

Replacement of Dietary Fishmeal by Black Soldier Fly Larvae (Hermetia illucens) Meal in Practical Diets for Juvenile Tench (Tinca tinca)

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish, Facilities, and Experimental Procedures
2.3. Diets and Feeding
2.4. Chemical Analysis of Diets and Fish
2.5. Data Collection
- -
- Survival rate (%) = (final number of juveniles/initial number of juveniles) × 100;
- -
- Specific growth rate, SGR (% d−1) = [(ln final W − ln initial W)/days)] × 100;
- -
- Fulton’s coefficient or condition factor, K = 100 × [final W/(TL3)];
- -
- Biomass gain, BG (g) = (final biomass/tank − initial biomass/tank);
- -
- Feed conversion ratio, FCR = (total feed provided per tank/BG [62].
2.6. Statistical Analysis
3. Results
3.1. Diets
3.2. Growth Performance
3.3. Juvenile Whole-Body Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Planning for Aquaculture Diversification: The Importance of Climate Change and Other drivers. In Proceedings of the FAO Technical Workshop, Bangkok, Thailand, 23–25 June 2016; Harvey, B., Soto, D., Carosfeld, J., Beveridge, M., Bartley, D.M., Eds.; FAO: Roma, Italy, 2017. 145p. [Google Scholar]
- Freyhof, J.; Kottelat, M. Tinca tinca. The IUCN Red List of Threatened Species 2008, e.T21912A9339248. Available online: https://www.iucnredlist.org/species/21912/9339248 (accessed on 21 November 2022).
- Kamiński, R.; Sikorska, J.; Wolkini, J. Diet and water temperature affect growth and body deformities in juvenile tench Tinca tinca (L.) reared under controlled conditions. Aquac. Res. 2017, 48, 1327–1337. [Google Scholar] [CrossRef]
- Pula, H.J.; Trenzado, C.E.; García-Mesa, S.; Fallola, C.; Sanz, A. Effects of different culture systems on growth, immune status, and other physiological parameters of tench (Tinca tinca L.). Aquaculture 2018, 485, 101–110. [Google Scholar] [CrossRef]
- Wedekind, H.; Rennert, B.; Kohlmann, K. Product quality in different strains of tench (Tinca tinca L.) tested under controlled environmental conditions. J. Appl. Ichthyol. 2003, 19, 174–176. [Google Scholar] [CrossRef]
- González-Rodríguez, Á.; Celada, J.D.; Carral, J.M.; Sáez-Royuela, M.; Fuertes, J.B. Effects of varying protein level in practical diets on survival, growth, feed utilization and body composition of juvenile tench (Tinca tinca L.). Aquac. Int. 2014, 22, 1723–1735. [Google Scholar] [CrossRef]
- Celada, J.D.; Aguilera, A.; García, V.; Carral, J.M.; Saez-Royuela, M.; González, R.; González, A. Rearing juvenile tench (Tinca tinca L.) under controlled conditions using Artemia nauplii as supplement to a dry diet. Aquac. Int. 2009, 17, 565–570. [Google Scholar] [CrossRef]
- García, V.; Celada, J.D.; Carral, J.M.; Sáez-Royuela, M.; González, R.; González, Á. Decapsulated Artemia cysts: A suitable dietary supplement for juvenile tench (Tinca tinca L.). J. Appl. Aquac. 2010, 22, 57–65. [Google Scholar] [CrossRef]
- González-Rodríguez, Á.; Celada, J.D.; Carral, J.M.; Sáez-Royuela, M.; Fuertes, J.B. Evaluation of a practical diet for juvenile tench (Tinca tinca L.) and substitution possibilities of fish meal by feather meal. Anim. Feed. Sci. Technol. 2014, 187, 61–67. [Google Scholar] [CrossRef]
- García, V.; Celada, J.D.; González, R.; Carral, J.M.; Sáez-Royuela, M.; González, Á. Response of juvenile tench (Tinca tinca L.) fed practical diets with different protein contents and substitution levels of fish meal by soybean meal. Aquac. Res. 2015, 46, 28–38. [Google Scholar] [CrossRef]
- EUMOFA—European Market Observatory for Fisheries and Aquaculture Products. Fishmeal and Fish Oil: Production and Trade Flows in the UE; Publications Office of the European Union: Luxembourg, 2021; 31p. [Google Scholar]
- OECD/FAO. OECD-FAO Agricultural Outlook 2022–2031; OECD Publishing: Paris, France, 2022; 363p. [Google Scholar] [CrossRef]
- Boyd, C.E.; D’Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.J.; Teletchea, F.; Tomasso, J.R.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. J. World Aquac. Soc. 2020, 51, 578–633. [Google Scholar] [CrossRef]
- Cottrell, R.S.; Blanchard, J.L.; Halpern, B.S.; Metian, M.; Froehlichet, H.E. Global adoption of novel aquaculture feeds could substantially reduce forage fish demand by 2030. Nat. Food 2020, 1, 301–308. [Google Scholar] [CrossRef]
- Campanati, C.; Willer, D.; Schubert, J.; Aldridge, D.C. Sustainable intensification of aquaculture through nutrient recycling and circular economies: More fish, less waste, blue growth. Rev. Fish. Sci. Aquac. 2022, 30, 143–169. [Google Scholar] [CrossRef]
- Tschirner, M.; Kloas, W. Increasing the sustainability of aquaculture systems: Insects as alternative protein source for fish diets. GAIA-Ecol. Perspect. Sci. Soc. 2017, 26, 332–340. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed. Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal Alternative Protein Sources for Aquaculture Feeds. In Feeds for the Aquaculture Sector; Current Situation and Alternative Sources; Springer Briefs in Molecular Science: Chemistry of Foods; Springer Nature: Cham, Switzerland, 2018; pp. 1–28. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2017/893 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32017R0893&rid=1 (accessed on 21 November 2022).
- Commission Regulation (EU) 2021/1925 of 5 November 2021 Amending Certain Annexes to Regulation (EU) No 142/2011 as Regards the Requirements for Placing on the Market of Certain Insect Products and the Adaptation of a Containment Method. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32021R1925 (accessed on 21 November 2022).
- Barragán-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illuscens L.) and its suitability as animal feed—A review. J. Insects Food Feed. 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the black soldier fly (Hermetia illucens) as fish meal substitute—growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364–365, 345–352. [Google Scholar] [CrossRef]
- Ido, A.; Ali, M.-F.-Z.; Takahashi, T.; Miura, C.; Miura, T. Growth of yellowtail (Seriola quinqueradiata) fed on a diet including partially or completely defatted black soldier fly (Hermetia illucens) larvae meal. Insects 2021, 12, 722. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Daskalopoulou, E.; Vogiatzis, I.; Rumbos, C.; Mente, E.; Athanassiou, C.G. Substitution of fishmeal by fly Hermetia illuscens prepupae meal in the diet of gilthead seabream (Sparus aurata). In Proceedings of the HydroMedit 2014, Volos, Greece, 13–15 November 2014; pp. 110–114. [Google Scholar]
- Madibana, M.J.; Mwanza, M.; Lewis, B.R.; Fouché, C.H.; Toefy, R.; Mlambo, V. Black Soldier Fly Larvae Meal as a Fishmeal Substitute in Juvenile Dusky Kob Diets: Effect on feed utilization, growth performance, and blood parameters. Sustainability 2020, 12, 9460. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, Å.; Lock, E.-J. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]
- Hua, K. A Meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture 2020, 530, 735732. [Google Scholar] [CrossRef]
- Fabrikov, D.; Vargas-García, M.d.C.; Barroso, F.G.; Sánchez-Muros, M.J.; Cacua Ortíz, S.M.; Morales, A.E.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F. Effect on intermediary metabolism and digestive parameters of the high substitution of fishmeal with insect meal in Sparus aurata feed. Insects 2021, 12, 965. [Google Scholar] [CrossRef]
- Magalhães, R.; Sánchez-López, A.; Silva Leal, R.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Khallaf, M.A.; Abdel-Latif, H.M.R. Effects of black soldier fly (Hermetia illucens L.) larvae meal on growth performance, organs-somatic indices, body composition, and hemato-biochemical variables of European sea bass, Dicentrarchus labrax. Aquaculture 2020, 522, 735136. [Google Scholar] [CrossRef]
- Wang, G.; Peng, K.; Hu, J.; Yi, C.; Chen, X.; Wu, H.; Huang, Y. Evaluation of defatted black soldier fly (Hermetia illucens L.) larvae meal as an alternative protein ingredient for juvenile Japanese seabass (Lateolabrax japonicus) diets. Aquaculture 2019, 507, 144–154. [Google Scholar] [CrossRef]
- Li, X.; Qin, C.; Fang, Z.; Sun, X.; Shi, H.; Wang, Q.; Zhao, H. Replacing dietary fish meal with defatted black soldier fly (Hermetia illucens) larvae meal affected growth, digestive physiology and muscle quality of Tongue Sole (Cynoglossus semilaevis). Front. Physiol. 2022, 13, 855957. [Google Scholar] [CrossRef] [PubMed]
- Takakuwa, F.; Tanabe, R.; Nomura, S.; Inui, T.; Yamada, S.; Biswas, A.; Tanaka, H. Availability of black soldier fly meal as an alternative protein source to fish meal in red sea bream (Pagrus major, Temminck & Schlegel) fingerling diets. Aquac. Res. 2022, 53, 36–49. [Google Scholar] [CrossRef]
- Lock, E.R.; Arsiwalla, T.; Waagbø, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbø, R.; Krogdahl, Å.; Lock, E.J. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Sealey, W.M.; Gaylord, T.G.; Barrows, F.T.; Tomberlin, J.K.; McGuire, M.A.; Ross, C.; St-Hilaire, S. Sensory analysis of rainbow trout, Oncorhynchus mykiss, fed enriched black soldier fly prepupae, Hermetia illucens. J. World Aquac. Soc. 2011, 42, 34–45. [Google Scholar] [CrossRef]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Anim. Sci. Biotechnol. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Bruni, L.; Secci, G.; Husein, Y.; Faccenda, F.; Lira de Medeiros, A.C.; Parisi, G. Is it possible to cut down fishmeal and soybean meal use in aquafeed limiting the negative effects on rainbow trout (Oncorhynchus mykiss) fillet quality and consumer acceptance? Aquaculture 2021, 543, 736996. [Google Scholar] [CrossRef]
- Caimi, C.; Renna, M.; Lussiana, C.; Bonaldo, A.; Gariglio, M.; Meneguz, M.; Dabbou, S.; Schiavone, A.; Gai, F.; Elia, A.C.; et al. First insights on Black Soldier Fly (Hermetia illucens L.) larvae meal dietary administration in Siberian sturgeon (Acipenser baerii Brandt) juveniles. Aquaculture 2020, 515, 734539. [Google Scholar] [CrossRef]
- Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. Black soldier fly full-fat larvae meal as an alternative to fish meal and fish oil in Siberian sturgeon nutrition: The effects on physical properties of the feed, animal growth performance, and feed acceptance and utilization. Animals 2020, 10, 2119. [Google Scholar] [CrossRef]
- Muin, H.; Taufek, N.M.; Kamarudin, M.S.; Razak, S.A. Growth performance, feed utilization and body composition of Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) fed with different levels of black soldier fly, Hermetia illucens (Linnaeus, 1758) maggot meal diet. Iran. J. Fish. Sci. 2017, 16, 567–577. [Google Scholar]
- Devic, E.; Leschen, W.; Murray, F.; Little, D.C. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing Black Soldier Fly (Hermetia illucens) larvae meal. Aquac. Nutr. 2018, 24, 416–423. [Google Scholar] [CrossRef]
- Wachira, M.N.; Osuga, I.M.; Munguti, J.M.; Ambula, M.K.; Subramanian, S.; Tanga, C.M. Efficiency and improved profitability of insect-based aquafeeds for farming Nile tilapia fish (Oreochromis niloticus L.). Animals 2021, 11, 2599. [Google Scholar] [CrossRef]
- Tippayadara, N.; Dawood, M.A.O.; Krutmuang, P.; Hoseinifar, S.H.; Doan, H.V.; Paolucci, M. Replacement of fish meal by black soldier fly (Hermetia illucens) larvae meal: Effects on growth, haematology, and skin mucus immunity of Nile tilapia, Oreochromis niloticus. Animals 2021, 11, 193. [Google Scholar] [CrossRef]
- Adeoye, A.A.; Akegbejo-Samsons, Y.; Fawole, F.J.; Davies, S.J. Preliminary assessment of black soldier fly (Hermetia illucens) larval meal in the diet of African catfish (Clarias gariepinus): Impact on growth, body index, and hematological parameters. J. World Aquac. Soc. 2020, 51, 1024–1033. [Google Scholar] [CrossRef]
- Fawole, F.J.; Adeoye, A.A.; Tiamiyu, L.O.; Ajala, K.I.; Obadara, S.O.; Ganiyu, I.O. Substituting fishmeal with Hermetia illucens in the diets of African catfish (Clarias gariepinus): Effects on growth, nutrient utilization, haemato-physiological response, and oxidative stress biomarker. Aquaculture 2020, 518, 734849. [Google Scholar] [CrossRef]
- Vongvichith, B.; Morioka, S.; Sugita, T.; Phousavanh, N.; Phetsanghanh, N.; Chanthasone, P.; Pommachan, P.; Nakamura, S. Evaluation of the efficacy of aquaculture feeds for the climbing perch Anabas testudineus: Replacement of fishmeal by black soldier fly Hermetia illucens prepupae. Fish. Sci. 2020, 86, 145–151. [Google Scholar] [CrossRef]
- Kattakdad, S.; Suratip, N.; Yuangsoi, B.; Kasamawut, K.; Udduang, S. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in climbing perch (Anabas testudineus) diet. Aquacult. Aquar. Conserv. Legis. 2022, 15, 68–82. [Google Scholar]
- Xiao, X.; Jin, P.; Zheng, L.; Cai, M.; Yu, Z.; Yu, J.; Zhang, J. Effects of black soldier fly (Hermetia illucens) larvae meal protein as a fishmeal replacement on the growth and immune index of yellow catfish (Pelteobagrus fulvidraco). Aquac. Res. 2018, 49, 1569–1577. [Google Scholar] [CrossRef]
- Khieokhajonkhet, A.; Uanlam, P.; Ruttarattanamongkol, K.; Aeksiri, N.; Tatsapong, P.; Kaneko, G. Replacement of fish meal by black soldier fly larvae meal in diet for goldfish Carassius auratus: Growth performance, hematology, histology, total carotenoids, and coloration. Aquaculture 2022, 561, 738618. [Google Scholar] [CrossRef]
- Kamalii, A.; Antony, C.; Ahilan, B.; Uma, A.; Prabu, E. Dietary protein replacement of fish meal with black soldier fly larvae meal: Effects on growth, whole-body composition, digestive enzyme activity, muscle-growth-related gene expression and haemato-biochemical responses of juvenile goldfish, Carassius auratus. Turk. J. Fish. Aquat. Sci. 2023, 23, 21837. [Google Scholar] [CrossRef]
- Zhou, J.S.; Liu, S.S.; Ji, H.; Yu, H.B. Effect of replacing dietary fish meal with black soldier fly larvae meal on growth and fatty acid composition of Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2018, 24, 424–433. [Google Scholar] [CrossRef]
- Katya, K.; Borsra, M.Z.S.; Ganesan, D.; Kuppusamy, G.; Herriman, M.; Salter, A.; Ali, S.A. Efficacy of insect larval meal to replace fish meal in juvenile barramundi, Lates calcarifer reared in freshwater. Int. Aquat. Res. 2017, 9, 303–312. [Google Scholar] [CrossRef]
- Sudha, C.; Ahilan, B.; Felix, N.; Uma, A.; Prabu, E. Effects of dietary protein substitution of fishmeal with black soldier fly larval meal on growth and physiological responses of juvenile striped catfish, Pangasianodon hypophthalmus. Aquac. Res. 2022, 53, 2204–2217. [Google Scholar] [CrossRef]
- Rodríguez, R.; Celada, J.D.; Sáez-Royuela, M.; Carral, J.M.; Aguilera, A.; Melendre, P.M. Artificial reproduction in 1-year-old tench (Tinca tinca L.). J. Appl. Ichthyol. 2004, 20, 542–544. [Google Scholar] [CrossRef]
- Cohen, S.A.; Michaud, D.P. Synthesis of a fluorescent derivatizing reagent, 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate, and its application for the analysis of hydrolysate amino acids via high-performance liquid chromatography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.A.; De Antonis, K.M. Applications of amino acid derivatization with 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate. J. Chromatogr. A 1994, 661, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Fornshell, G.; Hinshaw, J.M. Better Management Practices for Flow-Through Aquaculture Systems. In Environmental Best Management Practices for Aquaculture; Tucker, C.S., Hargreaves, J.A., Eds.; Blackwell Publishing: Ames, IA, USA, 2008; pp. 331–388. [Google Scholar]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect meals in fish nutrition. Rev. Aquac. 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Perrow, M.R.; Jowitt, A.J.D.; Johnsonf, S.R. Factors affecting the habitat selection of tench in a shallow eutrophic lake. J. Fish Biol. 1996, 48, 859–870. [Google Scholar] [CrossRef]
- Pyka, J. Daily feeding cycle tench, Tinca tinca (L.), in larval and fry stages in the conditions of pond culture. An attempt to determine daily food ration. Arch. Pol. Fish. 1997, 5, 279–290. [Google Scholar]
- Alaş, A.; Altindağ, A.; Yılmaz, M.; Kırpık, M.A.; Ak, A. Feeding habits of tench (Tinca tinca L., 1758) in Beyşehir lake (Turkey). Turk. J. Fish. Aquat. Sci. 2010, 10, 187–194. [Google Scholar] [CrossRef]
- Lu, S.; Taethaisong, N.; Meethip, W.; Surakhunthod, J.; Sinpru, B.; Sroichak, T.; Archa, P.; Thongpea, S.; Paengkoum, S.; Purba, R.A.P.; et al. Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: A review. Insects 2022, 13, 831. [Google Scholar] [CrossRef]
- Liland, N.S.; Araujo, P.; Xu, X.X.; Lock, E.J.; Radhakrishnan, G.; Prabhu, A.J.P.; Belghit, I. A meta-analysis on the nutritional value of insects in aquafeeds. J. Insects Food Feed 2021, 7, 743–759. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.-J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Maurer, V.; Holinger, M.; Amsler, Z.; Früh, B.; Wohlfahrt, J.; Stamer, A.; Leiber, F. Replacement of soybean cake by Hermetia illucens meal in diets for layers. J. Insects Food Feed 2016, 2, 83–90. [Google Scholar] [CrossRef]
- Basto, A.; Matos, I.; Valente, L.M.P. Nutritional value of different insect larvae meals as protein sources for European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2020, 21, 735085. [Google Scholar] [CrossRef]
- Jobling, M. Nutrient Partitioning and the Influence of Feed Composition on Body Composition. In Food Intake in Fish; Houlihan, D., Boujard, T., Jobling, M., Eds.; Blackwell: New York, NY, USA, 2001; pp. 354–375. [Google Scholar]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The Lipids. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 181–257. [Google Scholar]
- Ferrer Llagostera, P.; Kallas, Z.; Reig, L.; Amores de Gea, D. The use of insect meal as a sustainable feeding alternative in aquaculture: Current situation, Spanish consumers’ perceptions and willingness to pay. J. Clean. Prod. 2019, 229, 10–21. [Google Scholar] [CrossRef]
- Lugert, V.; Thaller, G.; Tetens, J.; Schulz, C.; Krieter, J. A review on fish growth calculation: Multiple functions in fish production and their specific application. Rev. Aquac. 2016, 8, 30–42. [Google Scholar] [CrossRef]
- Cahu, C.; Zambonino-Infante, J.; Takeuchi, T. 2003. Nutritional components affecting skeletal development in fish larvae. Aquaculture 2003, 227, 245–258. [Google Scholar] [CrossRef]
- Fontagné, S. The Impact of Nutritional Components on Rainbow Trout. In Control of Malformations in Fish Aquaculture: Science and Practice; Baeverfjord, G., Helland, S., Hough, C., Eds.; RapidPress: Luxembourg, 2009; pp. 73–83. [Google Scholar]
- Kamler, E.; Myszkowski, L.; Kaminski, R.; Korwin-Kossakowski, M.; Wolnicki, J. Does overfeeding affect tench Tinca tinca L. juvenile? Aquac. Int. 2006, 14, 99–111. [Google Scholar] [CrossRef]
- Wolnicki, J.; Myszkowski, L.; Korwin-Kossakowski, M.; Kaminski, R.; Stanny, L.A. Effects of different diets on juvenile tench Tinca tinca (L.) reared under controlled conditions. Aquac. Int. 2006, 14, 89–98. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Authors | Species | Initial Weight (g) | BSFLM (g kg−1) | FM Replacement (%) |
|---|---|---|---|---|
| Marine | ||||
| Kroeckel et al. [23] | Turbot (Scophthalmus maximus) | 54.9 | -a | 0 |
| Ido et al. [24] | Yellowtail (Seriola quinqueradiata) | 0.7 | - | 0 |
| Karapanagiotidis et al. [25] | Gilthead seabream (Sparus aurata) | 1.47 | -b | 0 |
| Fabrikov et al. [30] | Gilthead seabream | 6.76 | 109 * | 30% |
| Madibana et al. [26] | Dusky kob (Argyrosomus japonicus) | 25.5 | 200 * | 20% |
| Magalhães et al. [31] | European seabass (Dicentrarchus labrax) | 50.0 | 195 | 45% |
| Abdel-Tawwab et al. [32] | European seabass | 12.1 | 148 | 50% |
| Wang et al. [33] | Japanese seabass (Lateolabrax japonicus) | 14.1 | 192 | 64% |
| Li et al. [34] | Tongue sole (Cynoglossus semilaevis) | 543 | 144 | 25% |
| Takakuwa et al. [35] | Red sea bream (Pagrus major) | 17.9 | 293 | 41.7% |
| Lock et al. [36] | Atlantic salmon (Salmo salar) | 250 | 100 | 50% |
| Belghit et al. [27] | Atlantic salmon | 7.9 | 600 * | 100% |
| Belghit et al. [37] | Atlantic salmon | 1398 | 147 * | 100% |
| Freshwater | ||||
| St-Hilaire et al. [38] | Rainbow trout (Oncorhynchus mykiss) | 22.6 | 149 c | 25% |
| Sealey et al. [39] | Rainbow trout | 146 | 362 d | 50% |
| Renna et al. [40] | Rainbow trout | 179 | 400 | 50% |
| Dumas et al. [41] | Rainbow trout | 45.7 | 131 | 50% |
| Fabrikov et al. [26] | Rainbow trout | 55.0 | 109 * | 30% |
| Bruni et al. [42] | Rainbow trout | 54.2 | 450 * | 100% |
| Caimi et al. [43] | Siberian sturgeon (Acipenser baerii) | 24.2 | 185 | 25% |
| Rawski et al. [44] | Siberian sturgeon | 14.4 | 300 * | 63.1% |
| Muin et al. [45] | Nile tilapia (Oreochromis niloticus) | 3.0 | 150 | 50% |
| Devic et al. [46] | Nile tilapia | 5.7 | 80 | 70% |
| Wachira et al. [47] | Nile tilapia | 35 | 336 | 67% |
| Tippayadara et al. [48] | Nile tilapia | 14.8 | 100 * | 100% |
| Adeoye et al. [49] | African catfish (Clarias gariepinus) | 2.67 | 75 | 50% |
| Fawole et al. [50] | African catfish | 14 | 172 | 75% |
| Vongvichith et al. [51] | Climbing perch (Anabas testudiensis) | 4.42 | 290 * | 100% |
| Kattakdad et al. [52] | Climbing perch | 3.12 | 175 | 50% |
| Xiao et al. [53] | Yellow catfish (Pelteobagrus fulvidraco) | 48.5 | 223 | 48% |
| Khieokhajonkhet et al. [54] | Goldfish (Carassius auratus) | 9.5 | 210 * | 50.6% |
| Kamalii et al., [55] | Goldfish | 2.8 | 201 | 40% |
| Li et al. [28] | Jian carp (Cyprinus carpio var. Jian) | 34.8 | 106 * | 100% |
| Zhou et al. [56] | Jian carp | 10.1 | 140 * | 100% |
| Katya et al. [57] | Barramundi (Lates calcarifer) | 6.7 | 308 | 50% |
| Shuda et al. [58] | Striped catfish (Pangasianodon hypophthalmus) | 3.1 | 174 | 60% |
| Fabrikov et al. [30] | Tench (Tinca tinca) | 18.3 | 109 * | 30% |
| Fishmeal (FM) | Black Soldier Fly Larvae Meal (BSFLM) | |
|---|---|---|
| Proximate composition | ||
| Moisture | 76 ± 1.0 | 54.7 ± 0.9 |
| Crude protein | 668 ± 2.9 | 565 ± 1.5 |
| Crude fat | 96 ± 2.7 | 139 ± 3.4 |
| Carbohydrates | nd | 161.4 ± 3.6 |
| Ash | 162 ± 3.9 | 79.9 ± 1.2 |
| Essential amino acid (EAA) | ||
| Arginine | 38.2 ± 1.0 | 20.7 ± 0.8 |
| Histidine | 36.0 ± 1.1 | 29.5 ± 1.0 |
| Isoleucine | 23.3 ± 0.9 | 26.7 ± 1.0 |
| Leucine | 44.3 ± 1.6 | 35.6 ± 1.2 |
| Lysine | 51.5 ± 1.5 | 26.2 ± 1.1 |
| Methionine | 19.7 ± 0.8 | 8.7 ± 0.3 |
| Phenylalanine | 28.0 ± 1.2 | 18.6 ± 0.7 |
| Threonine | 24.1 ± 0.9 | 20.8 ± 0.8 |
| Tryptophan | 7.1 ± 0.4 | 4.2 ± 0.2 |
| Valine | 28.2 ± 1.1 | 34.1 ± 1.3 |
| Non-essential amino acid (NEAA) | ||
| Alanine | 43.4 ± 1.4 | 34.2 ± 1.2 |
| Aspartate | 67.8 ± 1.5 | 39.5 ± 1.3 |
| Cysteine | 6.0 ± 0.2 | 4.1 ± 0.2 |
| Glutamate | 85.4 ± 1.2 | 49.3 ± 1.2 |
| Glycine | 48.9 ± 1.6 | 29.0 ± 1.0 |
| Proline | 28.4 ± 1.2 | 34.8 ± 1.3 |
| Serine | 30.5 ± 1.1 | 22.5 ± 0.9 |
| Tyrosine | 23.1 ± 0.9 | 23.2 ± 0.8 |
| FM Replacement (%) | ||||||
|---|---|---|---|---|---|---|
| 0 | 15 | 30 | 45 | 60 | 75 | |
| Ingredients (g kg−1) | ||||||
| Fishmeal 1 | 645 | 548 | 451.5 | 355 | 258 | 162 |
| Hermetia illucens meal 2 | - | 117 | 232 | 348 | 464 | 579 |
| Corn meal 3 | 166 | 146 | 127.5 | 108 | 89 | 70 |
| Dried Artemia cysts 4 | 100 | 100 | 100 | 100 | 100 | 100 |
| Carboxymethyl cellulose 5 | 30 | 30 | 30 | 30 | 30 | 30 |
| Cod liver oil 6 | 20 | 20 | 20 | 20 | 20 | 20 |
| L-ascorbyl-2-monophosphate-N 7 | 5 | 5 | 5 | 5 | 5 | 5 |
| Dicalcium phosphate 7 | 10 | 10 | 10 | 10 | 10 | 10 |
| Choline chloride 7 | 3 | 3 | 3 | 3 | 3 | 3 |
| Soy lecithin 8 | 10 | 10 | 10 | 10 | 10 | 10 |
| Sodium chloride 9 | 1 | 1 | 1 | 1 | 1 | 1 |
| Mineral and Vitamin premix 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| FM Replacement (%) | ||||||
|---|---|---|---|---|---|---|
| Proximate composition (g kg−1) | 0 | 15 | 30 | 45 | 60 | 70 |
| Moisture | 61.0 ± 0.4 | 63.1 ± 0.5 | 60.4 ± 0.3 | 60.5 ± 0.2 | 60.0 ± 0.3 | 60.7 ± 0.4 |
| Crude protein | 501 ± 3.2 | 503 ± 2.8 | 502 ± 2.0 | 502 ± 0.9 | 501 ± 1.8 | 500 ± 1.8 |
| Crude lipid | 107.5 ± 2.1 | 108.3 ± 2.0 | 112.5 ± 2.2 | 118.3 ± 2.3 | 118.5 ± 2.3 | 123 ± 2.5 |
| Carbohydrates | 200.1 ± 2.8 | 201.4 ± 2.7 | 206.8 ± 3.0 | 208.8 ± 3.1 | 215.7 ± 3.3 | 218.7 ± 3.2 |
| Ash | 130.4 ± 1.6 | 124.2 ± 1.5 | 118.3 ± 1.2 | 110.4 ± 0.9 | 104.8 ± 0.9 | 97.6 ± 0.9 |
| Gross energy (MJ kg−1) | 15.8 ± 0.7 | 15.9 ± 0.6 | 16.0 ± 0.6 | 16.2 ± 0.5 | 16.1 ± 0.5 | 16.3 ± 0.6 |
| Essential amino acid (EAA) | ||||||
| Arginine | 42.3 ± 0.2 | 31.3 ± 0.3 | 28.4 ± 0.3 | 25.8 ± 0.4 | 21.3 ± 0.3 | 19.6 ± 0.2 |
| Histidine | 38.9 ± 0.6 | 38.7 ± 0.1 | 34.7 ± 0.2 | 34.5 ± 0.3 | 34.1 ± 0.1 | 32.9 ± 0.2 |
| Isoleucine | 18.6 ± 0.1 | 19.4 ± 0.1 | 20.6 ± 0.2 | 21.3 ± 0.2 | 21.7 ± 0.1 | 22.1 ± 0.1 |
| Leucine | 34.7 ± 0.3 | 33.9 ± 0.2 | 33.0 ± 0.2 | 32.2 ± 0.1 | 32.0 ± 0.2 | 31.5 ± 0.3 |
| Lysine | 38.5 ± 0.2 | 36.7 ± 0.5 | 33.5 ± 0.1 | 29.8 ± 0.1 | 27.6 ± 0.4 | 26.2 ± 0.3 |
| Methionine | 21.0 ± 0.2 | 18.4 ± 0.2 | 16.7 ± 0.4 | 14.5 ± 0.2 | 11.3 ± 0.3 | 10.5 ± 0.3 |
| Phenylalanine | 20.7 ± 0.4 | 19.5 ± 0.3 | 19.1 ± 0.1 | 18.4 ± 0.3 | 17.8 ± 0.2 | 17.3 ± 0.3 |
| Threonine | 19.5 ± 0.3 | 19.1 ± 0.1 | 18.8 ± 0.3 | 18.6 ± 0.4 | 17.5 ± 0.4 | 17.3 ± 0.3 |
| Tryptophan | 4.6 ± 0.3 | 4.2 ± 0.1 | 4.2 ± 0.2 | 4.0 ± 0.1 | 3.9 ± 0.1 | 3.8 ± 0.1 |
| Valine | 21.2 ± 0.2 | 22.3 ± 0.3 | 22.7 ± 0.5 | 23.4 ± 0.4 | 25.0 ± 0.1 | 25.3 ± 0.3 |
| ∑EAA | 259 | 243 | 232 | 222 | 212 | 206 |
| Non-essential amino acid (NEAA) | ||||||
| Alanine | 30.2 ± 0.2 | 29.8 ± 0.2 | 29.1 ± 0.4 | 28.6 ± 0.5 | 27.9 ± 0.4 | 27.1 ± 0.5 |
| Aspartate | 50.2 ± 0.2 | 49.6 ± 0.1 | 49.3 ± 0.4 | 48.2 ± 0.1 | 47.5 ± 0.5 | 46.5 ± 0.6 |
| Cysteine | 4.2 ± 0.2 | 4.0 ± 0.2 | 3.9 ± 0.3 | 3.5 ± 0.3 | 3.2 ± 0.2 | 3.0 ± 0.2 |
| Glutamate | 60.3 ± 0.3 | 57.4 ± 0.3 | 54.6 ± 0.4 | 47.5 ± 0.3 | 45.1 ± 0.3 | 43.9 ± 0.4 |
| Glycine | 33.5 ± 0.5 | 32.0 ± 0.3 | 31.3 ± 0.4 | 29.3 ± 0.1 | 28.0 ± 0.6 | 26.6 ± 0.5 |
| Proline | 20.9 ± 0.6 | 21.7 ± 0.4 | 23.2 ± 0.3 | 25.8 ± 0.5 | 26.6 ± 0.4 | 27.5 ± 0.4 |
| Serine | 21.9 ± 0.3 | 20.8 ± 0.5 | 20.4 ± 0.2 | 20.0 ± 0.1 | 19.5 ± 0.3 | 19.1 ± 0.2 |
| Tyrosine | 17.2 ± 0.2 | 17.1 ± 0.3 | 17.2 ± 0.2 | 17.4 ± 0.4 | 17.6 ± 0.4 | 17.8 ± 0.3 |
| ∑NEAA | 238 | 232 | 229 | 220 | 215 | 211 |
| ∑EAA/∑NEAA | 1.09 | 1.05 | 1.01 | 1.01 | 0.98 | 0.98 |
| FM Replacement (%) | Polynomial Contrasts | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 15 | 30 | 45 | 60 | 75 | SEM | ANOVA | Linear | Quadratic | Cubic | |
| Survival (%) | 100 | 97.9 | 95.8 | 99.0 | 100 | 96.9 | 0.58 | 0.31 | 0.72 | 0.18 | 0.87 |
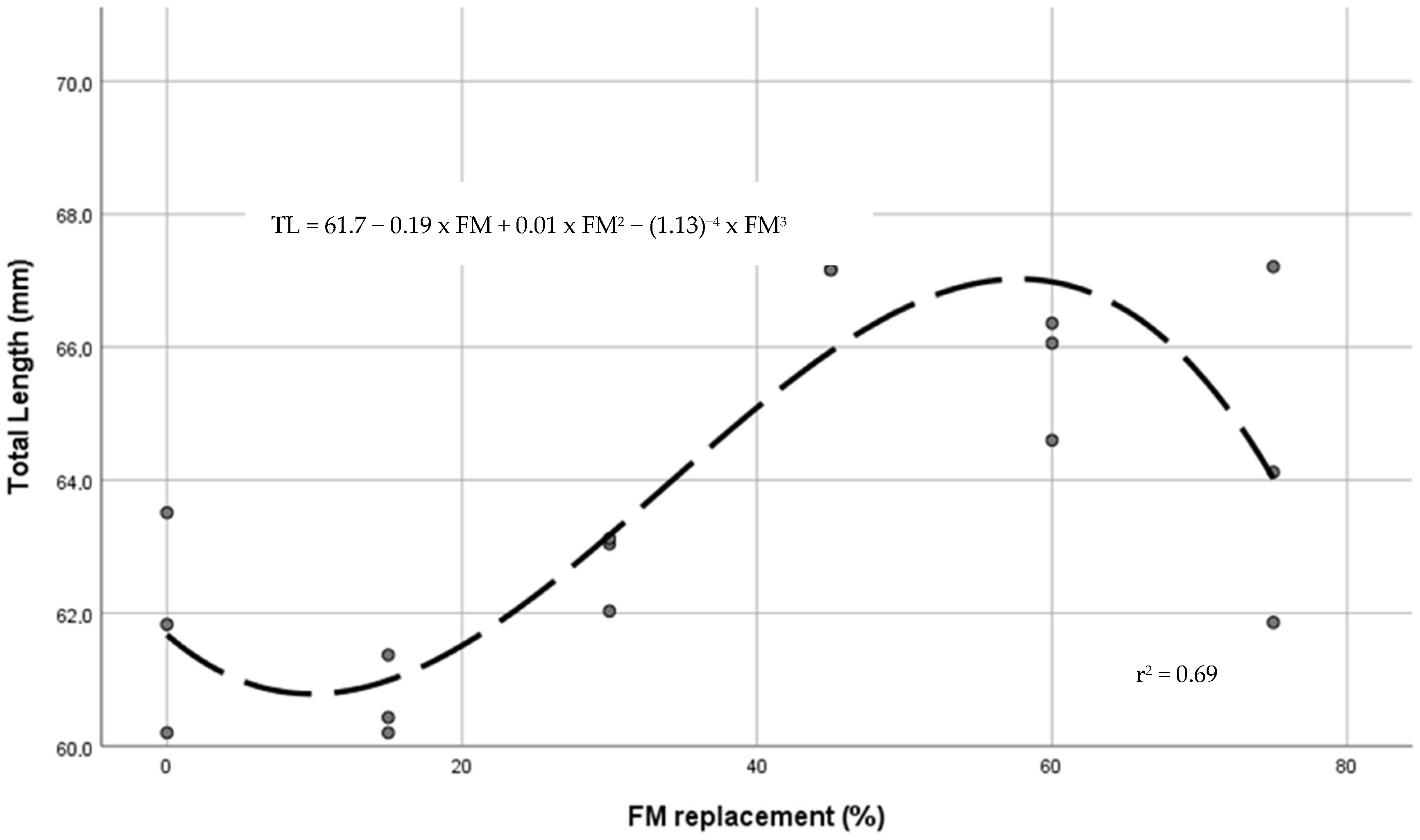
| Total length (mm) | 61.8 a,b | 60.7 a | 62.7 a,b | 67.5 c | 65.7 b,c | 64.4 a,b,c | 0.63 | 0.001 | 0.000 | 0.054 | 0.003 |
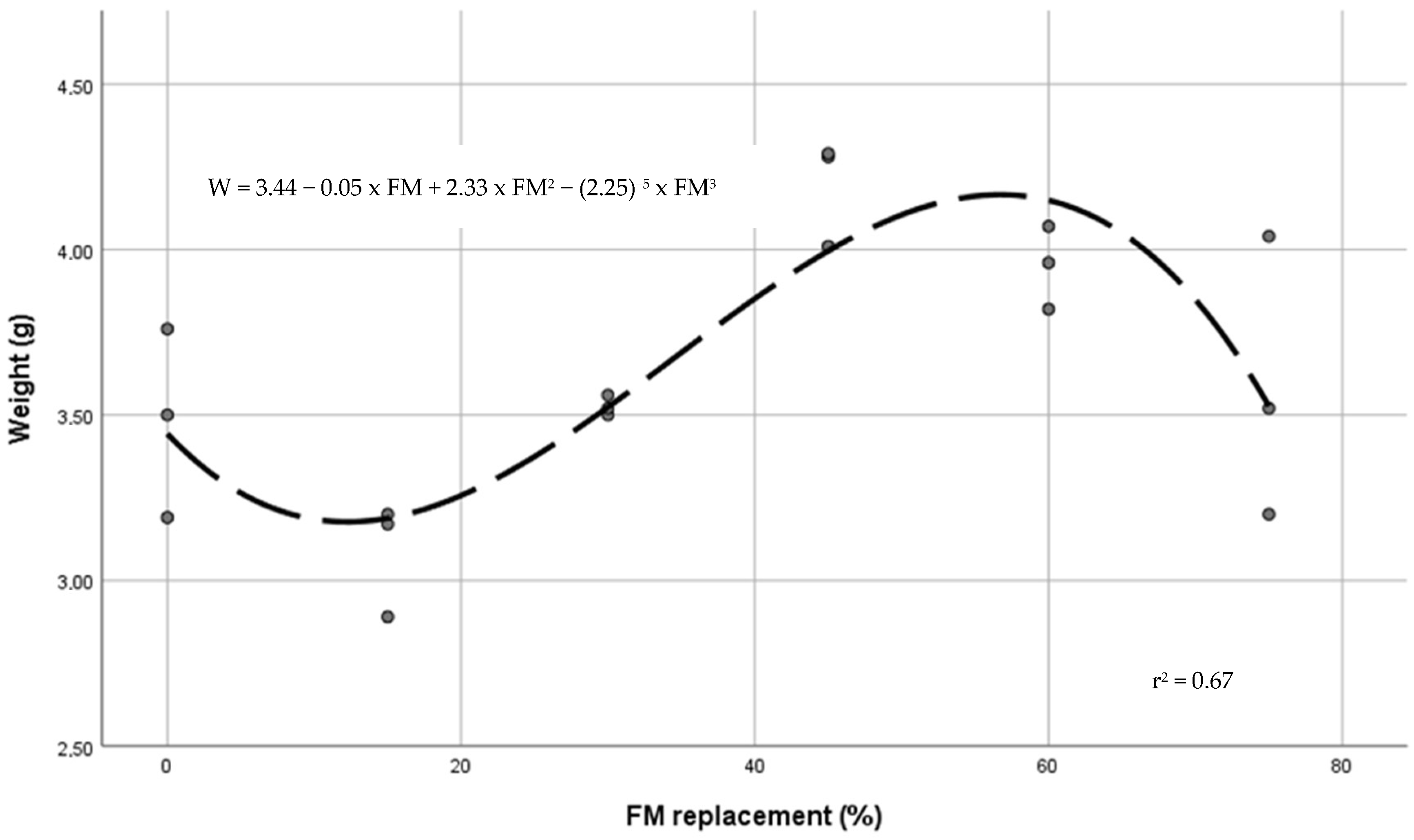
| Weight (g) | 3.48 a,b | 3.09 a | 3.53 a,b | 4.19 c | 3.95 b,c | 3.59 a,b,c | 0.10 | 0.001 | 0.006 | 0.003 | 0.001 |
| SGR 1 (% day−1) | 2.40 a,b | 2.31 a | 2.38 a,b | 2.62 c | 2.53 b,c | 2.45 a,b | 0.03 | 0.007 | 0.020 | 0.011 | 0.004 |
| K 2 | 1.38 a,b | 1.30 b | 1.32 a,b | 1.30 b | 1.28 b | 1.27 b | 0.01 | 0.005 | 0.000 | 0.228 | 0.183 |
| FCR 3 | 1.25 a,b,c | 1.46 a | 1.33 a,b | 1.08 c | 1.18 b,c | 1.30 a,b,c | 0.03 | 0.002 | 0.062 | 0.326 | 0.000 |
| BG 4 (g) | 92.5 a,b | 79.2 b | 90.3 a,b | 115 c | 106 b,c | 95.7 a,b | 3.12 | 0.001 | 0.000 | 0.078 | 0.000 |
| Deformed fish | 0.04 | 0.01 | 0.02 | 0.01 | 0 | 0.02 | 0.007 | 0.64 | 0.33 | 0.28 | 0.87 |
| FM Replacement (%) | Polynomial Contrasts | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial | 0 | 15 | 30 | 45 | 60 | 75 | SEM | ANOVA | Linear | Quadratic | Cubic | |
| Moisture | 767 | 732 | 732 | 753 | 756 | 757 | 760 | 3.98 | 0.06 | 0.006 | 0.31 | 0.62 |
| Protein | 133.5 | 160 | 160 | 159 | 159 | 162 | 158 | 1.45 | 0.93 | 0.71 | 0.71 | 0.54 |
| Lipid | 73.0 | 80.5 a | 78.9 a,b | 55.3 b,c | 55.1 b,c | 46.2 c | 45.7 c | 4.48 | 0.003 | <0.001 | 0.15 | 0.36 |
| Ash | 24.5 | 26.0 | 25.6 | 23.3 | 24.7 | 26.4 | 27.0 | 1.13 | 0.98 | 0.78 | 0.54 | 0.90 |
| Essential amino acids (EAA) | ||||||||||||
| Arginine | 9.5 | 11.6 | 11.9 | 12.1 | 12.5 | 12.7 | 11.4 | 0.16 | 0.11 | 0.49 | 0.02 | 0.08 |
| Histidine | 2.5 | 4.60 | 4.00 | 4.20 | 4.50 | 4.40 | 4.00 | 0.12 | 0.70 | 0.61 | 0.95 | 0.16 |
| Isoleucine | 20.1 | 20.2 a,b | 19.8 a | 21.1 a | 19.6 a,b | 18.9 b | 19.3 a,b | 0.24 | 0.04 | 0.02 | 0.25 | 0.14 |
| Leucine | 4.9 | 5.80 | 5.90 | 5.40 | 5.20 | 5.70 | 5.40 | 0.12 | 0.66 | 0.35 | 0.54 | 0.97 |
| Lysine | 10.2 | 11.3 | 11.8 | 11.2 | 11.3 | 12.3 | 12.6 | 0.19 | 0.09 | 0.03 | 0.12 | 0.58 |
| Methionine | 3.9 | 5.40 | 4.65 | 4.40 | 4.70 | 4.50 | 4.60 | 0.14 | 0.39 | 0.17 | 0.70 | 0.37 |
| Phenylalanine | 11.2 | 12.0 | 13.0 | 12.70 | 12.7 | 13.4 | 12.8 | 0.15 | 0.15 | 0.08 | 0.20 | 0.81 |
| Threonine | 5.5 | 5.80 | 6.70 | 6.60 | 7.55 | 6.80 | 6.20 | 0.19 | 0.10 | 0.29 | 0.02 | 0.60 |
| Tryptophan | 0.4 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.03 | 0.10 | 1.00 | 1.00 | 1.00 | 1.00 |
| Valine | 6.1 | 6.20 | 7.40 | 6.90 | 7.50 | 6.80 | 6.20 | 0.18 | 0.01 | 0.67 | 0.02 | 0.69 |
| ∑EAA | 74.3 | 83.5 | 85.7 | 85.2 | 86.1 | 86.1 | 82.5 | 0.43 | 0.06 | 0.35 | 0.08 | 0.12 |
| Non-essential amino acids (NEAA) | ||||||||||||
| Alanine | 7.7 | 8.8 a | 10.7 b | 10.3 a,b | 9.1 a,b | 10.6 a,b | 9.6 a,b | 0.24 | 0.02 | 0.39 | 0.02 | 0.07 |
| Aspartate | 13.0 | 13.9 a | 15.6 b | 15.6 b | 15.0 a,b | 15.3 a,b | 15.1 a,b | 0.19 | 0.03 | 0.09 | 0.02 | 0.03 |
| Cysteine | 0.2 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 0.02 | 0.29 | 1.00 | 0.04 | 1.00 |
| Glutamate | 17.7 | 20.1 a | 22.5 b | 20.7 a,b | 21.9 b,c | 21.5 a,b,c | 20.7 a,b | 0.26 | 0.006 | 0.61 | 0.006 | 0.20 |
| Glycine | 2.5 | 8.7 a | 9.8 ab | 12.4 d | 11.6 cd | 12.2 cd | 10.7 b,c | 0.41 | <0.001 | <0.001 | <0.001 | 0.37 |
| Proline | 4.9 | 5.3 a | 7.2 b | 7.7 b | 6.5 a,b | 7.9 b | 6.4 a,b | 0.28 | 0.007 | 0.04 | 0.003 | 0.27 |
| Serine | 6.3 | 6.4 | 6.9 | 6.7 | 7.1 | 7.3 | 6.2 | 0.14 | 0.15 | 0.80 | 0.04 | 0.18 |
| Tyrosine | 0.2 | 4.6 a | 0.2 b | 0.2 b | 0.1 b | 0.1 b | 0.1 b | 0.52 | <0.001 | <0.001 | <0.001 | <0.001 |
| ∑NEAA | 52.5 | 68.0 a | 73.0 b | 73.7 b,d | 71.4 c | 75.0 d | 69.0 a | 0.76 | 0.001 | 0.007 | <0.001 | 0.95 |
| ∑EAA/∑NEAA | 1.41 | 1.23 | 1.17 | 1.16 | 1.20 | 1.15 | 1.19 | 0.01 | 0.35 | 0.37 | 0.18 | 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carral, J.M.; Sáez-Royuela, M. Replacement of Dietary Fishmeal by Black Soldier Fly Larvae (Hermetia illucens) Meal in Practical Diets for Juvenile Tench (Tinca tinca). Fishes 2022, 7, 390. https://doi.org/10.3390/fishes7060390
Carral JM, Sáez-Royuela M. Replacement of Dietary Fishmeal by Black Soldier Fly Larvae (Hermetia illucens) Meal in Practical Diets for Juvenile Tench (Tinca tinca). Fishes. 2022; 7(6):390. https://doi.org/10.3390/fishes7060390
Chicago/Turabian StyleCarral, Jose M., and María Sáez-Royuela. 2022. "Replacement of Dietary Fishmeal by Black Soldier Fly Larvae (Hermetia illucens) Meal in Practical Diets for Juvenile Tench (Tinca tinca)" Fishes 7, no. 6: 390. https://doi.org/10.3390/fishes7060390
APA StyleCarral, J. M., & Sáez-Royuela, M. (2022). Replacement of Dietary Fishmeal by Black Soldier Fly Larvae (Hermetia illucens) Meal in Practical Diets for Juvenile Tench (Tinca tinca). Fishes, 7(6), 390. https://doi.org/10.3390/fishes7060390