
Transcriptomic Down-Regulation of Immune System Components in Barrier and Hematopoietic Tissues after Lipopolysaccharide Injection in Antarctic Notothenia coriiceps

 , , , , and
, , , , and 
Abstract
:
1. Introduction
2. Materials and Methods
3. Results
3.1. Blood Biochemistry
3.2. Transcriptome Assembly and Differentially Expressed Genes
3.3. Control Versus Sham Differentially Expressed Genes
3.4. LPS versus Sham Differentially Expressed Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nowlin, W.D.; Klinck, J.M. The Physics of the Antarctic Circumpolar Current. Rev. Geophys. 1986, 24, 469–491. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.; Murphy, E.J.; Meredith, M.P.; King, J.C.; Peck, L.S.; Barnes, D.K.A.; Smith, R.C. Climate Change and the Marine Ecosystem of the Western Antarctic Peninsula. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 149–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rintoul, S.R. Antarctic Circumpolar Current. In Encyclopedia of Ocean Sciences; Elsevier: Amsterdam, The Netherlands, 2009; pp. 178–190. [Google Scholar]
- Shin, S.C.; Ahn, D.H.; Kim, S.J.; Pyo, C.W.; Lee, H.; Kim, M.-K.; Lee, J.; Lee, J.E.; Detrich, H.W.; Postlethwait, J.H.; et al. The Genome Sequence of the Antarctic Bullhead Notothen Reveals Evolutionary Adaptations to a Cold Environment. Genome Biol. 2014, 15, 468. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.L.; Wohlschlag, D.E. Freezing Resistance in Some Antarctic Fishes. Science 1969, 163, 1073–1075. [Google Scholar] [CrossRef]
- DeVries, A.L.; Cheng, C.-H.C. Antifreeze Proteins and Organismal Freezing Avoidance in Polar Fishes. Fish Physiol. 2005, 22, 155–201. [Google Scholar]
- Cheng, C.-H.C.; Chen, L. Evolution of an Antifreeze Glycoprotein. Nature 1999, 401, 443–444. [Google Scholar] [CrossRef]
- Kim, B.M.; Amores, A.; Kang, S.; Ahn, D.H.; Kim, J.H.; Kim, I.C.; Lee, J.H.; Lee, S.G.; Lee, H.; Lee, J.; et al. Antarctic Blackfin Icefish Genome Reveals Adaptations to Extreme Environments. Nat. Ecol. Evol. 2019, 3, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Albertson, R.C.; Yan, Y.L.; Titus, T.A.; Pisano, E.; Vacchi, M.; Yelick, P.C.; Detrich, H.W.; Postlethwait, J.H. Molecular Pedomorphism Underlies Craniofacial Skeletal Evolution in Antarctic Notothenioid Fishes. BMC Evol. Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Detrich, H.W.; Johnson, K.A.; Marchese-Ragona, S.P. Polymerization of Antarctic Fish Tubulins at Low Temperatures: Energetic Aspects. Biochemistry 1989, 28, 10085–10093. [Google Scholar] [CrossRef]
- Detrich, H.W.; Parker, S.K.; Williams, J.; Nogales, E.; Downing, K.H. Cold Adaptation of Microtubule Assembly and Dynamics. Structural Interpretation of Primary Sequence Changes Present in the α- and β-Tubulins of Antarctic Fishes. J. Biol. Chem. 2000, 275, 37038–37047. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.M.; Sidell, B.D. The Interplay among Cardiac Ultrastructure, Metabolism and the Expression of Oxygen-Binding Proteins in Antarctic Fishes. J. Exp. Biol. 2000, 203, 1287–1297. [Google Scholar] [CrossRef] [PubMed]
- Place, S.P.; Hofmann, G.E. Comparison of Hsc70 Orthologs from Polar and Temperate Notothenioid Fishes: Differences in Prevention of Aggregation and Refolding of Denatured Proteins. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 288, R1195–R1202. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, G.E.; Buckley, B.; Airaksinen, S.; Keen, J.E.; Somero, G.N. Heat-Shock Protein Expression Is Absent in the Antarctic Fish Trematomus bernacchii (Family Nototheniidae). J. Exp. Biol. 2000, 203, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Furbino, L.E.; Godinho, V.M.; Santiago, I.F.; Pellizari, F.M.; Alves, T.M.A.; Zani, C.L.; Junior, P.A.S.; Romanha, A.J.; Carvalho, A.G.O.; Gil, L.H.V.G.; et al. Diversity Patterns, Ecology and Biological Activities of Fungal Communities Associated with the Endemic Macroalgae Across the Antarctic Peninsula. Microb. Ecol. 2014, 67, 775–787. [Google Scholar] [CrossRef]
- Grimaldi, W.W.; Seddon, P.J.; Lyver, P.O.; Nakagawa, S.; Tompkins, D.M. Infectious Diseases of Antarctic Penguins: Current Status and Future Threats. Polar Biol. 2015, 38, 591–606. [Google Scholar] [CrossRef]
- Duffy, G.A.; Coetzee, B.W.T.; Latombe, G.; Akerman, A.H.; McGeoch, M.A.; Chown, S.L. Barriers to Globally Invasive Species Are Weakening across the Antarctic. Divers. Distrib. 2017, 23, 982–996. [Google Scholar] [CrossRef] [Green Version]
- Cowan, D.A.; Chown, S.L.; Convey, P.; Tuffin, M.; Hughes, K.; Pointing, S.; Vincent, W.F. Non-Indigenous Microorganisms in the Antarctic: Assessing the Risks. Trends Microbiol. 2011, 19, 540–548. [Google Scholar] [CrossRef]
- Magor, B.G.; Magor, K.E. Evolution of Effectors and Receptors of Innate Immunity. Dev. Comp. Immunol. 2001, 25, 651–682. [Google Scholar] [CrossRef]
- Tort, L.; Balasch, J.C.; Mackenzie, S. Fish Immune System. A Crossroads between Innate and Adaptive Responses. Inmunologia 2003, 22, 277–286. [Google Scholar] [CrossRef]
- Magnadottir, B.; Gudmundsdottir, B.K.; Lange, S.; Steinarsson, A.; Oddgeirsson, M.; Bowden, T.; Bricknell, I.; Dalmo, R.A.; Gudmundsdottir, S. Immunostimulation of Larvae and Juveniles of Cod, Gadus Morhua L. J. Fish Dis. 2006, 29, 147–155. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The Mucosal Immune System of Fish: The Evolution of Tolerating Commensals While Fighting Pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunyer, J.O. Fishing for Mammalian Paradigms in the Teleost Immune System. Nat. Immunol. 2013, 14, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Nie, L.; Zhu, G.; Xiang, L.; Shao, J. Advances in Research of Fish Immune-Relevant Genes: A Comparative Overview of Innate and Adaptive Immunity in Teleosts. Dev. Comp. Immunol. 2013, 39, 39–62. [Google Scholar] [CrossRef]
- Bonneaud, C.; Wilson, R.S.; Seebacher, F. Immune-Challenged Fish up-Regulate Their Metabolic Scope to Support Locomotion. PLoS ONE 2016, 11, e0166028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abram, Q.H.; Dixon, B.; Katzenback, B.A. Impacts of Low Temperature on the Teleost Immune System. Biology 2017, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Bly, J.E.; Clem, L.W. Temperature and Teleost Immune Functions. Fish Shellfish Immunol. 1992, 2, 159–171. [Google Scholar] [CrossRef]
- Le Morvan, C.; Troutaud, D.; Deschaux, P. Differential Effects of Temperature on Specific and Nonspecific Immune Defences in Fish. J. Exp. Biol. 1998, 201, 165–168. [Google Scholar] [CrossRef]
- MacKenzie, S.; Balasch, J.C.; Novoa, B.; Ribas, L.; Roher, N.; Krasnov, A.; Figueras, A. Comparative Analysis of the Acute Response of the Trout, O. mykiss, Head Kidney to in Vivo Challenge with Virulent and Attenuated Infectious Hematopoietic Necrosis Virus and LPS-Induced Inflammation. BMC Genom. 2008, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, S.; Jiang, Z.; Wang, X.; Liu, Y. Chemokine Receptor CXCR3 in Turbot (Scophthalmus maximus): Cloning, Characterization and Its Responses to Lipopolysaccharide. Fish Physiol. Biochem. 2016, 42, 659–671. [Google Scholar] [CrossRef]
- Magnoni, L.; Roher, N.; Crespo, D.; Krasnov, A.; Planas, J. In Vivo Molecular Responses of Fast and Slow Muscle Fibers to Lipopolysaccharide in a Teleost Fish, the Rainbow Trout (Oncorhynchus mykiss). Biology 2015, 4, 67–87. [Google Scholar] [CrossRef] [Green Version]
- Le Morvan-Rocher, C.; Troutaud, D.; Deschaux, P. Effects of Temperature on Carp Leukocyte Mitogen-Induced Proliferation and Nonspecific Cytotoxic Activity. Dev. Comp. Immunol. 1995, 19, 87–95. [Google Scholar] [CrossRef]
- Le Morvan, C.; Clerton, P.; Deschaux, P.; Troutaud, D. Effects of Environmental Temperature on Macrophage Activities in Carp. Fish Shellfish Immunol. 1997, 7, 209–212. [Google Scholar] [CrossRef]
- Dexiang, C.; Ainsworth, A.J. Effect of Temperature on the Immune System of Channel Catfish (Ictalurus punctatus)—II. Adaptation of Anterior Kidney Phagocytes to 10 °C. Comp. Biochem. Physiol. Part A Physiol. 1991, 100, 913–918. [Google Scholar] [CrossRef]
- Silva, J. Phagocytosis and Giant Cell Formation at 0 °C by Macrophage (MØ) of Notothenia coriiceps. J. Fish Biol. 2002, 60, 466–478. [Google Scholar] [CrossRef]
- Silva, J.R.M.C.; Hernadez-Blazquez, F.J.; Barbieri, R.L. Induced Inflammatory Process in the Antarctic Fish Notothenia neglecta. Polar Biol. 1998, 20, 206–212. [Google Scholar] [CrossRef]
- Silva, J.R.M.C.; Staines, N.A.; Parra, O.M.; Hernandez-Blazquez, F.J. Experimental Studies on the Response of the Fish (Notothenia coriiceps Richardson, 1844) to Parasite (Pseudoterranova decipiens Krabbe, 1878) and Other Irritant Stimuli at Antarctic Temperatures. Polar Biol. 1999, 22, 417–424. [Google Scholar] [CrossRef]
- O’Neill, J.G.; White, M.G.; Sims, T.A.; Barber, D.L. The Inflammatory Response of the Antarctic Silverfish, Pleuragramma antarcticum Boulenger, 1902, to a Plerocercoid (Species Unknown) Infestation. J. Fish Biol. 1987, 31, 231–232. [Google Scholar] [CrossRef]
- Cunha da Silva, J.R.M.; Cooper, E.L.; Sinhorini, I.L.; Borges, J.C.S.; Jensch-Junior, B.E.; Porto-Neto, L.R.; Hernandez-Blazquez, F.J.; Vellutini, B.C.; Pressinotti, L.N.; Costa-Pinto, F.A. Microscopical Study of Experimental Wound Healing in Notothenia coriiceps (Cabeçuda) at 0 Degrees C. Cell Tissue Res. 2005, 321, 401–410. [Google Scholar] [CrossRef]
- Ahn, D.H.; Shin, S.C.; Park, H. Characterization of Toll-like Receptor Gene Expression and the Pathogen Agonist Response in the Antarctic Bullhead Notothen Notothenia coriiceps. Immunogenetics 2014, 66, 563–573. [Google Scholar] [CrossRef]
- Sousa, C.; Fernandes, S.A.; Cardoso, J.C.R.; Wang, Y.; Zhai, W.; Guerreiro, P.M.; Chen, L.; Canário, A.V.M.; Power, D.M. Toll-Like Receptor Evolution: Does Temperature Matter? Front. Immunol. 2022, 13, 27. [Google Scholar] [CrossRef]
- Martínez, D.P.; Sousa, C.; Oyarzún, R.; Pontigo, J.P.; Canario, A.V.M.; Power, D.M.; Vargas-Chacoff, L.; Guerreiro, P.M. LPS Modulates the Expression of Iron-Related Immune Genes in Two Antarctic Notothenoids. Front. Physiol. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Cheng, C.-H.C.; Hu, P.; Ye, H.; Chen, Z.; Cao, L.; Chen, L.; Shen, Y.; Chen, L. Adaptive Evolution of Hepcidin Genes in Antarctic Notothenioid Fishes. Mol. Biol. Evol. 2008, 25, 1099–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, D.-H.; Kang, S.; Park, H. Transcriptome Analysis of Immune Response Genes Induced by Pathogen Agonists in the Antarctic Bullhead Notothen Notothenia coriiceps. Fish Shellfish Immunol. 2016, 55, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Huang, S.; Zhai, W.; Jiang, S.; Li, W.; Wang, F.; Xu, Q. Comparative Proteomic Analysis of Head Kidney among Three Antarctic Fishes. Mar. Biol. 2022, 169, 58. [Google Scholar] [CrossRef]
- Nilsson, C.; Skoglund, A.; Moran, A.P.; Annuk, H.; Engstrand, L.; Normark, S. Lipopolysaccharide Diversity Evolving in Helicobacter Pylori Communities through Genetic Modifications in Fucosyltransferases. PLoS ONE 2008, 3, e3811. [Google Scholar] [CrossRef]
- Araya, M.A.; Valenzuela, T.; Inostroza, N.G.; Maruyama, F.; Jorquera, M.A.; Acuña, J.J. Isolation and Characterization of Cold-Tolerant Hyper-ACC-Degrading Bacteria from the Rhizosphere, Endosphere, and Phyllosphere of Antarctic Vascular Plants. Microorganisms 2020, 8, 1788. [Google Scholar] [CrossRef]
- Meredith, T.C.; Aggarwal, P.; Mamat, U.; Lindner, B.; Woodard, R.W. Redefining the Requisite Lipopolysaccharide Structure in Escherichia coli. ACS Chem. Biol. 2006, 1, 33–42. [Google Scholar] [CrossRef]
- Bishop, R.E. Fundamentals of Endotoxin Structure and Function. In Concepts in Bacterial Virulence; KARGER: Basel, Switzerland, 2004; Volume 12, pp. 1–27. [Google Scholar]
- Swain, P.; Nayak, S.K.; Nanda, P.K.; Dash, S. Biological Effects of Bacterial Lipopolysaccharide (Endotoxin) in Fish: A Review. Fish Shellfish Immunol. 2008, 25, 191–201. [Google Scholar] [CrossRef]
- Langston, A.L.; Johnstone, R.; Ellis, A.E. The Kinetics of the Hypoferraemic Response and Changes in Levels of Alternative Complement Activity in Diploid and Triploid Atlantic Salmon, Following Injection of Lipopolysaccharide. Fish Shellfish Immunol. 2001, 11, 333–345. [Google Scholar] [CrossRef]
- Chu, Q.; Bi, D.; Zheng, W.; Xu, T. MicroRNA Negatively Regulates NF-ΚB-Mediated Immune Responses by Targeting NOD1 in the Teleost Fish Miichthys miiuy. Sci. China Life Sci. 2021, 64, 803–815. [Google Scholar] [CrossRef]
- Haukenes, A.H.; Barton, B.A. Characterization of the Cortisol Response Following an Acute Challenge with Lipopolysaccharide in Yellow Perch and the Influence of Rearing Density. J. Fish Biol. 2004, 64, 851–862. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, M.; Fu, L.; Zhong, L.; Liu, G.; Zheng, Y.; Chen, X.; Bian, W. Liver Transcriptome Analysis and Cortisol Immune-Response Modulation in Lipopolysaccharide-Stimulated in Channel Catfish (Ictalurus punctatus). Fish Shellfish Immunol. 2020, 101, 19–50. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ellis, A.E. Lysozyme Assays. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., Van Muiswinkel, W.B., Eds.; SOS Publications: Cambridge, UK, 1990; pp. 101–103. [Google Scholar]
- Ellis, A.E. Innate Host Defense Mechanisms of Fish against Viruses and Bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Rotllant, J.; Guerreiro, P.M.; Anjos, L.; Redruello, B.; Canario, A.V.M.; Power, D.M. Stimulation of Cortisol Release by the N Terminus of Teleost Parathyroid Hormone-Related Protein in Interrenal Cells in Vitro. Endocrinology 2005, 146, 71–76. [Google Scholar] [CrossRef]
- Andrews, S. FASTQC A Quality Control Tool for High Throughput Sequence Data. Babraham Inst. 2015. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 January 2022).
- Krueger, F. Trim Galore: A Wrapper Tool around Cutadapt and FastQC. Babraham Inst. 2015. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 15 January 2022).
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-Free Quality Assessment of de Novo Transcriptome Assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl De Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [Green Version]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A Tool for Discovery and Visualization of Enriched GO Terms in Ranked Gene Lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A Web Server for Annotation and Identification of Enriched Pathways and Diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef] [PubMed]
- Kandalski, P.K.; de Souza, M.R.D.P.; Herrerias, T.; Machado, C.; Zaleski, T.; Forgati, M.; Guillen, A.C.; Viana, D.; Moura, M.O.; Donatti, L. Effects of Short-Term Thermal Stress on the Plasma Biochemical Profiles of Two Antarctic Nototheniid Species. Rev. Fish Biol. Fish. 2018, 28, 925–940. [Google Scholar] [CrossRef]
- Hanif, A.; Bakopoulos, V.; Leonardos, I.; Dimitriadis, G.J. The Effect of Sea Bream (Sparus aurata) Broodstock and Larval Vaccination on the Susceptibility by Photobacterium damsela Subsp. Piscicida and on the Humoral Immune Parameters. Fish Shellfish Immunol. 2005, 19, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.K.; Swain, P.; Nanda, P.K.; Dash, S.; Shukla, S.; Meher, P.K.; Maiti, N.K. Effect of Endotoxin on the Immunity of Indian Major Carp, Labeo rohita. Fish Shellfish Immunol. 2008, 24, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Sepulcre, M.P.; Alcaraz-Pérez, F.; López-Muñoz, A.; Roca, F.J.; Meseguer, J.; Cayuela, M.L.; Mulero, V. Evolution of Lipopolysaccharide (LPS) Recognition and Signaling: Fish TLR4 Does Not Recognize LPS and Negatively Regulates NF-ΚB Activation. J. Immunol. 2009, 182, 1836–1845. [Google Scholar] [CrossRef] [Green Version]
- Berczi, I.; Bertók, L.; Bereznai, T. Comparative Studies on the Toxicity of Escherichia Coli Lipopolysaccharide Endotoxin in Various Animal Species. Can. J. Microbiol. 1966, 12, 1070–1071. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Lee, J.-O. Recognition of Lipopolysaccharide Pattern by TLR4 Complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef] [Green Version]
- Obach, A.; Quentel, C.; Baudin Laurencin, F. Effects of Alpha-Tocopherol and Dietary Oxidized Fish Oil on the Immune Response of Sea Bass Dicentrarchus labrax. Dis. Aquat. Org. 1993, 15, 175–185. [Google Scholar] [CrossRef]
- Yildiz, H.Y.; Altunay, S. Physiological Stress and Innate Immune Response in Gilthead Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax) Exposed to Combination of Trimethoprim and Sulfamethoxazole (TMP-SMX). Fish Physiol. Biochem. 2011, 37, 401–409. [Google Scholar] [CrossRef]
- Christybapita, D.; Divyagnaneswari, M.; Dinakaran Michael, R. Oral Administration of Eclipta Alba Leaf Aqueous Extract Enhances the Non-Specific Immune Responses and Disease Resistance of Oreochromis mossambicus. Fish Shellfish Immunol. 2007, 23, 840–852. [Google Scholar] [CrossRef]
- Lie, Ø.; Evensen, Ø.; Sørensen, A.; Frøysadal, E. Study on Lysozyme Activity in Some Fish Species. Dis. Aquat. Org. 1989, 6, 55–61. [Google Scholar] [CrossRef]
- Simide, R.; Richard, S.; Prévot-D’Alvise, N.; Miard, T.; Gaillard, S. Assessment of the Accuracy of Physiological Blood Indicators for the Evaluation of Stress, Health Status and Welfare in Siberian Sturgeon (Acipenser baerii) Subject to Chronic Heat Stress and Dietary Supplementation. Int. Aquat. Res. 2016, 8, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Garate, J.A.; Oostenbrink, C. Lipid a from Lipopolysaccharide Recognition: Structure, Dynamics and Cooperativity by Molecular Dynamics Simulations. Proteins Struct. Funct. Bioinform. 2013, 81, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Lerouge, I. O-Antigen Structural Variation: Mechanisms and Possible Roles in Animal/Plant–Microbe Interactions. FEMS Microbiol. Rev. 2002, 26, 17–47. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.S.; Jagannadham, M.V.; Ray, M.K. Low-Temperature-Induced Changes in Composition and Fluidity of Lipopolysaccharides in the Antarctic Psychrotrophic Bacterium Pseudomonas syringae. J. Bacteriol. 2002, 184, 6746–6749. [Google Scholar] [CrossRef] [Green Version]
- Wu, E.L.; Engström, O.; Jo, S.; Stuhlsatz, D.; Yeom, M.S.; Klauda, J.B.; Widmalm, G.; Im, W. Molecular Dynamics and NMR Spectroscopy Studies of E. Coli Lipopolysaccharide Structure and Dynamics. Biophys. J. 2013, 105, 1444–1455. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, M.K. Mechanism of Bacterial Adaptation to Low Temperature. J. Biosci. 2006, 31, 157–165. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Ohno, N.; Morrison, D.C. Lipopolysaccharide Interactions with Lysozyme Differentially Affect Lipopolysaccharide Immunostimulatory Activity. Eur. J. Biochem. 1989, 186, 629–636. [Google Scholar] [CrossRef]
- Sanders, G.E.; Batts, W.N.; Winton, J.R. Susceptibility of Zebrafish (Danio rerio) to a Model Pathogen, Spring Viremia of Carp Virus. Comp. Med. 2003, 53, 514–521. [Google Scholar]
- Cheng, A.C.; Cheng, S.A.; Chen, Y.Y.; Chen, J.C. Effects of Temperature Change on the Innate Cellular and Humoral Immune Responses of Orange-Spotted Grouper Epinephelus coioides and Its Susceptibility to Vibrio alginolyticus. Fish Shellfish Immunol. 2009, 26, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Ndong, D.; Chen, Y.-Y.; Lin, Y.-H.; Vaseeharan, B.; Chen, J.-C. The Immune Response of Tilapia Oreochromis mossambicus and Its Susceptibility to Streptococcus iniae under Stress in Low and High Temperatures. Fish Shellfish Immunol. 2007, 22, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Avunje, S.; Oh, M.J.; Jung, S.J. Impaired TLR2 and TLR7 Response in Olive Flounder Infected with Viral Haemorrhagic Septicaemia Virus at Host Susceptible 15 °C but High at Non-Susceptible 20 °C. Fish Shellfish Immunol. 2013, 34, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Xu, D.; Tian, L.; Chen, R.; Wang, L.; Tan, P.; You, Q. Overwinter Mortality in Yellow Drum (Nibea albiflora): Insights from Growth and Immune Responses to Cold and Starvation Stress. Fish Shellfish Immunol. 2019, 92, 341–347. [Google Scholar] [CrossRef]
- Magnadottir, B.; Audunsdottir, S.S.; Bragason, B.T.; Gisladottir, B.; Jonsson, Z.O.; Gudmundsdottir, S. The Acute Phase Response of Atlantic Cod (Gadus morhua): Humoral and Cellular Response. Fish Shellfish Immunol. 2011, 30, 1124–1130. [Google Scholar] [CrossRef]
- Palstra, A.P.; Kals, J.; Garcia, A.B.; Dirks, R.P.; Poelman, M. Immunomodulatory Effects of Dietary Seaweeds in LPS Challenged Atlantic Salmon Salmo salar as Determined by Deep RNA Sequencing of the Head Kidney Transcriptome. Front. Physiol. 2018, 9, 625. [Google Scholar] [CrossRef]
- Lu, X.-J.; Ning, Y.-J.; Liu, H.; Nie, L.; Chen, J. A Novel Lipopolysaccharide Recognition Mechanism Mediated by Internalization in Teleost Macrophages. Front. Immunol. 2018, 9, 2758. [Google Scholar] [CrossRef] [Green Version]
- Zoccola, E.; Kellie, S.; Barnes, A.C. Immune Transcriptome Reveals the Mincle C-Type Lectin Receptor Acts as a Partial Replacement for TLR4 in Lipopolysaccharide-Mediated Inflammatory Response in Barramundi (Lates calcarifer). Mol. Immunol. 2017, 83, 33–45. [Google Scholar] [CrossRef]
- Chiaranunt, P.; Burrows, K.; Ngai, L.; Mortha, A. Isolation of mononuclear phagocytes from the mouse gut. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 2020; Volume 632, pp. 67–90. [Google Scholar] [CrossRef]
- Fritz, J.H.; Girardin, S.E. How Toll-like Receptors and Nod-like Receptors Contribute to Innate Immunity in Mammals. J. Endotoxin Res. 2005, 11, 390–394. [Google Scholar] [CrossRef]
- Hong, J.; Yoon, D.; Nam, Y.; Seo, D.; Kim, J.H.; Kim, M.S.; Lee, T.Y.; Kim, K.S.; Ko, P.W.; Lee, H.W.; et al. Lipopolysaccharide Administration for a Mouse Model of Cerebellar Ataxia with Neuroinflammation. Sci. Rep. 2020, 10, 13337. [Google Scholar] [CrossRef]
- Yang, I.V.; Wade, C.M.; Kang, H.M.; Alper, S.; Rutledge, H.; Lackford, B.; Eskin, E.; Daly, M.J.; Schwartz, D.A. Identification of Novel Genes That Mediate Innate Immunity Using Inbred Mice. Genetics 2009, 183, 1535–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Ye, Z.; Zhang, D.; Gao, C.; Su, B.; Song, L.; Tan, F.; Song, H.; Wang, Y.; Li, C. Characterization and Expression Profiling of NOD-like Receptor C3 (NLRC3) in Mucosal Tissues of Turbot (Scophthalmus maximus L.) Following Bacterial Challenge. Fish Shellfish Immunol. 2017, 66, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Li, J.; He, J.; Liu, W.; Jiang, L.; Ye, Y.; Wu, C. Anti-Infective Mannose Receptor Immune Mechanism in Large Yellow Croaker (Larimichthys crocea). Fish Shellfish Immunol. 2016, 54, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Bin Park, S.; Hikima, J.-I.; Suzuki, Y.; Ohtani, M.; Nho, S.W.; Cha, I.S.; Bin Jang, H.; Kondo, H.; Hirono, I.; Aoki, T.; et al. Molecular cloning and functional analysis of nucleotide-binding oligomerization domain 1 (NOD1) in olive flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2011, 36, 680–687. [Google Scholar] [CrossRef]
- Swain, B.; Basu, M.; Samanta, M. NOD1 and NOD2 Receptors in Mrigal (Cirrhinus mrigala): Inductive Expression and Downstream Signalling in Ligand Stimulation and Bacterial Infections. J. Biosci. 2013, 38, 533–548. [Google Scholar] [CrossRef]
- Li, J.; Kong, L.; Gao, Y.; Wu, C.; Xu, T. Characterization of NLR-A Subfamily Members in Miiuy Croaker and Comparative Genomics Revealed NLRX1 Underwent Duplication and Lose in Actinopterygii. Fish Shellfish Immunol. 2015, 47, 397–406. [Google Scholar] [CrossRef]
- Aplin, A.C.; Ligresti, G.; Fogel, E.; Zorzi, P.; Smith, K.; Nicosia, R.F. Regulation of Angiogenesis, Mural Cell Recruitment and Adventitial Macrophage Behavior by Toll-like Receptors. Angiogenesis 2014, 17, 147–161. [Google Scholar] [CrossRef]
- Ni, H.; Zhao, W.; Kong, X.; Li, H.; Ouyang, J. Celastrol Inhibits Lipopolysaccharide-Induced Angiogenesis by Suppressing TLR4-Triggered Nuclear Factor-Kappa B Activation. Acta Haematol. 2014, 131, 102–111. [Google Scholar] [CrossRef]
- Eggestøl, H.; Lunde, H.S.; Knutsen, T.M.; Haugland, G.T. Interleukin-1 Ligands and Receptors in Lumpfish (Cyclopterus lumpus L.): Molecular Characterization, Phylogeny, Gene Expression, and Transcriptome Analyses. Front. Immunol. 2020, 11, 502. [Google Scholar] [CrossRef]
- Pelegrín, P.; García-Castillo, J.; Mulero, V.; Meseguer, J. Interleukin-1β Isolated from a Marine Fish Reveals up-Regulated Expression in Macrophages Following Activation with Lipopolysaccharide and Lymphokines. Cytokine 2001, 16, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Abos, B.; Wang, T.; Castro, R.; Granja, A.G.; Leal, E.; Havixbeck, J.; Luque, A.; Barreda, D.R.; Secombes, C.J.; Tafalla, C. Distinct Differentiation Programs Triggered by IL-6 and LPS in Teleost IgM+ B Cells in the Absence of Germinal Centers. Sci. Rep. 2016, 6, 30004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Bao, B.; He, Q.; Peatman, E.; He, C.; Liu, Z. Characterization and Expression Analysis of Bactericidal Permeability-Increasing Protein (BPI) Antimicrobial Peptide Gene from Channel Catfish Ictalurus punctatus. Dev. Comp. Immunol. 2005, 29, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Liang, Y.; Jiang, Y.; Liu, S.; Zhang, F.; He, X.; Wang, T.; Zhou, Y.; Zhong, H.; Yan, J. Identification and Expression Analysis on Bactericidal Permeability-Increasing Protein/Lipopolysaccharide-Binding Protein of Blunt Snout Bream, Megalobrama amblycephala. Fish Shellfish Immunol. 2015, 45, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, S.; Qi, J.; Echtenkamp, S.F.; Chatterjee, R.; Wang, M.; Boons, G.-J.; Dziarski, R.; Gupta, D. Zebrafish Peptidoglycan Recognition Proteins Are Bactericidal Amidases Essential for Defense against Bacterial Infections. Immunity 2007, 27, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Shan, S.; Xu, X.; Wang, Y.; Zhang, Y.; Yin, M.; Yang, G. Molecular Characterization and Expression Analysis of Two Peptidoglycan Recognition Proteins (CcPGRP5, CcPGRP6) in Larvae Ontogeny of Common Carp Cyprinus carpio L. and upon Immune Stimulation by Bacteria. BMC Vet. Res. 2019, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Briolat, V.; Jouneau, L.; Carvalho, R.; Palha, N.; Langevin, C.; Herbomel, P.; Schwartz, O.; Spaink, H.P.; Levraud, J.-P.; Boudinot, P. Contrasted Innate Responses to Two Viruses in Zebrafish: Insights into the Ancestral Repertoire of Vertebrate IFN-Stimulated Genes. J. Immunol. 2014, 192, 4328–4341. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Li, B.; Wu, L.; Yin, X.; Zhong, X.; Li, Y.; Wang, Y.; Guo, Z.; Ye, J. Interleukin-6 Gets Involved in Response to Bacterial Infection and Promotes Antibody Production in Nile Tilapia (Oreochromis niloticus). Dev. Comp. Immunol. 2018, 89, 141–151. [Google Scholar] [CrossRef]
- Liu, Q.-N.; Xin, Z.-Z.; Chai, X.-Y.; Jiang, S.-H.; Li, C.-F.; Zhang, H.-B.; Ge, B.-M.; Zhang, D.-Z.; Zhou, C.-L.; Tang, B.-P. Characterization of Immune-Related Genes in the Yellow Catfish Pelteobagrus fulvidraco in Response to LPS Challenge. Fish Shellfish Immunol. 2016, 56, 248–254. [Google Scholar] [CrossRef]
- Gauthier, A.E.; Chandler, C.E.; Poli, V.; Gardner, F.M.; Tekiau, A.; Smith, R.; Bonham, K.S.; Cordes, E.E.; Shank, T.M.; Zanoni, I.; et al. Deep-Sea Microbes as Tools to Refine the Rules of Innate Immune Pattern Recognition. Sci. Immunol. 2021, 6, eabe0531. [Google Scholar] [CrossRef]
- Gurung, P.; Lukens, J.R.; Kanneganti, T.-D. Mitochondria: Diversity in the Regulation of the NLRP3 Inflammasome. Trends Mol. Med. 2015, 21, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-Y.; Wang, Y.-Y.; Shao, T.; Fan, D.-D.; Lin, A.-F.; Xiang, L.-X.; Shao, J.-Z. The Zebrafish NLRP3 Inflammasome Has Functional Roles in ASC-Dependent Interleukin-1β Maturation and Gasdermin E–Mediated Pyroptosis. J. Biol. Chem. 2020, 295, 1120–1141. [Google Scholar] [CrossRef]
- Reyes-Cerpa, S.; Maisey, K.; Reyes-Lpez, F.; Toro-Ascuy, D.; Mara, A.; Imarai, M. Fish Cytokines and Immune Response. In New Advances and Contributions to Fish Biology; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Lulijwa, R.; Alfaro, A.C.; Merien, F.; Burdass, M.; Venter, L.; Young, T. In Vitro Immune Response of Chinook Salmon (Oncorhynchus tshawytscha) Peripheral Blood Mononuclear Cells Stimulated by Bacterial Lipopolysaccharide. Fish Shellfish Immunol. 2019, 94, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.Y.; Cho, S.H.; Kwon, S.H.; Eom, C.Y.; Jeong, M.S.; Lee, W.W.; Kim, S.Y.; Heo, S.J.; Ahn, G.; Lee, K.P.; et al. The Roles of NF-ΚB and ROS in Regulation of pro-Inflammatory Mediators of Inflammation Induction in LPS-Stimulated Zebrafish Embryos. Fish Shellfish Immunol. 2017, 68, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, S.; Pulpipat, T.; Wang, P.C.; Chen, S.C. Transcriptome Analysis of Immune- and Iron-Related Genes after Francisella noatunensis Subsp. Orientalis Infection in Nile Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 111, 36–48. [Google Scholar] [CrossRef]
- Qin, C.; Zhao, W.; Deng, D.; Lu, R.; Yang, G.; Yan, X.; Meng, X.; Nie, G. Effects of Glucose, Insulin, Glucagon, LPS and Poly (I:C) on Adiponectin Receptors Expression in Grass Carp (Ctenophcuyngodon idellus). Aquaculture 2020, 528, 735501. [Google Scholar] [CrossRef]
- Xie, J.; Chen, X.; He, X.; Niu, J. Cloning and Expression Analysis of Keap1 of Golden Pompano (Trachinotus ovatus) and Response to Oxidized Fish Oil and LPS Administration. Aquac. Rep. 2020, 18, 100527. [Google Scholar] [CrossRef]
- Congleton, J.L.; Wagner, E.J. Acute-Phase Hypoferremic Response to Lipopolysaccharide in Rainbow Trout (Oncorhynchus mykiss). Comp. Biochem. Physiol.-Part A Physiol. 1991, 98, 195–200. [Google Scholar] [CrossRef]
- Freedman, A.H.; Clamp, M.; Sackton, T.B. Error, Noise and Bias in de Novo Transcriptome Assemblies. Mol. Ecol. Resour. 2021, 21, 18–29. [Google Scholar] [CrossRef]


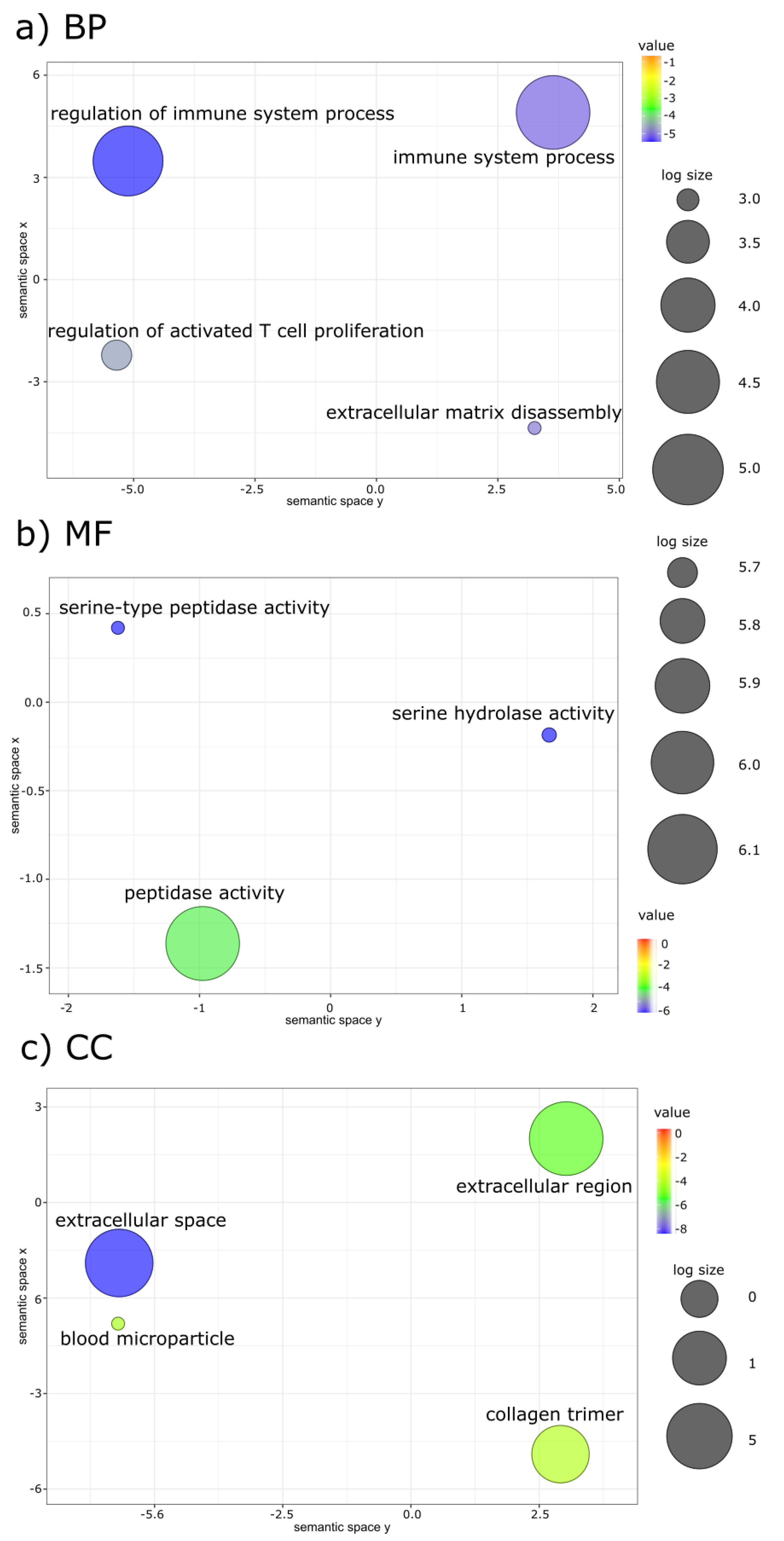
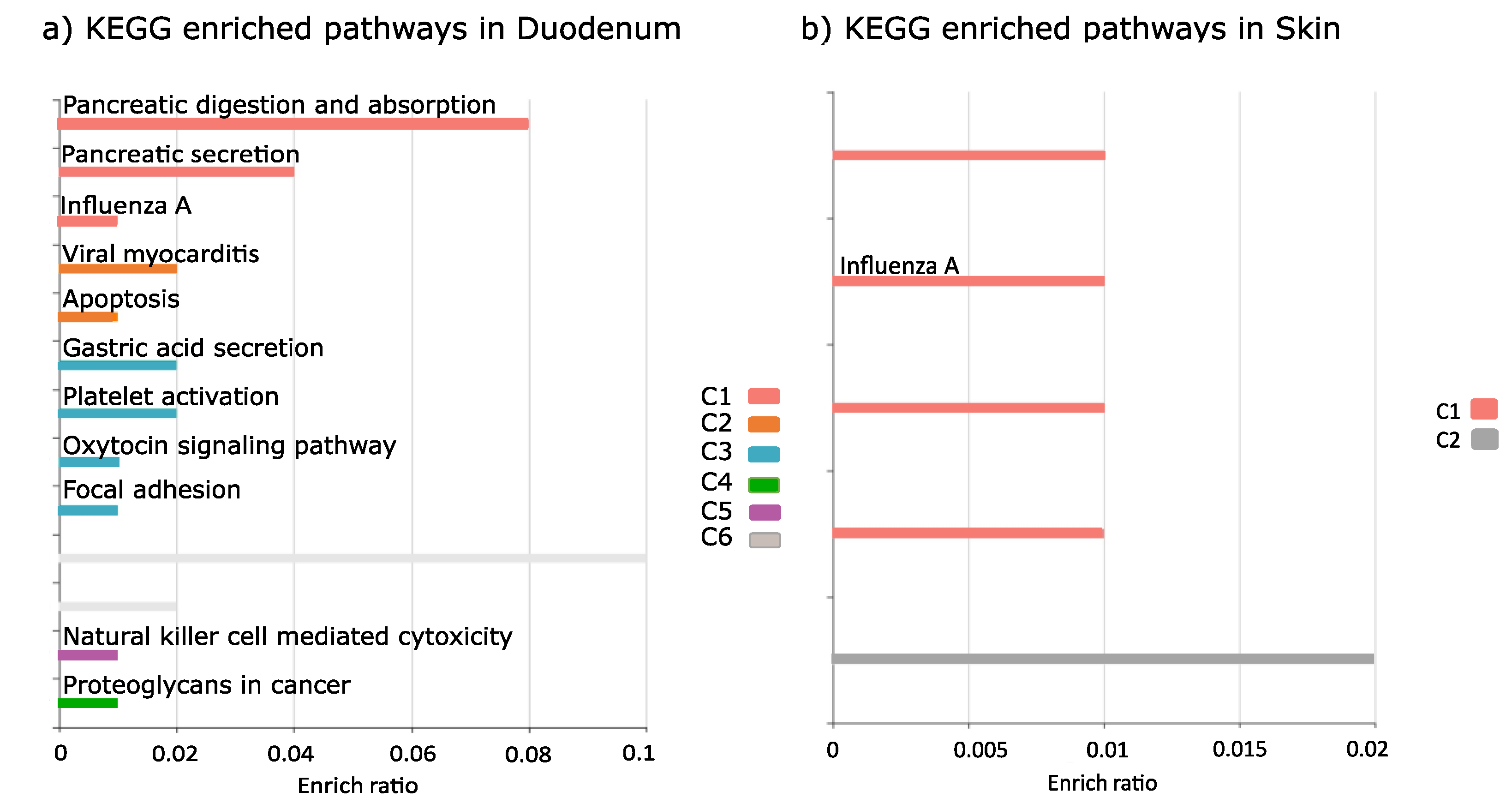

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TRINITY ID | logFC | UniProtKB ID | Gene Definition | Gene Symbol |
|---|---|---|---|---|
| Head-kidney | ||||
| TRINITY_DN159360_c4_g1 | 9.78 | F1RAW5 | Shootin 3 | shtn3 |
| TRINITY_DN158576_c1_g3 | 8.91 | N/A | Zinc-binding protein A33-like | za33l |
| TRINITY_DN163380_c1_g1 | 8.84 | Q8ND71 | GTPase IMAP family member 8 | gimap8 |
| TRINITY_DN159230_c9_g2 | 8.51 | Q9Y4D1 | Disheveled-associated activator of morphogenesis 1 | daam1 |
| TRINITY_DN142659_c1_g6 | 8.40 | A8MUP2 | Methyltransferase-like protein 12, mitochondrial | cskmt |
| TRINITY_DN159639_c0_g1 | 8.38 | Q01130 | Serine and arginine rich splicing factor 2a | srsf2 |
| TRINITY_DN144517_c5_g2 | 8.25 | Q9Y3Z3 | SAM domain and HD domain 1 | samhd1 |
| TRINITY_DN137183_c4_g1 | 8.19 | Q8WWR8 | Sialidase 4 | neu4 |
| TRINITY_DN122607_c0_g1 | 7.94 | Q9NV96 | Cell cycle control protein 50A-like | tmem30a |
| TRINITY_DN155301_c3_g7 | 7.86 | Q6ZSJ9 | Protein shisa-6 | shisa6 |
| TRINITY_DN148291_c0_g2 | −2.18 | P62241 | 40S ribosomal protein S8 | rps |
| TRINITY_DN143420_c2_g1 | −2.43 | O60462 | Neuropilin 2 | nrp2 |
| TRINITY_DN155536_c2_g1 | −2.94 | Q53G44 | Interferon-induced protein 44-like | ifi44l |
| TRINITY_DN175530_c0_g1 | −3.21 | Q969X5 | Endoplasmic reticulum-golgi intermediate compartment 1 | ergic1 |
| TRINITY_DN162708_c0_g2 | −3.34 | O95398 | Rap guanine nucleotide exchange factor (GEF) 3 | rapgef3 |
| TRINITY_DN142663_c1_g1 | −3.86 | N/A | Peptidoglycan-recognition protein LB-like | pgrp-lbl |
| TRINITY_DN149357_c4_g1 | −3.89 | N/A | Galactose-specific lectin nattectin-like | lgals3l |
| TRINITY_DN140144_c4_g1 | −4.14 | Q6IMW7 | Parvalbumin 4 | pvalb4 |
| TRINITY_DN156139_c0_g1 | −1.07 × 10 | P17213 | Bactericidal permeability-increasing protein | bpi |
| TRINITY_DN157841_c3_g1 | −1.44 × 10 | Q7RTR2 | NLR family CARD domain-containing protein 3 | nlrc3 |
| Duodenum | ||||
| TRINITY_DN153528_c0_g2 | 9.89 | P07478 | Trypsin-2 | prss2 |
| TRINITY_DN160991_c2_g2 | 9.70 | Q7TST0 | Butyrophilin-like protein 1 | btnl1 |
| TRINITY_DN153528_c0_g3 | 9.66 | P35030 | Trypsin-3 | prss3 |
| TRINITY_DN123020_c0_g1 | 9.47 | Q9UNI1 | Chymotrypsin-like elastase family, member 1 | cela1 |
| TRINITY_DN134620_c1_g2 | 9.39 | E7F0X0 | High choriolytic enzyme 1-like | hce2l2 |
| TRINITY_DN151930_c0_g3 | 9.36 | P01833 | Polymeric immunoglobulin receptor | pigr |
| TRINITY_DN152124_c5_g2 | 9.15 | Q9BQ24 | Zinc finger, FYVE domain containing 21 | zfyve21 |
| TRINITY_DN140296_c3_g2 | 9.13 | P08217 | Chymotrypsin-like elastase family member 2A | cela2a |
| TRINITY_DN153528_c0_g1 | 9.09 | P07477 | Trypsin 1 | prss1 |
| TRINITY_DN138531_c1_g2 | 8.88 | Q7Z2Y8 | Interferon-induced very large GTPase 1 | gvinp1 |
| TRINITY_DN155812_c1_g1 | −2.07 | Q7Z6B7 | SLIT-ROBO Rho GTPase activating protein 1 | srgap1 |
| TRINITY_DN162540_c4_g1 | −2.68 | Q16653 | Myelin-oligodendrocyte glycoprotein | mog |
| TRINITY_DN92606_c0_g1 | −2.94 | P09382 | Beta-galactoside-binding lectin | lgals1 |
| TRINITY_DN153065_c2_g2 | −2.97 | P18847 | Activating transcription factor 3 | atf3 |
| TRINITY_DN6238_c0_g1 | −3.02 | P01344 | Insulin-like growth factor 2 | igf2 |
| TRINITY_DN135989_c1_g5 | −3.07 | Q9NPI9 | Inward rectifier potassium channel 16 | kcnj16 |
| TRINITY_DN139165_c1_g1 | −3.37 | Q01995 | Transgelin | tagln |
| Skin | ||||
| TRINITY_DN159639_c0_g1 | 8.87 | U3JA80 | Serine and arginine rich splicing factor 2a | srsf2a |
| TRINITY_DN150109_c2_g2 | 8.44 | P07477 | Cationic trypsin | prss1 |
| TRINITY_DN141237_c0_g7 | 7.65 | Q14863 | POU domain, class 6, transcription factor 1-like | pou6d1 |
| TRINITY_DN137203_c7_g3 | 7.10 | A0A1S3L397 | Nuclear factor 7, brain-like | nf7bl |
| TRINITY_DN139257_c0_g1 | 4.05 | Q96C55 | Zinc finger protein 524 | znf524 |
| TRINITY_DN162280_c2_g2 | 3.26 | Q96A28 | SLAM family member 9 | slamf9 |
| TRINITY_DN129892_c0_g1 | 3.15 | Q7Z2Y8 | Interferon-induced very large GTPase 1 | gvinp1 |
| TRINITY_DN158576_c1_g3 | −3.94 | N/A | Zinc-binding protein A33-like | za33l |
| TRINITY_DN159712_c1_g3 | −7.06 | Q96P20 | NACHT, LRR and PYD domains-containing protein 3 | nlrp3 |
| TRINITY_DN143097_c2_g4 | −7.44 | Q13216 | Excision repair cross-complementation group 8 | ercc8 |
| TRINITY_DN159613_c2_g1 | −7.60 | Q14258 | E3 ubiquitin/ISG15 ligase TRIM25 | trim25 |
| TRINITY_DN158530_c2_g1 | −7.63 | Q6BDS2 | UHRF1 binding protein 1 | uhrf1bp1 |
| TRINITY_DN149926_c4_g3 | −7.68 | Q9PWM2 | Homeobox B9a | hxb9a |
| TRINITY_DN151272_c3_g5 | −7.81 | N/A | Cytochrome P450 2K1-like | cy2k1l |
| TRINITY_DN151209_c9_g1 | −7.85 | Q7RTR2 | NLR family CARD domain-containing protein 3 | nlrc3 |
| TRINITY_DN156424_c1_g2 | −8.13 | Q8NHV1 | GTPase IMAP family member 7 | gimap7 |
| TRINITY_DN137852_c4_g3 | −8.71 | Q9UGT4 | Sushi domain containing 2 | susd2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, C.S.V.; Power, D.M.; Guerreiro, P.M.; Louro, B.; Chen, L.; Canário, A.V.M. Transcriptomic Down-Regulation of Immune System Components in Barrier and Hematopoietic Tissues after Lipopolysaccharide Injection in Antarctic Notothenia coriiceps. Fishes 2022, 7, 171. https://doi.org/10.3390/fishes7040171
Sousa CSV, Power DM, Guerreiro PM, Louro B, Chen L, Canário AVM. Transcriptomic Down-Regulation of Immune System Components in Barrier and Hematopoietic Tissues after Lipopolysaccharide Injection in Antarctic Notothenia coriiceps. Fishes. 2022; 7(4):171. https://doi.org/10.3390/fishes7040171
Chicago/Turabian StyleSousa, Cármen S. V., Deborah M. Power, Pedro M. Guerreiro, Bruno Louro, Liangbiao Chen, and Adelino V. M. Canário. 2022. "Transcriptomic Down-Regulation of Immune System Components in Barrier and Hematopoietic Tissues after Lipopolysaccharide Injection in Antarctic Notothenia coriiceps" Fishes 7, no. 4: 171. https://doi.org/10.3390/fishes7040171
APA StyleSousa, C. S. V., Power, D. M., Guerreiro, P. M., Louro, B., Chen, L., & Canário, A. V. M. (2022). Transcriptomic Down-Regulation of Immune System Components in Barrier and Hematopoietic Tissues after Lipopolysaccharide Injection in Antarctic Notothenia coriiceps. Fishes, 7(4), 171. https://doi.org/10.3390/fishes7040171