; Li, Y.T.; Wang, G.; Zhang, D.Z.; Yang, H.; Tang, B.P.; He, W.F. Differentially expressed genes involved in immune pathways from yellowhead catfish (Tachysurus fulvidraco) after poly (I:C) challenge. Int. J. Biol. Macromol. 2021, 183, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wesener, D.A.; Wangkanont, K.; McBride, R.; Song, X.; Kraft, M.B.; Hodges, H.L.; Zarling, L.C.; Splain, R.A.; Smith, D.F.; Cummings, R.D.; et al. Recognition of microbial glycans by human intelectin-1. Nat. Struct. Mol. Biol. 2015, 22, 603–610. [Google Scholar] [CrossRef]
- Yan, J.; Xu, L.; Zhang, Y.; Zhang, C.; Zhang, C.; Zhao, F.; Feng, L. Comparative genomic and phylogenetic analyses of the intelectin gene family: Implications for their origin and evolution. Dev. Comp. Immunol. 2013, 41, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, K.; Maeda, K.; Saigo, C.; Kito, Y.; Yoshida, K.; Takeuchi, T. Adiponectin and intelectin-1: Important adipokine players in obesity-related colorectal carcinogenesis. Int. J. Mol. Sci. 2017, 18, 866. [Google Scholar] [CrossRef]
- Ding, Z.; Zhao, X.; Zhan, Q.; Cui, L.; Sun, Q.; Lin, L.; Wang, W.; Liu, H. Characterization and expression analysis of an intelectin gene from Megalobrama amblycephala with excellent bacterial binding and agglutination activity. Fish Shellfish Immunol. 2017, 61, 100–110. [Google Scholar] [CrossRef]
- Chen, L.; Yan, J.; Sun, W.; Zhang, Y.; Sui, C.; Qi, J.; Du, Y.; Feng, L. A zebrafish intelectin ortholog agglutinates Gram-negative and Gram-positive bacteria with binding capacity to bacterial polysaccharideS. Fish Shellfish Immunol. 2016, 55, 729–736. [Google Scholar] [CrossRef]
- Takano, T.; Sha, Z.; Peatman, E.; Terhune, J.; Liu, H.; Kucuktas, H.; Li, P.; Edholm, E.-S.; Wilson, M.; Liu, Z. The two channel catfish intelectin genes exhibit highly differential patterns of tissue expression and regulation after infection with Edwardsiella ictaluri. Dev. Comp. Immunol. 2008, 32, 693–705. [Google Scholar] [CrossRef]
- Qiao, G.; Zhang, M.; Li, Y.; Xu, C.; Xu, D.H.; Zhao, Z.; Zhang, J.; Li, Q. Biofloc technology (BFT): An alternative aquaculture system for prevention of Cyprinid herpesvirus 2 infection in gibel carp (Carassius auratus gibelio). Fish Shellfish Immunol. 2018, 83, 140–147. [Google Scholar] [CrossRef]
- Xue, Z.; Pang, Y.; Liu, X.; Zheng, Z.; Xiao, R.; Jin, M.; Han, Y.; Su, P.; Lv, L.; Wang, J.; et al. First evidence of protein G-binding protein in the most primitive vertebrate: Serum lectin from lamprey (Lampetra japonica). Dev. Comp. Immunol. 2013, 41, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, C.; Zhang, Y.; Li, K.; Xu, L.; Guo, L.; Kong, Y.; Feng, L. Characterization and comparative analyses of two amphioxus intelectins involved in the innate immune response. Fish Shellfish Immunol. 2013, 34, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Nonnecke, E.B.; Castillo, P.A.; Dugan, A.E.; Almalki, F.; Underwood, M.A.; De La Motte, C.A.; Yuan, W.; Lu, W.; Shen, B.; Johansson, M.E.V.; et al. Human intelectin-1 (ITLN1) genetic variation and intestinal expression. Sci. Rep. 2021, 11, 12889. [Google Scholar] [CrossRef]
- Nonnecke, E.B.; Castillo, P.A.; Johansson, M.E.V.; Hollox, E.J.; Shen, B.; Lönnerdal, B.; Bevins, C.L. Human intelectin-2 (ITLN2) is selectively expressed by secretory Paneth cells. FASEB J. 2022, 36, e22200. [Google Scholar] [CrossRef]
- Lu, Z.H.; di Domenico, A.; Wright, S.H.; Knight, P.A.; Whitelaw, C.B.; Pemberton, A.D. Strain-specific copy number variation in the intelectin locus on the 129 mouse chromosome 1. BMC Genom. 2011, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Fänge, R.; Nilsson, S. The fish spleen: Structure and function. Experientia 1985, 41, 152–158. [Google Scholar] [CrossRef]
- Yan, J.; Wang, J.; Zhao, Y.; Zhang, J.; Bai, C.; Zhang, C.; Zhang, C.; Li, K.; Zhang, H.; Du, X.; et al. Identification of an amphioxus intelectin homolog that preferably agglutinates gram-positive over gram-negative bacteria likely due to different binding capacity to LPS and PGN. Fish Shellfish Immunol. 2012, 33, 11–20. [Google Scholar] [CrossRef]
- Gerwick, L.; Corley-Smith, G.; Bayne, C.J. Gene transcript changes in individual rainbow trout livers following an inflammatory stimulus. Fish Shellfish Immunol. 2007, 22, 157–171. [Google Scholar] [CrossRef]
- Pemberton, A.D.; Knight, P.A.; Gamble, J.; Colledge, W.H.; Lee, J.K.; Pierce, M.; Miller, H.R. Innate BALB/c enteric epithelial responses to Trichinella spiralis: Inducible expression of a novel goblet cell lectin, intelectin-2, and its natural deletion in C57BL/10 mice. J. Immunol. 2004, 173, 1894–1901. [Google Scholar] [CrossRef]
- Bai, L.; Yu, G.; Liu, Y.; Aizaz, M.; Yang, G.; Chen, L. Common carp intelectin 3 (cITLN3) plays a role in the innate immune response. Fish Shellfish Immunol. 2023, 141, 109057. [Google Scholar] [CrossRef]
- Andresen, S.; Fantone, K.; Chapla, D.; Rada, B.; Moremen, K.W.; Pierce, M.; Szymanski, C.M. Human intelectin-1 promotes cellular attachment and neutrophil killing of Streptococcus pneumoniae in a serotype-dependent manner. Infect Immun. 2022, 90, e0068221. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, S.; Yamashita, M.; Hoffman, D.R.; Nishiyama, A.; Shinohara, T.; Ohtsu, T.; Shibata, Y. Capture of heat-killed Mycobacterium bovis bacillus Calmette-Guérin by intelectin-1 deposited on cell surfaces. Glycobiology 2009, 19, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Pemberton, A.D.; Verdon, B.; Inglis, N.F.; Pearson, J.P. Sheep intelectin-2 co-purifies with the mucin Muc5ac from gastric mucus. Res. Vet. Sci. 2011, 91, e53–e57. [Google Scholar] [CrossRef]
- Nagata, S. Identification and characterization of a novel intelectin in the digestive tract of Xenopus laevis. Dev. Comp. Immunol. 2016, 59, 229–239. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).