
Yolk Absorption Rate and Mouth Development in Larvae of Dormitator latifrons (Perciformes: Eleotridae)

 , ,
, ,  and
and 
Abstract
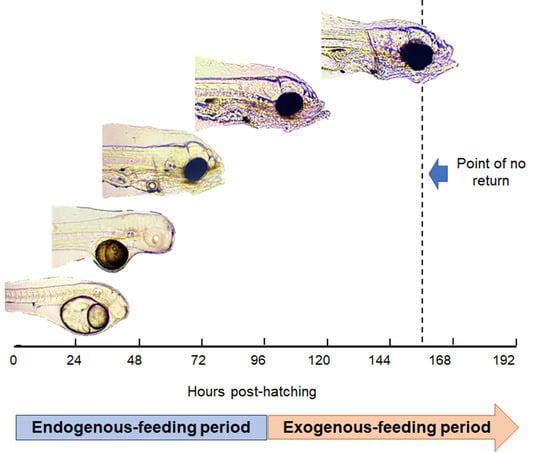
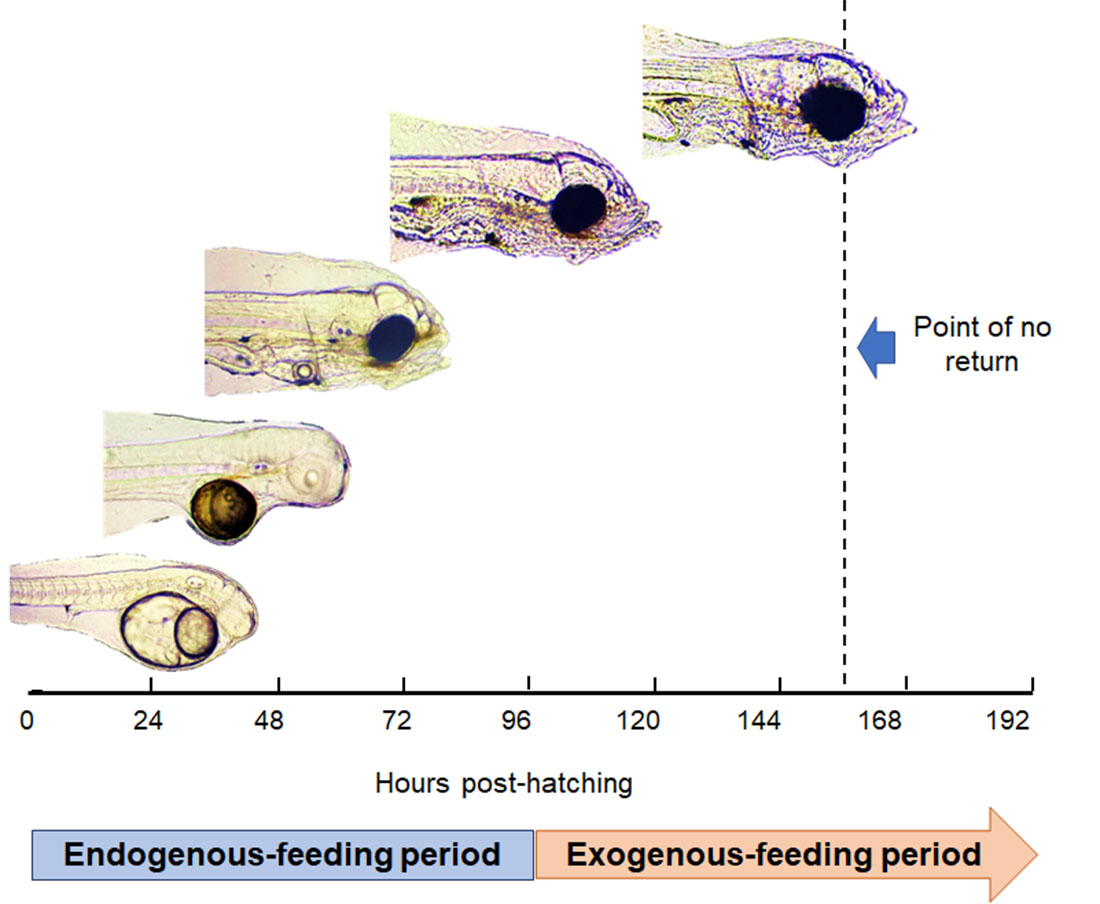
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Spawning and Larval Rearing
2.3. Data Analysis
3. Results
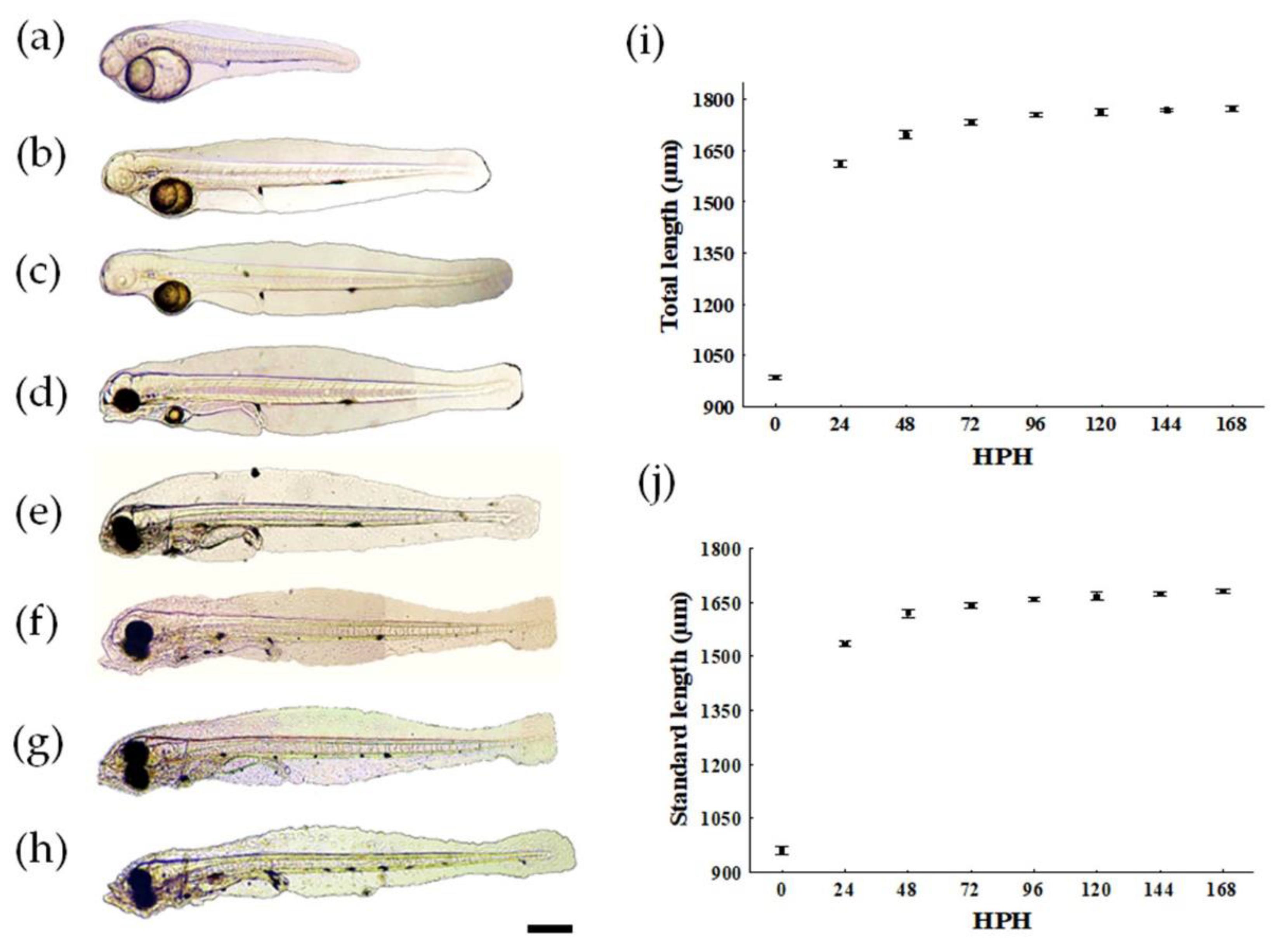
3.1. Larval Growth
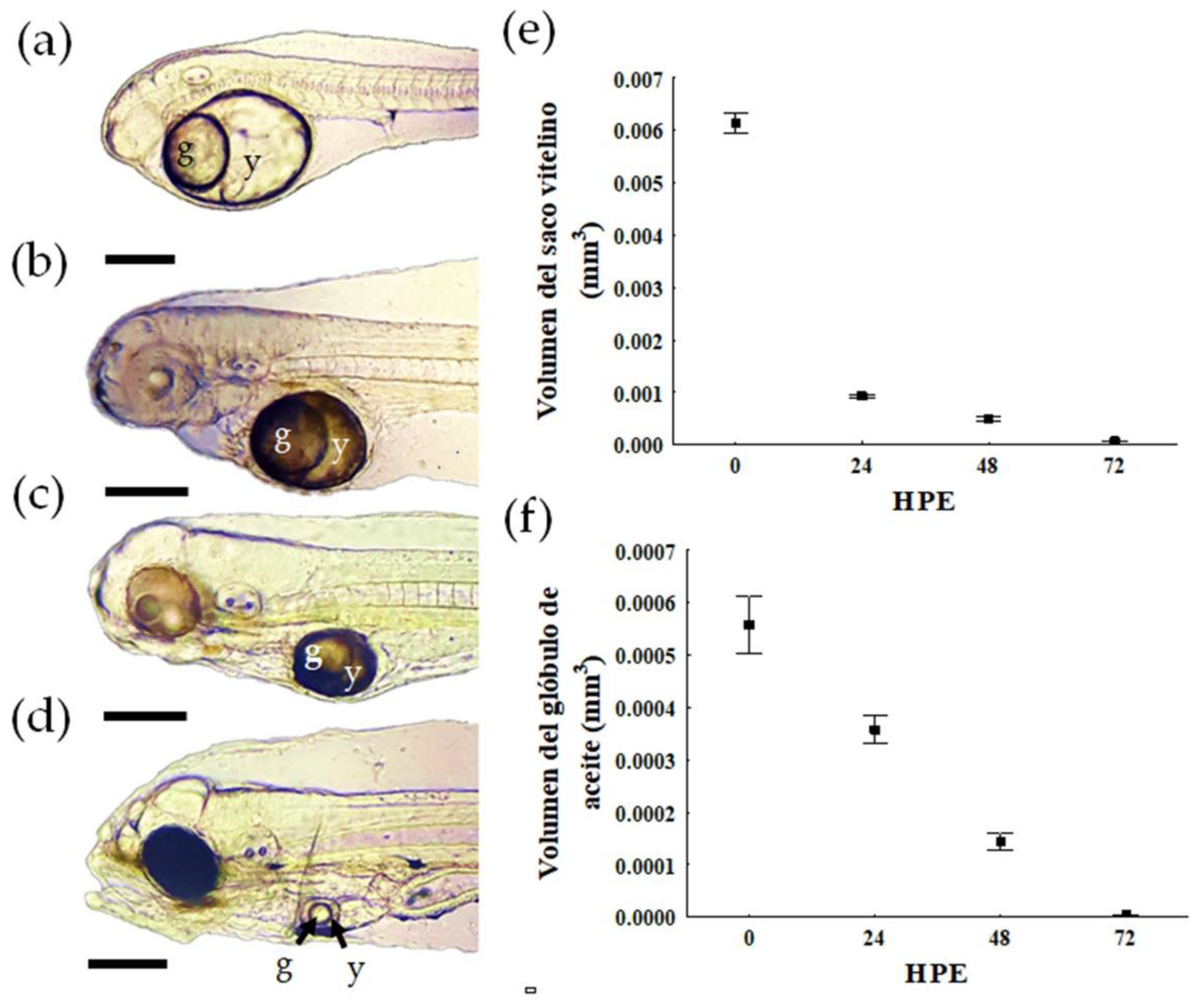
3.2. Yolk Absorption
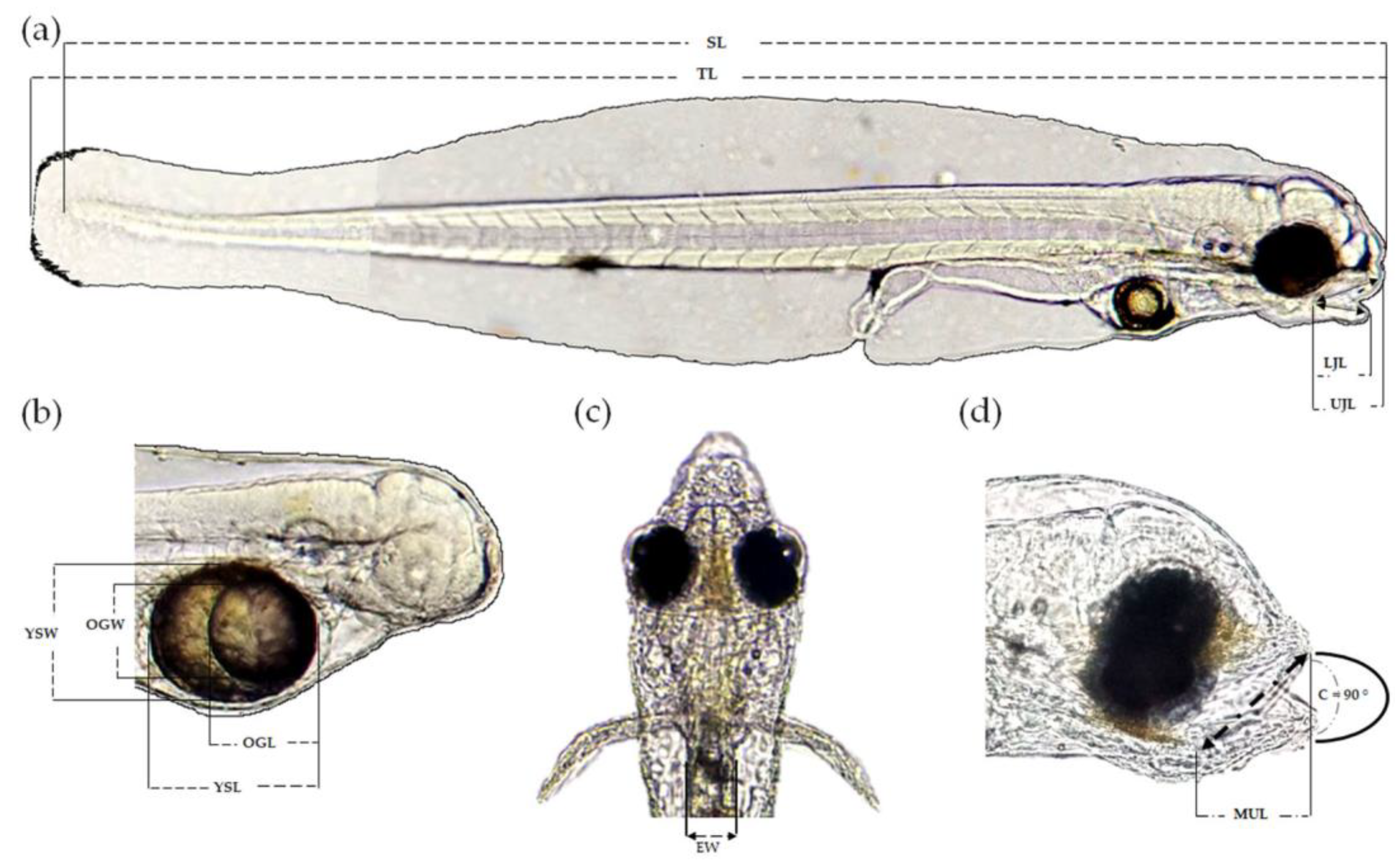
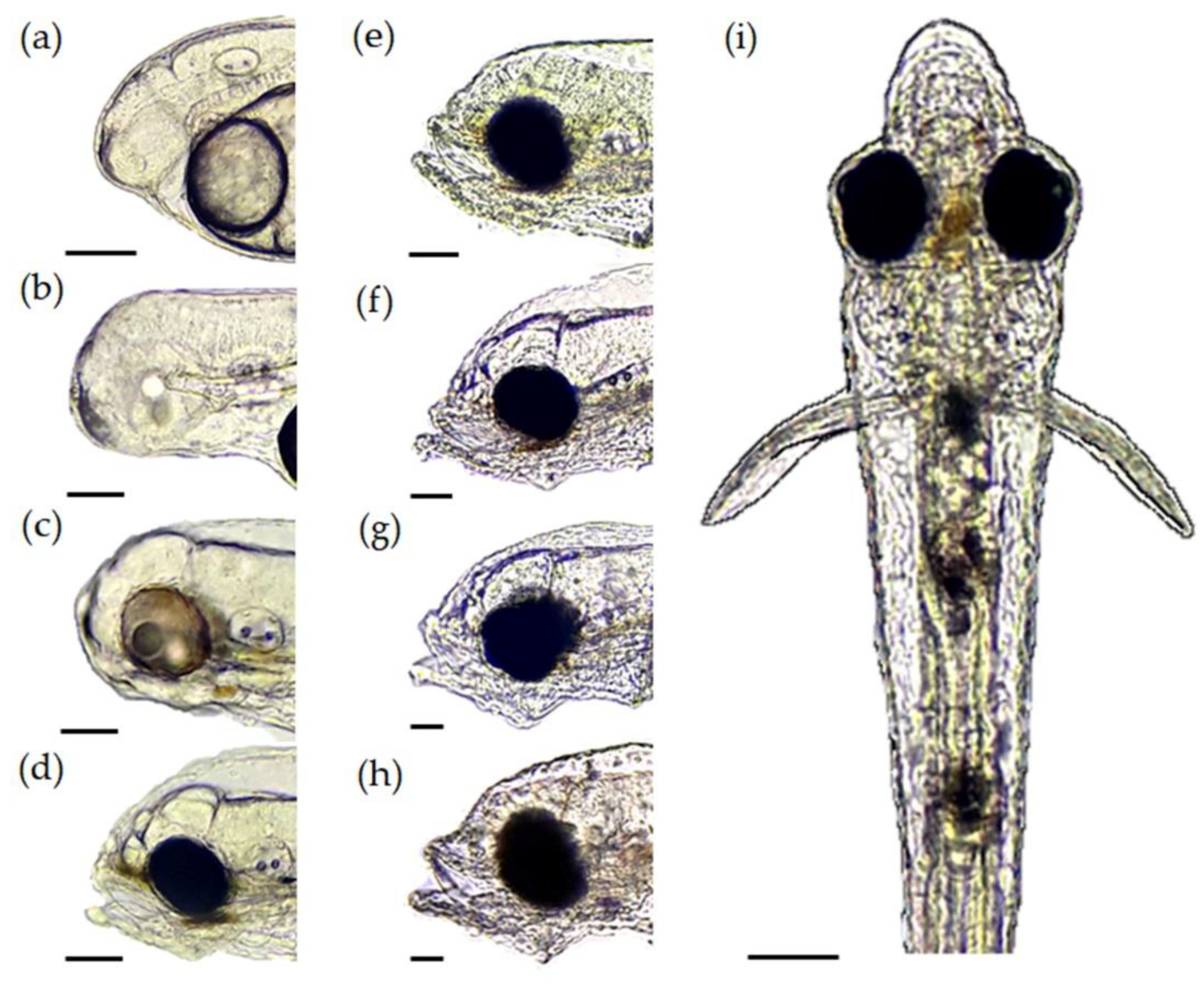
3.3. Mouth Morphometry
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braithwaite, V.; Salvanes, A.G. Aquaculture and restocking: Implications for conservation and welfare. Anim. Welf. 2010, 19, 139–149. [Google Scholar]
- Takeuchi, T. Progress on larval and juvenile nutrition to improve the quality and health of seawater fish: A review. Fish. Sci. 2014, 80, 389–403. [Google Scholar] [CrossRef]
- Yúfera, M.; Darias, M. The onset of feeding in marine fish larvae. Aquaculture 2007, 268, 53–63. [Google Scholar] [CrossRef]
- Blaxter, J.H.S.; Hempel, G. The Influence of Egg Size on Herring Larvae (Clupea harengus L.). ICES J. Mar. Sci. 1963, 28, 211–240. [Google Scholar] [CrossRef]
- Gisbert, E.; Conklin, D.B.; Piedrahita, R.H. Effects of delayed first feeding on the nutritional condition and mortality of California halibut larvae. J. Fish. Biol. 2004, 64, 116–132. [Google Scholar] [CrossRef]
- Galván-Quesada, S.; Doadrio, I.; Alda, F.; Perdices, A.; Reina, R.G.; García Varela, M.; Hernández, N.; Mendoza, A.C.; Bermingham, E.; Domínguez-Domínguez, O. Molecular Phylogeny and Biogeography of the Amphidromous Fish Genus Dormitator Gill 1861 (Teleostei: Eleotridae). PLoS ONE 2016, 11, e0153538. [Google Scholar] [CrossRef]
- Gonzalez-Martinez, A.; Lopez, M.; Molero, H.M.; Rodriguez, J.; González, M.; Barba, C.; García, A. Morphometric and Meristic Characterization of Native Chame Fish (Dormitator latifrons) in Ecuador Using Multivariate Analysis. Animals 2020, 10, 1805. [Google Scholar] [CrossRef]
- Chang, B.D. Tolerances to salinity and air exposure of Dormitator latifrons (Pisces: Eleotridae). Rev. Biol. Trop. 1984, 32, 155–157. [Google Scholar]
- Milton, D.A. Living in Two Worlds: Diadromous Fishes, and Factors Affecting Population Connectivity Between Tropical Rivers and Coasts. In Ecological Connectivity among Tropical Coastal Ecosystems; Nagelkerken, I., Ed.; Springer: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Chang, B.D.; Navas, W. Seasonal variations in growth, condition and gonads of Dormitator latifrons (Richardson) in the Chone River Basin, Ecuador. J. Fish Biol. 1984, 24, 637–648. [Google Scholar] [CrossRef]
- Flores-Nava, A.; Brown, A. Peces Nativos de Agua Dulce de América del SUR de Interés Para la Acuicultura: Una Síntesis del Estado de Desarrollo Tecnológico de su Cultivo. In Serie Acuicultura en Latinoamérica; FAO: Rome, Italy, 2010; Volume 1, pp. 63–70. [Google Scholar]
- Rodríguez-Montes de Oca, G.; Medina-Hernández, E.; Velázquez-Sandoval, J.; López-López, V.; Reyes, J.C.; Dabrowski, K.; Haws, M. Production of “‘Chame’” (Dormitator latifrons, Pisces: Eleotridae) larvae using GnRHa and LHRHa. Rev. Colomb. Cienc. Pecu. 2012, 25, 422–429. [Google Scholar]
- López-López, V.; Rodríguez-Montes de Oca, G.; Galaviz, M.; Reyes, J.C.; Medina-Hernández, E.; Dabrowski, K.; Haws, M.C. Descripción histológica comparativa del desarrollo del sistema digestivo y visual de larvas de chame Dormitator latifrons (Pisces: Eleotridae). Lat. Am. J. Aquat. Res. 2015, 43, 484–494. [Google Scholar] [CrossRef]
- Reyes, J.C.; Monteon, C.; Castillo, H.; Monroy Dosta, M.; Rodríguez-Montes de Oca, G. Population growth and protein and energy content of Proales similis (Rotifera: Monogononta) reared at different salinities. Turk. J. Fish. Aquat. Sci. 2017, 17, 767–775. [Google Scholar]
- Hepher, B. Ingestion, digestion and absorption of food. In Nutrition of Pond Fishes; Hepher, B., Ed.; Cambridge University Press: Cambrigde, UK, 1988; pp. 16–63. [Google Scholar]
- Arriaga, L.; Montaño, M.; Vásconez, J. Integrated management perspectives of the Bahía de Caráquez zone and Chone River estuary, Ecuador. Ocean Coast. Manag. 1999, 42, 229–241. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Shirota, A. Studies on the Mouth Size of Fish larvae. Bull. Jpn. Soc. Sci. Fish. 1970, 36, 353–368. [Google Scholar] [CrossRef]
- Ritz, C.; Jensen, S.M.; Gerhard, D.; Streibig, J.C. Dose-Response Analysis Using R, 1st ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2019; pp. 95–143. [Google Scholar]
- Farris, D.A. A Change in the Early Growth Rates of Four Larval Marine Fishes1. Limnol. Oceanogr. 1959, 4, 29–36. [Google Scholar] [CrossRef]
- Auty, E. Reproductive Behaviour and Early Development of the Empire Fish Hypseleotris compressus (Eleotridae). Mar. Freshw. Res. 1978, 29, 585–597. [Google Scholar] [CrossRef]
- Llewellyn, L. Spawning and development of the Flat-headed Gudgeon Philypnodon grandiceps (Krefft, 1864) Teleostei: Eleotridae). Aust. Zool. 2007, 34, 1–21. [Google Scholar] [CrossRef]
- Patimar, R.; Amouei, M.; Langroudi, S.M.M.-A. New data on the biology of Cobitis cf. satunini from the southern Caspian basin (northern Iran). Folia. Zool. 2011, 60, 308–314. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Litvak, M.K. Stock and parental effects on embryonic and early larval development of winter flounder Pseudopleuronectes americanus (Walbaum). J. Fish Biol. 2007, 70, 1070–1087. [Google Scholar] [CrossRef]
- Garcia, L.M.B.; Sayco, M.J.P.; Aya, F.A. Point-of-no-return and delayed feeding mortalities in first-feeding larvae of the silver therapon Leiopotherapon plumbeus (Kner) (Teleostei: Terapontidae). Aquac. Rep. 2020, 16, 100288. [Google Scholar] [CrossRef]
- Houde, E.D. Patterns and trends in larval-stage growth and mortality of teleost fish*. J. Fish Biol. 1997, 51, 52–83. [Google Scholar] [CrossRef]
- Tan, O.K.K.; Lam, T.J. Induced breeding and early development of the Marble Goby (Oxyeleotris marmorata, Blk.). Aquaculture 1973, 2, 411–423. [Google Scholar] [CrossRef]
- Dou, S.; Masuda, R.; Tanaka, M.; Tsukamoto, K. Feeding resumption, morphological changes and mortality during starvation in Japanese flounder larvae. J. Fish Biol. 2002, 60, 1363–1380. [Google Scholar] [CrossRef]
- Wegner, A.; Ostaszewska, T.; Rożek, W. The ontogenetic development of the digestive tract and accessory glands of sterlet (Acipenser ruthenus L.) larvae during endogenous feeding. Rev. Fish Biol. Fish. 2009, 19, 431. [Google Scholar] [CrossRef]
- Yúfera, M.; Pascual, E.; Polo, A.; Sarasquete, M.C. Effect of starvation on the feeding ability of gilthead seabream (Spams aurata L.) larvae at first feeding. J. Exp. Mar. Biol. Ecol. 1993, 169, 259–272. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Jaw Length (µm) | Maximum Mouth Opening (µm) | Width of the Oesophagus (µm) | |
|---|---|---|---|---|
| Upper | Lower | |||
| 0 | Closed mouth | Closed mouth | Closed mouth | Closed mouth |
| 24 | Closed mouth | Closed mouth | Closed mouth | Closed mouth |
| 48 | Closed mouth | Closed mouth | Closed mouth | Closed mouth |
| 72 | 43.22 ± 5.12 | 40.72 ± 4.96 | 61.13 ± 7.28 | 53.87 ± 6.65 |
| 96 | 95.83 ± 12.15 | 88.48 ± 13.34 | 135.53 ± 17.69 | 65.67 ± 4.04 |
| 120 | 147.45 ± 24.54 | 160.06 ± 23.79 | 208.54 ± 34.70 | 80.88 ± 8.30 |
| 144 | 148.70 ± 24.54 | 164.88 ± 26.26 | 210.29 ± 34.70 | 72.12 ± 8.42 |
| 168 | 153.56 ± 24.02 | 174.50 ± 25.14 | 217.17 ± 33.96 | 74.19 ± 8.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Mero, B.M.; Santana-Piñeros, A.M.; Muñoz-Chumo, L.G.; Cruz-Quintana, Y.; Gisbert, E. Yolk Absorption Rate and Mouth Development in Larvae of Dormitator latifrons (Perciformes: Eleotridae). Fishes 2022, 7, 375. https://doi.org/10.3390/fishes7060375
Reyes-Mero BM, Santana-Piñeros AM, Muñoz-Chumo LG, Cruz-Quintana Y, Gisbert E. Yolk Absorption Rate and Mouth Development in Larvae of Dormitator latifrons (Perciformes: Eleotridae). Fishes. 2022; 7(6):375. https://doi.org/10.3390/fishes7060375
Chicago/Turabian StyleReyes-Mero, Byron Manuel, Ana María Santana-Piñeros, Leonela Griselda Muñoz-Chumo, Yanis Cruz-Quintana, and Enric Gisbert. 2022. "Yolk Absorption Rate and Mouth Development in Larvae of Dormitator latifrons (Perciformes: Eleotridae)" Fishes 7, no. 6: 375. https://doi.org/10.3390/fishes7060375
APA StyleReyes-Mero, B. M., Santana-Piñeros, A. M., Muñoz-Chumo, L. G., Cruz-Quintana, Y., & Gisbert, E. (2022). Yolk Absorption Rate and Mouth Development in Larvae of Dormitator latifrons (Perciformes: Eleotridae). Fishes, 7(6), 375. https://doi.org/10.3390/fishes7060375