
Utilization of Fishery-Processing By-Product Squid Pens for Scale-Up Production of Phenazines via Microbial Conversion and Its Novel Potential Antinematode Effect


 , ,
, ,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Phenazine Production via Microbial Fermentation Experiments in Small Flasks
2.2.2. Scale-Up Production of Phenazines via Using a 14 L Bioreactor System
2.2.3. Quantitives and Purification of Phenazines
2.3. Assay of Nematodes Inhibition
2.4. Statistical Analysis
3. Results and Discussion
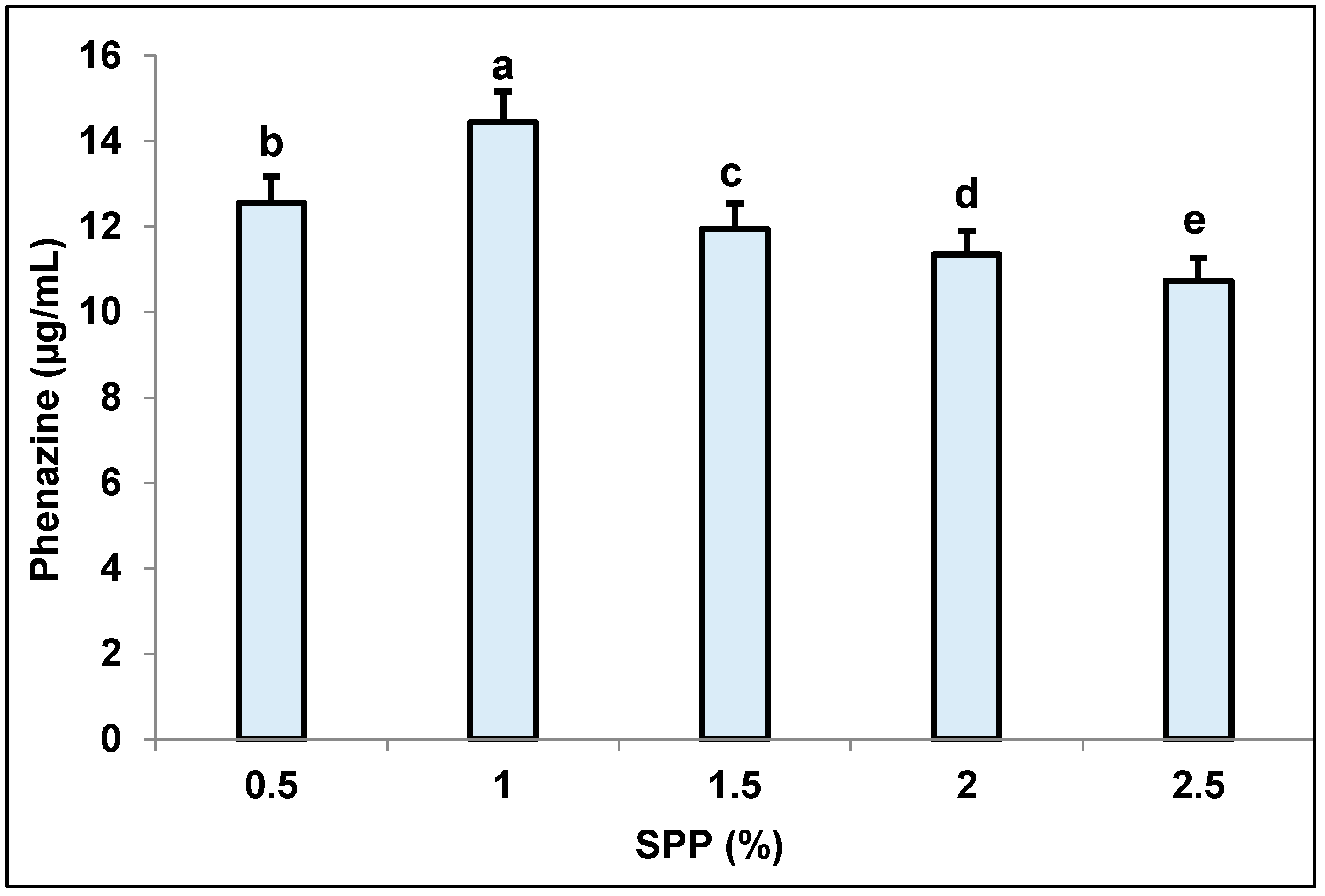
3.1. New Records of Fishery-Processing By-Product Squid Pens as Sole Carbon/Nitrogen Source for Phenazine Production via Fermentation
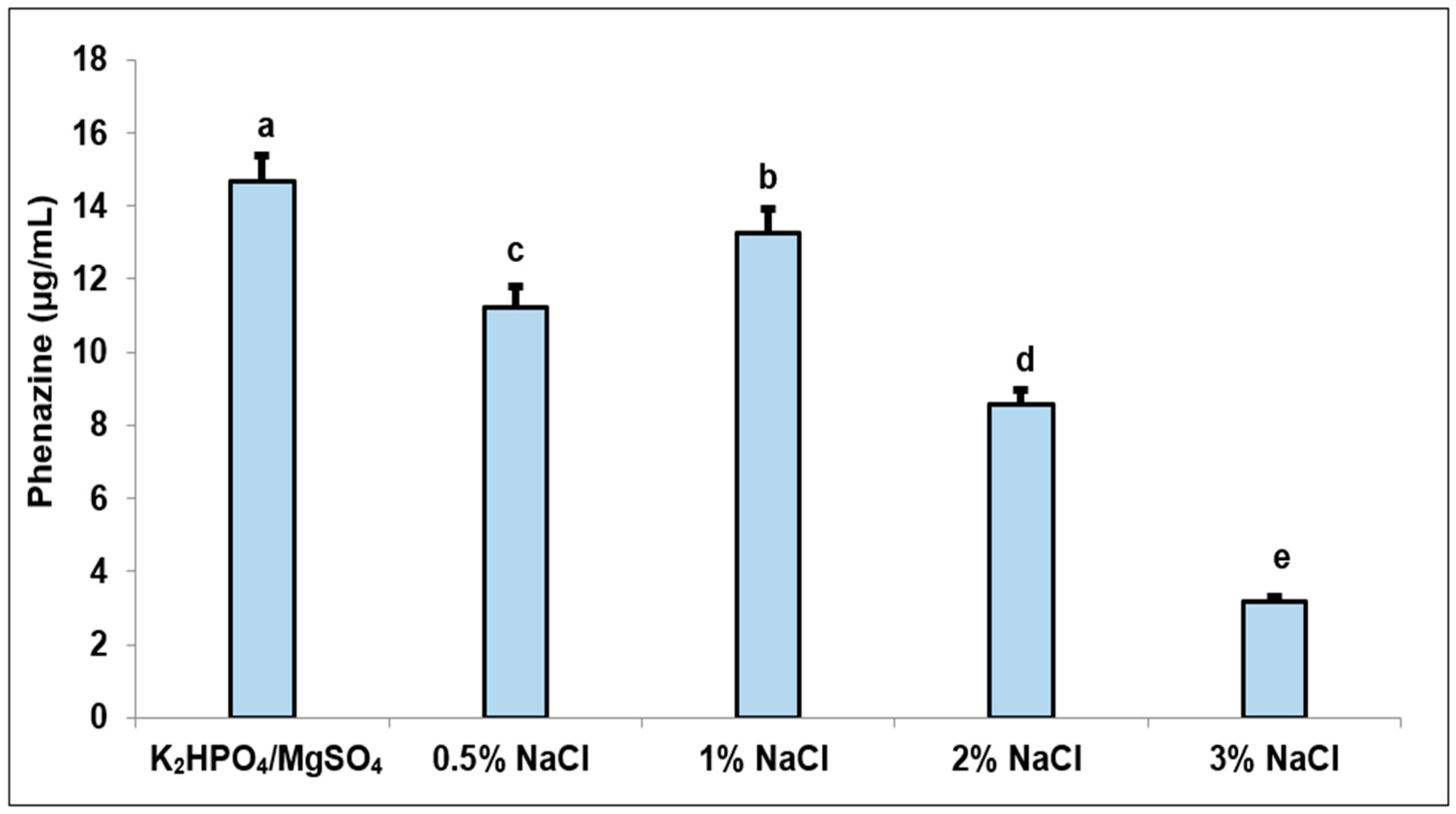
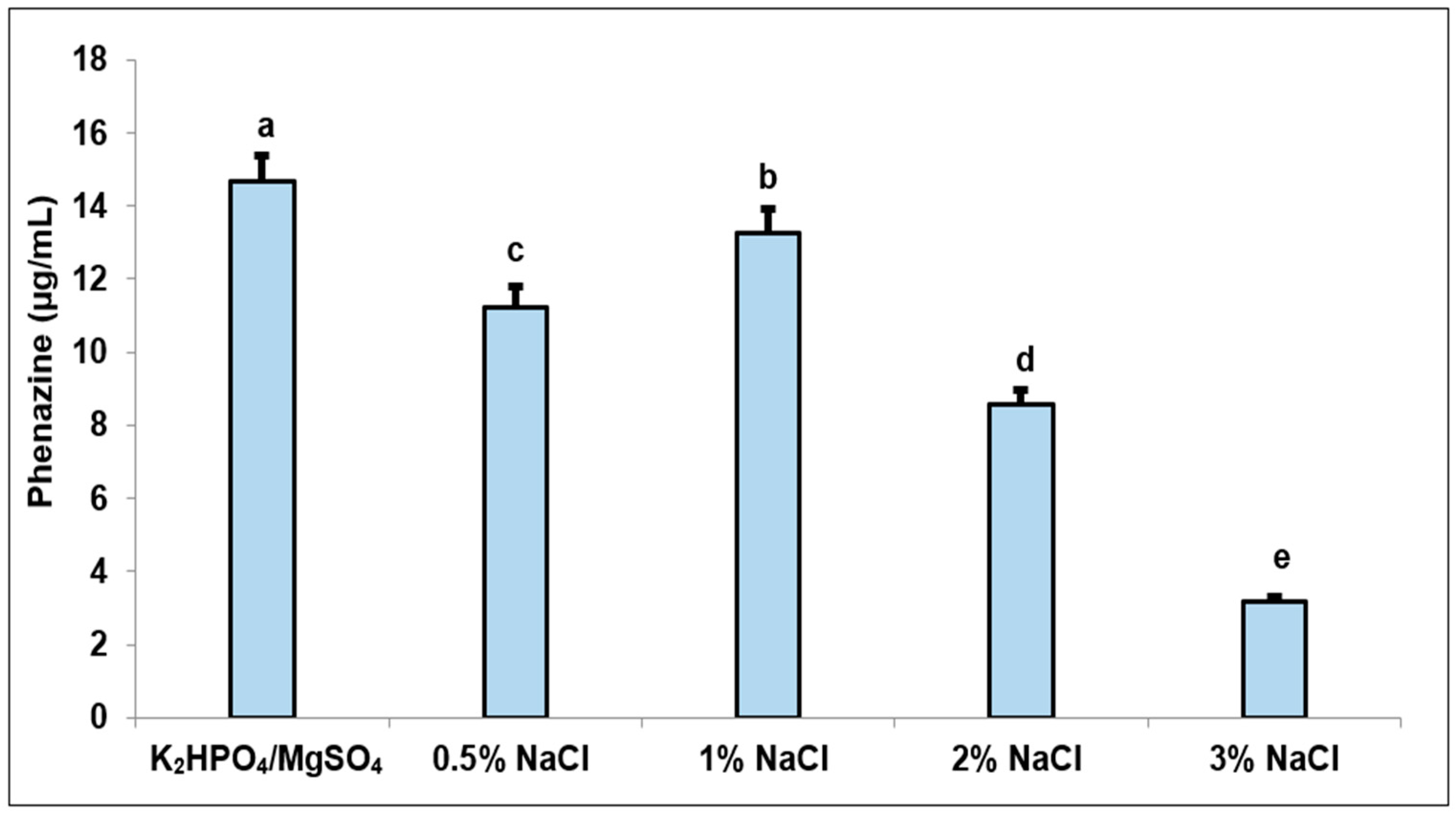
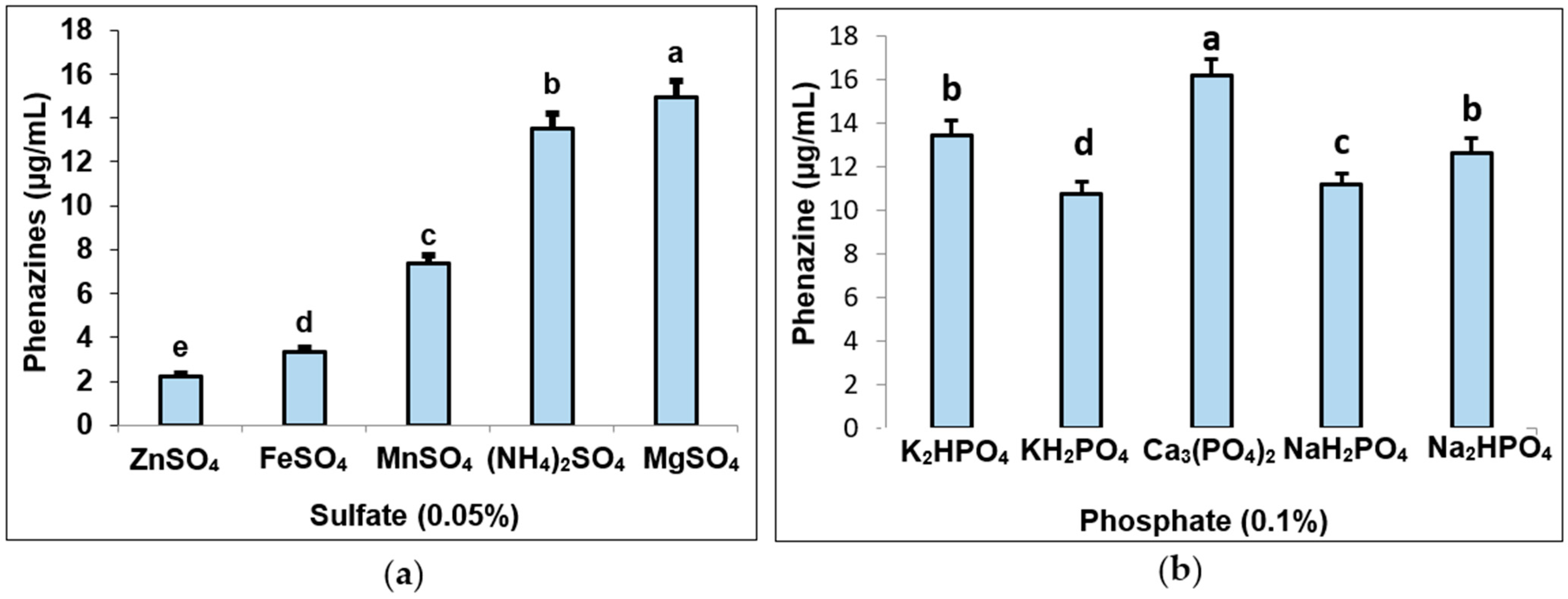
3.2. The Effect of Salt Composition on Phenazine Productivity in Fermentation
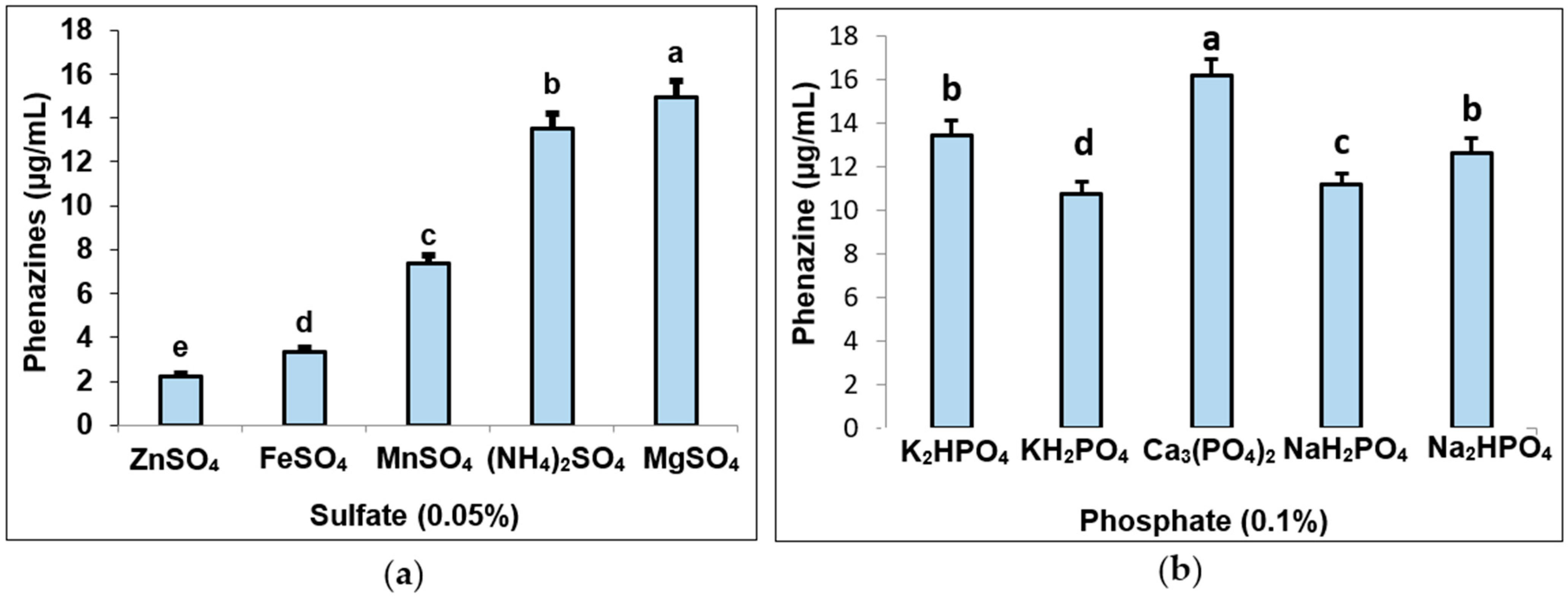
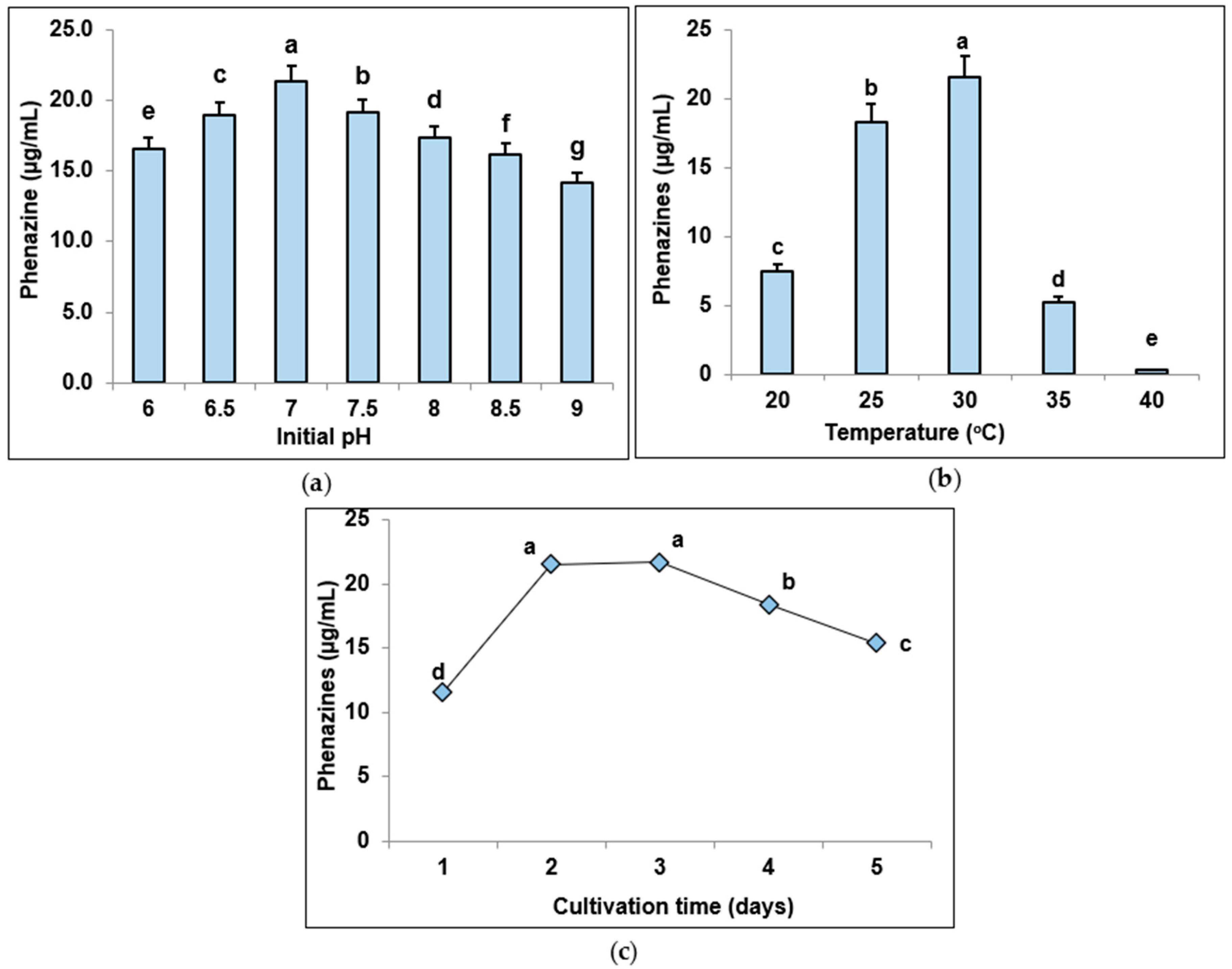
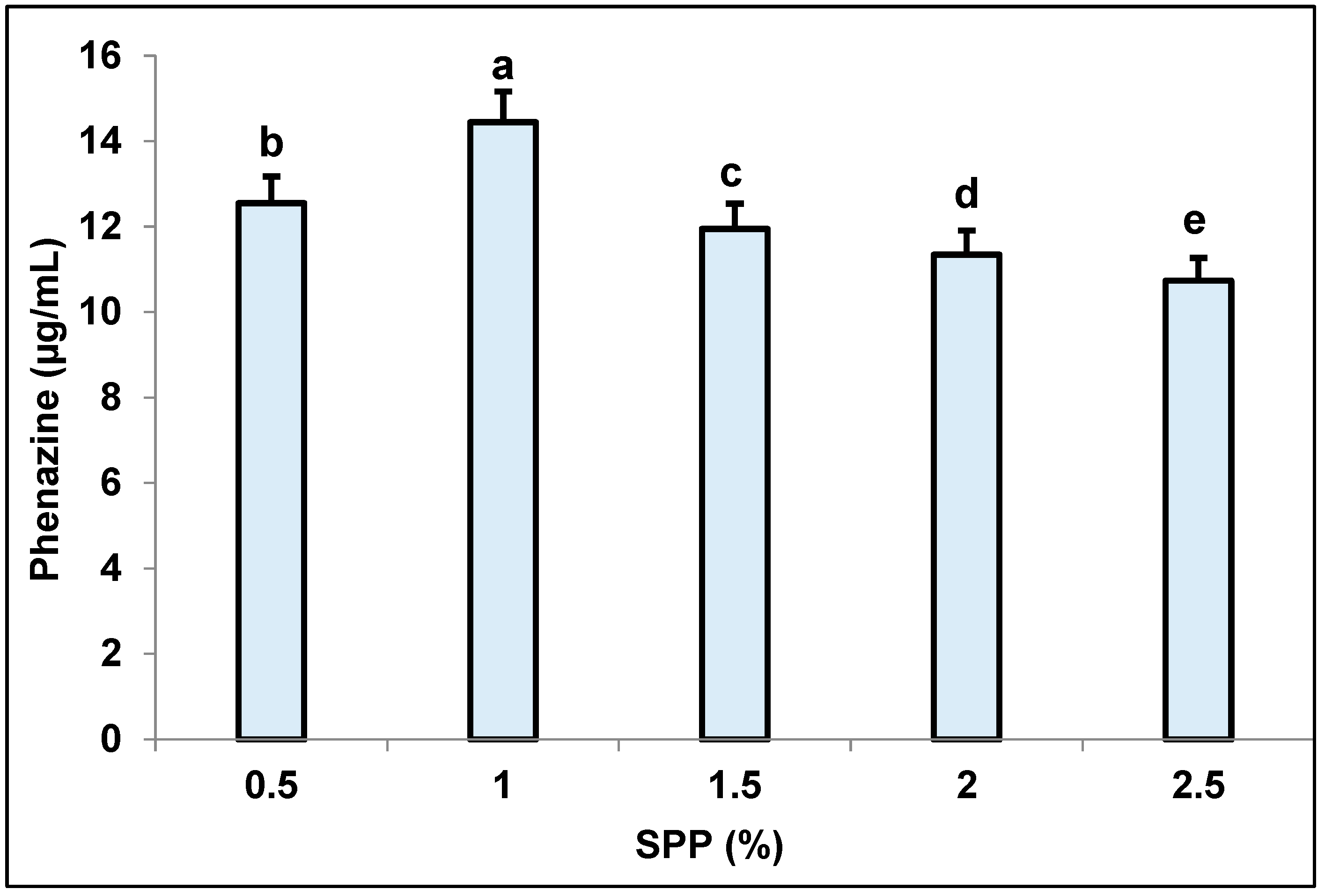
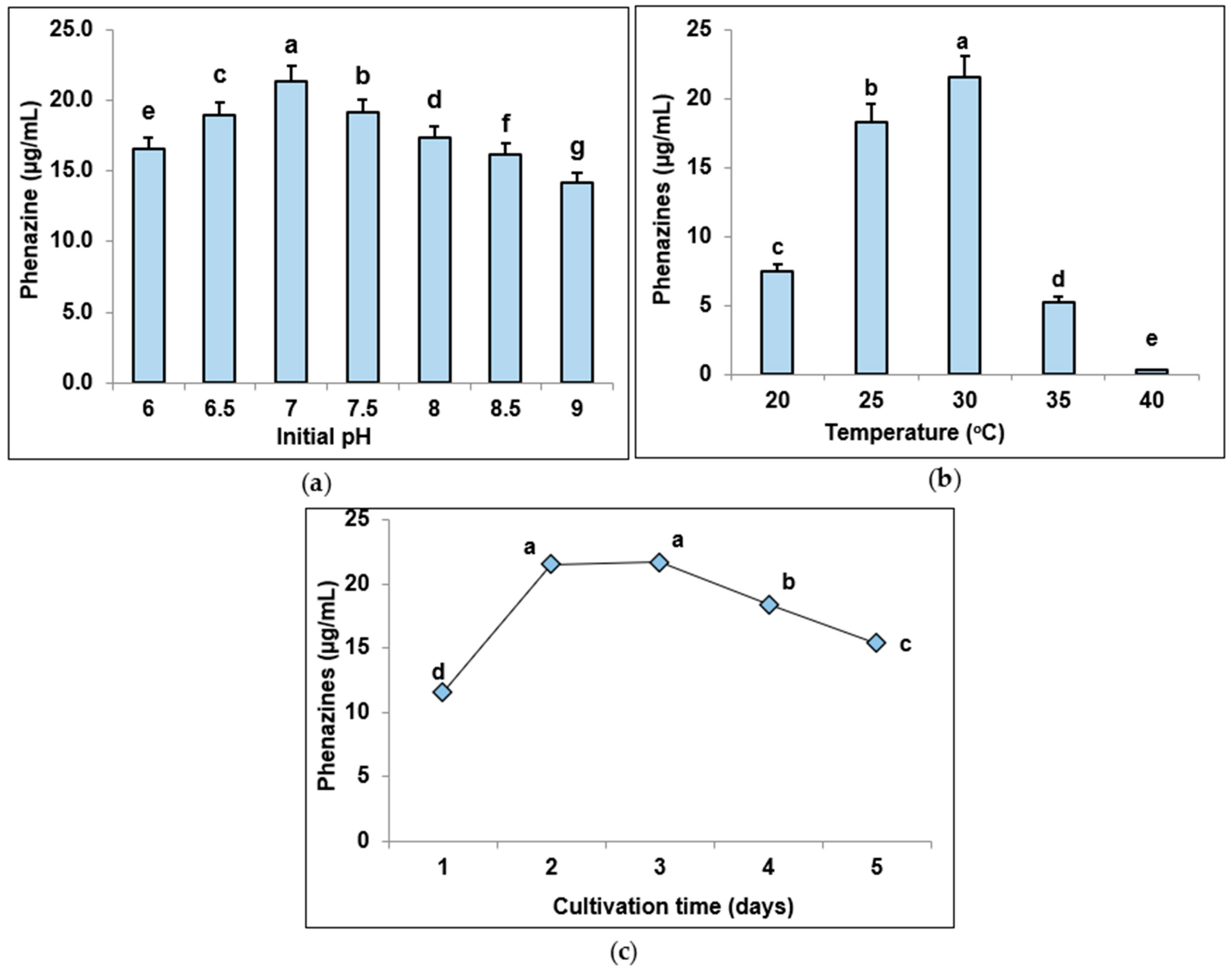
3.3. Determination of Suitable Culture Parameters for Pigment Biosynthesis
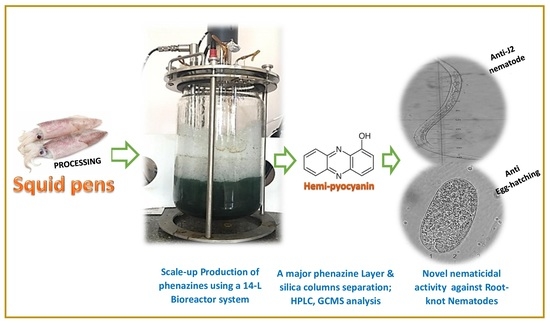
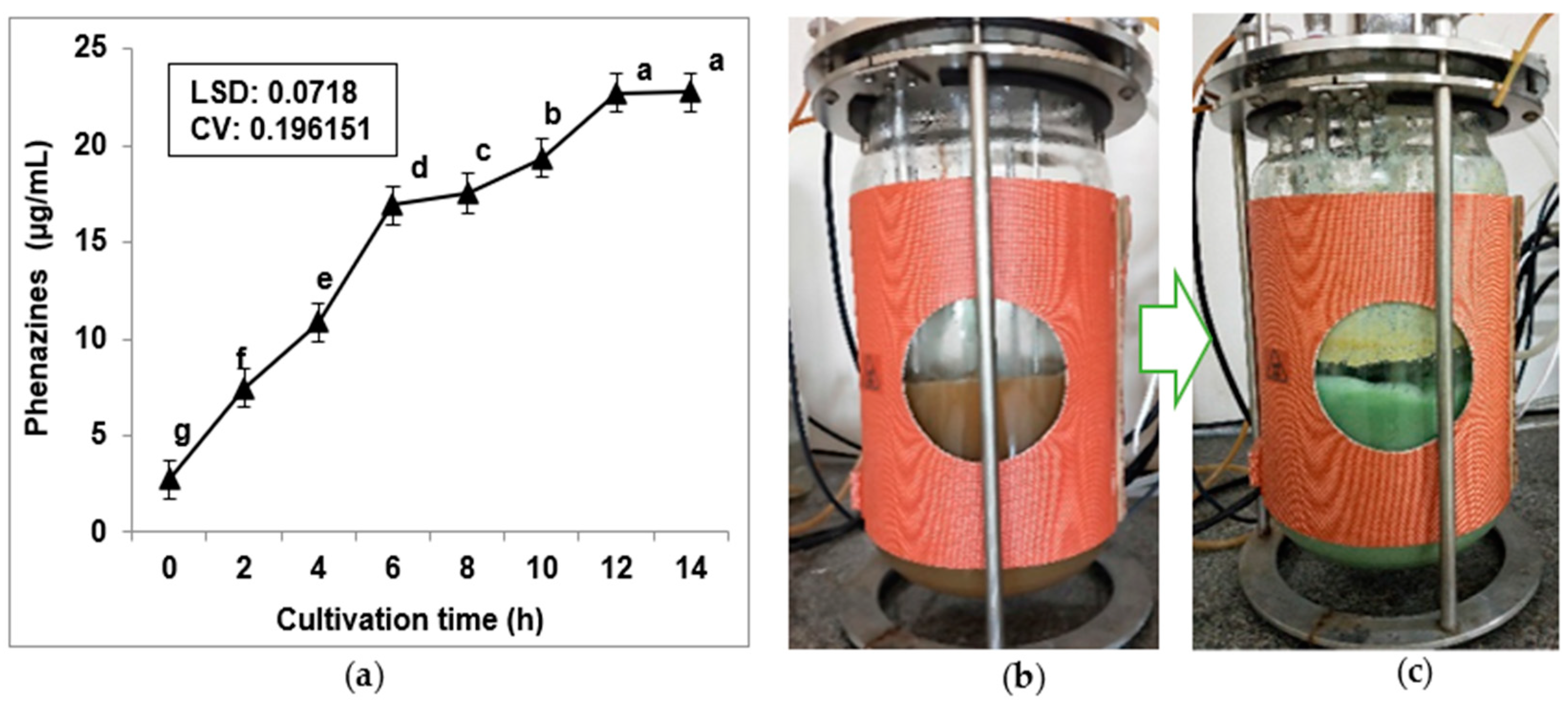
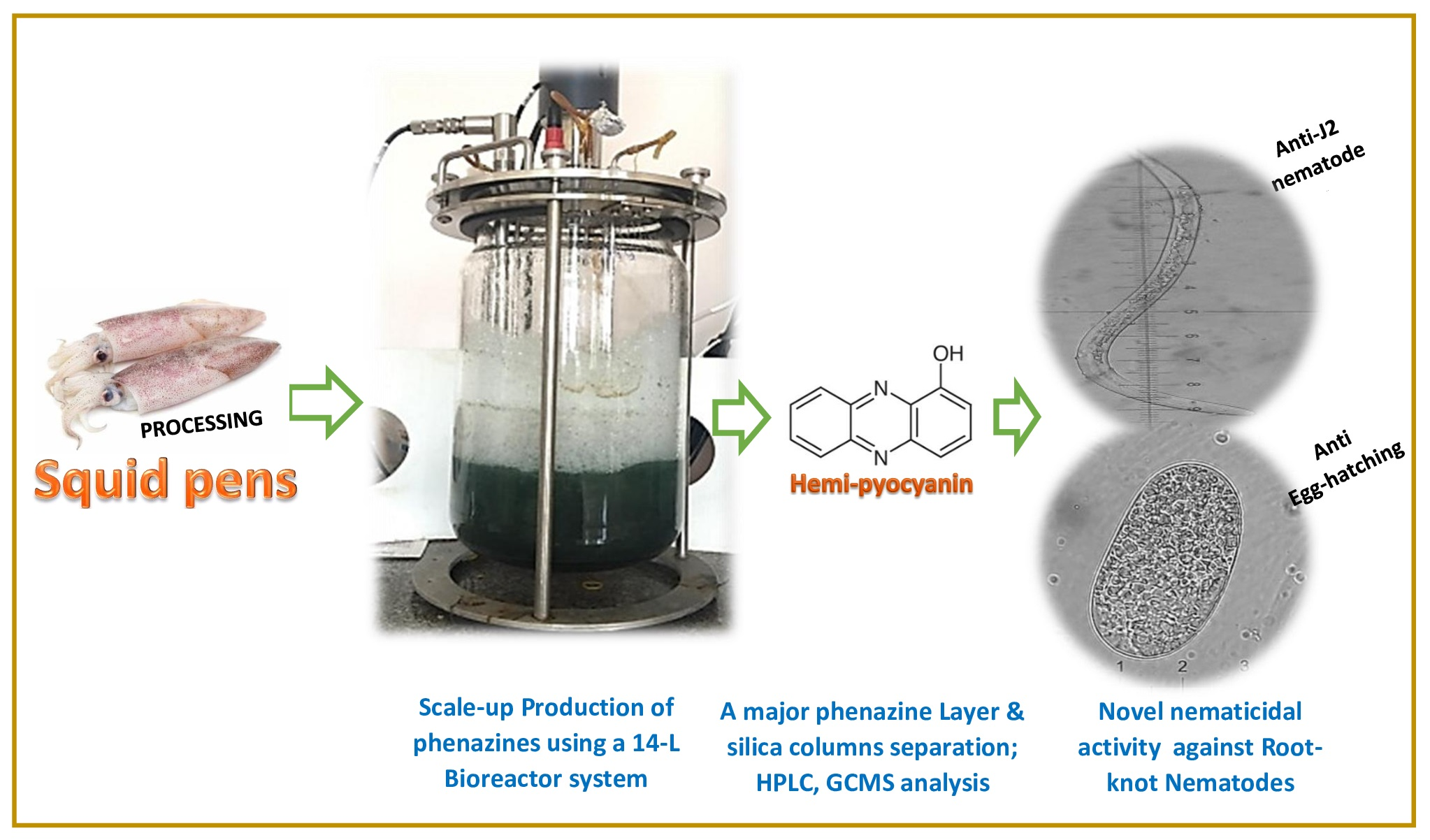
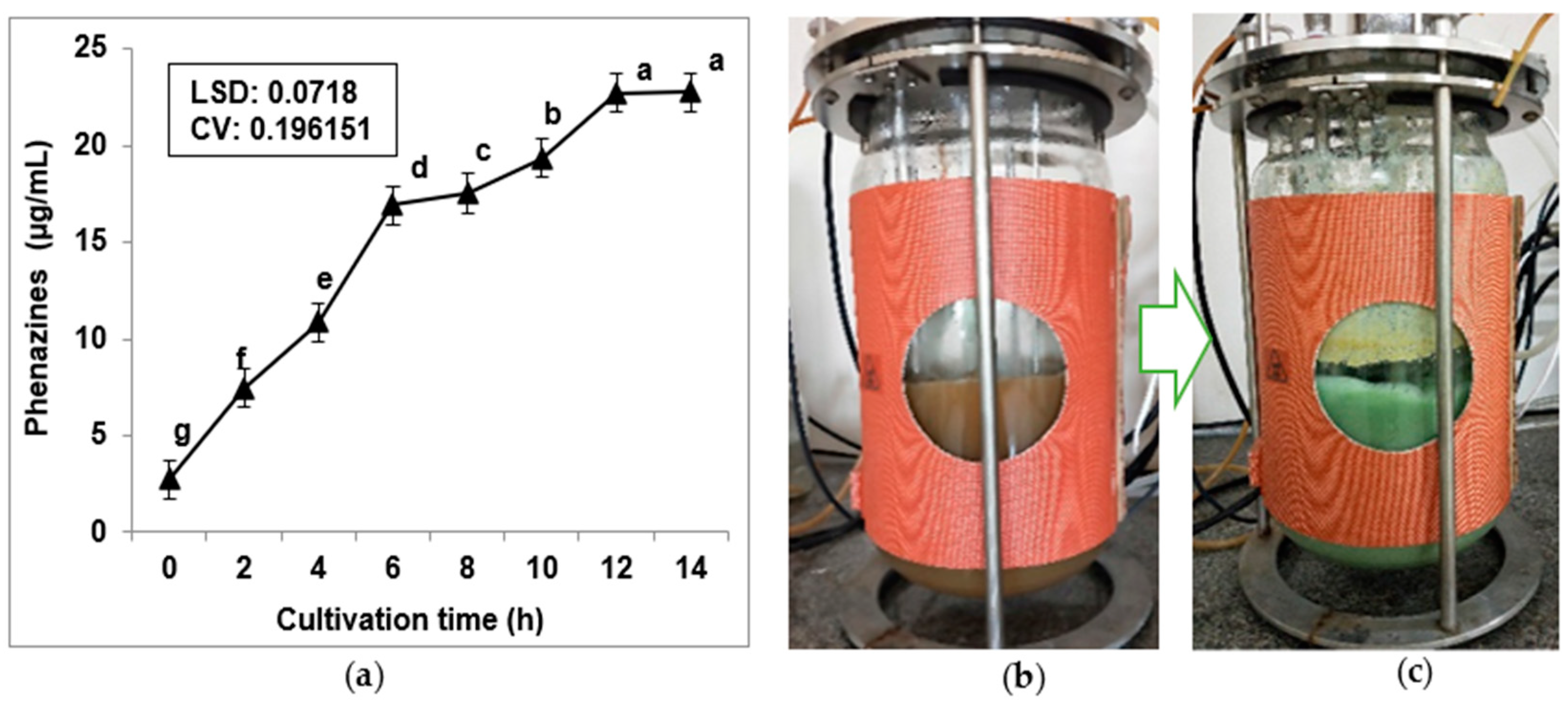
3.4. Scale-Up Production of Phenazines via Using a 14 L Bioreactor System and Purification of Phenazines
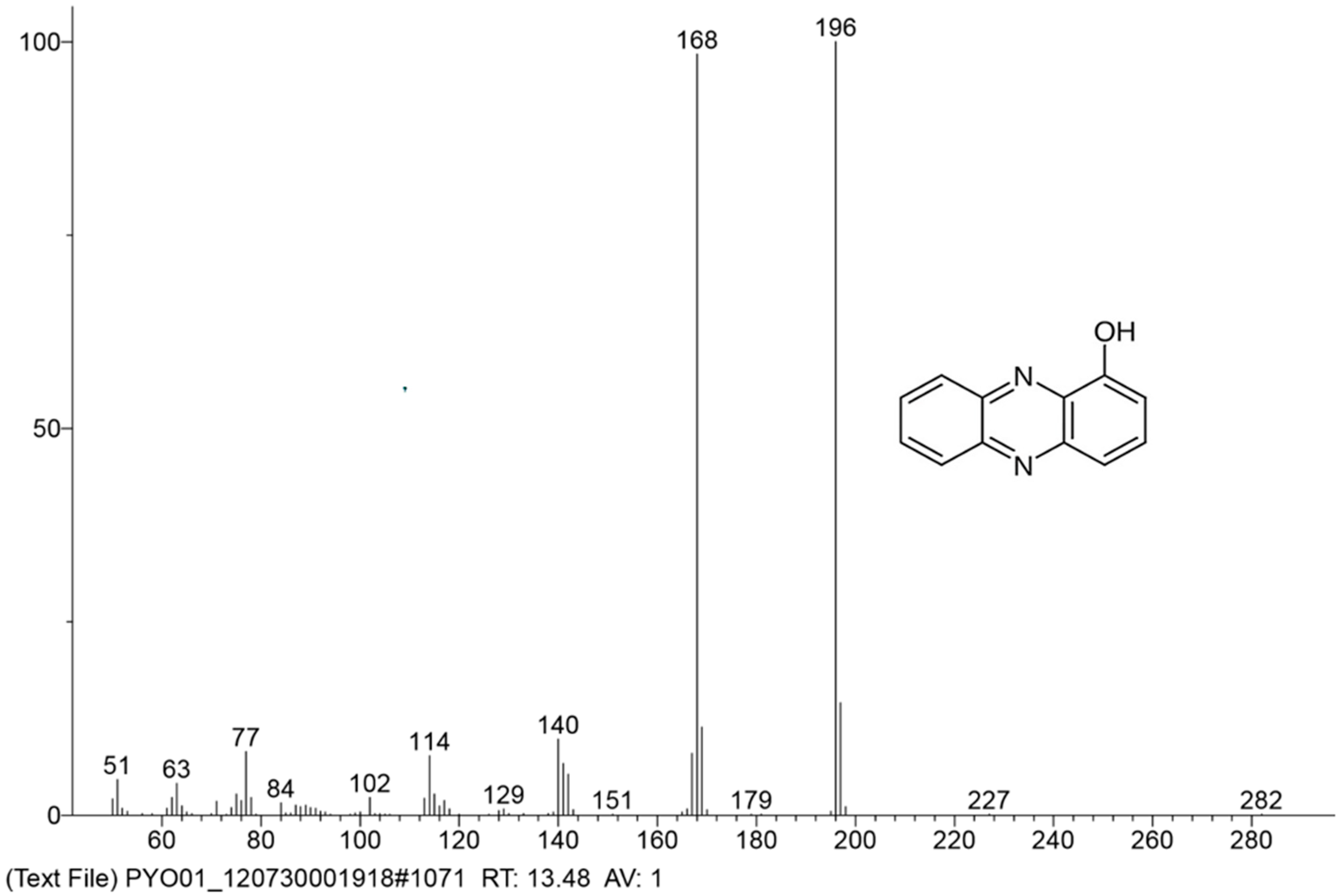
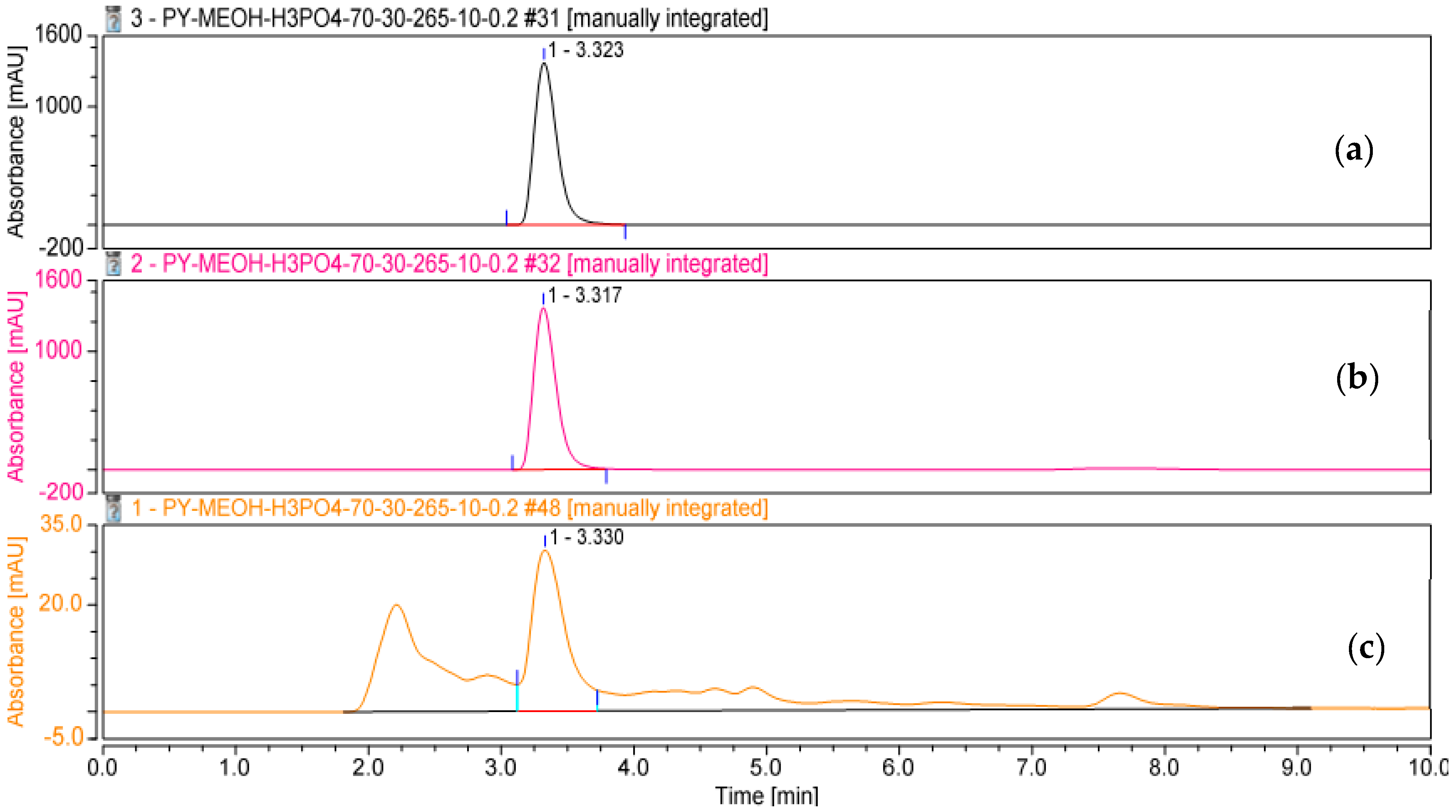
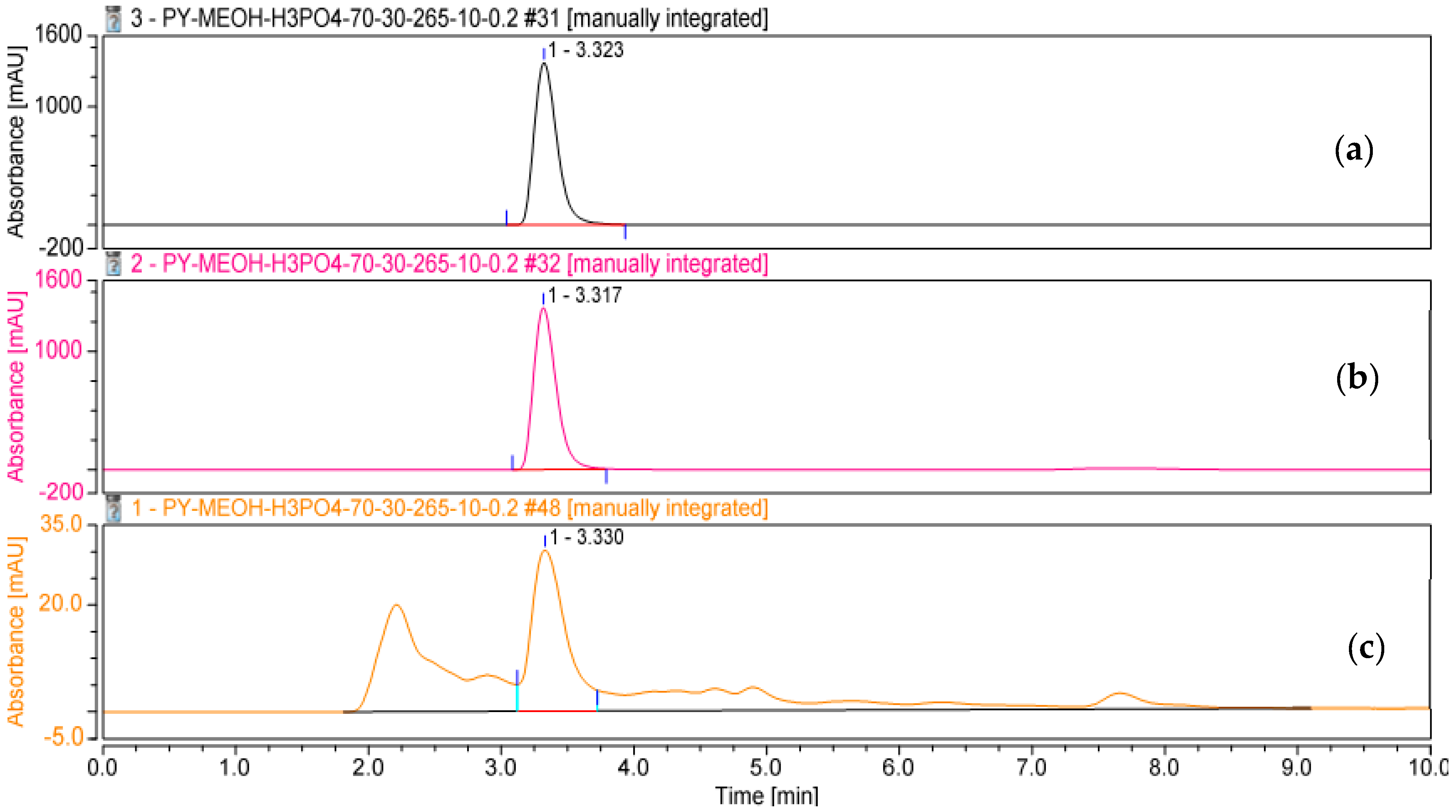
3.5. Determination, Extraction, and Purification of the Main Phenazine Compound from the Culture Broth
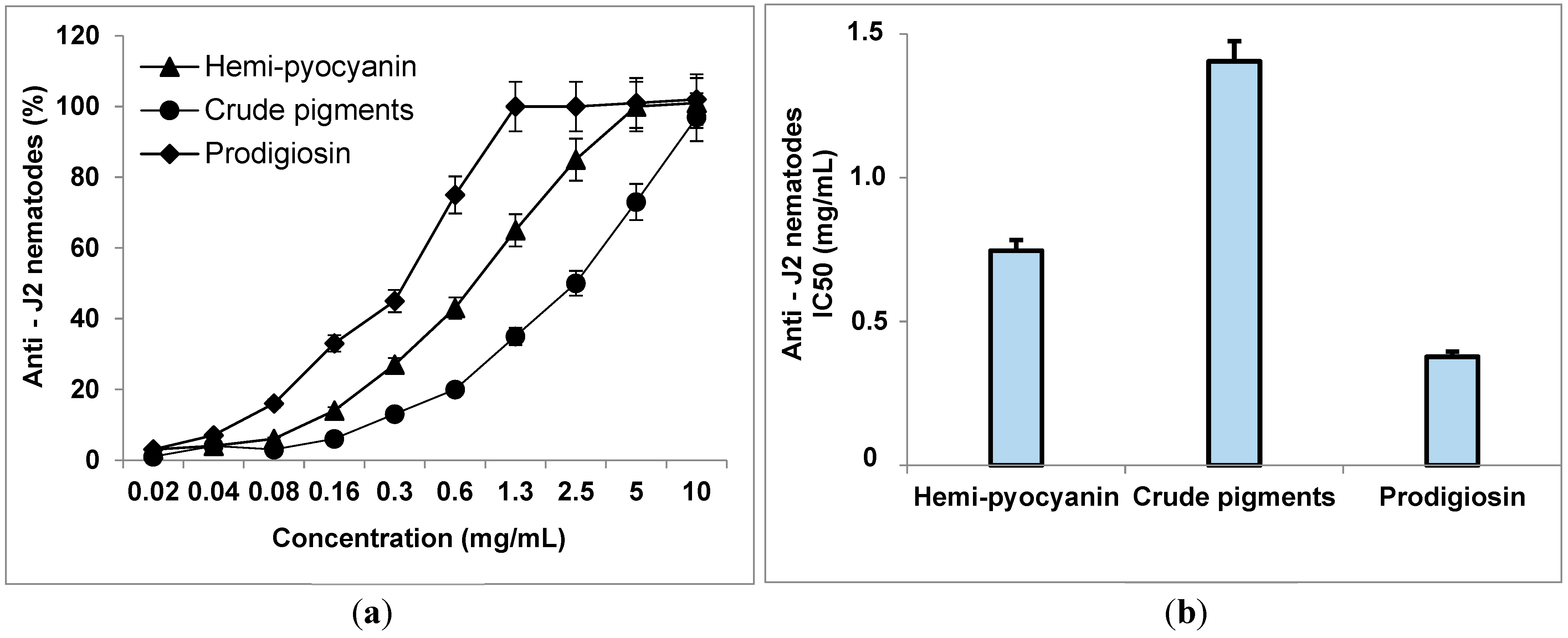
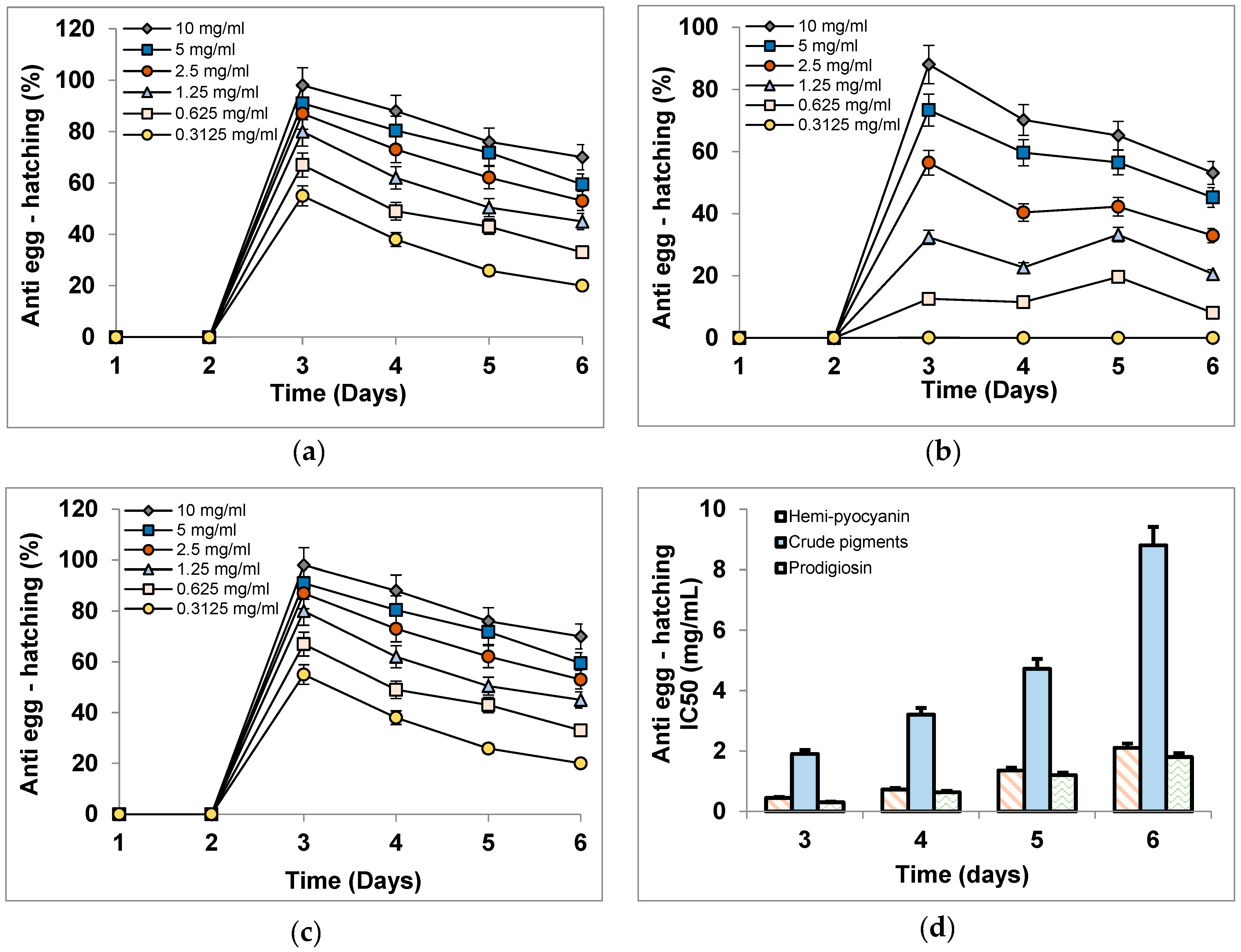
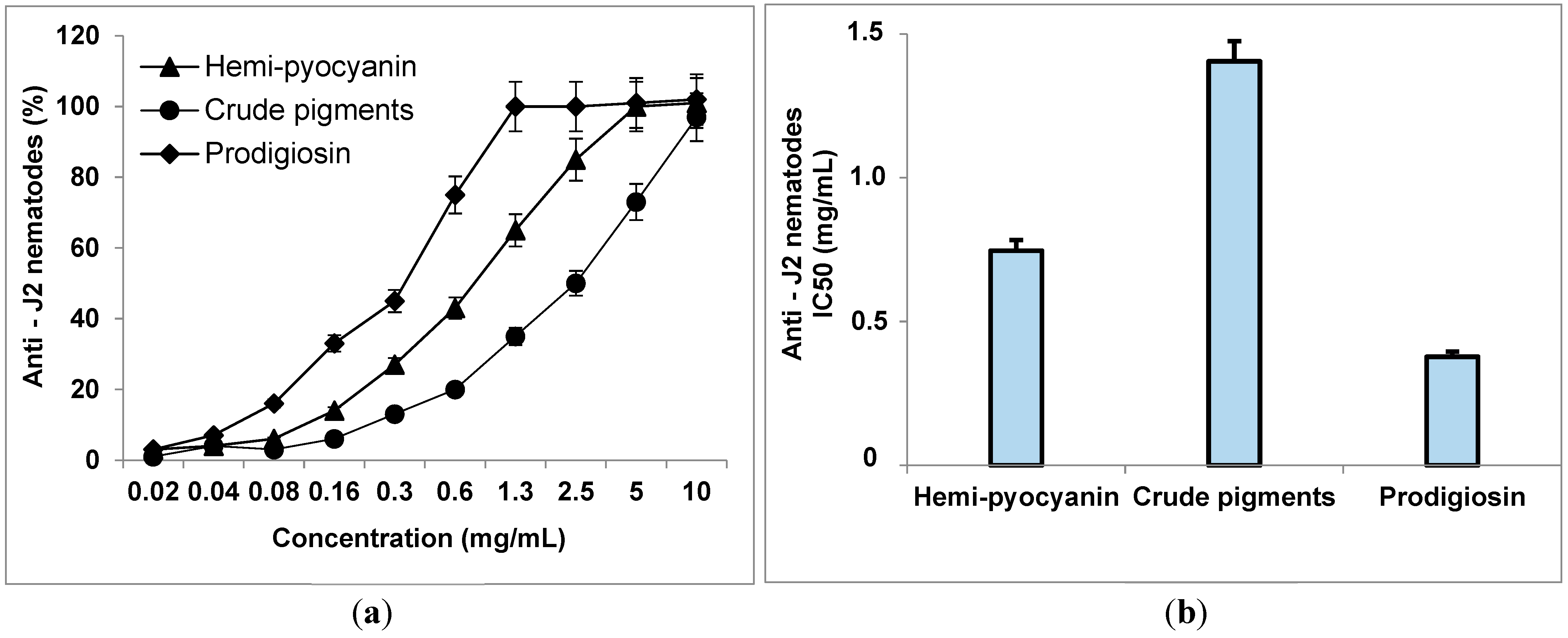
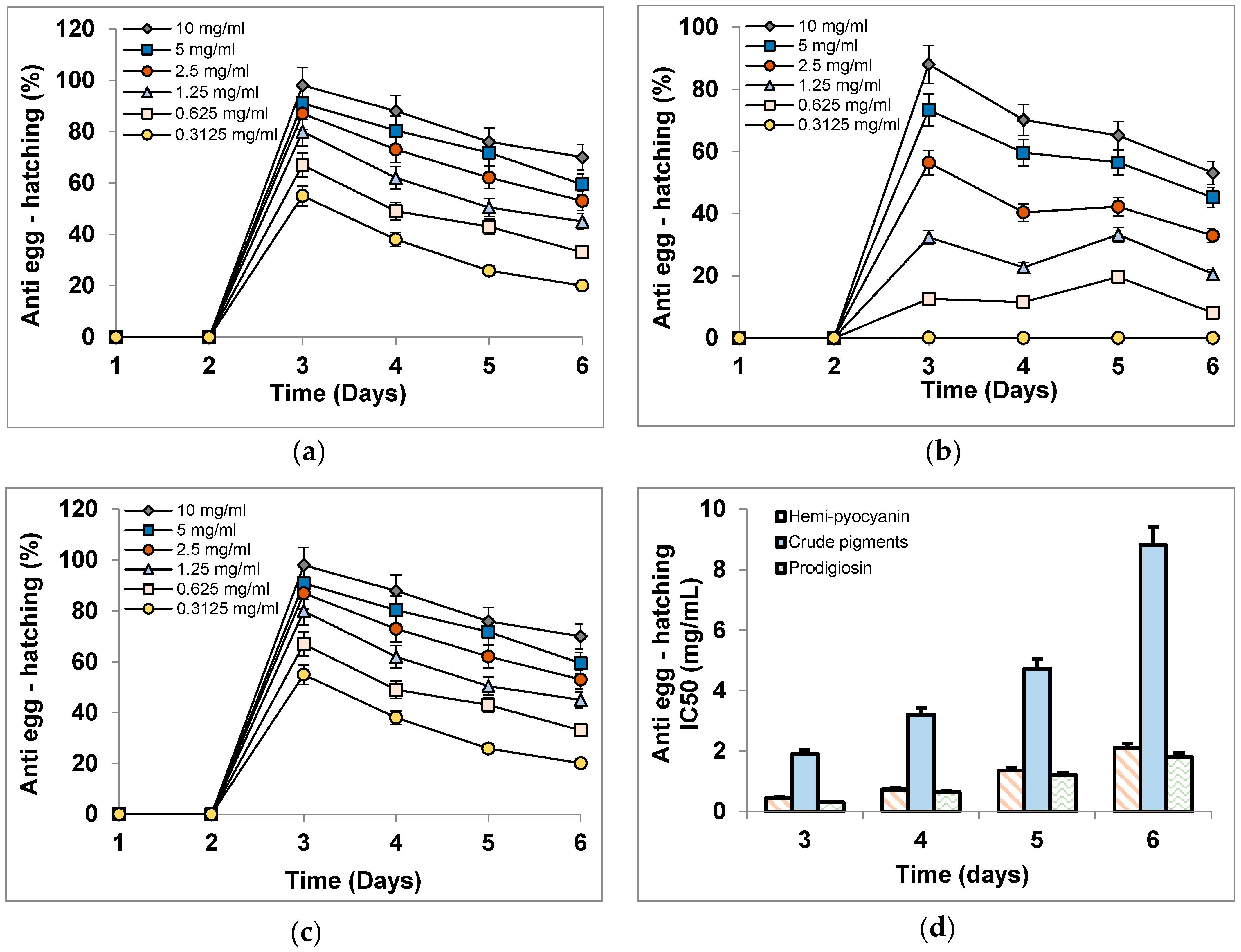
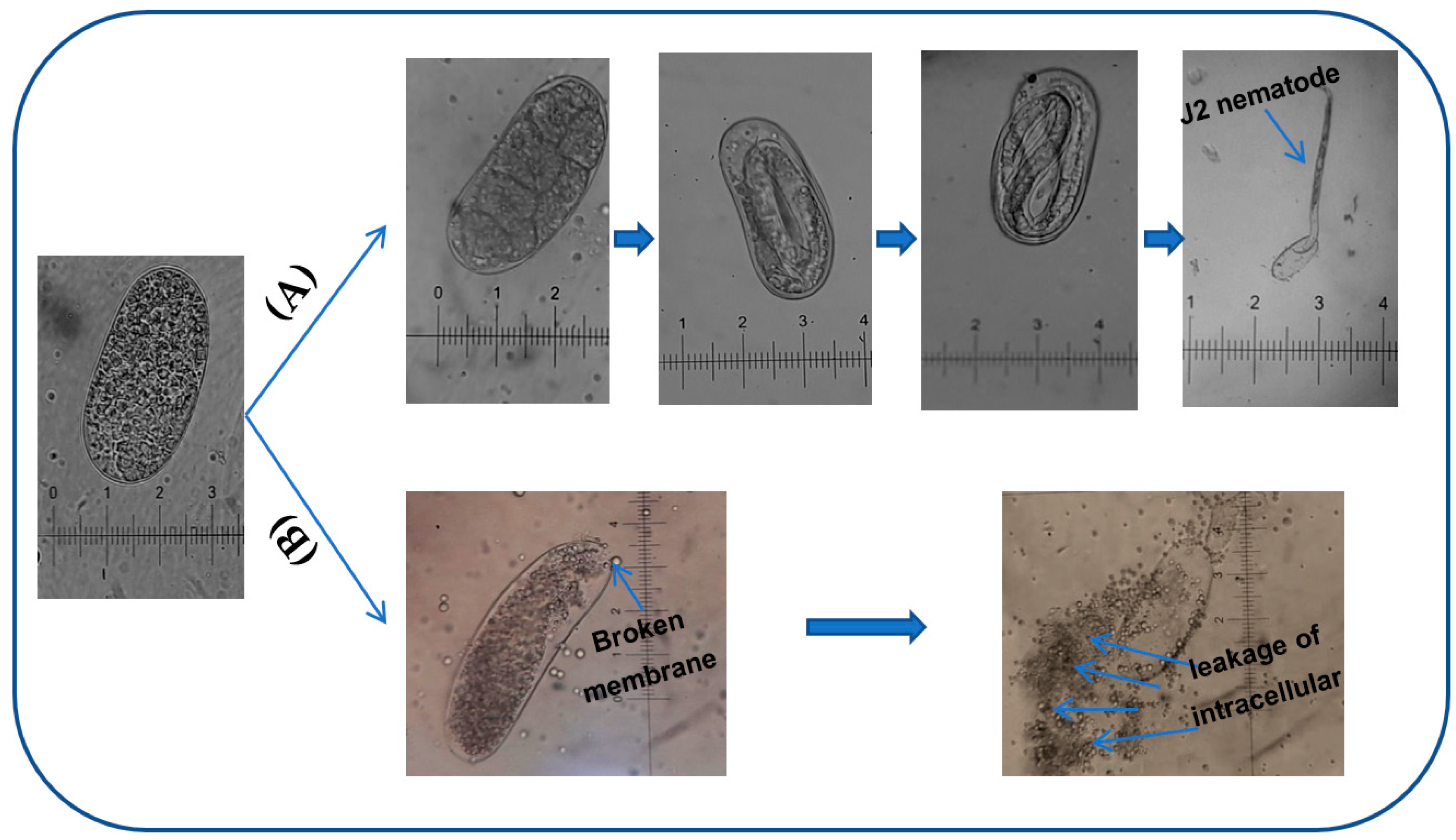
3.6. The Inhibitory Activity of Phenazines on Black Pepper Nematodes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Santos, V.P.; Marques, N.S.S.; Maia, P.C.S.V.; Lima, M.A.B.; Franco, L.O.; Campos-Takaki, G.M. Seafood waste as attractive source of chitin and chitosan production and their applications. Int. J. Mol. Sci. 2020, 21, 4290. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery wastes as a yet undiscovered treasure from the sea: Biomolecules sources, extraction methods and valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Doan, C.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Reclamation of fishery processing waste: A mini-review. Molecules 2019, 24, 2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradit, S.; Noppradit, P.; Goh, B.P.; Sornplang, K.; Ong, M.C.; Towatana, P. Occurrence of microplastics and trace metals in fishand shrimp from Songkhla Lake, Thailand during the COVID-19 pandemic. Appl. Ecol. Environ. Res. 2021, 19, 1085–1106. [Google Scholar] [CrossRef]
- Hue, H.T.T.; Pradit, S.; Lim, A.; Goncalo, C.; Nitiratsuwan, T. Shrimp and fish catch landing trends in Songkhla Lagoon, Thailandduring 2003–2016. Appl. Ecol. Environ. Res. 2018, 16, 3061–3078. [Google Scholar] [CrossRef]
- Venugopal, V. Valorization of seafood processing discards: Bioconversion and biorefinery approaches. Front. Sustain. Food Syst. 2021, 5, 611835. [Google Scholar] [CrossRef]
- Sanuddin, M.; Yulianis, Y.; Annisaq, N. Synthesis chitosan from squid pens waste. ALKIMIA J. Ilmu Kim. Dan Terap. 2020, 4, 6–11. [Google Scholar]
- Huang, Y.L.; Tsai, Y.H. Extraction of chitosan from squid pen waste by high hydrostatic pressure: Effects on physicochemical properties and antioxidant activities of chitosan. Int. J. Biol. Macromol. 2020, 160, 677–687. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L.; Nguyen, A.D.; Phan, T.Q.; Techato, K.; Pradit, S. Bioproduction of prodigiosin from fishery processingwaste shrimp heads and evaluation of its potential bioactivities. Fishes 2021, 6, 30. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wang, S.L.; Nguyen, D.N.; Nguyen, A.D.; Nguyen, T.H.; Doan, M.D.; Ngo, V.A.; Doan, C.T.; Kuo, Y.H.; Nguyen, V.B. Bioprocessing of marine chitinous wastes for the production of bioactive prodigiosin. Molecules 2021, 26, 3138. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, D.N.; Nguyen, A.D.; Ngo, V.A.; Ton, T.Q.; Doan, C.T.; Pham, T.P.; Tran, T.P.H.; Wang, S.L. Utilization of crab waste for cost-effective bioproduction of prodigiosin. Mar. Drugs 2020, 18, 523. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, D.N.; Wang, S.L. Microbial reclamation of chitin and protein-containing marine by-products for the production of prodigiosin and the evaluation of its bioactivities. Polymers 2020, 12, 1328. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Chen, S.P.; Nguyen, T.H.; Nguyen, M.T.; Tran, T.T.T.; Doan, C.T.; Nguyen, A.D.; Kuo, Y.H.; Wang, S.L. Novel efficient bioprocessing of marine chitins into active anticancer prodigiosin. Mar. Drugs 2019, 18, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, L.T.; Techato, K.; Nguyen, V.B.; Wang, S.L.; Nguyen, A.D.; Phan, T.Q.; Doan, M.D.; Phoungthong, K. Utilization of cassava wastewater for low-cost production of prodigiosin via Serratia marcescens TNU01 fermentation and its novel potent α-glucosidase inhibitory effect. Molecules 2021, 26, 6270. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Wang, S.L.; Doan, M.D.; Nguyen, T.H.; Tran, T.H.T.; Tran, T.N.; Doan, C.T.; Ngo, V.A.; Ho, N.D.; Do, V.C.; et al. Utilization of by-product of groundnut oil processing for production of prodigiosin by microbial fermentation and its novel potent anti-nematodes effect. Agronomy 2022, 12, 41. [Google Scholar] [CrossRef]
- Nikolaus, G.; Wulf, B.; Rolf, B. Recent developments in the isolation, biological function, biosynthesis, and synthesis of phenazine natural products. Bioorg. Med. Chem. 2017, 25, 6149–6166. [Google Scholar]
- Alka, R.; Wamik, A. An overview on biosynthesis and applications of extracellular pyocyanin pigment and its role in Pseudomonas aeruginosa pathogenesis. Ann. Phytomed. 2019, 8, 28–42. [Google Scholar]
- Pierson, L.S.; Pierson, E.A. Metabolism and function of phenazines in bacteria: Impacts on the behaviour of bacteria in the environment and biotechnological process. Appl. Microbiol. Biotechnolol. 2010, 86, 1659–1670. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Ye, F.C.; Pang, C.P.; Yong, T.Q.; Tang, W.D.; Xiao, J.; Shang, C.H.; Lu, Z.J. Isolation and identifcation of bioactive substance 1-hydroxyphenazine from Pseudomonas aeruginosa and its antimicrobial activity. Lett. Appl. Microbiol. 2020, 71, 303–310. [Google Scholar] [CrossRef]
- Gruber, J.D.; Wei, C.; Stuart, P.; Kevin, B.; Peter, M.; Patrick, A.F.; Zhang, Y.M. The role of 2,4-dihydroxyquinoline (DHQ) in Pseudomonas aeruginosa pathogenicity. PeerJ 2016, 4, e1495. [Google Scholar] [CrossRef] [Green Version]
- Barakat, K.M.; Mattar, M.Z.; Sabae, S.Z.; Darwesh, O.M.; Hassan, S.H. Production and characterization of bioactive pyocyanin pigment by marine Pseudomonas aeruginosa Osh1. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 933–943. [Google Scholar]
- Devnath, P.; Uddin, M.K.; Ahmed, F.; Hossain, M.T.; Manchur, M.A. Extraction, purification and characterization of pyocyanin produced by Pseudomonas aeruginosa and evaluation for its antimicrobial activity. Int. Res. J. Biol. Sci. 2017, 6, 1–7. [Google Scholar]
- Ozdal, M.; Gurkok, S.; Ozdal, O.G.; Kurbanoglu, E.B. Enhancement of pyocyanin production by Pseudomonas aeruginosa via the addition of n-hexane as an oxygen vector. Biocatal. Agric. Biotechnol. 2019, 22, 101365. [Google Scholar] [CrossRef]
- Ozdal, M. A new strategy for the efficient production of pyocyanin, a versatile pigment, in Pseudomonas aeruginosa OG1 via toluene addition. 3 Biotech. 2019, 9, 370. [Google Scholar] [CrossRef] [PubMed]
- El-Fouly, M.Z.; Sharaf, A.M.; Shahin, A.A.M.; El-Bialy, H.A.; Omara, A.M.A. Biosynthesis of pyocyanin pigment by Pseudomonas aeruginosa. J. Radiat. Res. Appl. Sci. 2015, 8, 36–48. [Google Scholar] [CrossRef] [Green Version]
- DeBritto, S.; Gajbar, T.D.; Satapute, P.; Lalitha, S.; Ramachandra, Y.L.; Sudisha, J.; Shin-ichi, I. Isolation and characterization of nutrient dependent pyocyanin from Pseudomonas aeruginosa and its dye and agrochemical properties. Sci. Rep. 2020, 10, 1542. [Google Scholar] [CrossRef] [Green Version]
- Hüseyin, K.; Cennet, C.K. Pyocyanine production, twitching motility and hydrophobicity of different wastes on Pseudomonas aeruginosa. Pol. J. Environ. Stud. 2021, 30, 1–5. [Google Scholar]
- Bianca, T.M.O.; Patrik, S.Z.B.; Thiago, G.C.; Ian, P.G.A.; Ulrich, V. Craft beer waste as substrate for pyocyanin synthesis. IOSR-JPBS 2019, 14, 21–25. [Google Scholar]
- Nguyen, D.N.; Wang, S.L.; Nguyen, A.D.; Doan, M.D.; Tran, D.M.; Nguyen, T.H.; Ngo, V.A.; Doan, C.T.; Tran, T.N.; Do, V.C.; et al. Potential application of rhizobacteria isolated from the central highland of Vietnam as an effective biocontrol agent of robusta coffee nematodes and as a bio-fertilizer. Agronomy 2021, 11, 1887. [Google Scholar] [CrossRef]
- Wang, S.L.; Yeh, P.Y. Production of a surfactant and solvent-stable alkaliphilic protease by bioconversion of shrimp shell wastes fermented by Bacillus subtilis TKU007. Process. Biochem. 2006, 41, 1545–1552. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wang, S.-L.; Nguyen, A.D.; Doan, M.D.; Tran, T.N.; Doan, C.T.; Nguyen, V.B. Novel α-Amylase inhibitor hemi-pyocyanin produced by microbial conversion of chitinous discards. Mar. Drugs 2022, 20, 283. [Google Scholar] [CrossRef]
- Elbargisy, R.M. Optimization of nutritional and environmental conditions for pyocyanin production by urine isolates of Pseudomonas aeruginosa. Saudi J. Biol. Sci. 2021, 28, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Francisco, J.B.V.; Jesús, A.P.G.; Mayra, L.F.M.; Fabricio, E.A.; Luis, A.O.F.; Yolanda, R.V. Optimized production of a redox metabolite (pyocyanin) by Pseudomonas aeruginosa NEJ01R using a maize by-product. Microorganisms 2020, 8, 1559. [Google Scholar]
- Khan, Z.; Kim, S.G.; Jeon, Y.H.; Khan, H.U.; Son, S.H.; Kim, Y.H. A plant growth promoting rhizobacterium, Paenibacillus polymyxastrain GBR-1, suppresses root-knot nematode. Bioresour. Technol. 2008, 99, 3016–3302. [Google Scholar] [CrossRef] [PubMed]
- Onbasli, D.; Aslim, A. Determination of antimicrobial activity and production of some metabolites by Pseudomonas aeruginosa B1 and B2 in sugar beet molasses. Afr. J. Biotechnol. 2008, 7, 4614–4619. [Google Scholar]
- Onbasli, D.; Aslim, B.; Yuvalicelik, G. Investigation of metabolite productions and degradation of hazardous organic pollutants by Pseudomonas spp. J. Appl. Biol 2011, 5, 9–14. [Google Scholar]
- Li, S.; Mou, Q.; Feng, N.; Leung, P.H.M. A selective medium for pyocyanin-dependent fast electrochemical detection of Pseudomonas aeruginosa in environmental microbial samples. Int. J. Electrochem. Sci. 2018, 13, 3789–3798. [Google Scholar] [CrossRef]
- Hicham, D.; Rachid, D.; Saïd, N. Antimicrobial, antioxidant and hemolytic effects of Pyocyanin produced by Pseudomonas aeruginosa isolated from saline soil of Mina river, Algeria. Int. J. Biosci. 2016, 9, 134–143. [Google Scholar]
- Burton, M.O.; Eagles, B.A.; Campbell, J.R. The amino acid requirements for pyocyanin production. Can. J. Res. 1947, 25, 121–128. [Google Scholar] [CrossRef]
- Georgia, F.R.; Poe, C.F. Study of bacterial fluorescence in various media. The production of fluorescence in media made from peptones. J. Bacteriol. Res. 1932, 23, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Yupeng, W.; Hongchen, L.; Mo, X.; Wei, H. Biosynthesis and metabolic engineering of 1-hydroxyphenazine in Pseudomonas chlororaphis H18. Microb. Cell Factories 2021, 20, 235. [Google Scholar]
- Ling, H.; Xunzhi, Z.; Shixing, Z.; Zhenrui, C.; Kai, S.; Chi, Z.; Hua, S. Phthalic acid esters: Natural sources and biological. Toxins 2021, 13, 495. [Google Scholar]
- Wiratno, M.S.; Ankardiansyah, P.P.; Ahmed, I.A.Y. Biological control of root-knot nematode (Meloidogyne spp.) in pepper plants utilizing endophytic bacteria Pseudomonas sp. and Micrococcus sp. J. Pepper Ind. 2018, 9, 11–22. [Google Scholar]
- Thuy, T.T.T.; Chi, N.T.M.; Yen, N.T.; Anh, L.T.N.; Te, L.L.; De Waele, D. Fungi associated with black pepper plants in Quang Tri province (Vietnam), and interaction between Meloidogyne incognita and Fusarium solani. Int. J. Sci. Technol. Res. 2013, 46, 470–482. [Google Scholar]
- Singh, H.B.; Keswani, C.; Reddy, M.S.; Sansinenea, E.; Carlos, G.E. Biological control of nematodes by plant growth promoting rhizobacteria: Secondary metabolites involved and potential applications. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Harikesh, B.S., Chetan, K., Reddy, M.S., Estibaliz, S., Carlos, G.E., Eds.; Springer: Singapore, 2019; Chapter 13; pp. 253–264. [Google Scholar]
- Javed, N.; Gowen, S.R.; Inam, M.; Abdullah, K.; Shahina, F. Systemic and persistent effect of neem (Azadirachta indica) formulations against root-knot nematodes, Meloidogyne javanica and their storage life. Crop. Prot. 2007, 26, 911–916. [Google Scholar] [CrossRef]
- Bogner, C.W.; Kamdem, R.S.; Sichtermann, G.; Matthäus, C.; Hölscher, D.; Popp, J.; Proksch, P.; Grundler, F.M.; Schouten, A. Bioactive secondary metabolites with multiple activities from a fungal endophyte. Microb. Biotechnol. 2017, 10, 175–188. [Google Scholar] [CrossRef]
- Zhao, D.; Zhu, X.; Chen, L.; Liu, W.; Chen, J.; Wang, S.; Zang, J.; Duan, Y.; Liu, X. Toxicity of a secondary metabolite produced by Simplicillium chinense Snef5 against the root-knot nematode Meloidogyne incognita. Acta. Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 550–555. [Google Scholar] [CrossRef]
- Puja, O.; Satinder, K.P. Effect of phenolic compounds on nematodes—A review. J. Nat. Appl. Sci. 2010, 2, 344–350. [Google Scholar]
- Sabrina, C.; Carolyn, A.B.; Ulrike, M. Functions of flavonoids in plant–nematode interactions. Plants 2018, 7, 85. [Google Scholar]
- Nguyen, T.L.T.; Jang, J.Y.; Kim, T.Y.; Yu, N.H.; Park, A.R.; Lee, S.; Bae, C.H.; Yeo, J.H.; Hur, J.S.; Park, H.W.; et al. Nematicidal activity of verrucarin A and roridin A isolated from Myrothecium verrucaria against Meloidogyne incognita. Pestic. Biochem. Phys. 2018, 148, 133–143. [Google Scholar] [CrossRef]
- Kim, T.Y.; Jang, J.Y.; Jeon, S.J.; Lee, H.W.; Bae, C.H.; Yeo, J.H.; Lee, H.B.; Kim, I.S.; Park, H.W.; Kim, J.C. Nematicidal activity of kojic acid produced by Aspergillus oryzae against Meloidogyne incognita. J. Microbiol. Biotechnol. 2016, 26, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Linsong, W.; Yukun, Q.; Zhaoqian, F.; Kun, G.; Jiang, Z.; Ronge, X.; Song, L.; Pengcheng, L. Novel lead compound discovery from Aspergillus fumigatus 1T-2 against Meloidogyne incognita based on a chemical ecology study. J. Agric. Food Chem. 2022, 70, 4644–4657. [Google Scholar]
- Zeng, Q.; Huang, H.; Zhu, J.; Fang, Z.; Sun, Q.; Bao, S. A new nematicidal compound produced by Streptomyces albogriseolus HA10002. Antonie Leeuwenhoek 2013, 103, 1107–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brent, C.; Nawaporn, V.; Daniel, G.L.; Grace, J.Y.; Alan, S.; Frederick, M.A. Identification of Pseudomonas aeruginosa phenazines that kill Caenorhabditis elegans. PLoS Pathog. 2014, 9, e1003101. [Google Scholar]
- Zahraa, J.J.; Anaam, F.H.; Muthana, A.A.; Nuha, F.A.; Eman, S.A. Bioactivity of pyocyanin of Pseudomonas aeruginosa clinical isolates against a variety of human pathogenic bacteria and fungi species. Int. J. Antimicrob. Agents 2017, 7, 1–7. [Google Scholar]
- Sunish, K.R.; Ayyadurai, N.; Pandiaraja, P.; Reddy, A.V.; Venkateswarlu, Y.; Prakash, O.; Sakthivel, N. Characterization of anti-fungal metabolite produced by a new strain Pseudomonas aeruginosa PUPa3 that exhibits broad spectrum antifungal activity and biofertilizing traits. J. Appl. Microbiol. 2004, 98, 145–154. [Google Scholar]
- Simionato, A.S.; Navarro, M.O.; Jesus, M.; Barazetti, A.R.; Silva, C.S.; Simões, G.C.; Balbi-Peña, M.I.; Mello, J.C.; Panagio, L.A.; Almeida, R.S.; et al. The Effect of Phenazine-1-carboxylic acid on mycelial growth of Botrytis cinerea produced by Pseudomonas aeruginosa LV strain. Front. Microbiol. 2017, 8, 1102. [Google Scholar] [CrossRef]
- Makarand, R.R.; Prashant, D.S.; Bhushan, L.C.; Sudhir, B.C. Detection, isolation and identification of phenazine-1-carboxylic acid produced by biocontrol strains of Pseudomonas aeruginosa. J. Sci. Ind. Res. 2007, 66, 627–631. [Google Scholar]
- Jasim, B.; Anisha, C.; Rohini, S.; Kurian, J.M.; Jyothis, M.; Radhakrishnan, E.K. Phenazine carboxylic acid production and rhizome protective effect of endophytic Pseudomonas aeruginosa isolated from Zingiber officinale. World J. Microbiol. Biotechnol. 2014, 30, 1649–1654. [Google Scholar] [CrossRef]
- Lee, K.W.; Omar, D.; Cheng, G.L.E.; Nasehi, A.; Wong, M.Y. Characterization of phenazine and phenazine-1-carboxylic acid isolated from Pseudomonas aeruginosa UPMP3 and their antifungal activities against Ganoderma boninense. Pertanika J. Trop. Agric. Sci. 2018, 41, 1795–1809. [Google Scholar]
- Aunchalee, N.; Sukanya, A.; Chanokporn, P.; Paweena, P.; Saksit, C.; Chalerm, R. Synthesis, isolation of phenazine derivatives and their antimicrobial activities. Walailak J. Sci. Tech. 2009, 6, 79–91. [Google Scholar]
- Zhang, L.; Tian, X.; Shan, K.; Liu, G.; Zhang, C.; Sun, C. Antagonistic activity and mode of action of Phenazine-1-carboxylic acid, produced by marine bacterium Pseudomonas aeruginosa PA31x, against Vibrio anguillarum in vitro and in a Zebrafish in vivo model. Front. Microbiol. 2017, 8, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaresan, K.; Subramanian, M.; Vaithiyanathan, S.; Sevagaperumal, N.; Gopal, C.; Fernando, W.G.D. Broad spectrum action of phenazine against active and dormant structures of fungal pathogens and root knot nematode. Arch. Phytopathol. Pflanzenschutz 2005, 38, 69–76. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Phenazine Content (PYO, µg/mL) |
|---|---|
| Marine by-products (1) | |
| Squid pen powder (SPP) | 14.52 ± 0.34 |
| Shrimp shell powder (SSP) | - |
| Crab shell powder (CSP) | - |
| Commercial free protein (2) | |
| Yeast extract | - |
| Peptone | 6.91 ± 0.054 |
| Casein | 3.54 ± 0.041 |
| Gelatin | - |
| By-product + peptone (3) | |
| SPP + Peptone | 14.31 ± 0.213 |
| SSP + Peptone | 5.21 ± 0.136 |
| CSP + Peptone | 3.12 ± 0.075 |
| Bacteria Strains | Substrate Sources | Phenazine Content (PYO, µg/mL) | References |
|---|---|---|---|
| Commercial nutrition | |||
| P. aeruginosa OSh1 | King’s B | 25.5 | [21] |
| Peptone water | 11.7 | ||
| Tryptone water | 9.5 | ||
| Nutrient broth | 5.2 | ||
| P. aeruginosa PU10 | Glycerol, D-alanine, peptone | 9.45 | [22] |
| P. aeruginosa | Nutrient broth, n-hexan | 11.96 | [23] |
| Nutrient broth, n-dodecane | 7.65 | ||
| Nutrient broth, n-hexadecane | 7.99 | ||
| P. aeruginosa OG1 | Nutrient broth | 8 | [24] |
| Nutrient broth, 0.2% toluene | 33 | ||
| Nutrient broth, 0.3% chloroform | 20 | ||
| Nutrient broth, 0.2% TriptonX-100 | 13 | ||
| Nutrient broth, 0.1% Tween 80 | 8 | ||
| Nutrient broth, 0.1% Tween 20 | 13 | ||
| P. aeruginosa R1 | King’s A | 4.5 | [25] |
| Glycerol-supplemented mineral medium | 3.2 | ||
| P. aeruginosa P21 | Nutrient broth | 20.7 | [32] |
| Low-cost organic | |||
| P. aeruginosa R1 | Corn steep liquor | ND | [25] |
| Cottonseed meal | 4.0 | ||
| Grape seed | 3.1 | ||
| Peat moss | ND | ||
| Taro leaves | 2.5 | ||
| P. aeruginosa | Corn | 0.34 | [26] |
| Soya bean | 0.17 | ||
| Sweet potato | 1.7 | ||
| Watermelon seeds | 1.3 | ||
| Groundnut | 0.8 | ||
| By-product/waste | |||
| P. aeruginosa TUN03 | Squid pen powder | 14.52 | In this study |
| P. aeruginosa (ATCC 27853) | Waste cheese whey | 1.62 | [27] |
| Waste frying oil | 2 | ||
| Sugar beet molasses | 9.36 | ||
| Turkish coffee waste | ND | ||
| Tea wastewater | 3 | ||
| P. aeruginosa NEJ01R | Maize wastewater | 0.7–3.2 | [33] |
| P. aeruginosa R1 | Olive waste | 1.3 | [25] |
| Pea pods | 1.6 | ||
| Potato-washing water | ND | ||
| Vegetable frying oil | ND | ||
| P. aeruginosa B1 | Sugar beet molasses 5% | 17.1 | [35] |
| Pseudomonas spp. | 1% benzene, 2% gasoline | 0.02–1.30 | [36] |
| P. aeruginosa | Craft beer waste | 21–58 | [28] |
| No | Retention Time (RT) | Peak Areas (%) | Exact Mass (g·mol−1) | Chemical Formula | Identified Scientific Name |
|---|---|---|---|---|---|
| 1 | 13.44 | 11.28 | 196.063663 | C12H8N2O | hemi-pyocyanin |
| 2 | 16.93 | 2.67 | 180.068748 | C12H8N2 | Phenazine |
| 3 | 17.66 | 2.11 | 306.18311 | C18H26O4 | Phthalic acid, monodecyl ester |
| 4 | 19.42 | 71.82 | 362.24571 | C22H34O4 | Phthalic acid, hex-3-yl octyl ester |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.H.; Wang, S.-L.; Nguyen, T.H.; Doan, M.D.; Tran, T.H.T.; Ngo, V.A.; Ho, N.D.; Tran, T.N.; Doan, C.T.; Do, V.C.; et al. Utilization of Fishery-Processing By-Product Squid Pens for Scale-Up Production of Phenazines via Microbial Conversion and Its Novel Potential Antinematode Effect. Fishes 2022, 7, 113. https://doi.org/10.3390/fishes7030113
Nguyen TH, Wang S-L, Nguyen TH, Doan MD, Tran THT, Ngo VA, Ho ND, Tran TN, Doan CT, Do VC, et al. Utilization of Fishery-Processing By-Product Squid Pens for Scale-Up Production of Phenazines via Microbial Conversion and Its Novel Potential Antinematode Effect. Fishes. 2022; 7(3):113. https://doi.org/10.3390/fishes7030113
Chicago/Turabian StyleNguyen, Thi Hanh, San-Lang Wang, Thi Huyen Nguyen, Manh Dung Doan, Thi Ha Trang Tran, Van Anh Ngo, Nhat Duoc Ho, Thi Ngoc Tran, Chien Thang Doan, Van Chung Do, and et al. 2022. "Utilization of Fishery-Processing By-Product Squid Pens for Scale-Up Production of Phenazines via Microbial Conversion and Its Novel Potential Antinematode Effect" Fishes 7, no. 3: 113. https://doi.org/10.3390/fishes7030113
APA StyleNguyen, T. H., Wang, S.-L., Nguyen, T. H., Doan, M. D., Tran, T. H. T., Ngo, V. A., Ho, N. D., Tran, T. N., Doan, C. T., Do, V. C., Nguyen, A. D., & Nguyen, V. B. (2022). Utilization of Fishery-Processing By-Product Squid Pens for Scale-Up Production of Phenazines via Microbial Conversion and Its Novel Potential Antinematode Effect. Fishes, 7(3), 113. https://doi.org/10.3390/fishes7030113