
Water Hardness Alters the Gene Expression Response and Copper Toxicity in Daphnia magna


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Test Media and Chemical Analysis
2.2. Acute Toxicity Test
2.3. Speciation Modeling of Cu in Different Hardness Conditions
2.4. Gene Expression Analysis
2.5. Data Process and Statistical Analysis
3. Results
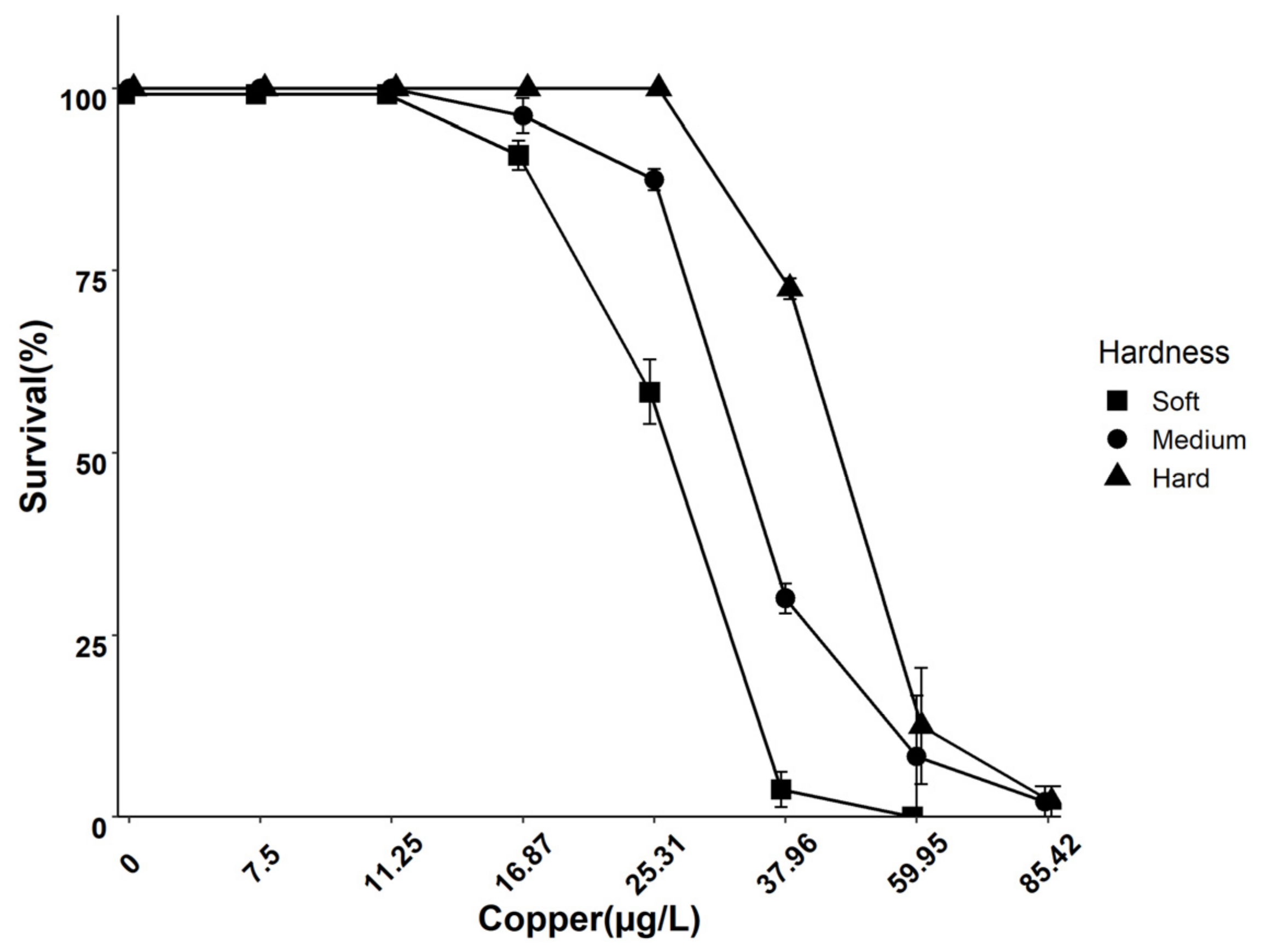
3.1. Effect of Water Hardness on Cu Speciation and 96 h Survival
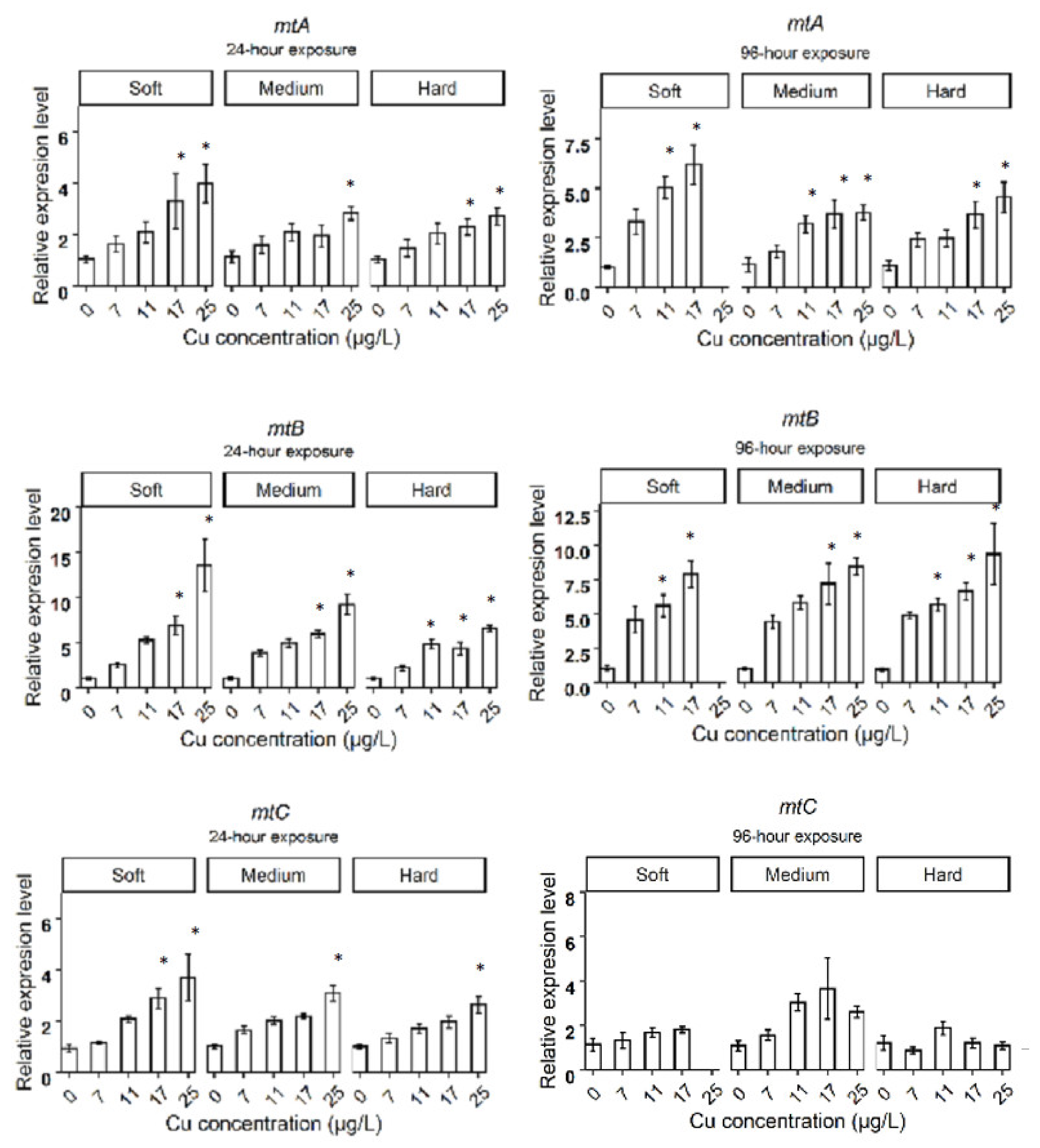
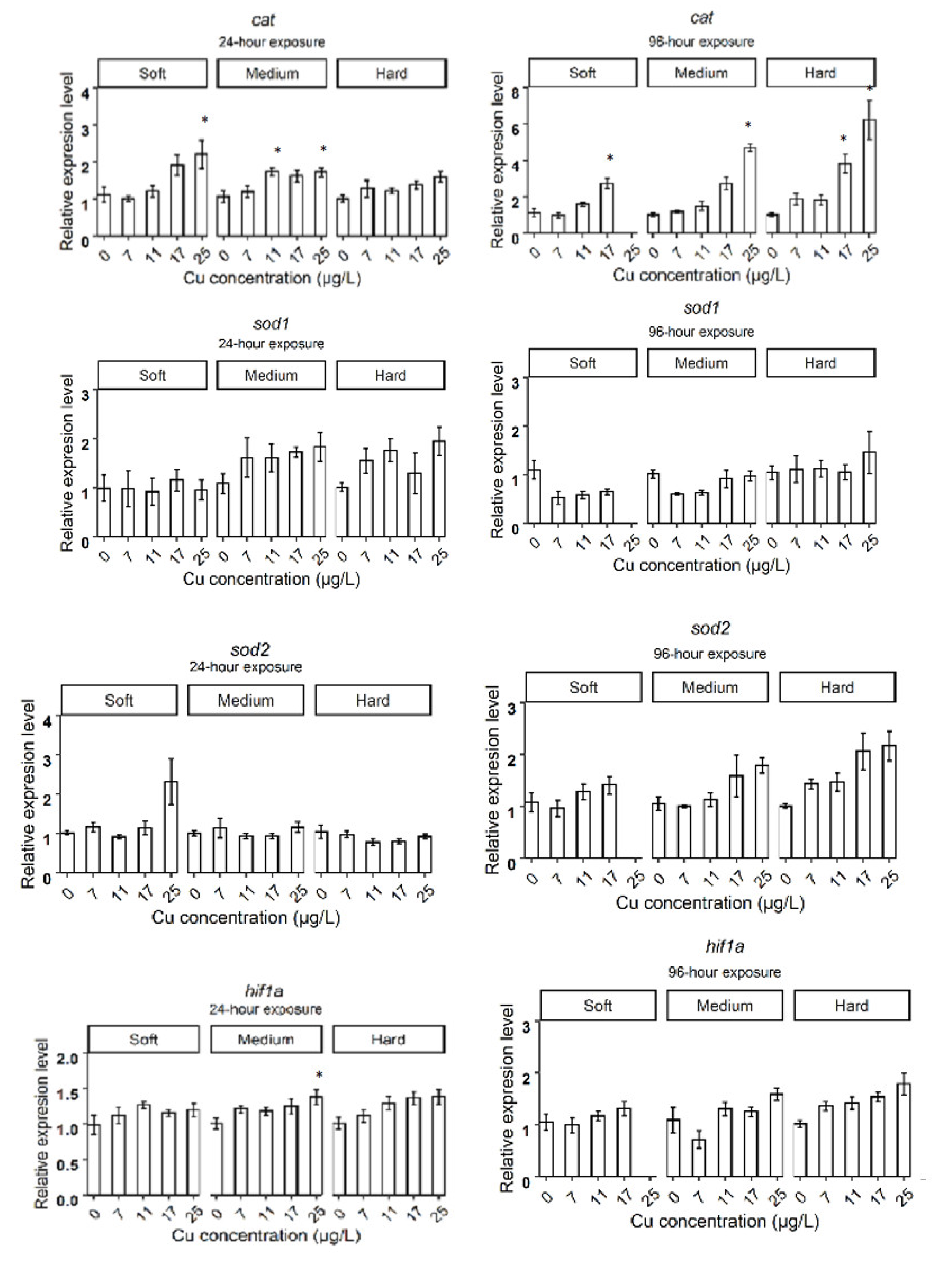
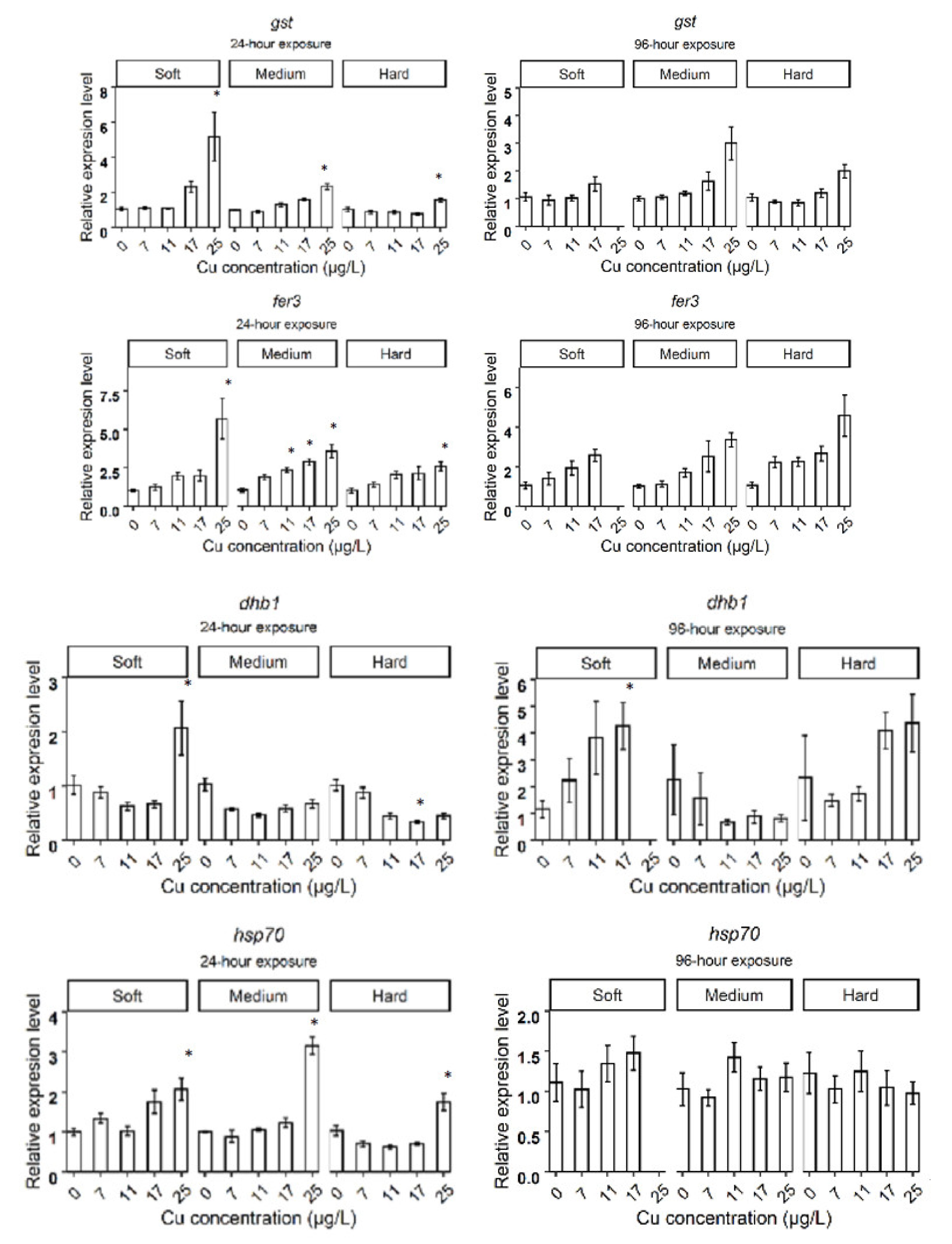
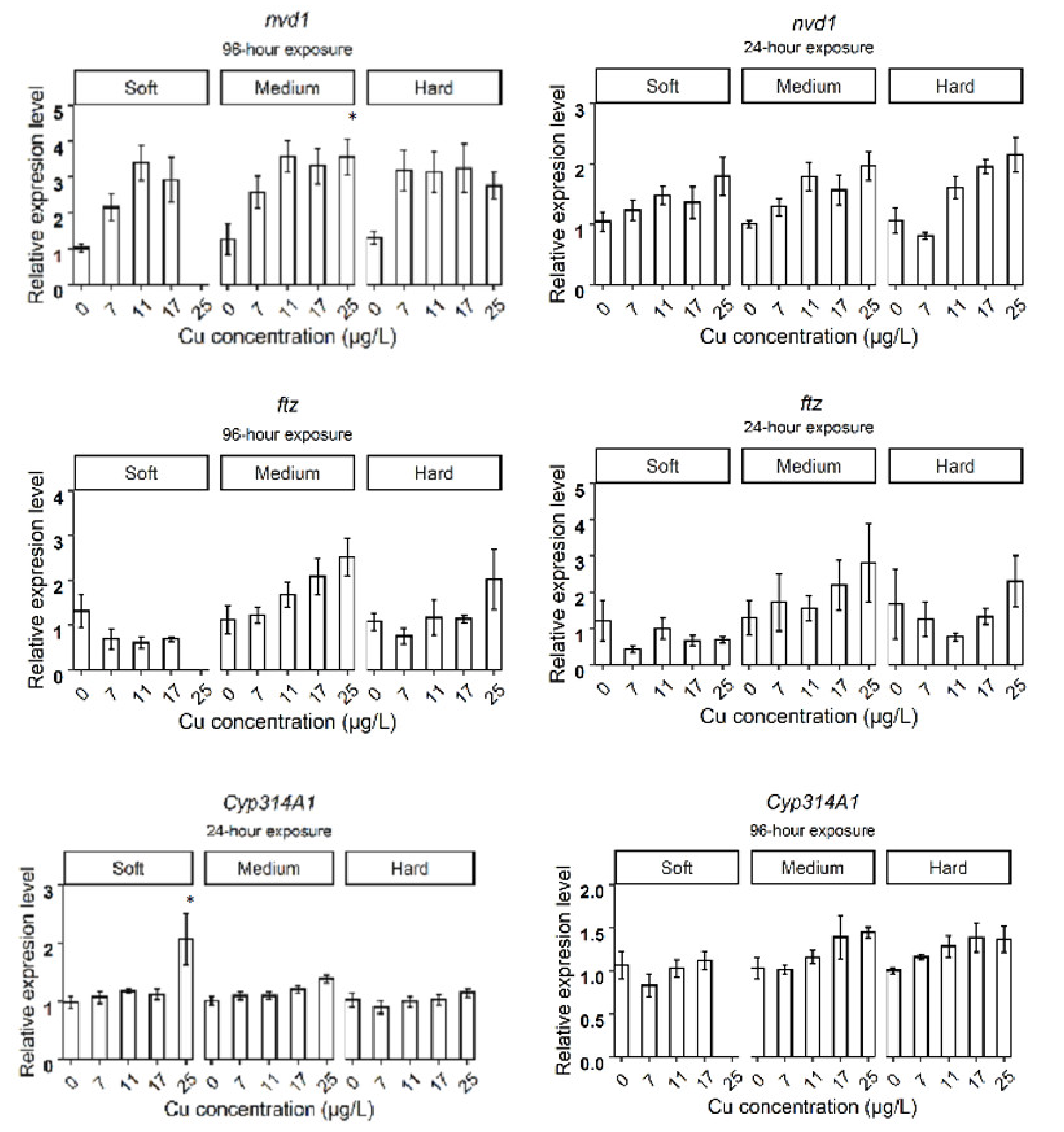
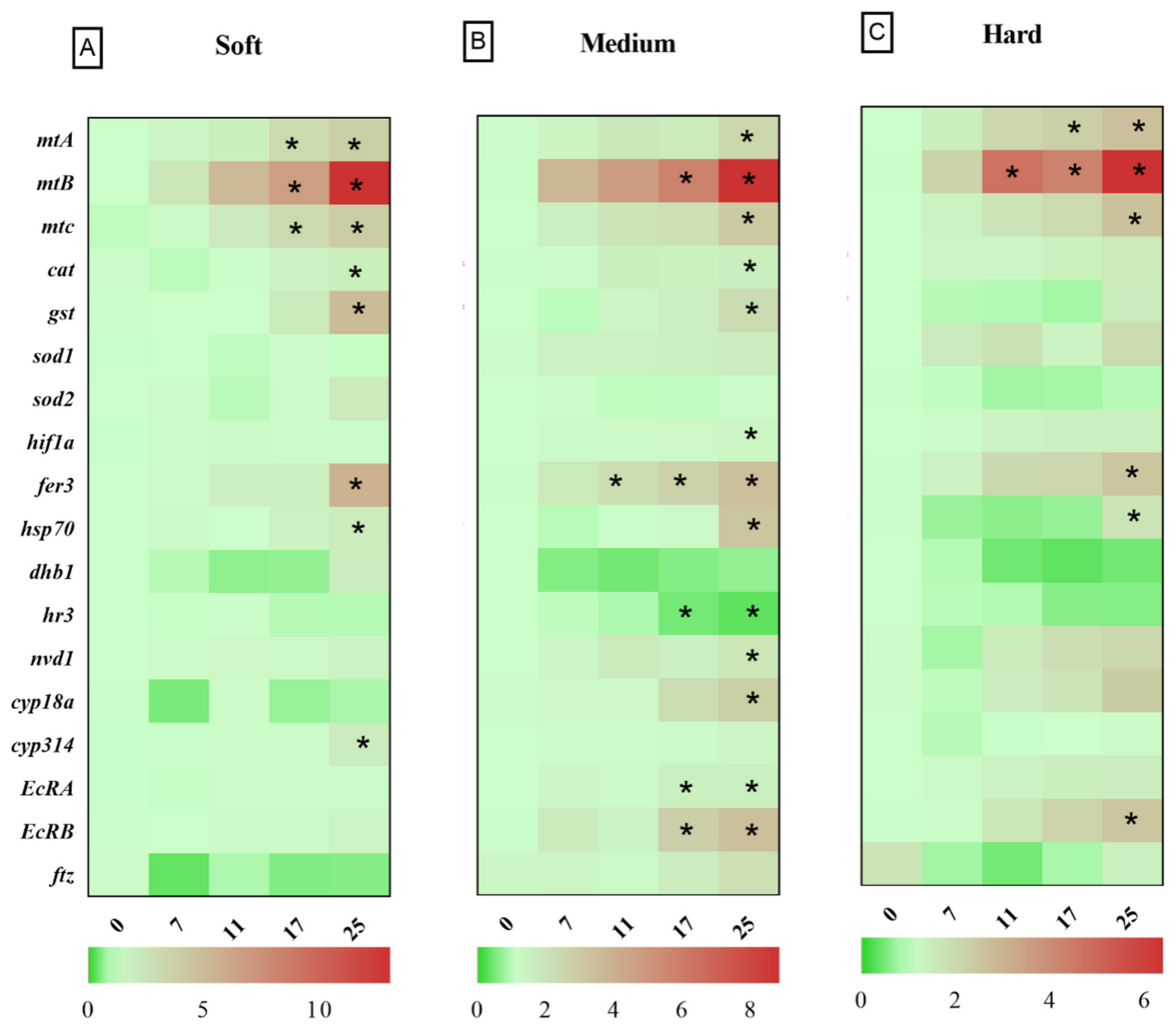
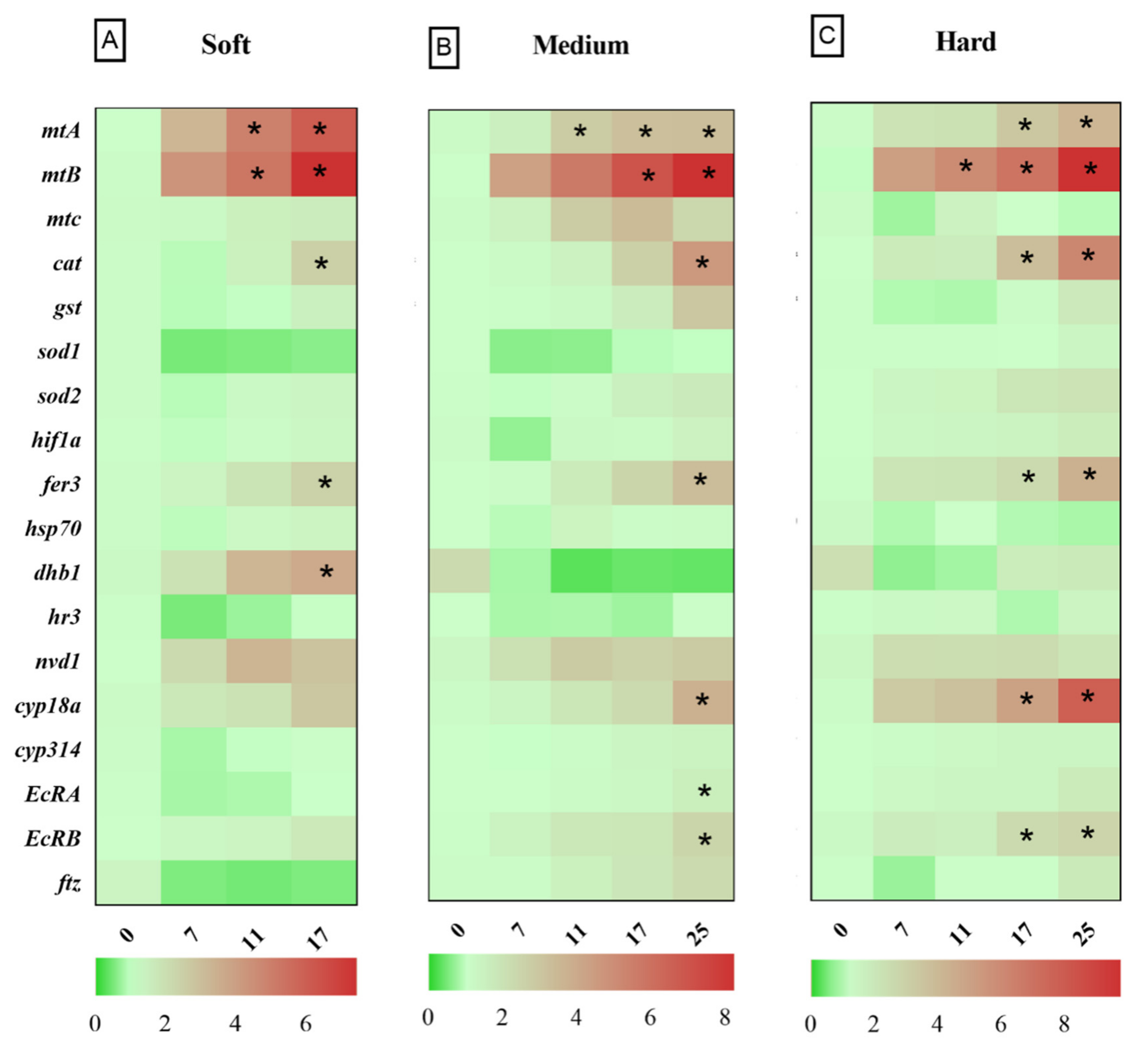
3.2. Relative Gene Expression in D. magna Exposed to Cu in Soft Water
3.3. Relative Gene Expression in D. magna Exposed to Cu in Medium Water
3.4. Relative Gene Expression in D. magna Exposed to Cu in Hard Water
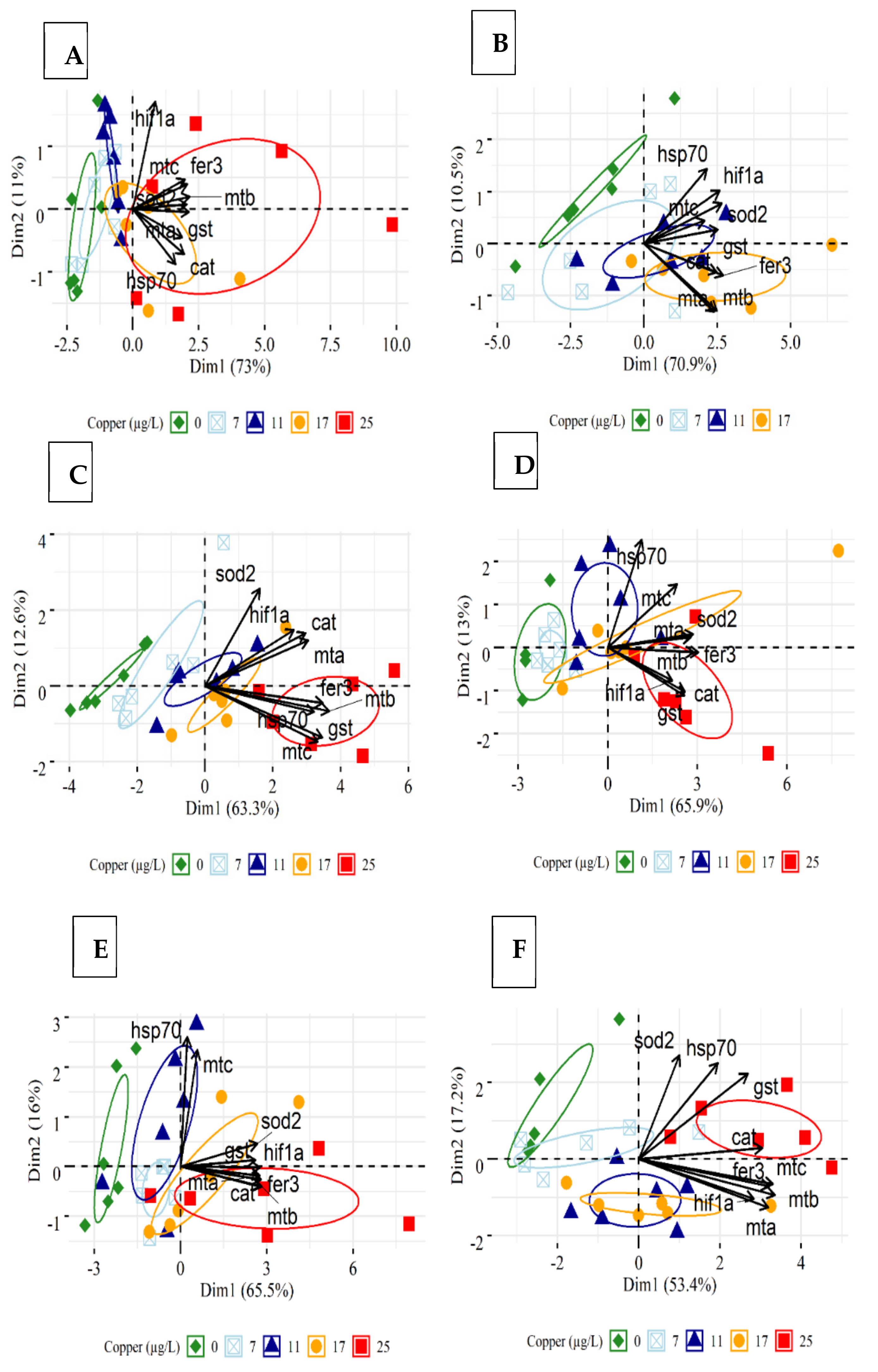
3.5. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Na | Mg | K | Ca | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hardness mg/L as CaCO3 | Replicates | Theoretical (μg/L) | Measured (μg/L) | Recovery (%) | Theoretical (μg/L) | Measured (μg/L) | Recovery (%) | Theoretical (μg/L) | Measured (μg/L) | Recovery (%) | Theoretical (μg/L) | Measured (μg/L) | Recovery (%) |
| 50 (Soft) | 1 | 18,530 | 18,060.5 | 97.5 | 5500 | 5463.2 | 99.3 | 3000 | 3516.4 | 117.2 | 11,000 | 9749.1 | 88.6 |
| 2 | 18,425.7 | 99.4 | 5542.6 | 100.8 | 3120.3 | 104.0 | 9807.9 | 89.2 | |||||
| 3 | 18,029.0 | 97.3 | 5418.9 | 98.5 | 2986.2 | 99.5 | 9431.4 | 85.7 | |||||
| 100 (Medium) | 1 | 18,530 | 18,966.4 | 102.4 | 11,000 | 11,578.0 | 105.3 | 3000 | 3427.2 | 114.2 | 22,000 | 19,957.4 | 90.7 |
| 2 | 18,894.6 | 102.0 | 11,268.6 | 102.4 | 3279.4 | 109.3 | 19,983.5 | 90.8 | |||||
| 3 | 18,616.9 | 100.5 | 11,308.3 | 102.8 | 3237.9 | 107.9 | 19,772.7 | 89.9 | |||||
| 200 (Hard) | 1 | 18,530 | 18,957.3 | 102.3 | 22,000 | 22,574.1 | 102.6 | 3000 | 3260.5 | 108.7 | 44,000 | 40,153.2 | 91.3 |
| 2 | 18,752.9 | 101.2 | 22,500.3 | 102.3 | 3126.0 | 104.2 | 39,374.6 | 89.5 | |||||
| 3 | 18,728.2 | 101.1 | 22,836.2 | 103.8 | 3224.4 | 107.5 | 39,467.6 | 89.7 | |||||
| Soft | Medium | Hard | |||||
|---|---|---|---|---|---|---|---|
| Theoretical (μg/L) | Measured (μg/L) | Recovery (%) | Measured (μg/L) | Recovery (%) | Measured (μg/L) | Recovery (%) | |
| Time 0 h | 0 | 1.2 | 1.4 | 1.1 | |||
| 7.5 | 6.8 | 90.53 | 6.6 | 87.91 | 7.1 | 94.89 | |
| 11.25 | 11.7 | 103.79 | 11.1 | 98.39 | 11.4 | 101.59 | |
| 16.87 | 17.3 | 102.56 | 17.3 | 102.65 | 17.2 | 102.06 | |
| 25.313 | 24.9 | 98.20 | 23.5 | 92.99 | 25.4 | 100.35 | |
| Time 24 h | Control | 0.8 | 3.1 | 1.7 | |||
| 7.5 | 7.6 | 101.04 | 7.1 | 94.76 | 7.7 | 102.94 | |
| 11.25 | 10.7 | 95.44 | 10.5 | 93.49 | 9.0 | 80 | |
| 16.87 | 16.1 | 95.70 | 18.0 | 106.69 | 14.7 | 87.10 | |
| 25.313 | 26.2 | 103.3 | 22.2 | 87.65 | 23.6 | 93.41 | |
| Gene Name | Gene Symbol | Forward (5′→3′) | Reverse (5′→3′) | |
|---|---|---|---|---|
| Actin | act | CCCCACGCCATCCTCCGTCT | GGTGGAGGCAGCAGCAGTGG | |
| Catalase | cat | TGGCGGAGAAAGCGGTTCAGC | GTGCGTGGTCTCTGGGCGAA | |
| Cu/zn-superoxide dismutase | sod1 | TCGTCGATACCGTCGTCTCACTTAGCG | ATTGCCCGTCTTGGTGCTGCCTTCGTT | |
| Cytochrome P450 18A1 family | cyp18A1 | TACCCGATCGTCGGTTACCT | GAGCGCCGTCAGCTCTTC | |
| Cytochrome P450 314 family (CYP314) | cyp314 | TCTTGGGTCGGCGTCTGGGA | TCGCGGGTGTCAACGCCTTC | |
| Ecdysone receptor A | EcRA | CAGGCACATCAACATCAACAA | GGCGACATGGAATCGACA | |
| Ecdysone receptor B | EcRB | CACCACAACCAACTGCATTTAC | CCATTAATGTCAAGATCCCACA | |
| Ferritin | fer3 | GCTGGTATCGCCAAGAACCTCAAA | TGGCGAAGAATTTCTGGATGCGGG | |
| Glutathione-s-transferase | gst | TCAGGCTGGTGTTGAGTTTG | GAGCAAGCATTTGTCCATCA | |
| Heat shock protein 70 | hsp70 | CGACGGCGGGAGATACGCAC | CCACGGAAAAGGTCGGCGCA | |
| HR3 nuclear hormone receptor | hr3 | AGTCATCACCTGCGAGGGC | GAACTTTGCGACCGCCG | |
| Hypoxia Inducible Factor 1 Alpha | hif1α | GGTCCAGACCCAAGCAGCCAGGC | GTCCAGGAGCAGCAGCCAGC | |
| Metallothionein A | mtA | GAGCGCCATGCCAAAATCCC | TCGTCGTTGTAAAATCCGCCT | |
| Metallothionein B | mtB | TGGAACCGAATGCAAATGCG | CGGACTTGCATGGACAACTG | |
| Metallothionein C | mtC | AAAGTGTGCCCTCGTTGTCA | CTTACAGTCGTCCCCACACG | |
| Zn-superoxide dismutase | sod2 | CCTAATGGAGGTGAACCAGAGGGAGAG | CCTAGCCAACCCCATCCTGAACCTTGA | |
| Hemoglobin 1 | dhb1 | GCTGGTATCGCCAAGAACCTCAAA | TGGCGAAGAATTTCTGGATGCGGG | |
| Fushi tarazu factor-1 | ftz | GTTGCACAATCACCTGCCTG | TAACGTCGTGAAGGGGTTCG | |
| Gene name | Gene symbol | Forward (5′→3′) | Reverse (5′→3′) | Reference |
| Neverland 1 | ndv1 | AGCACAAGGCGGGAAGAGT | GCTTCCCATTTCACCTTCCA | Sumiya et al., 2016 [60] |
| Species Name (% of total concentration) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Hardness | Copper (µg/L) | Cu+2 | CuOH+ | Cu(OH)2 (aq) | Cu2(OH)2+2 | CuCl+ | CuSO4 (aq) | CuCO3 (aq) | CuHCO3+ | Cu(CO3)2−2 |
| Soft | 7 | 11.76 | 11.89 | 0.73 | 0.01 | 0.01 | 0.36 | 74.48 | 0.43 | 0.34 |
| Medium | 12.47 | 12.05 | 0.73 | 0.01 | 0.02 | 0.65 | 73.29 | 0.43 | 0.34 | |
| Hard | 12.18 | 12.48 | 0.83 | 0.01 | 0.04 | 1.02 | 72.65 | 0.39 | 0.41 | |
| Soft | 11 | 11.76 | 11.89 | 0.73 | 0.02 | 0.01 | 0.36 | 74.47 | 0.43 | 0.34 |
| Medium | 12.47 | 12.05 | 0.73 | 0.02 | 0.02 | 0.65 | 73.28 | 0.43 | 0.34 | |
| Hard | 12.18 | 12.48 | 0.83 | 0.02 | 0.04 | 1.02 | 72.64 | 0.39 | 0.41 | |
| Soft | 17 | 11.75 | 11.89 | 0.73 | 0.03 | 0.01 | 0.36 | 74.46 | 0.43 | 0.34 |
| Medium | 12.47 | 12.05 | 0.73 | 0.03 | 0.02 | 0.65 | 73.28 | 0.43 | 0.34 | |
| Hard | 12.18 | 12.48 | 0.83 | 0.03 | 0.04 | 1.02 | 72.64 | 0.39 | 0.41 | |
| Soft | 25 | 11.75 | 11.89 | 0.73 | 0.04 | 0.01 | 0.36 | 74.45 | 0.43 | 0.34 |
| Medium | 12.47 | 12.05 | 0.73 | 0.04 | 0.02 | 0.65 | 73.26 | 0.43 | 0.34 | |
| Hard | 12.18 | 12.48 | 0.83 | 0.05 | 0.04 | 1.02 | 72.62 | 0.39 | 0.41 | |
Appendix B

References
- Puig, S.; Thiele, D.J. Molecular mechanisms of copper uptake and distribution. Curr. Opin. Chem. Biol. 2002, 6, 171–180. [Google Scholar] [CrossRef]
- De Oliveira-Filho, E.C.; Lopes, R.M.; Paumgartten, F.J.R. Comparative study on the susceptibility of freshwater species to copper-based pesticides. Chemosphere 2004, 56, 369–374. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Collaboration between the World Health Organization and the National Institute of Environmental Health Sciences: Highlights from 30 Years of Partnership; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Stern, B.R. Essentiality and toxicity in copper health risk assessment: Overview, update and regulatory considerations. J. Toxicol. Environ. 2010, 73, 114–127. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency (US EPA). Aquatic Life Ambient Freshwater Quality Criteria-Copper. 2007 Revision. EPA-822-R-07-001. Available online: https://www.federalregister.gov/documents/2007/02/22/E7-3007/aquatic-life-ambient-freshwater-quality-criteria-copper-2007-revision (accessed on 10 June 2021).
- Fu, Z.; Wu, F.; Chen, L.; Xu, B.; Feng, C.; Bai, Y.; Liao, H.; Sun, S.; Giesy, J.P.; Guo, W. Copper and zinc, but not other priority toxic metals, pose risks to native aquatic species in a large urban lake in Eastern China. Environ. Pollut. 2016, 219, 1069–1076. [Google Scholar] [CrossRef]
- Roux, D.; Jooste, S.; MacKay, H.M. Substance-specific water quality criteria for the protection of South African freshwater ecosystems: Methods for derivation and initial results for some inorganic toxic substances. S. Afr. J. Sci. 1996, 92, 198–206. [Google Scholar]
- Borgmann, U.; Couillard, Y.; Doyle, P.; Dixon, D.G. Toxicity of sixty-three metals and metalloids to Hyalella azteca at two levels of water hardness. Environ. Toxicol. 2005, 24, 641–652. [Google Scholar] [CrossRef]
- Rai, L.; Gaur, J.; Kumar, H.D. Phycology and heavy-metal pollution. Biol. Rev. 1981, 56, 99–151. [Google Scholar] [CrossRef]
- Kramer, K.J.; Jak, R.G.; van Hattum, B.; Hooftman, R.N.; Zwolsman, J.J.G. Copper toxicity in relation to surface water-dissolved organic matter: Biological effects to Daphnia magna. Environ. Toxicol. 2004, 23, 2971–2980. [Google Scholar] [CrossRef]
- Andrew, R.; Biesinger, K.; Glass, G.E. Effects of inorganic complexing on the toxicity of copper to Daphnia magna. Water Res. 1977, 11, 309–315. [Google Scholar] [CrossRef]
- De Schamphelaere, K.A.C.; Janssen, C.R. Effects of dissolved organic carbon concentration and source, pH, and water hardness on chronic toxicity of copper to Daphnia magna. Environ. Sci. Technol. 2004, 23, 1115–1122. [Google Scholar] [CrossRef]
- Park, E.J.; Jo, H.J.; Jung, J. Combined effects of pH, hardness and dissolved organic carbon on acute metal toxicity to Daphnia magna. Ind. Eng. Chem. 2009, 15, 82–85. [Google Scholar] [CrossRef]
- Okamoto, A.; Yamamuro, M.; Tatarazako, N. Acute toxicity of 50 metals to Daphnia magna. J. Appl. Toxicol. 2015, 35, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.H.; Arbildua, J.; Villavicencio, G.; Mejías, R.; Urrestarazu, P.; Jiménez, M. Copper acute and chronic toxicity to D. magna: Sensitivity at three different hardness at pH 6.3 (MES buffered) in the presence of 2 mg/L DOC. In Chilean Mining and Metallurgy Research Center, Laboratory of Ecotoxicology and Chemistry of Trace Metals; Centro de Investigación Minera y Metalúrgica: Santiago, Chile, 2012; Available online: http://hdl.handle.net/1854/LU-5902885 (accessed on 10 June 2021).
- Bui, T.-K.L.; Do-Hong, L.C.; Dao, T.-S.; Hoang, T.C. Copper toxicity and the influence of water quality of Dongnai River and Mekong River waters on copper bioavailability and toxicity to three tropical species. Chemosphere 2016, 144, 872–878. [Google Scholar] [CrossRef] [PubMed]
- De Schamphelaere, K.A.; Janssen, C.R. A biotic ligand model predicting acute copper toxicity for Daphnia magna: The effects of calcium, magnesium, sodium, potassium, and pH. Environ. Sci. Technol. 2002, 36, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.M.; Wood, C.M.; McClelland, G.B. Water chemistry alters gene expression and physiological end points of chronic waterborne copper exposure in zebrafish, Danio rerio. Environ. Sci. Technol. 2010, 44, 2156–2162. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, version 4.0.2; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 10 June 2021).
- RStudio Team. RStudio: Integrated Development for R, version 1.2.5042; PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com (accessed on 10 June 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kozlova, T.; Wood, C.M.; McGeer, J.C. The effect of water chemistry on the acute toxicity of nickel to the cladoceran Daphnia pulex and the development of a biotic ligand model. Aquat. Toxicol. 2009, 91, 221–228. [Google Scholar] [CrossRef]
- De Paiva Magalhães, D.; da Costa Marques, M.R.; Baptista, D.F.; Buss, D.F. Metal bioavailability and toxicity in freshwaters. Environ. Chem. Lett. 2015, 13, 69–87. [Google Scholar] [CrossRef]
- McWilliams, P.G.; Potts, W.T.W. The effects of pH and calcium concentrations on gill potentials in the brown trout, Salmo trutta. Comp. Biochem. Physiol. B Biochem. 1978, 126, 277–286. [Google Scholar] [CrossRef]
- Markich, S.; Batley, G.; Stauber, J.; Rogers, N.; Apte, S.; Hyne, R.; Bowles, K.; Wilde, K.; Creighton, N.M. Hardness corrections for copper are inappropriate for protecting sensitive freshwater biota. Chemosphere 2005, 60, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hessen, D.O.; Alstad, N.E.; Skardal, L. Calcium limitation in Daphnia magna. J. Plankton Res. 2000, 22, 553–568. [Google Scholar] [CrossRef]
- Jesus, F.T.; Martins, C.; Nogueira, A.J.A. Changes in life-history parameters of Daphnia longispina (Cladocera, Crustacea) as a function of water chemistry. J. Limnol. 2014, 73, 340–349. [Google Scholar] [CrossRef]
- Poynton, H.C.; Loguinov, A.V.; Varshavsky, J.R.; Chan, S.; Perkins, E.J.; Vulpe, C.D. Gene expression profiling in Daphnia magna part I: Concentration-dependent profiles provide support for the no observed transcriptional effect level. Environ. Sci. Technol. 2008, 42, 6250–6256. [Google Scholar] [CrossRef] [PubMed]
- Asselman, J.; Shaw, J.R.; Glaholt, S.P.; Colbourne, J.K.; De Schamphelaere, K.A.C. Transcription patterns of genes encoding four metallothionein homologs in Daphnia pulex exposed to copper and cadmium are time-and homolog-dependent. Aquat. Toxicol. 2013, 142, 422–430. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, G.; Ngo, T.T.H.; Van Campenhout, K.; Blust, R. Differential metallothionein induction patterns in three freshwater fish during sublethal copper exposure. Aquat. Toxicol. 2003, 65, 413–424. [Google Scholar] [CrossRef]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Olsson, P.E.; Kling, P.; Hogstrand, C. Mechanisms of heavy metal accumulation and toxicity in fish. In Metal Metabolism in Aquatic Environments; Bebianno, W.J., Langston, M.J., Eds.; Chapman and Hall: London, UK, 1998; pp. 321–350. [Google Scholar] [CrossRef]
- Mayer, G.D.; Leach, A.; Kling, P.; Olsson, P.E.; Hogstrand, C. Activation of the rainbow trout metallothionein-A promoter by silver and zinc. Comp. Biochem. Physiol. B Biochem. 2003, 134, 181–188. [Google Scholar] [CrossRef]
- Amiard, J.-C.; Amiard-Triquet, C.; Barka, S.; Pellerin, J.; Rainbow, P.S. Metallothioneins in aquatic invertebrates: Their role in metal detoxification and their use as biomarkers. Aquat. Toxicol. 2006, 76, 160–202. [Google Scholar] [CrossRef]
- Thornalley, P.J.; Vašák, M. Possible role for metallothionein in protection against radiation-induced oxidative stress. Kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. Biochim. Biophys. Acta 1985, 827, 36–44. [Google Scholar] [CrossRef]
- Iszard, M.B.; Liu, J.; Klaassen, C.D. Effect of several metallothionein inducers on oxidative stress defense mechanisms in rats. Toxicology 1995, 104, 25–33. [Google Scholar] [CrossRef]
- Sun, X.; Zhou, Z.; Kang, J. Attenuation of doxorubicin chronic toxicity in metallothionein-overexpressing transgenic mouse heart. Cancer Res. 2001, 61, 3382–3387. [Google Scholar] [PubMed]
- Kling, P.; Olsson, P.E. Metallothionein: Structure and regulation. In Biochemistry and Molecular Biology of Fishes; Mommsen, T.W.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 6, pp. 289–302. [Google Scholar] [CrossRef]
- Mosleh, Y.Y.; Paris-Palacios, S.; Ahmed, M.T.; Mahmoud, F.; Osman, M.; Biagianti-Risbourg, S. Effects of chitosan on oxidative stress and metallothioneins in aquatic worm Tubifex tubifex (Oligochaeta, Tubificidae). Chemosphere 2007, 67, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Barata, C.; Varo, I.; Navarro, J.C.; Arun, S.; Porte, C. Antioxidant enzyme activities and lipid peroxidation in the freshwater cladoceran Daphnia magna exposed to redox cycling compounds. Comp. Biochem. Physiol. C Toxicol. 2005, 140, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Chain, F.J.; Finlayson, S.; Crease, T.; Cristescu, M. Variation in transcriptional responses to copper exposure across Daphnia pulex lineages. Aquat. Toxicol. 2019, 210, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Pulford, D.J. The glutathione S-transferase supergene family: Regulation of GST and the contribution of the lsoenzymes to cancer chemoprotection and drug resistance part I. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–520. [Google Scholar] [CrossRef] [PubMed]
- Poynton, H.C.; Varshavsky, J.R.; Chang, B.; Cavigiolio, G.; Chan, S.; Holman, P.S.; Loguinov, A.V.; Bauer, D.J.; Komachi, K.; Theil, E.C.; et al. Daphnia magna ecotoxicogenomics provides mechanistic insights into metal toxicity. Environ. Sci. Technol. 2007, 41, 1044–1050. [Google Scholar] [CrossRef]
- Tsuji, Y.; Ayaki, H.; Whitman, S.P.; Morrow, C.S.; Torti, S.V.; Torti, F.M. Coordinate transcriptional and translational regulation of ferritin in response to oxidative stress. Mol. Cell. Biol. 2000, 20, 5818–5827. [Google Scholar] [CrossRef]
- Padmini, E.; Geetha, B.V. Impact of season on liver mitochondrial oxidative stress and the expression of HSP70 in grey mullets from contaminated estuary. Ecotoxicology 2009, 18, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Efremova, S.M.; Margulis, B.A.; Guzhova, I.V.; Itskovich, V.B.; Lauenroth, S.; Müller, W.E.; Schröder, H.C. Heat shock protein Hsp70 expression and DNA damage in Baikalian sponges exposed to model pollutants and wastewater from Baikalsk Pulp and Paper Plant. Aquat. Toxicol. 2002, 57, 267–280. [Google Scholar] [CrossRef]
- Connon, R.; Hooper, H.L.; Sibly, R.M.; Lim, F.L.; Heckmann, L.H.; Moore, D.J.; Watanabe, H.; Soetaert, A.; Cook, K.; Maund, S.J.; et al. Linking molecular and population stress responses in Daphnia magna exposed to cadmium. Environ. Sci. Technol. 2008, 42, 2181–2188. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, S.; Mini, J.; Munuswamy, N. Effects of heavy metals on antioxidants and expression of HSP70 in different tissues of Milk fish (Chanos chanos) of Kaattuppalli Island, Chennai, India. Ecotoxicol. Environ. Saf. 2013, 98, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.S.; Kim, P.J.; Won, E.J.; Lee, Y.M. Response of antioxidant enzymes to Cd and Pb exposure in water flea Daphnia magna: Differential metal and age—Specific patterns. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 209, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Cowgill, U.; Emmel, H.; Hopkins, D.; Applegath, S.; Takahashi, I.T. The influence of water on reproductive success and chemical composition of laboratory reared populations of Daphnia magna. Water Res. 1986, 20, 317–323. [Google Scholar] [CrossRef]
- Alstad, N.E.; Skardal, L.; Hessen, D.O. The effect of calcium concentration on the calcification of Daphnia magna. Limnol. Oceanogr. 1999, 44, 2011–2017. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Mousa, M.A.; Ahmad, M.H.; Sakr, S.F.M. The use of calcium pre-exposure as a protective agent against environmental copper toxicity for juvenile Nile tilapia, Oreochromis niloticus (L.). Aquaculture 2007, 264, 236–246. [Google Scholar] [CrossRef]
- Li, H.S.; Zhou, Y.N.; Li, L.; Li, S.F.; Long, D.; Chen, X.L.; Zhang, J.B.; Feng, L.; Li, Y.P. HIF-1α protects against oxidative stress by directly targeting mitochondria. Redox Biol. 2019, 25, 101109. [Google Scholar] [CrossRef]
- Chen, S.; McKinney, G.J.; Nichols, K.M.; Colbourne, J.K.; Sepulveda, M.S. Novel cadmium responsive microRNAs in Daphnia pulex. Environ. Sci. Technol. 2015, 49, 14605–14613. [Google Scholar] [CrossRef]
- Mottet, D.; Michel, G.; Renard, P.; Ninane, N.; Raes, M.; Michiels, C. Role of ERK and calcium in the hypoxia-induced activation of HIF-1. J. Cell. Physiol. 2002, 94, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Sumiya, E.; Ogino, Y.; Toyota, K.; Miyakawa, H.; Miyagawa, S.; Iguchi, T. Neverland regulates embryonic moltings through the regulation of ecdysteroid synthesis in the water flea Daphnia magna, and may thus act as a target for chemical disruption of molting. J. Appl. Toxicol. 2016, 36, 1476–1485. [Google Scholar] [CrossRef] [PubMed]
| 24 h | 96 h | |||||
|---|---|---|---|---|---|---|
| Soft | Medium | Hard | Soft | Medium | Hard | |
| mtA | 0.60 * | 0.59 * | 0.62 * | 0.79 * | 0.64 * | 0.70 * |
| mtB | 0.79 * | 0.89 * | 0.83 * | 0.81 * | 0.78 * | 0.72 * |
| mtC | 0.70 * | 0.85 * | 0.74 * | 0.40 | 0.35 | 0.04 |
| cat | 0.60 * | 0.55 * | 0.49 * | 0.71 * | 0.89 * | 0.75 * |
| gst | 0.66 * | 0.85 * | 0.49 * | 0.34 | 0.65 * | 0.58 * |
| sod1 | 0.01 | 0.35 | 0.33 | −0.45 | 0.14 | 0.26 |
| sod2 | 0.50 * | 0.09 | −0.21 | 0.33 | 0.52 * | 0.62 * |
| hif1a | 0.26 | 0.53 * | 0.53 * | 0.31 | 0.48 * | 0.64 * |
| fer3 | 0.68 * | 0.84 * | 0.67 * | 0.63 * | 0.67 * | 0.65 * |
| hsp70 | 0.61 * | 0.78 * | 0.49 * | 0.28 | 0.15 | −0.15 |
| dhb1 | 0.41 | −0.39 | −0.69 * | 0.50 | −0.28 | 0.35 |
| hr3 | −0.17 | −0.82 * | −0.64 * | −0.03 | 0.07 | 0.20 |
| nvd1 | 0.42 | 0.54 * | 0.68 * | 0.59 * | 0.52 * | 0.33 |
| cyp18a | −0.09 | 0.68 * | 0.56 * | 0.77 * | 0.71 * | 0.73 * |
| cyp314 | 0.54 * | 0.62 * | 0.26 | 0.09 | 0.47 * | 0.48 * |
| ecra | 0.22 | 0.72 * | 0.54 * | −0.03 | 0.63 * | 0.56 * |
| ecrb | 0.46 | 0.81 * | 0.75 * | 0.56 * | 0.64 * | 0.70 * |
| ftz | −0.18 | 0.30 | 0.19 | −0.40 | 0.57 * | 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chupani, L.; Sjöberg, V.; Jass, J.; Olsson, P.-E. Water Hardness Alters the Gene Expression Response and Copper Toxicity in Daphnia magna. Fishes 2022, 7, 248. https://doi.org/10.3390/fishes7050248
Chupani L, Sjöberg V, Jass J, Olsson P-E. Water Hardness Alters the Gene Expression Response and Copper Toxicity in Daphnia magna. Fishes. 2022; 7(5):248. https://doi.org/10.3390/fishes7050248
Chicago/Turabian StyleChupani, Latifeh, Viktor Sjöberg, Jana Jass, and Per-Erik Olsson. 2022. "Water Hardness Alters the Gene Expression Response and Copper Toxicity in Daphnia magna" Fishes 7, no. 5: 248. https://doi.org/10.3390/fishes7050248
APA StyleChupani, L., Sjöberg, V., Jass, J., & Olsson, P.-E. (2022). Water Hardness Alters the Gene Expression Response and Copper Toxicity in Daphnia magna. Fishes, 7(5), 248. https://doi.org/10.3390/fishes7050248