
Molecular Cloning and Characterization of tlr1, tlr5s, tlr5m, and tlr14 Genes in the Swamp Eel, Monopterus albus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Acquisition of cDNA Sequences
2.3. Sequence Analysis
2.4. Phylogenetic Analysis
2.5. Syntenic Analysis of TLR Genes in Swamp Eel and Other Teleost Fish
2.6. Tissue Collections, Total RNA Extraction, and Reverse Transcription
2.7. Gene Clone
2.8. Quantitative PCR (qPCR) and Statistical Analysis
2.9. Bacterial Challenge Experiment
3. Results
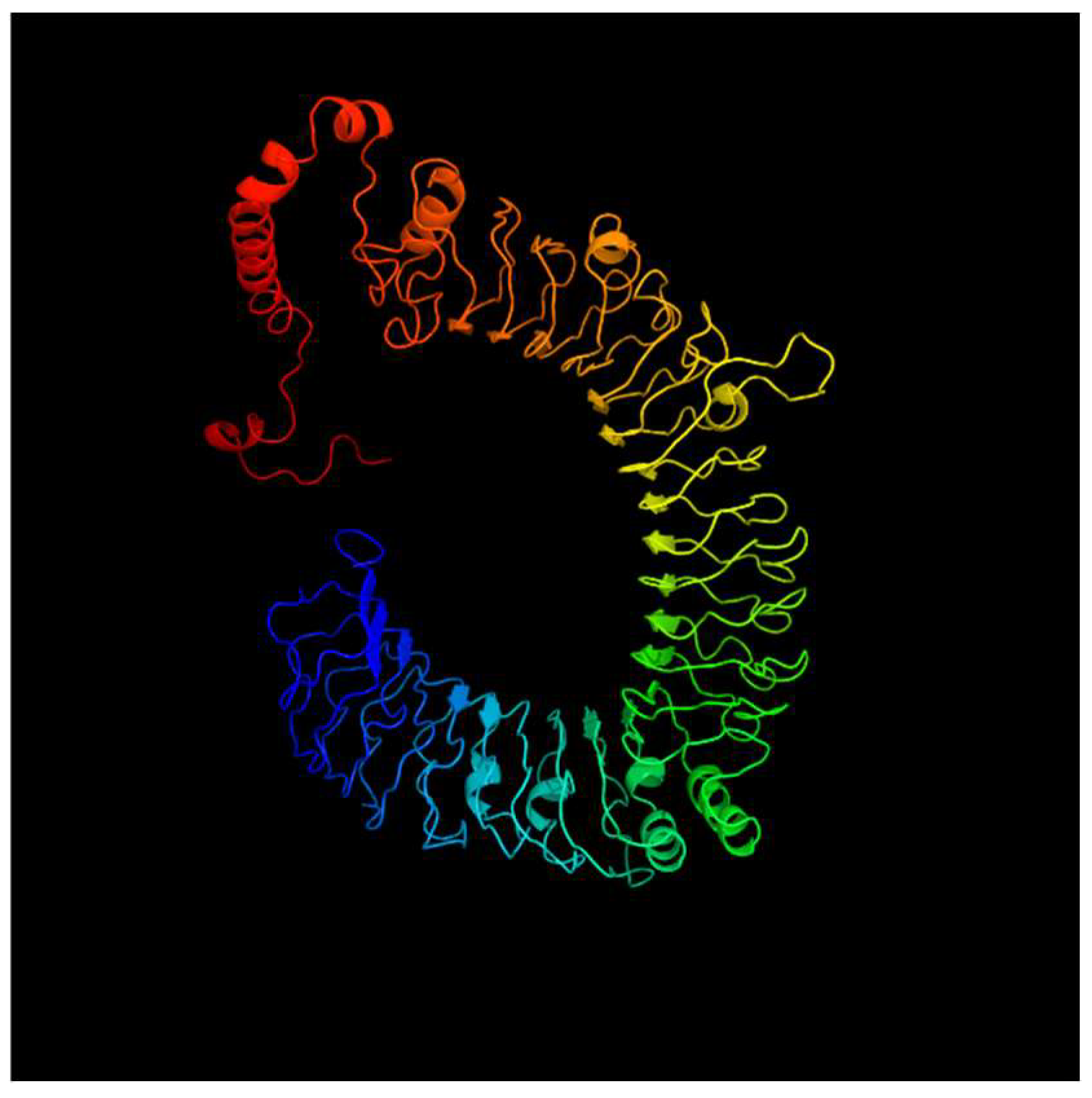
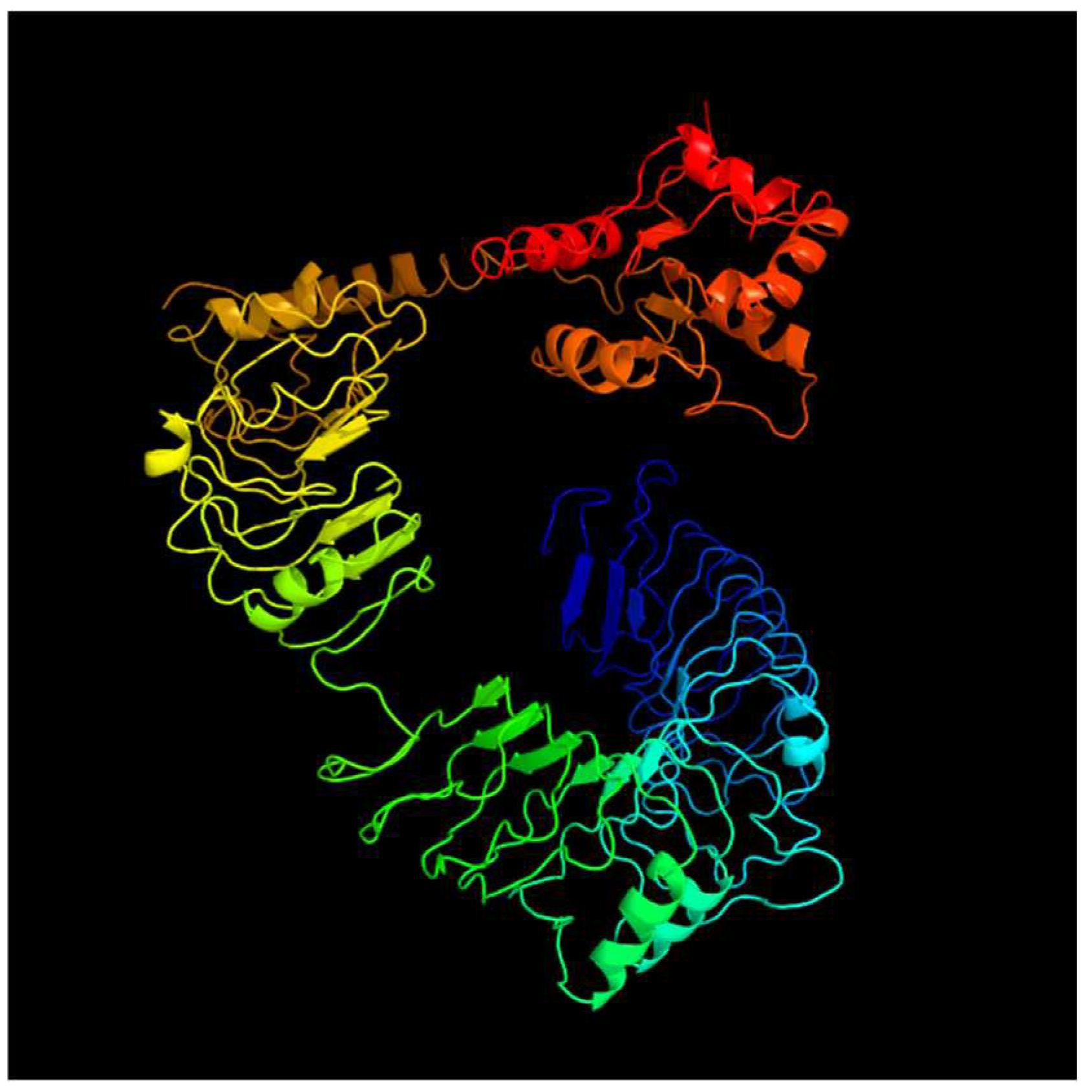
3.1. Identification and Analysis of TLR Genes
3.2. Phylogeny and Collinearity Analysis
3.3. Collinearity Analysis
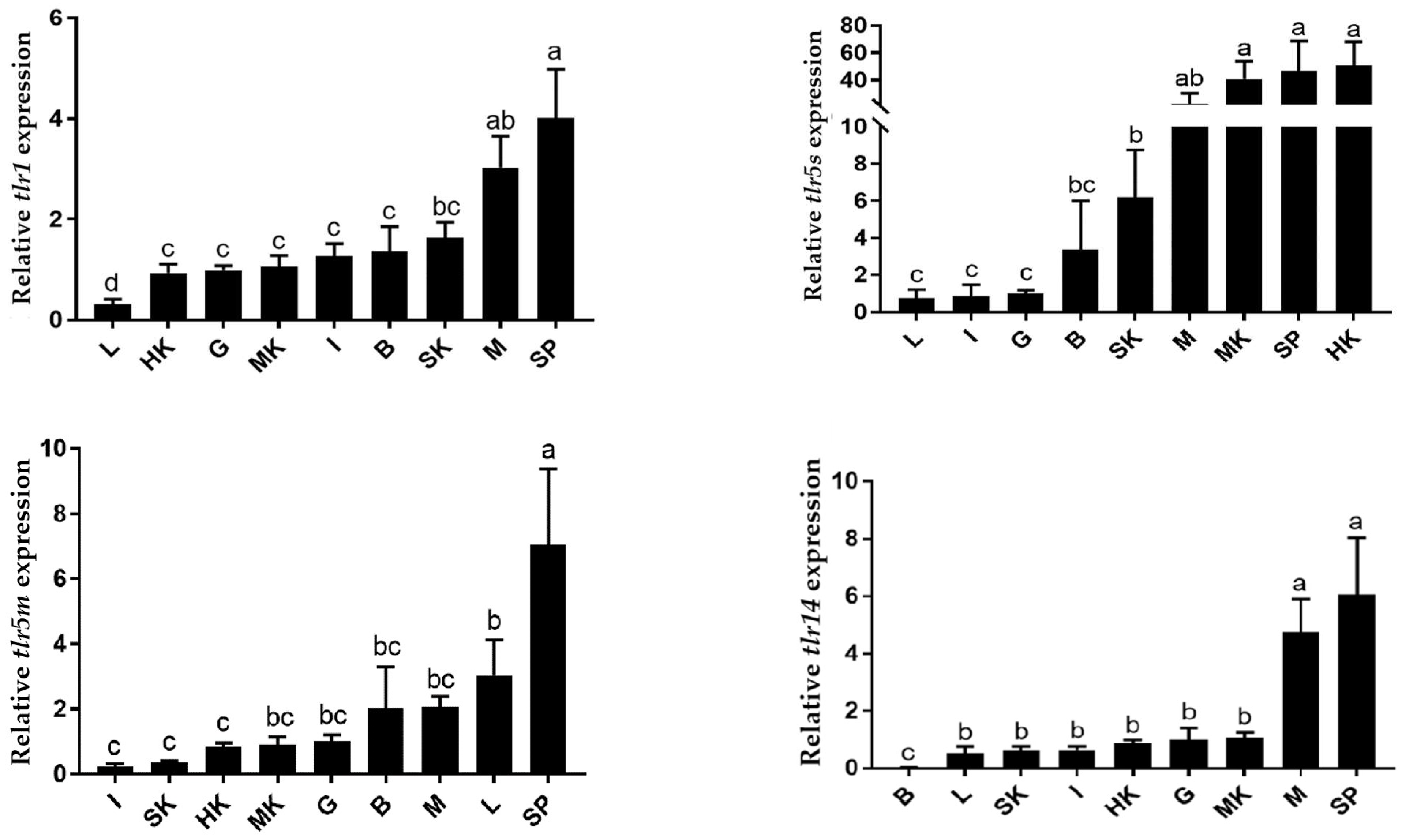
3.4. Expression Patterns of TLR Genes in Different Tissues
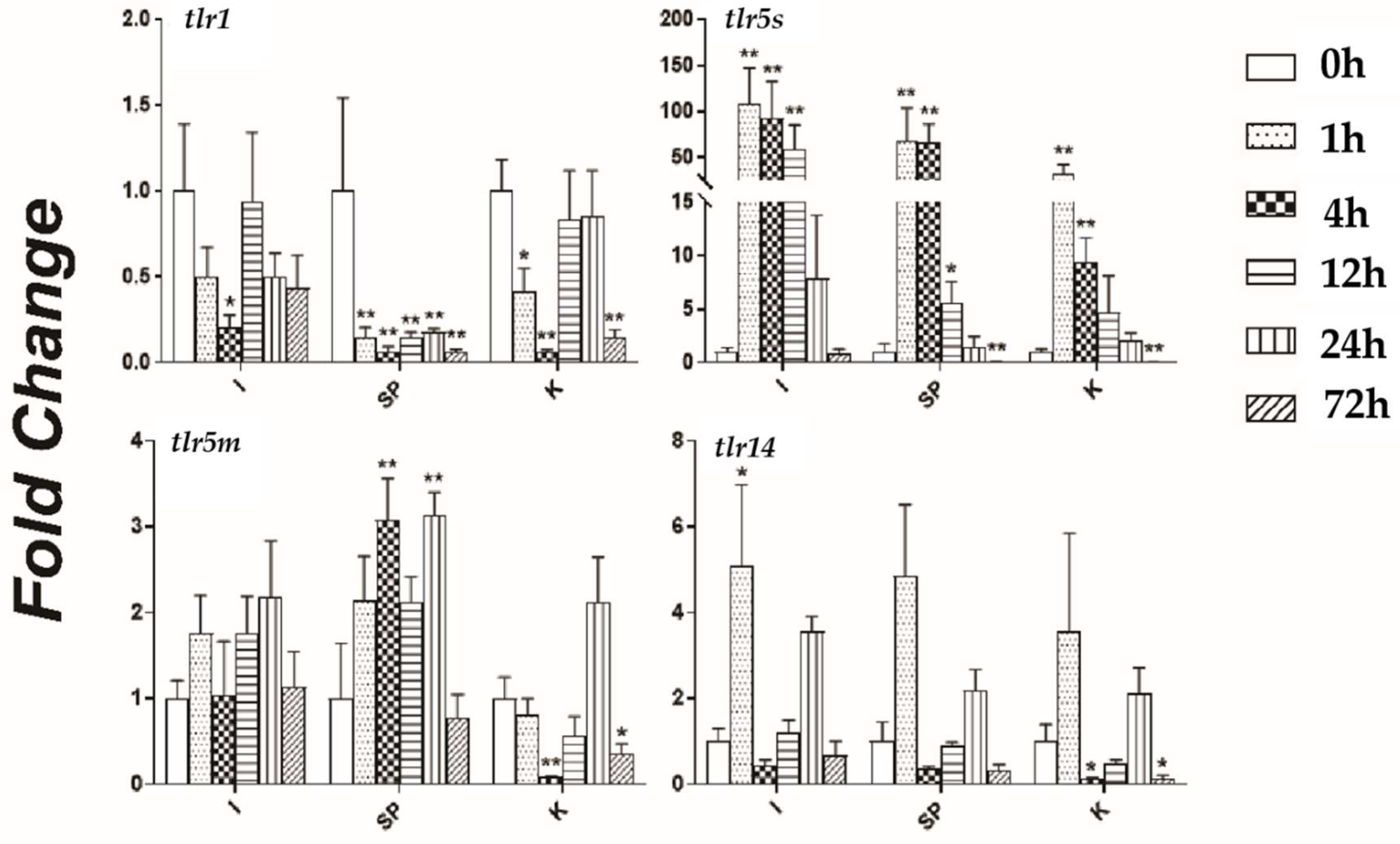
3.5. Responses of TLR Genes to Bacterial Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, R.; Mosser, D.M. Pattern recognition receptors in innate immunity, host defense, and immunopathology. Adv. Physiol. Educ. 2013, 37, 284–291. [Google Scholar] [CrossRef]
- Sahoo, B.R. Structure of fish Toll-like receptors (TLR) and NOD-like receptors (NLR). Int. J. Biol. Macromol. 2020, 161, 1602–1617. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netea, M.G.; Wijmenga, C.; O’Neill, L.A.J. Genetic variation in Toll-like receptors and disease susceptibility. Nat. Immunol. 2012, 13, 535–542. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Broz, P.; Monack, D.M. Newly described pattern recognition receptors team up against intracellular pathogens. Nat. Rev. Immunol. 2013, 13, 551–565. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Bowie, A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef]
- Blasius, A.L.; Beutler, B. Intracellular Toll-like Receptors. Immunity 2010, 32, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Goubau, D.; Deddouche, S.; Reis e Sousa, C. Cytosolic Sensing of Viruses. Immunity 2013, 38, 855–869. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Lin, Z.; Pang, X.; Shan, P.; Wang, J. MicroRNA regulation of Toll-like receptor signaling pathways in teleost fish. Fish Shellfish Immunol. 2018, 75, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kong, X.; Zhou, C.; Li, L.; Nie, G.; Li, X. Toll-like receptor recognition of bacteria in fish: Ligand specificity and signal pathways. Fish Shellfish Immunol. 2014, 41, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Rauta, P.R.; Samanta, M.; Dash, H.R.; Nayak, B.; Das, S. Toll-like receptors (TLRs) in aquatic animals: Signaling pathways, expressions and immune responses. Immunol. Lett. 2013, 158, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Yan, X.; Yang, N.; Fu, Q.; Xue, T.; Zhao, S.; Hu, J.; Li, Q.; Song, L.; Zhang, X.; et al. Genome-wide characterization of Toll-like receptors in black rockfish Sebastes schlegelii: Evolution and response mechanisms following Edwardsiella tarda infection. Int. J. Biol. Macromol. 2020, 164, 949–962. [Google Scholar] [CrossRef]
- Gong, Y.; Feng, S.; Li, S.; Zhang, Y.; Zhao, Z.; Hu, M.; Xu, P.; Jiang, Y. Genome-wide characterization of Toll-like receptor gene family in common carp (Cyprinus carpio) and their involvement in host immune response to Aeromonas hydrophila infection. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 24, 89–98. [Google Scholar] [CrossRef]
- Campanella, J.J.; Bitincka, L.; Smalley, J. MatGAT: An application that generates similarity/identity matrices using protein or DNA sequences. BMC Bioinform. 2003, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, I.; Sepulcre, M.P.; Meseguer, J.; Mulero, V. Molecular cloning, phylogenetic analysis and functional characterization of soluble Toll-like receptor 5 in gilthead seabream, Sparus aurata. Fish Shellfish Immunol. 2013, 35, 36–45. [Google Scholar] [CrossRef]
- Takano, T.; Hwang, S.D.; Kondo, H.; Hirono, I.; Aoki, T.; Sano, M. Evidence of Molecular Toll-like Receptor Mechanisms in Teleosts. Fish Pathol. 2010, 45, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Yang, P.; Liu, L.; Luo, Y.; Sun, Y.; Wang, W.; Chen, N.; Zhao, J. Advances in Intestinal Mucosal Immunoglobulins of Teleost Fish (A Review). Isr. J. Aquac.-Bamidgeh 2019, 71, 1617. Available online: http://hdl.handle.net/10524/62917 (accessed on 25 June 2022).
- Zhang, X.-T.; Zhang, G.-R.; Shi, Z.-C.; Yuan, Y.-J.; Zheng, H.; Lin, L.; Wei, K.-J.; Ji, W. Expression analysis of nine Toll-like receptors in yellow catfish (Pelteobagrus fulvidraco) responding to Aeromonas hydrophila challenge. Fish Shellfish Immunol. 2017, 63, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.; Chen, S.N.; Huo, H.J.; Nie, P. Identification and expression analysis of sixteen Toll-like receptor genes, TLR1, TLR2a, TLR2b, TLR3, TLR5M, TLR5S, TLR7–9, TLR13a–c, TLR14, TLR21–23 in mandarin fish Siniperca chuatsi. Dev. Comp. Immunol. 2021, 121, 104100. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting Edge: Bacterial Flagellin Activates Basolaterally Expressed TLR5 to Induce Epithelial Proinflammatory Gene Expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.-S.; Li, Y.-W.; Deng, Y.; Huang, Y.-Q.; He, S.-H.; Dai, J.; Zhao, S.-Z.; Dan, X.-M.; Luo, X.-C. Molecular identification and expression analysis of TLR5M and TLR5S from orange-spotted grouper (Epinepheluscoioides). Fish Shellfish Immunol. 2017, 63, 97–102. [Google Scholar] [CrossRef]
- Tsujita, T.; Tsukada, H.; Nakao, M.; Oshiumi, H.; Matsumoto, M.; Seya, T. Sensing Bacterial Flagellin by Membrane and Soluble Orthologs of Toll-like Receptor 5 in Rainbow Trout (Onchorhynchus mikiss). J. Biol. Chem. 2004, 279, 48588–48597. [Google Scholar] [CrossRef] [Green Version]
- Tsujita, T.; Ishii, A.; Tsukada, H.; Matsumoto, M.; Che, F.-S.; Seya, T. Fish soluble Toll-like receptor (TLR)5 amplifies human TLR5 response via physical binding to flagellin. Vaccine 2006, 24, 2193–2199. [Google Scholar] [CrossRef]
- Yu, J.; Liu, X.; Yang, N.; Wang, B.; Su, B.; Fu, Q.; Zhang, M.; Tan, F.; Li, C. Characterization of toll-like receptor 1 (TLR1) in turbot (Scophthalmus maximus L.). Fish Shellfish Immunol. 2021, 115, 27–34. [Google Scholar] [CrossRef]
- Wu, X.-Y.; Xiang, L.-X.; Huang, L.; Jin, Y.; Shao, J.-Z. Characterization, expression and evolution analysis of Toll-like receptor 1 gene in pufferfish (Tetraodon nigroviridis). Int. J. Immunogenet. 2008, 35, 215–225. [Google Scholar] [CrossRef]
- Jin, M.S.; Kim, S.E.; Heo, J.Y.; Lee, M.E.; Kim, H.M.; Paik, S.-G.; Lee, H.; Lee, J.-O. Crystal Structure of the TLR1-TLR2 Heterodimer Induced by Binding of a Tri-Acylated Lipopeptide. Cell 2007, 130, 1071–1082. [Google Scholar] [CrossRef] [Green Version]
- Kasamatsu, J.; Oshiumi, H.; Matsumoto, M.; Kasahara, M.; Seya, T. Phylogenetic and expression analysis of lamprey toll-like receptors. Dev. Comp. Immunol. 2010, 34, 855–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.D.; Kondo, H.; Hirono, I.; Aoki, T. Molecular cloning and characterization of Toll-like receptor 14 in Japanese flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2011, 30, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-W.; Xu, D.-D.; Li, X.; Mo, Z.-Q.; Luo, X.-C.; Li, A.-X.; Dan, X.-M. Identification and characterization of three TLR1 subfamily members from the orange-spotted grouper, Epinephelus coioides. Dev. Comp. Immunol. 2016, 61, 180–189. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Number | Sequence (5′ to 3′) | Usage | Length (bp) |
|---|---|---|---|---|
| tlr1-QF tlr1-QR | XM_020624433.1 | ACAGCAACTCCACCGAGAAGA CGGCTAGGTTAAGGACCACGA | Quantitative real-time PCR | 209 |
| tlr1-F tlr1-R | ATGAAGCCCATGACTGCCACCT TCACTCATCTATTTCTGTGGCT | ORF cloning | 2406 | |
| tlr5s-QF tlr5s-QR | M_020603847.1 | TGTAGCCAACTGTGCCTTCCG CACATTCATGCCGAGCACCAG | Quantitative real-time PCR | 219 |
| tlr5s-F tlr5s-R | ATGAAAGAAGATTTAAGGTG TTACTTCTGTTCGACCTGAG | ORF cloning | 1962 | |
| tlr5m-QF tlr5m-QR | XM_020617012.1 | GGCTCTTCAGTTGGCTTTCAC ATGCTGGGAGGCAGAATAGGT | Quantitative real-time PCR | 132 |
| tlr5m-F tlr5m-R | ATGTGGACACTGGCTCTTCA TCACATGGCAATTGCTCTGA | ORF cloning | 2667 | |
| tlr14-QF tlr14-QR | XM_020615508.1 | GACTTTGCTCGTTCACCCACAT TACCCAGGCATCACAAGGCAC | Quantitative real-time PCR | 201 |
| tlr14-F tlr14-R | ATGATTTGGAAGTTGATTCA TCAAACTACCTCCAGAGGGA | ORF cloning | 2601 | |
| EF-1α-QF EF-1α-QR | KC011266.1 | CCTTCTTGGGTATGGAATCTTGC CAGAGTATTTACGCTCAGGTGGG | Quantitative real-time PCR | 190 |
| Feature | tlr1 | tlr5s | tlr5m | tlr14 |
|---|---|---|---|---|
| cDNA length (bp) | 2876 | 2174 | 3400 | 3319 |
| ORF (bp) | 2406 | 1962 | 2667 | 2601 |
| Amino acids (aa) | 801 | 653 | 888 | 866 |
| Signal peptide | 22 | 20 | 22 | |
| LRR motifs | 6 | 12 | 11 | 7 |
| LRR-CT | 0 | 1 | 1 | 1 |
| LRR-NT | 0 | 1 | 0 | 0 |
| TM domain | 0 | 0 | 1 | 1 |
| TIR domain | 1 | 0 | 1 | 1 |
| Molecular weight (kDa) | 108.12 | 80.87 | 127.47 | 125.001 |
| pI | 6.50 | 8.72 | 5.79 | 8.59 |
| Instability index | 41.68 | 39.97 | 38.08 | 38.83 |
| Aliphatic index | 103.83 | 104.37 | 101.76 | 91.26 |
| Gene ID | XM_020624433.1 | XM_020603847.1 | XM_020617012.1 | XM_020615508.1 |
| Species | tlr1 | tlr5s | tlr5m | tlr14 |
|---|---|---|---|---|
| Homo sapiens | 37.2% | 38.5% | ||
| Gallus | 36.5% | 40.3% | ||
| Xenopus tropicalis | 33.9% | |||
| Danio rerio | 47.5% | 41.2% | ||
| Oncorhynchus mykiss | 58.8% | 52.2% | ||
| Ictalurus punctatus | 45.8% | 43.2% | ||
| Siniperca chuatsi | 74.0% | 74.9% | 74.5% | 75.5% |
| Epinephelus coioides | 68.4% | 68.5% | 64.4% | |
| Scophthalmus maximus | 72.2% | 72.9% | ||
| Salmo salar | 53.1% | |||
| Takifugu rubripes | 62.3% | 62.5% | 71.7% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, J.; Zhang, Z.; Guo, H.; Xu, Q.; Yang, D.; Yuan, H. Molecular Cloning and Characterization of tlr1, tlr5s, tlr5m, and tlr14 Genes in the Swamp Eel, Monopterus albus. Fishes 2022, 7, 177. https://doi.org/10.3390/fishes7040177
Xiang J, Zhang Z, Guo H, Xu Q, Yang D, Yuan H. Molecular Cloning and Characterization of tlr1, tlr5s, tlr5m, and tlr14 Genes in the Swamp Eel, Monopterus albus. Fishes. 2022; 7(4):177. https://doi.org/10.3390/fishes7040177
Chicago/Turabian StyleXiang, Jing, Zhehua Zhang, Huizhi Guo, Qiaoqing Xu, Daiqin Yang, and Hanwen Yuan. 2022. "Molecular Cloning and Characterization of tlr1, tlr5s, tlr5m, and tlr14 Genes in the Swamp Eel, Monopterus albus" Fishes 7, no. 4: 177. https://doi.org/10.3390/fishes7040177
APA StyleXiang, J., Zhang, Z., Guo, H., Xu, Q., Yang, D., & Yuan, H. (2022). Molecular Cloning and Characterization of tlr1, tlr5s, tlr5m, and tlr14 Genes in the Swamp Eel, Monopterus albus. Fishes, 7(4), 177. https://doi.org/10.3390/fishes7040177