
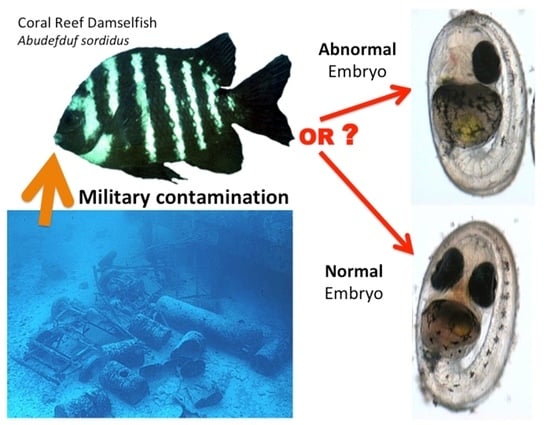
Damselfish Embryos as a Bioindicator for Military Contamination on Coral Reefs at Johnston Atoll, Pacific Ocean


Abstract
:
1. Introduction
1.1. Biomonitoring in Coral Reef Habitats
1.2. PCBs
1.3. Study Objectives
2. Materials and Methods
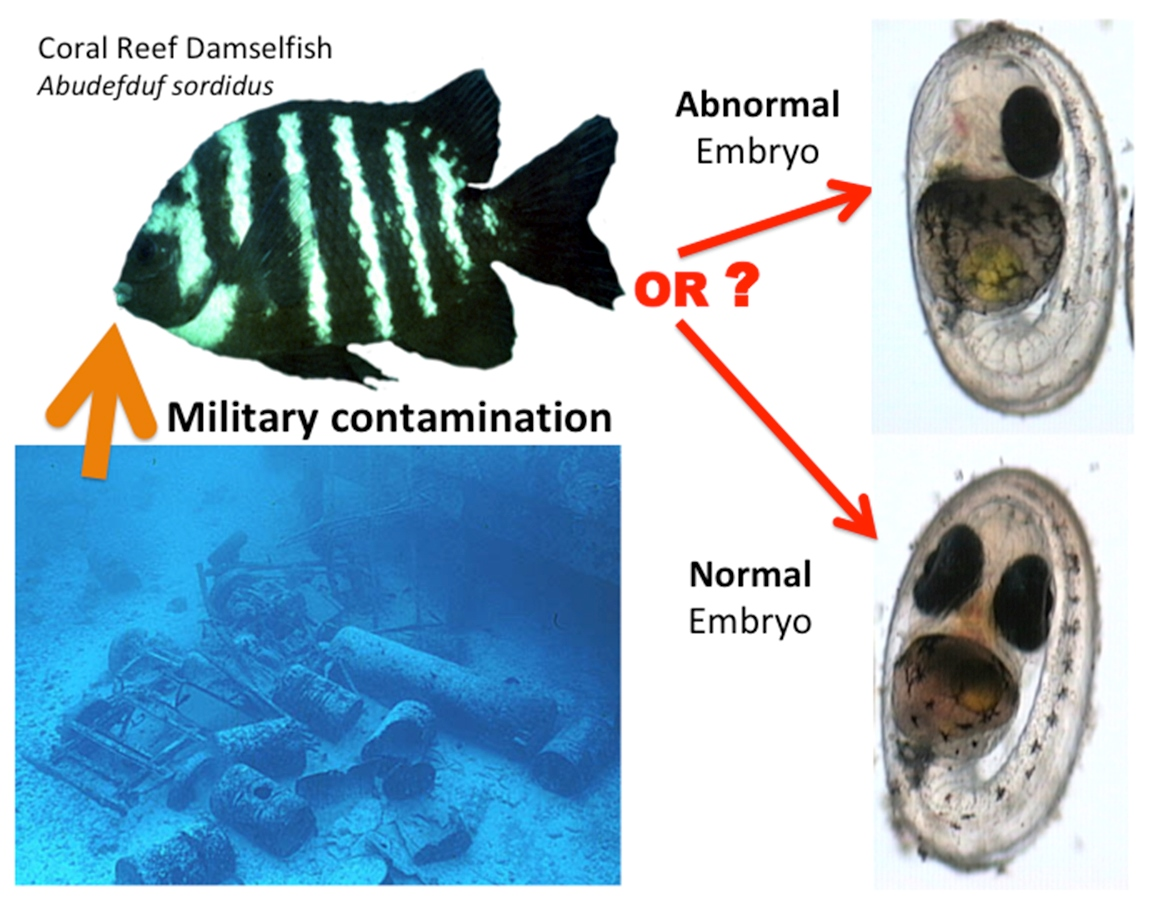
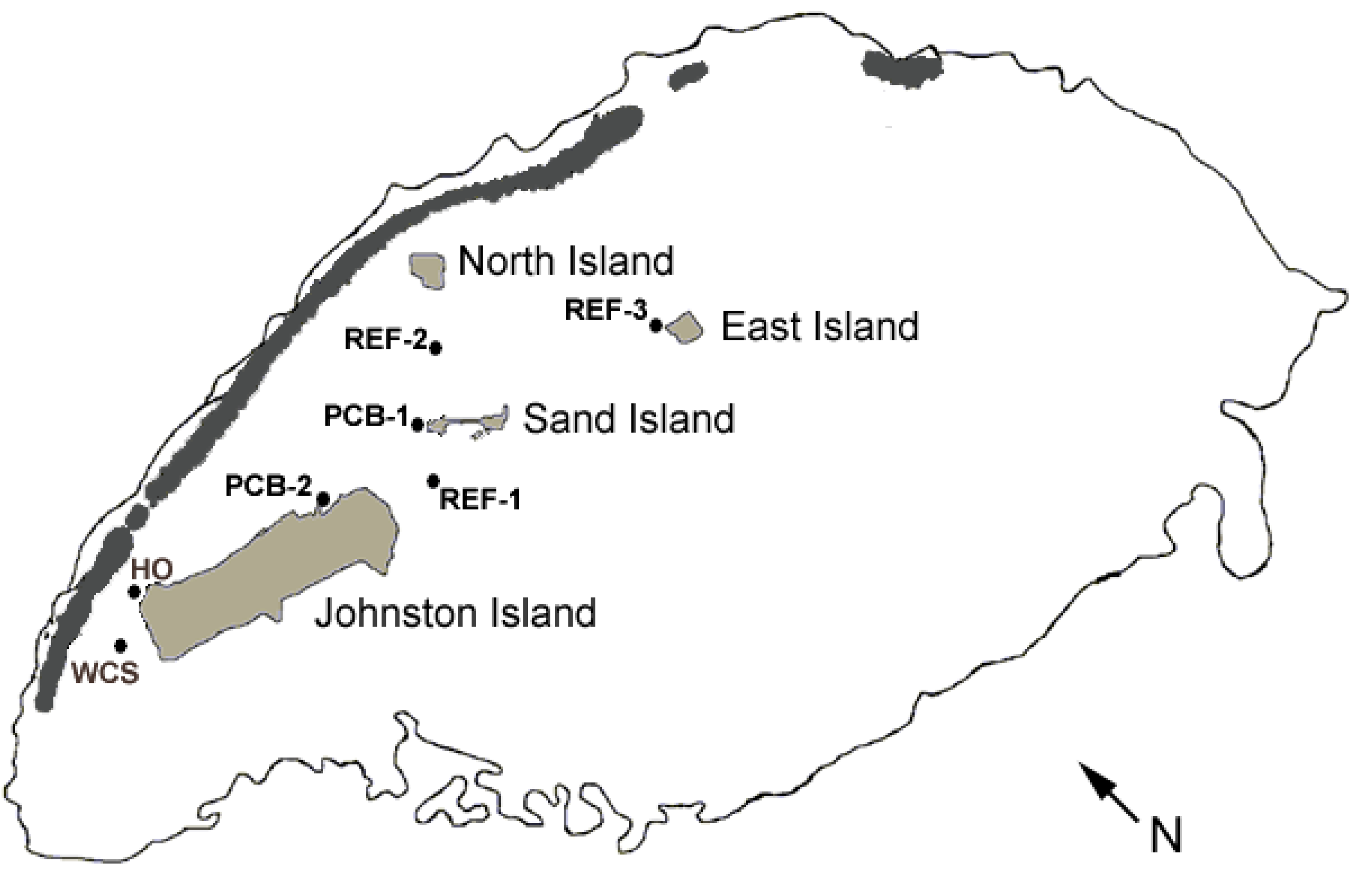
2.1. Study Site and Study Organism
2.2. Data and Sample Collection
2.3. Embryo Incubations
2.4. Chemical Analyses
2.5. Statistical Analyses
3. Results
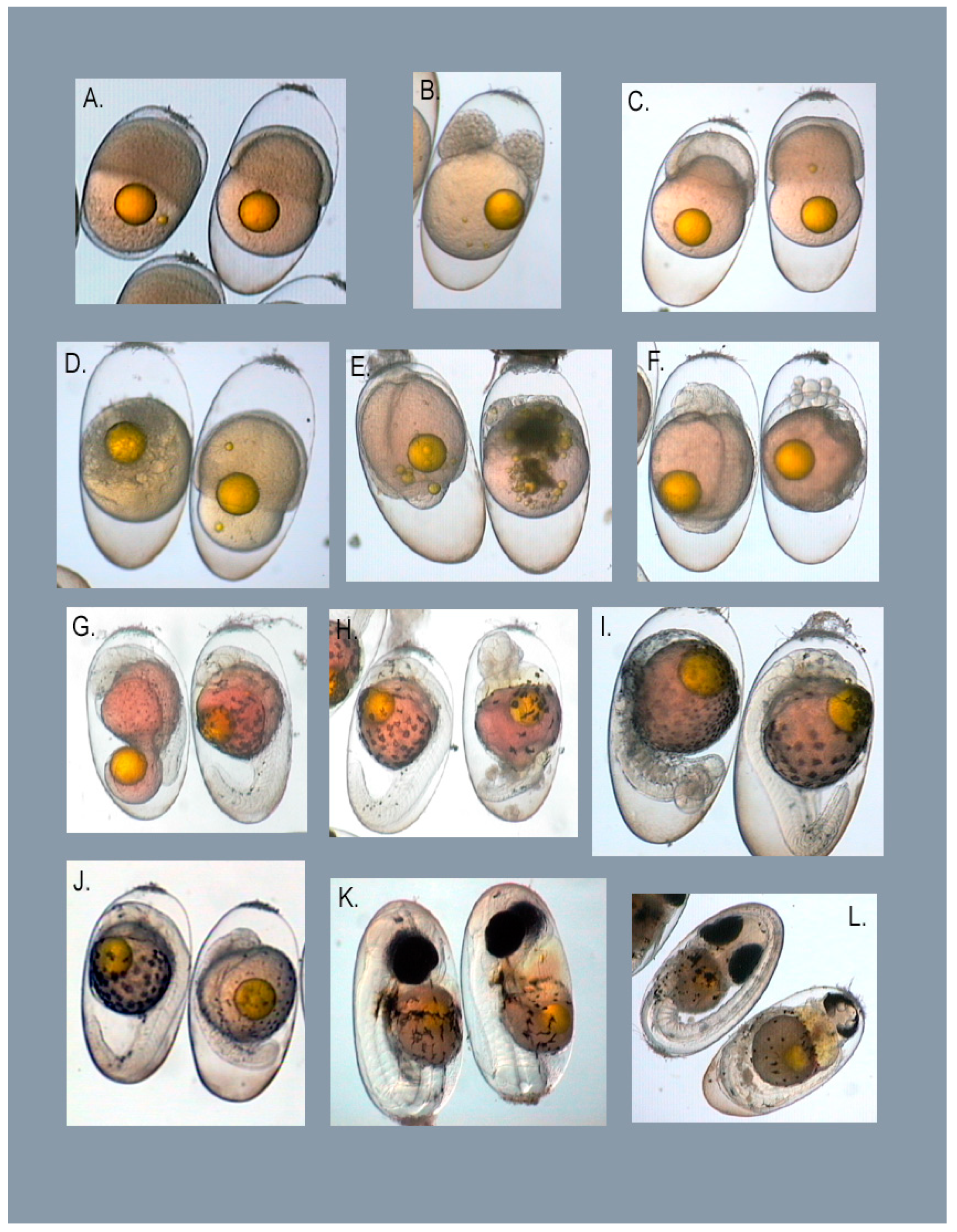
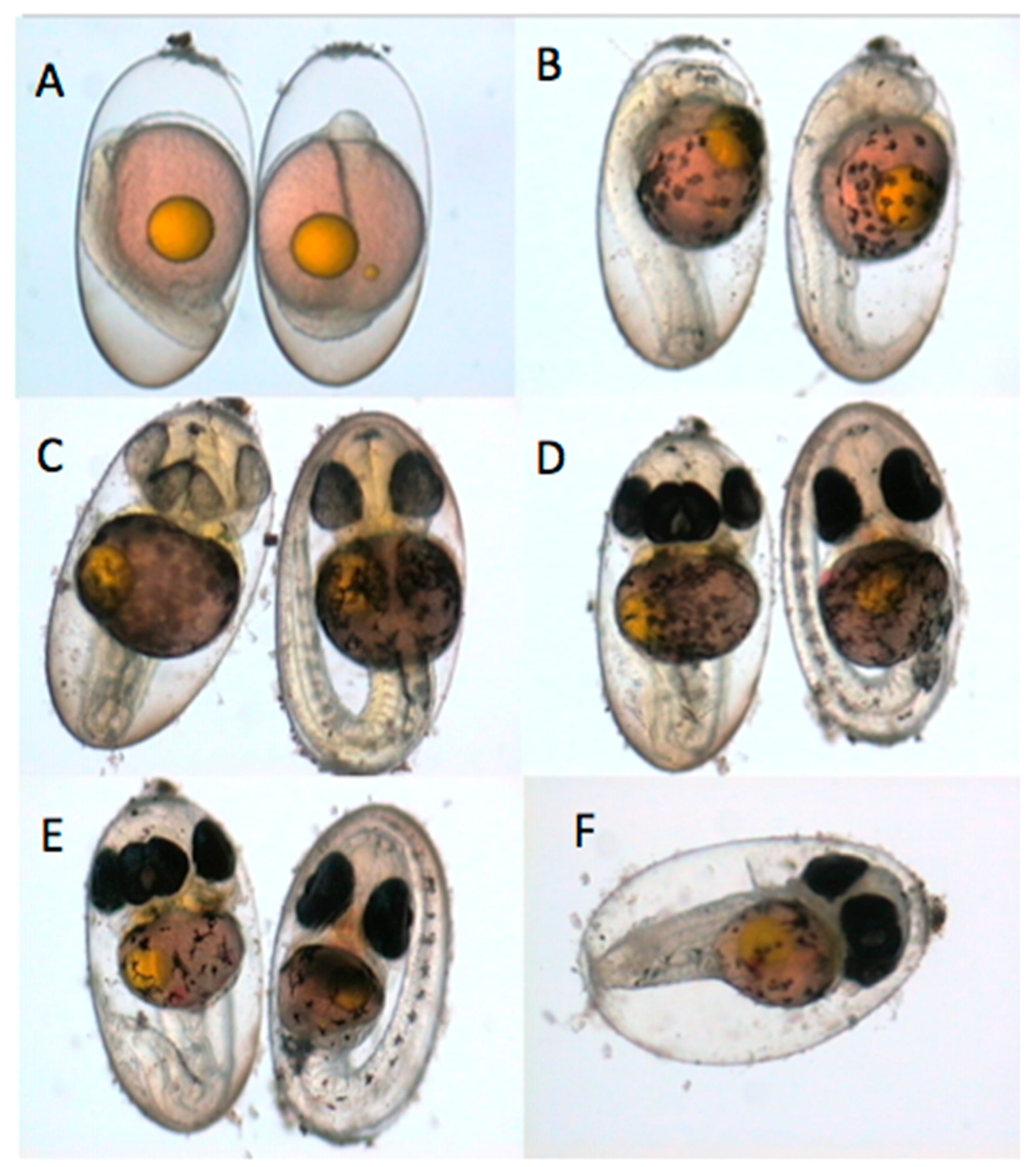
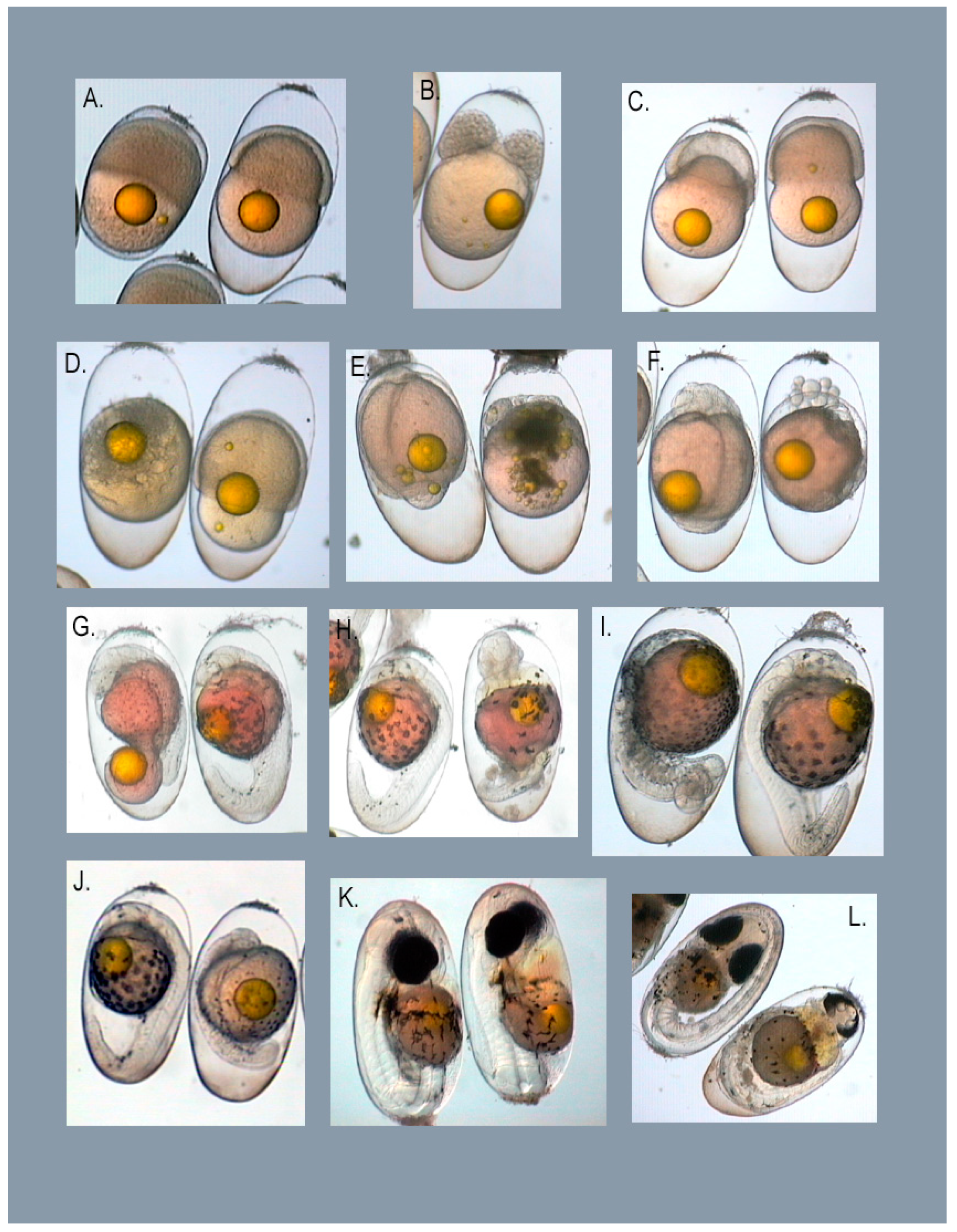
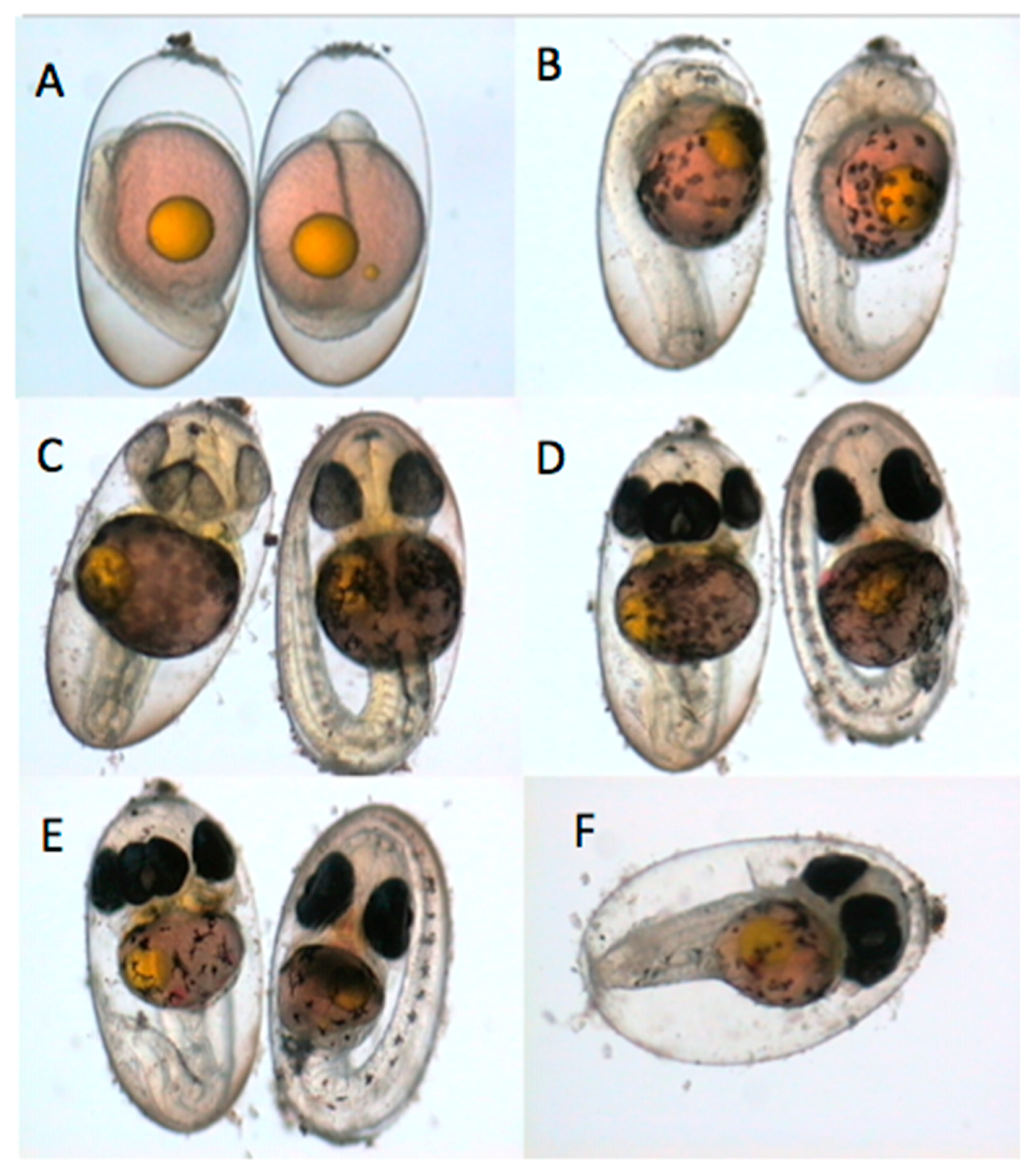
3.1. Types of Embryo Abnormalities Observed
3.2. Embryo Incubation
3.3. The Gonadosomatic and Hepatosomatic Indices (GSI and HSI)
3.4. Gut Contents
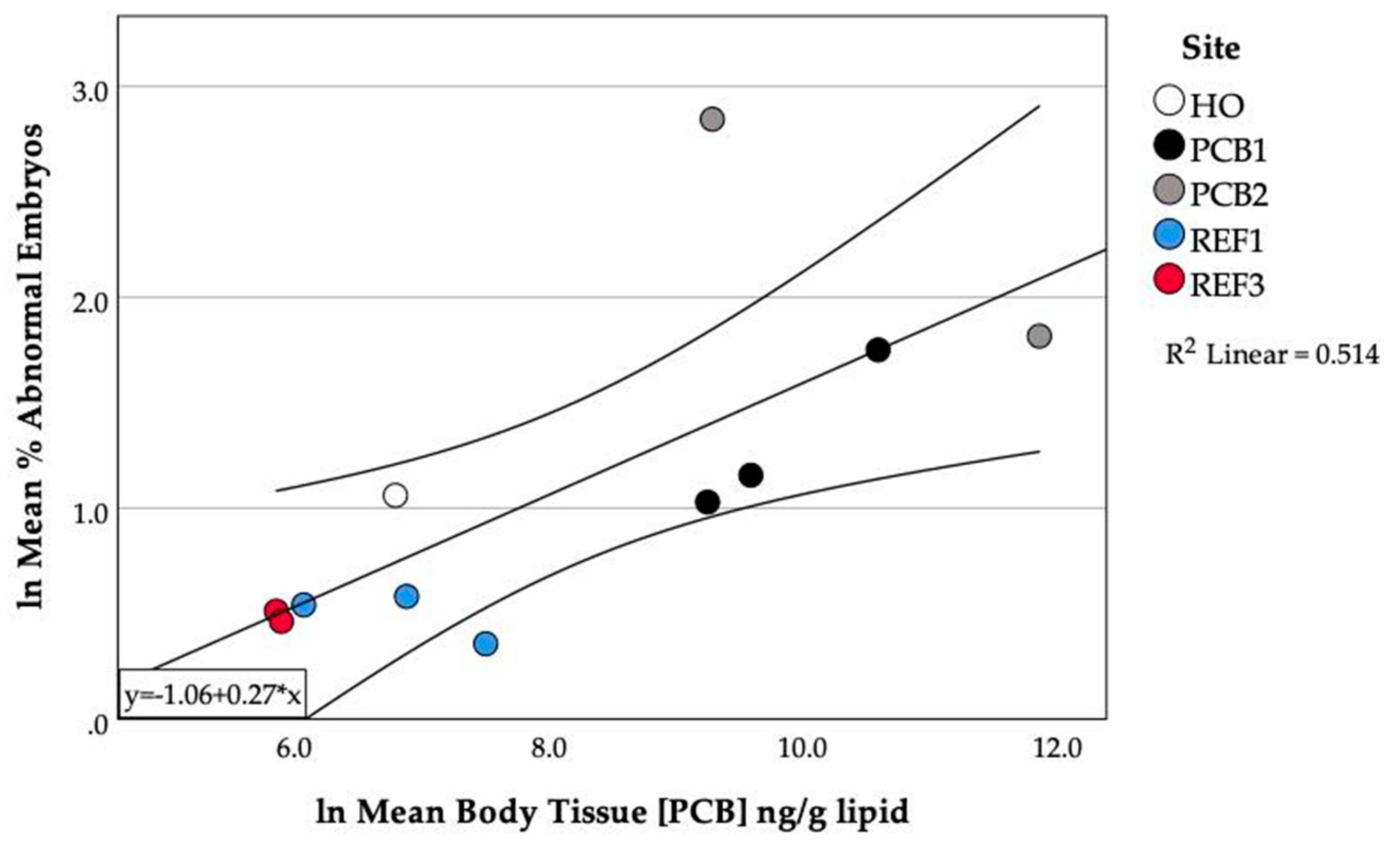
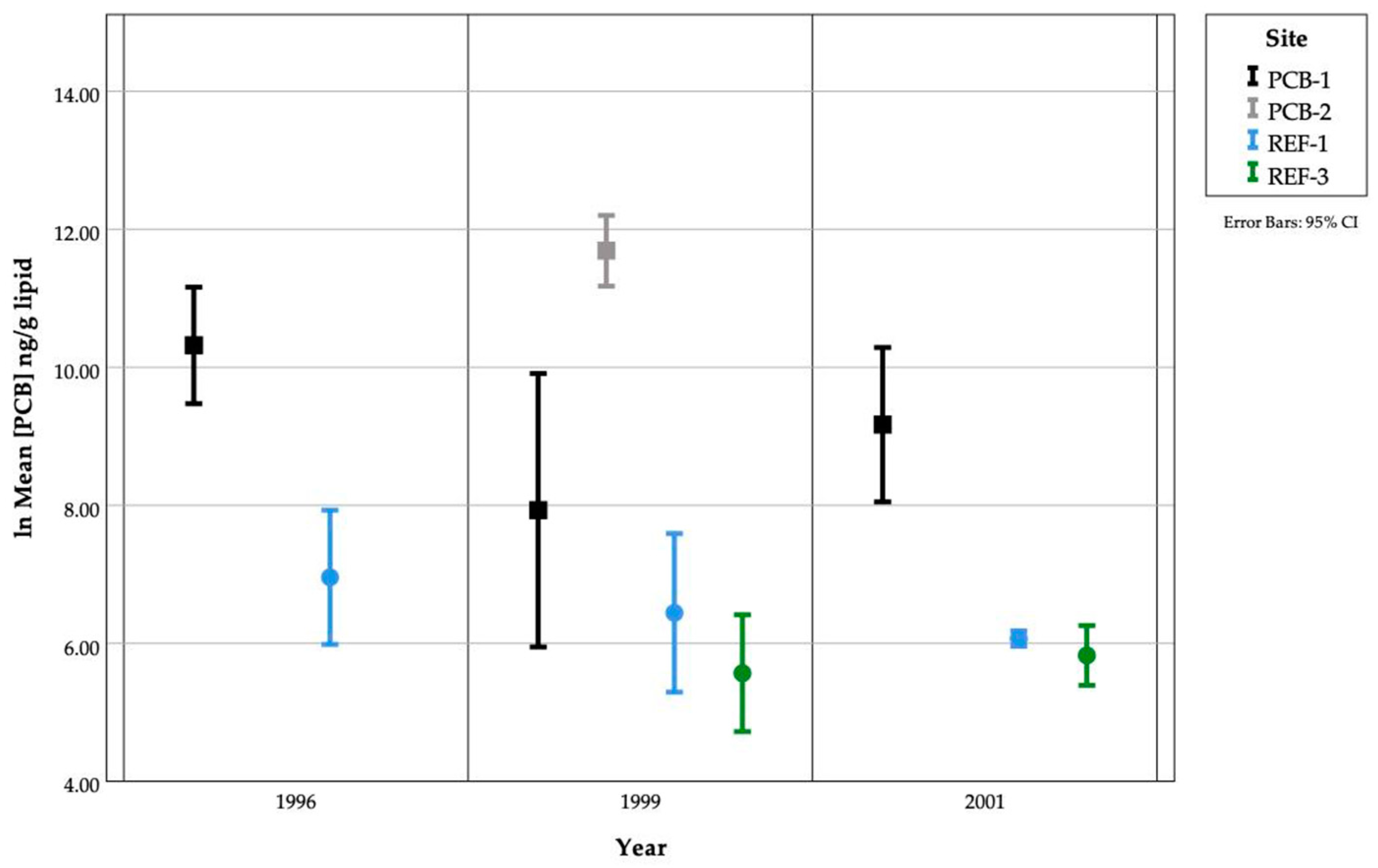
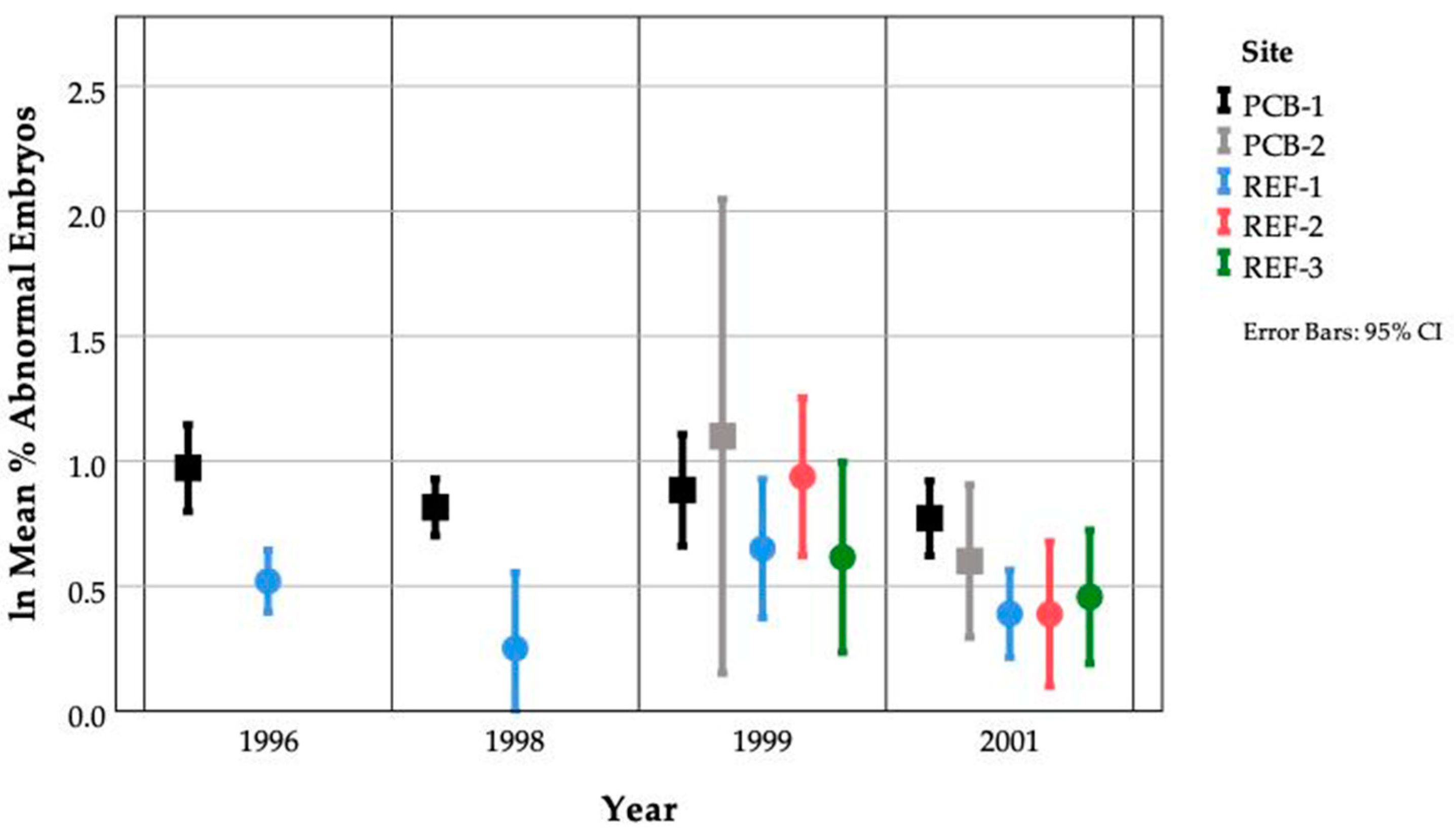
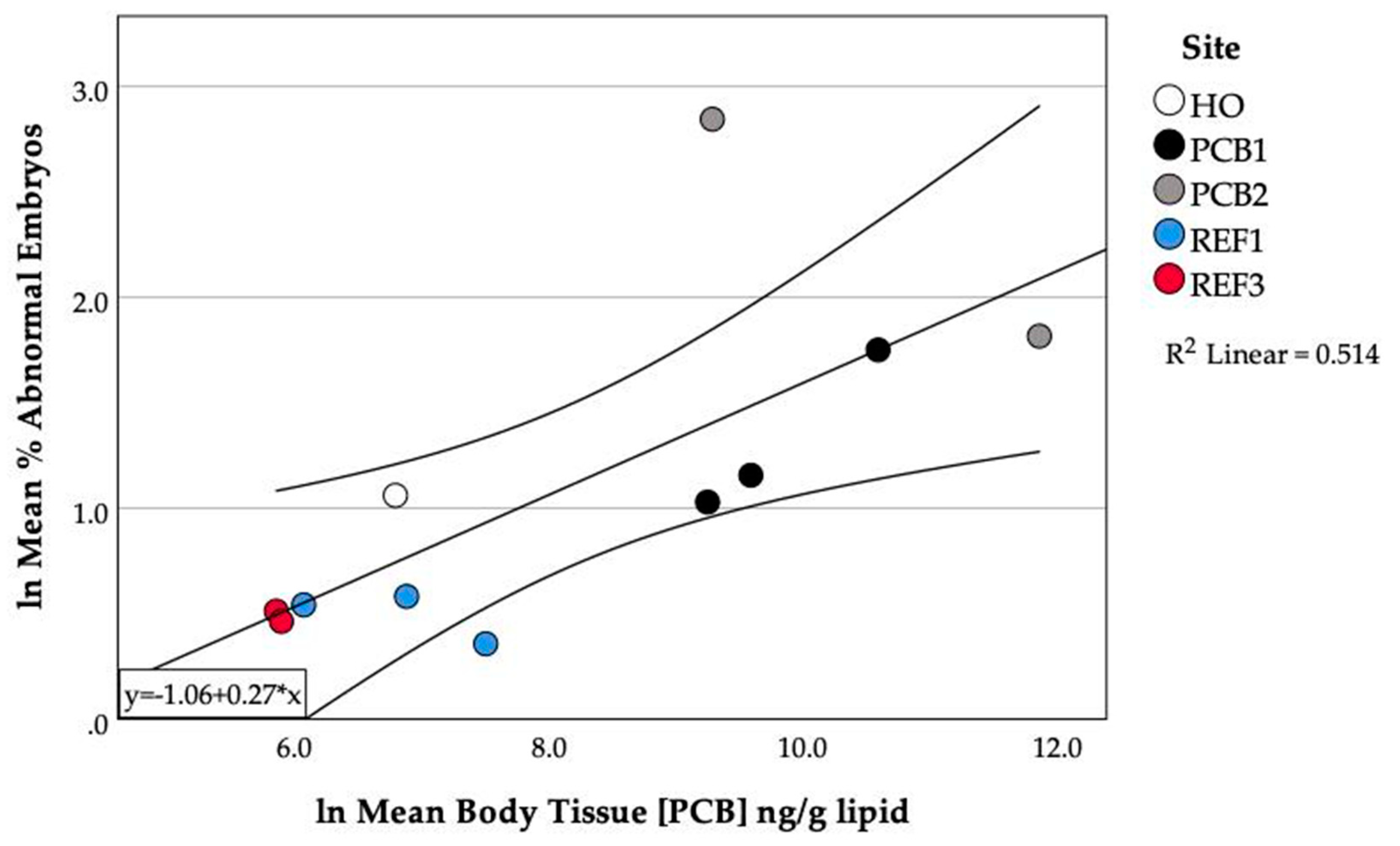
3.5. PCB Accumulation and Embryo Abnormalities
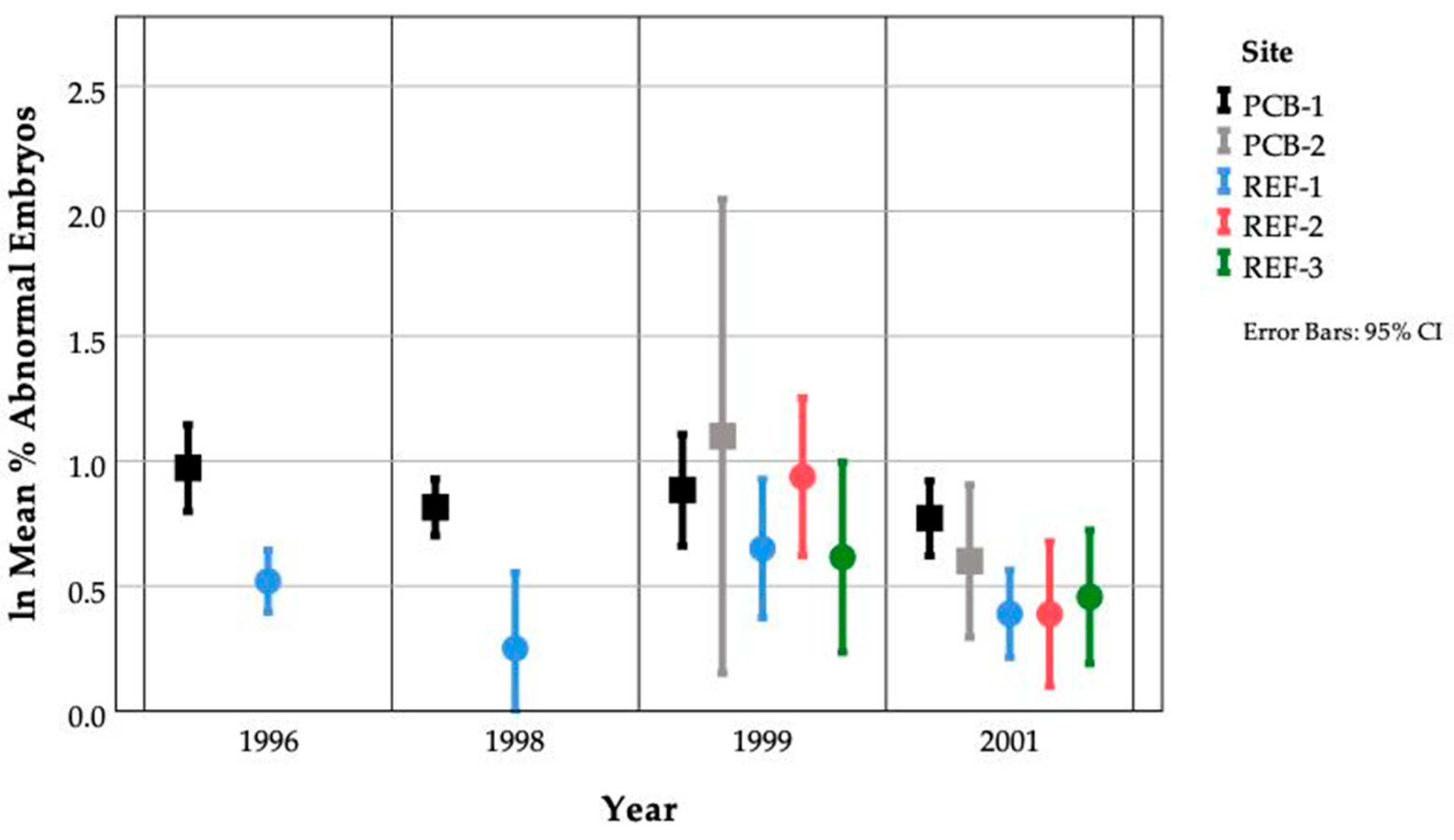
3.6. Within Site Variation
3.7. Between Site Variation
3.8. Relationship between PCBs in Fish and Abnormalities
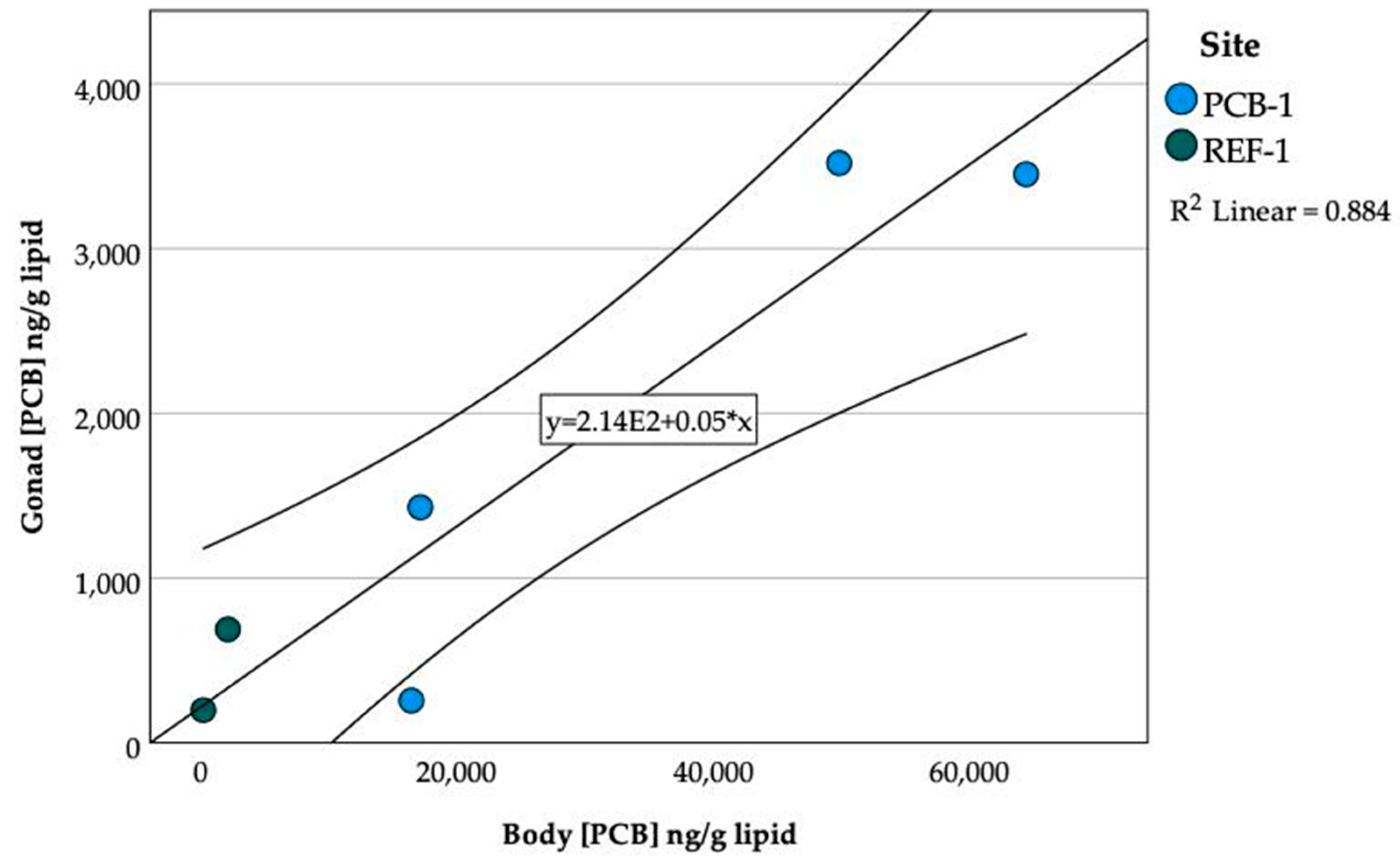
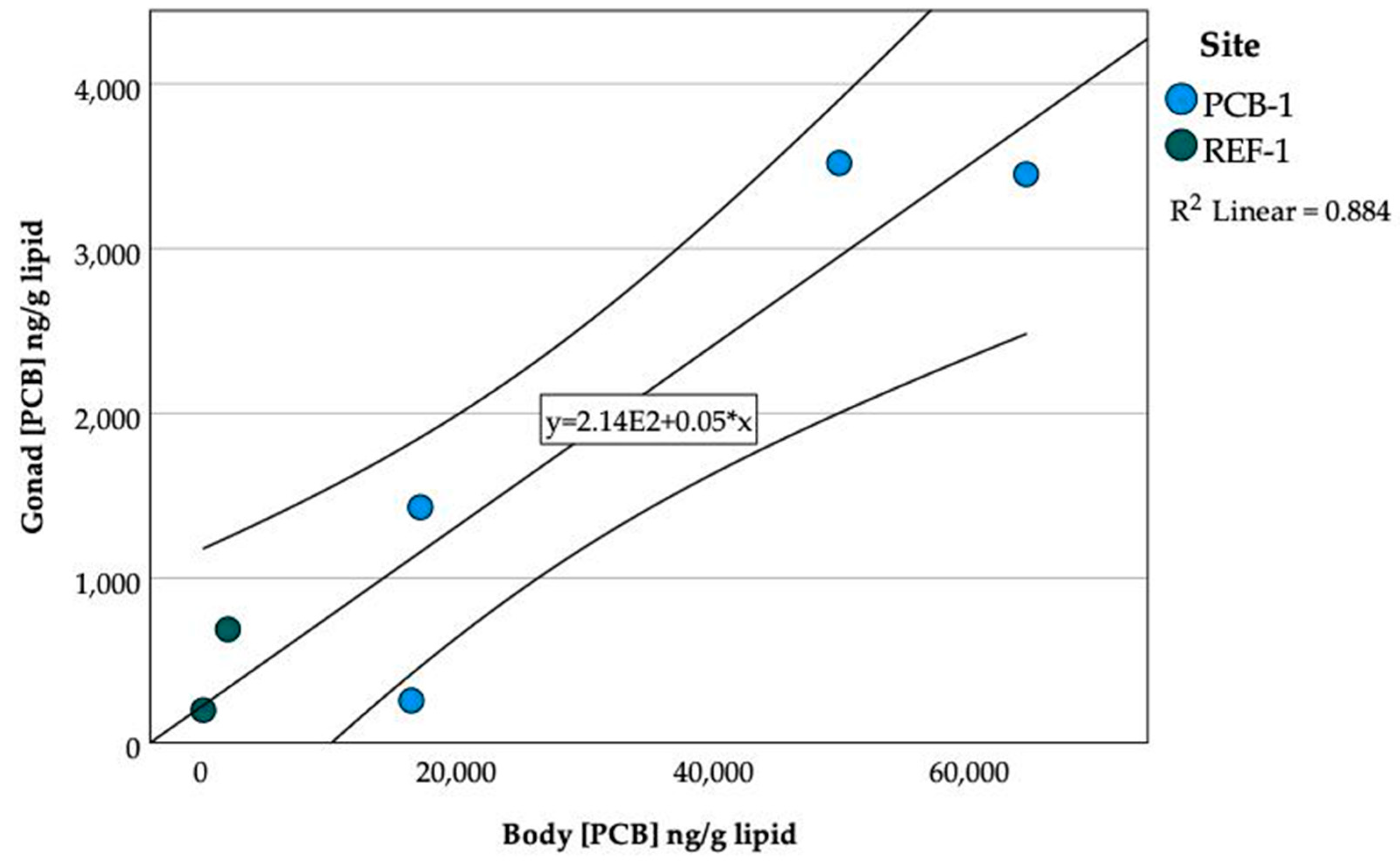
3.9. PCBs in Gonads and Embryos
3.10. Inter-Annual Site Variation
3.11. Other Contaminants
4. Discussion
4.1. Comparison of Tropical Species to Temperate-Zone Species
4.2. Comparison to Studies That Used Planktonic Embryos
4.3. Natural or “Background” Levels of Abnormalities
4.4. Johnston Atoll
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Adams, S.M.; Ham, K.D. Current Challenges in Contaminant Effects Monitoring: Multiple Stressors and Ecological Significance (No. CONF-9607154-1); Oak Ridge National Lab.: Oak Ridge, TN, USA, 1996. [Google Scholar]
- Jameson, S.C.; Erdmann, M.V.; Gibson, G.R., Jr.; Potts, K.W. Development of biological criteria for coral reef ecosystem assessment. Atoll Res. Bull. 1998, 450, 102. [Google Scholar] [CrossRef] [Green Version]
- Johannes, R.E.; Betzer, S.B. Introduction: Marine communities respond differently to pollution in the tropics than at higher latitudes. In Elsevier Oceanography Series; Elsevier: Amsterdam, The Netherlands, 1975; Volume 12, pp. 1–12. [Google Scholar]
- Peters, E.C.; Gassman, N.J.; Firman, J.C.; Richmond, R.H.; Power, E.A. Ecotoxicology of tropical marine ecosystems. Environ. Toxicol. Chem. 1997, 16, 12–40. [Google Scholar] [CrossRef]
- Erdmann, M.V.; Caldwell, R.L. Stomatopod crustaceans as bioindicators of marine pollution stress on coral reefs. In Proceedings of the Eighth International Coral Reef Symposium, Panama City, Panama, 24–29 June 1997; Volume 2, pp. 1521–1526. [Google Scholar]
- Hourigan, T.F.; Timothy, C.T.; Reese, E.S. Coral reef fishes as indicators of environmental stress in coral reefs. In Marine Organisms as Indicators; Springer: New York, NY, USA, 1988; pp. 107–135. [Google Scholar]
- Crosby, M.P.; Reese, E. A Manual for Monitoring Coral Reefs with Indicator Species: Butterflyfishes as Indicators of Change on Indo-Pacific Reefs; Office of Ocean and Coastal Resource Management, National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 1996; 45p. [Google Scholar]
- Doherty, P.J. Spatial and temporal patterns in recruitment. In The Ecology of Fishes on Coral Reefs; Sale, P., Ed.; Academic Press: San Diego, CA, USA, 1991; pp. 261–293. [Google Scholar]
- Alcolado, P.M.; Herrera-Moreno, A.; Martinez-Estalella, N. Sessile communities as environmental bio-monitors in Cuban coral reefs. Oceanogr. Lit. Rev. 1995, 8, 680. [Google Scholar]
- Stephens, J.S.; Hose, J.E.; Love, M.S. Fish assemblages as indicators of environmental change in nearshore environments. In Marine Organisms as Indicators; Springer: New York, NY, USA, 1988; pp. 91–105. [Google Scholar]
- Brown, B.E. Assessing environmental impacts on coral reefs. In Proceedings of the Sixth International Coral Reef Symposium, Townsville, Australia, 8–12 August 1988; Volume 1, pp. 71–80. [Google Scholar]
- Wenner, A.M. Crustaceans and other invertebrates as indicators of beach pollution. In Marine Organisms as Indicators; Springer: New York, NY, USA, 1988; pp. 199–229. [Google Scholar]
- Donaldson, E.M. Reproductive indices as measures of the effects of environmental stressors in fish. Am. Fish. Soc. Symp. 1990, 8, 109–122. [Google Scholar]
- McIntyre, A.D.; Pearce, J.B.; Longwell, A.C.; Hughes, J.B. Cytologic, cytogenetic, and developmental state of Atlantic mackerel eggs from sea surface waters of the New York Bight, and prospects for biological effects monitoring with ichthyoplankton [pollution, cytogenetic observations]. Rapp. Proces-Verbaux Reun. 1980, 179, 275–291. [Google Scholar]
- Chang, S.; Longwell, A. Examining statistical associations of malformation cyto-pathology and cytogenic abnormality of Atlantic mackerel embryos with indictor levels of environmental contaminants in the New York Bight. CM/ICES 1984, 11, 1–9. [Google Scholar]
- Hardy, J.; Kiesser, S.; Antrim, L.; Stubin, A.; Kocan, R.; Strand, J. The sea-surface microlayer of Puget Sound: Part I. Toxic effects on fish eggs and larvae. Mar. Environ. Res. 1987, 23, 227–249. [Google Scholar]
- Westernhagen, H.V.; Dethlefsen, V.; Cameron, P.; Berg, J.; Fürstenberg, G. Developmental defects in pelagic fish embryos from the western Baltic. Helgol. Mar. Res. 1988, 42, 13–36. [Google Scholar] [CrossRef] [Green Version]
- Klumpp, D.W.; Von Westernhagen, H. Biological effects of pollutants in Australian tropical coastal waters: Embryonic malformations and chromosomal aberrations in developing fish eggs. Mar. Pollut. Bull. 1995, 30, 158–165. [Google Scholar] [CrossRef]
- Kingsford, M.J.; Gray, C.A. Influence of pollutants and oceanography on abundance and deformities of wild fish larvae. In Detecting Ecological Impacts: Concepts and Applications in Coastal Habitats; Schmitt, R.J., Osenberg, C.W., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 235–255. [Google Scholar]
- Seitz, A.; Ratte, H.T. Aquatic ecotoxicology: On the problems of extrapolation from laboratory experiments with individuals and populations to community effects in the field. Comp. Biochem. Physiol. 1991, 100, 301–304. [Google Scholar] [CrossRef]
- Chapman, P.M. Extrapolating laboratory toxicity results to the field. Environ. Toxicol. Chem. 1996, 14, 927–930. [Google Scholar] [CrossRef]
- EPA. Ecological Risk Assessment Guidance for Superfund: Process for Designing and Conducting Ecological Risk Assessments—Interim Final, EPA 540/R-97-006; Office of Solid Waste and Emergency Response, U.S. Environmental Protection Agency: Washington, DC, USA, 1997.
- EPA. EPA 821/R-97-001 Method 1668: Toxic Polychlorinated Biphenyls by Isotope Dilution High Resolution Gas Chromatography/High Resolution Mass Spectrometry; U.S. Environmental Protection Agency: Washington, DC, USA, 1997.
- Allen, G.R. Damselfishes of the World; Mergus Publishers: Melle, Germany, 1991; p. 271. [Google Scholar]
- Lobel, L.K. Toxic caviar: Using fish embryos to monitor contaminant impacts. In Diving for Science 2011. Proceedings of the American Acade My of Underwater Sciences 30th Symposium; Pollock, N.W., Ed.; AAUS: Dauphin Island, AL, USA, 2011; pp. 34–39. [Google Scholar]
- Lobel, L.K.; Lobel, P.S. Choosing organisms for monitoring contaminant exposure on coral reefs: Case studies from Johnston Atoll, Central Pacific Ocean. In Proceedings of the American Academy of Underwater Sciences 34th Symposium; Lobel, L.K., Ed.; Diving for Science 2015: Key West, FL, USA, 2015; pp. 48–64. [Google Scholar]
- Kerr, L.M. Embryonic Abnormalities and Reproductive Output for the Damselfish, Abudefduf sordidus (Pomacentridae), Relative to Environmental Contamination at Johnston Atoll, Central Pacific Ocean. Master’s Thesis, Boston University, Boston, MA, USA, 1997. [Google Scholar]
- Kerr, L.M.; Lobel, P.S.; Ingoglia, J.M. Evaluation of a reporter gene system biomarker for detecting contamination in tropical marine sediments. Biol. Bull. 1999, 197, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Lobel, L.K.; Lobel, P.S. Junkyard damselfishes: Spawning behavior and nest site selection. In Proceedings of the AAUS/EDSP Joint International Diving Symposium, Curaçao, Netherlands Antilles, 24–27 October 2013; pp. 167–178. [Google Scholar]
- Kerr Lobel, L.; Drown, D.M.; Barber, P.H.; Lobel, P.S. A genetic assessment of parentage in the blackspot sergeant damselfish, Abudefduf sordidus (Pisces: Pomacentridae). Fishes 2019, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Shadrin, A.M.; Emel’yanova, N.G. Embryonic-larval development and some data on the reproductive biology of Abudefduf sexfasciatus (Pomacentridae: Perciformes). J. Ichthyol. 2007, 47, 67–80. [Google Scholar] [CrossRef]
- Wittenrich, M.L.; Turingan, R.G.; Cassiano, E.J. Rearing tank size effects feeding performance, growth, and survival of sergeant major, Abudefduf saxatilis, larvae. Aquac. Aquar. Conserv. Legis. 2012, 5, 393–402. [Google Scholar]
- Cairns, T.; Doose, G.M.; Froberg, J.E.; Jacobson, R.A.; Siegmund, E.G. Analytical chemistry of PCBs. PCBs Environ. 1986, 1, 1–45. [Google Scholar]
- Weis, J.S.; Weis, P. Effects of environmental pollutants on early fish development. Crit. Rev. Aquat. Sci. 1989, 1, 45–73. [Google Scholar]
- Freeman, H.C.; Sangalang, G.; Flemming, B. The sublethal effects of a polychlorinated biphenyl (Aroclor 1254) diet on the Atlantic cod (Gadus morhua). Sci. Total Environ. 1982, 24, 1–11. [Google Scholar] [CrossRef]
- Matsuyama, H.; Yano, T. Effect of PCB on the Steroid Hormone Metabolism in Goldfish Liver Microsome; Science Bulletin of the Faculty of Agriculture, Kyushu University: Fukuoka, Japan, 1987; Volume 42, pp. 1–7. [Google Scholar]
- Thomas, P. Reproductive endocrine function in female Atlantic croaker exposed to pollutants. Mar. Environ. Res. 1988, 24, 179–183. [Google Scholar] [CrossRef]
- Janssen, P.A.H.; Lambert, J.G.D.; Goos, H.T. The annual ovarian cycle and the influence of pollution on vitellogenesis in the flounder, Pleuronectes flesus. J. Fish Biol. 1995, 47, 509–523. [Google Scholar] [CrossRef]
- Hose, J.E.; Hannah, J.B.; Landolt, M.L.; Miller, B.S.; Felton, S.P.; Iwaoka, W.T. Uptake of benzo [α] pyrene by gonadal tissue of flatfish (family Pleuronectidae) and its effects on subsequent egg development. J. Toxicol. Environ. Health Part A Curr. Issues 1981, 7, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Von Westernhagen, H.; Rosenthal, H.; Dethlefsen, V.; Ernst, W.; Harms, U. Bioaccumulating substances and reproductive success in Baltic flounder Platichthys flesus. Aquat. Toxicol. 1981, 1, 85–99. [Google Scholar] [CrossRef]
- Hansen, P.D.; Von Westernhagen, H.; Rosenthal, H. Chlorinated hydrocarbons and hatching success in Baltic herring spring spawners. Mar. Environ. Res. 1985, 15, 59–76. [Google Scholar] [CrossRef]
- Halter, M.T.; Johnson, H.E. Acute toxicities of a polychlorinated biphenyl (PCB) and DDT alone and in combination to early life stages of coho salmon (Oncorhynchus kisutch). J. Fish. Board Can. 1974, 31, 1543–1547. [Google Scholar] [CrossRef]
- Schimmel, S.C.; Hansen, D.J.; Forester, J. Effects of aroclor (R) 1254 on laboratory-reared embryos and fry of sheepshead minnows (Cyprinodon variegatus). Trans. Am. Fish. Soc. 1974, 103, 582–586. [Google Scholar] [CrossRef]
- Billsson, K.; Westerlund, L.; Tysklind, M.; Olsson, P.E. Developmental disturbances caused by polychlorinated biphenyls in zebrafish (Brachydanio rerio). Mar. Environ. Res. 1998, 46, 461–464. [Google Scholar] [CrossRef]
- Black, D.E.; Phelps, D.K.; Lapan, R.L. The effect of inherited contamination on egg and larval winter flounder, Pseudopleuronectes americanus. Mar. Environ. Res. 1988, 25, 45–62. [Google Scholar] [CrossRef]
- Black, D.E.; Gutjahr-Gobell, R.; Pruell, R.J.; Bergen BMcElroy, A.E. Effects of non-ortho and mono-ortho polychlorinated biphenyls on reproduction in Fundulus heteroclitus (Linnaeus). Environ. Toxicol. Chem. 1998, 17, 1396–1404. [Google Scholar] [CrossRef]
- Black, D.E.; Gutjahr-Gobell, R.; Pruell, R.J.; Bergen, B.; Mills, L.; McElroy, A.E. Reproduction and polychlorinated biphenyls in Fundulus heteroclitus (Linnaeus) from new bedford harbor, massachusetts, USA. Environ. Toxicol. Chem. 1998, 17, 1405–1414. [Google Scholar] [CrossRef]
- Spies, R.B.; Rice, D.W. Effects of organic contamination on reproduction of the starry flounder (Platichthys stellatus) in San Francisco Bay. II. Reproductive success of fish captured in San Francisco Bay and spawned in the laboratory. Mar. Biol. 1988, 98, 191–200. [Google Scholar]
- Lobel, L.K.; Lobel, P.S. Contaminants in Fishes from Johnston Atoll. In 11th International Coral Reef Symposium; ReefBase: Fort Lauderdale, FL, USA, 2008. [Google Scholar]
- Lobel, L.K.; Lobel, P.S. Current status of the US military atolls in the Pacific: Johnston and Wake. In World Seas: An Environmental Evaluation; Academic Press: Cambridge, MA, USA, 2019; pp. 645–659. [Google Scholar]
- Long, E.R.; MacDonald, D.D.; Smith, S.L.; Calder, F.D. Incidence of adverse biological effects within ranges of chemical concentrations in marine and estuarine sediments. Environ. Manag. 1995, 19, 81–97. [Google Scholar] [CrossRef]
- O’Connor, T.P. The sediment quality guideline, ERL, is not a chemical concentration at the threshold of sediment toxicity. Mar. Pollut. Bull. 2004, 49, 383–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobel, P.S.; Lobel, L.K. Aspects of the biology and geomorphology of Johnston and Wake atolls, Pacific Ocean. In Coral Reefs of the USA; Springer: Dordrecht, The Netherlands, 2008; pp. 655–689. [Google Scholar]
- Randall, J.E.; Lobel, P.S.; Chave, E.H. Annotated checklist of the fishes of Johnston Island. Pac. Sci. 1985, 39, 24–80. [Google Scholar]
- Kosaki, R.K.; Pyle, R.L.; Randall, J.E.; Irons, D.K. New records of fishes from Johnston Atoll, with notes on biogeography. Pac. Sci. 1991, 45, 186–203. [Google Scholar]
- Lobel, P.S. Marine life of Johnston Atoll, Central Pacific Ocean; Natural World Press: Vida, OR, USA, 2003. [Google Scholar]
- Lobel, P.S.; Kerr, L.M. Status of contaminants in Johnston Atoll lagoon sediments after 70 years of US military activity. In Proceedings of the Ninth International Coral Reef Symposium, Bali, Indonesia, 23–27 October 2000; Volume 2, pp. 861–866. [Google Scholar]
- Shaw, E.S. The embryology of the sergeant major, Abudefduf saxatilis. Copeia 1955, 1955, 85–89. [Google Scholar] [CrossRef]
- Fritzsche, R.A. Chaetodontidae through Ophidiidae; in Development of Fishes of the Mid-Atlantic Bight; Fish and Wildlife Service, US Department of the Interior: Washington, DC, USA, 1978; Volume 4, 340p. [Google Scholar]
- Cameron, P.; Berg, J. Morphological and chromosomal aberrations during embryonic development in dab, Limanda. Mar. Ecol. Prog. Ser. 1992, 91, 163–169. [Google Scholar] [CrossRef]
- Van den Berg, M.; Birnbaum, L.; Bosveld, A.T.C.; Brunstrom, B.; Cook, P.; Freeley, M.; Giesey, J.P.; Hanberg, A.; Hasegawa, R.; Kennedy, S.W.; et al. Toxic equivalency factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ. Health Perspect. 1998, 106, 775–792. [Google Scholar] [CrossRef]
- Zabel, E.W.; Cook, P.M.; Peterson, R.E. Toxic equivalency factors of polychlorinated dibenzo-p-dioxin, dibenzofuran and biphenyl congeners based on early life stage mortality in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 1995, 31, 315–328. [Google Scholar] [CrossRef]
- Lobel, L.M.K.; Davis, E.A. Immunohistochemical detection of polychlorinated biphenyls in field collected damselfish (Abudefduf sordidus; Pomacentridae) embryos and larvae. Environ. Pollut. 2002, 120, 529–532. [Google Scholar] [CrossRef]
- Pruell, R.J.; Rubinstein, N.I.; Taplin, B.K.; LiVolsi, J.A.; Bowen, R.D. Accumulation of polychlorinated organic contaminants from sediment by three benthic marine species. Arch. Environ. Contam. Toxicol. 1993, 24, 290–297. [Google Scholar] [CrossRef]
- Crimmins, B.S.; Brown, P.D.; Kelso, D.P.; Foster, G.D. Bioaccumulation of PCBs in aquatic biota from a tidal freshwater marsh ecosystem. Arch. Environ. Contam. Toxicol. 2002, 42, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Cameron, P.; Berg, J.; Dethlefsen, V.; Von Westernhagen, H. Developmental defects in pelagic embryos of several flatfish species in the southern North Sea. Neth. J. Sea Res. 1992, 29, 239–256. [Google Scholar] [CrossRef]
- Longwell, A.C.; Chang, S.; Hebert, A.; Hughes, J.B.; Perry, D. Pollution and developmental abnormalities of Atlantic fishes. Environ. Biol. Fishes 1992, 35, 1–21. [Google Scholar] [CrossRef]
- Atlas, E.; Bidleman, T.; Giam, C.S. Atmospheric transport of PCBs to the oceans. In PCBs and the Environment; CRC Press: Boca Raton, FL, USA, 1986; Volume 1, pp. 79–100. [Google Scholar]
- Stout, V.F. What is happening to PCBs. Elements of Effective Environmental Monitoring as Illustrated by an Analysis of PCB Trends in Terrestrial and Aquatic Organisms. In PCBs and the Environment; CRC Press: Boca Raton, FL, USA, 1986; Volume 1, pp. 163–206. [Google Scholar]
- Yannopoulos, A.; Yannopoulos, C. Fish egg mortality and abnormal embryogenesis. Rapp. Proces Verbaux Reun. Comm. Int. Pour I’Explor. 1981, 27, 143–146. [Google Scholar]
- Rønnestad, I.; Robertson, R.; Fyhn, H.J. Free amino acids and protein content in pelagic and demersal eggs of tropical marine fishes. In The Fish Egg; MacKinlay, D.D., Eldridge, M., Eds.; American Fisheries Society: Vancouver, BC, Canada, 1996; pp. 81–84. ISBN 0-9698631-0-7. [Google Scholar]
- Rosenthal, H.; Alderdice, D.F. Sublethal effects of environmental stressors, natural and pollutional, on marine fish eggs and larvae. J. Fish. Res. Board Can. 1976, 33, 2047–2065. [Google Scholar]
- Cross, J.N.; Ellen, H.J. Evidence for impaired reproduction in white croaker (Genyonemus lineatus) from contaminated areas off southern California. Mar. Environ. Res. 1988, 24, 185–188. [Google Scholar] [CrossRef]
- Forbes, V.E.; Moller, V.; Depledge, M.H. Intrapopulation variability in sublethal response to heavy metal stress in sexual and asexual gastropod populations. Funct. Ecol. 1995, 9, 477–484. [Google Scholar] [CrossRef]
- Callaghan, A.; Holloway, G.J. The relationship between environmental stress and variance. Ecol. Appl. 1999, 9, 456–462. [Google Scholar] [CrossRef]
- Forbes, V.E.; Depledge, M.H. Environmental stress and the distribution of traits within populations. In ECOtoxicology: Ecological Dimensions; Springer: Dordrecht, The Netherlands, 1996; pp. 71–86. [Google Scholar]
- Kerr, L.M.; Lang, K.L.; Lobel, P.S. PCB contamination relative to age for a Pacific damselfish, Abudefduf sordidus (Pomacentridae). Biol. Bull. 1997, 193, 279–281. [Google Scholar] [CrossRef]
- Itzkowitz, M. Habitat selection and subsequent reproductive success in the beaugregory damselfish. Environ. Biol. Fishes 1991, 30, 287–293. [Google Scholar] [CrossRef]
- Itzkowitz, M.; Itzkowitz, D.E.; Shelly, D. Territory use and disuse in the beaugregory damselfish. Bull. Mar. Sci. 1995, 57, 653–662. [Google Scholar]
- Eskelinen, P. Effects of different diets on egg production and egg quality of Atlantic salmon (Salmo salar L.). Aquaculture 1989, 79, 275–281. [Google Scholar] [CrossRef]
- Lovell, T. Nutrition and Feeding of Fish; Van Nostrand Reinhold: New York, NY, USA, 1989; Volume 260. [Google Scholar]
- Craik, J.C.A. Egg quality and egg pigment content in salmonid fishes. Aquaculture 1985, 47, 61–88. [Google Scholar] [CrossRef]
- Hubbs, C.; Stavenhagen, L. Effects of maternal carotenoid deficiency on the viability of darter (Osteichthyes) offspring. Physiol. Zool. 1958, 31, 280–283. [Google Scholar] [CrossRef]
- Bodammer, J.E. The teratological and pathological effects of contaminants on embryonic and larval fishes exposed as embryos: A brief review. In Water Quality and the Early Life Stages of Fishes; Fuiman, L.A., Ed.; American Fisheries Society: Bethesda, MD, USA, 1993; pp. 77–84. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | PCBs (ng/g) | TEQ (ng/g) | PAHs (ng/g) | As | Sb | Ba | Cd | Cu | Cr | Pb | Hg | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCB-1 | 28.6 (61.0) | 0.6 (0.8) | NA | 0.6 (0.4) | 0.8 (0.5) | NA | NA | 8.1 (14.3) | NA | 31.0 (35.1) | 0.02 (0.02) | 12.9 (16.8) |
| PCB-2 | 86.6 (126.9) | NA | ND | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| REF-1 | 0.45 (0.1) | NA | J | 0.4 (0) | NA | 172.2 (30.6) | ND | ND | 10.4 (0.7) | 39.2 (1.5) | ND | ND |
| REF-2 | 0.6 (0.2) | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| REF-3 | 0.7 (0.4) | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| HO | 2.4 (2.0) | 16.9 (113.6) | 589.6 (1845.6) | 0.6 (0.3) | NA | 76.3 (67.3) | ND | 5.1 (3.9) | 10.2 (9.0) | 10.1 (9.5) | 0.01 (0.003) | 13.0 (23.7) |
| Site | Nest | # Abnormal Embryos | # Normal Embryos | % Abnormal Hatching | % Normal Hatching | Abnormality Type |
|---|---|---|---|---|---|---|
| PCB-1 | sp5 | 6 | 14 | 17 | 100 | Developmental retardation, no differentiation, abnormal gastrulation |
| PCB-1 | e1 | 8 | 0 | 13 | NA | Abnormal cleavage |
| PCB-1 | h | 7 | 21 | 43 | 95 | Loose cells, enlarged segmentation cavity, abnormal gastrulation |
| PCB-1 | p | 1 | 3 | 0 | 100 | Abnormal gastrulation |
| PCB-1 | f2 | 4 | 12 | 0 | 100 | Developmental retardation, axial, abnormal gastrulation |
| PCB-1 | v | 3 | 9 | 0 | 89 | No differentiation, enlarged segmentation cavity, abnormal gastrulation |
| PCB-1 | k | 4 | 18 | 0 | 100 | Loose cells, enlarged segmentation cavity, abnormal blastodisc |
| PCB-1 | ag | 4 | 12 | 25 | 100 | Enlarged segmentation cavity, no differentiation |
| PCB-1 | d | 2 | 6 | 0 | 100 | Abnormal keel, early blastopore closure |
| PCB-1 | 3 | 4 | 12 | 25 | 100 | Enlarged segmentation cavity |
| PCB-1 | q | 2 | 6 | 50 | 100 | Enlarged segmentation cavity, abnormal gastrulation |
| PCB-1 | e2 | 14 | 34 | 0 | 88 | Loose cells, enlarged segmentation cavity |
| PCB-1 | q | 4 | 9 | 25 | 89 | Enlarged segmentation cavity, abnormal gastrulation |
| PCB-1 | b | 12 | 35 | 8 | 94 | Abnormal keel, abnormal gastrulation |
| PCB-1 | g | 24 | 40 | 0 | 63 | Axial, abnormal cell proliferations, abnormal keel, no differentiation |
| PCB-1 | o | 8 | 23 | 0 | 87 | Loose cells, abnormal gastrulation |
| PCB-1 | d | 8 | 25 | 0 | 80 | Loose cells, enlarged segmentation cavity |
| REF-1 | b14-5 | 8 | 24 | 0 | 100 | Abnormal yolk, no differentiation, abnormal keel |
| REF-2 | tug10 | 12 | 20 | 0 | 100 | Loose cells, enlarged segmentation cavity |
| REF-2 | tug | 16 | 16 | 31 | 88 | Loose cells |
| REF-3 | east | 15 | 15 | 40 | 100 | Abnormal yolk, abnormal cell proliferations, Abnormal gastrulation |
| Total | 166 | 354 | 6.7 | 90.4 |
| Site | Sex | SL (mm) | Total Weight (g) | Gonad Weight (g) | GSI | Liver Weight (g) | LSI |
|---|---|---|---|---|---|---|---|
| PCB-1 | M | 140 | 173. 8 | 1.1 | 0.63 | 1.2 | 0.71 |
| F | 142 | 188.1 | 3.9 | 2.08 | 1.2 | 0.65 | |
| F | 135 | 148.5 | 3.0 | 2.01 | 1.4 | 0.94 | |
| M | 146 | 199.5 | 1.8 | 0.89 | 1.9 | 0.98 | |
| M | 135 | 157.7 | 1.1 | 0.72 | 1.3 | 0.82 | |
| M | 165 | 238.0 | 1.4 | 0.59 | 2.2 | 0.94 | |
| Mean (SD) | 143.8 (11.2) | 184.3 (32.4) | 2.0 (1.15) | 1.15 (0.70) | 1.6 (0.4) | 0.84 (0.13) | |
| REF-1 | M | 145 | 204.8 | 0.9 | 0.46 | 1.7 | 0.81 |
| M | 149 | 196.7 | 0.9 | 0.46 | 1.6 | 0.79 | |
| M | 143 | 178.5 | 1.3 | 0.73 | 1.7 | 0.94 | |
| M | 160 | 238.5 | 1.9 | 0.81 | 2.8 | 1.15 | |
| M | 137 | 157.7 | 1.3 | 0.79 | 1.4 | 0.90 | |
| M | 145 | 188.5 | 1.1 | 0.57 | 1.9 | 1.02 | |
| Mean (SD) | 146.5 (7.7) | 194.1 (27.2) | 1.2 (0.4) | 0.64 (0.16) | 1.8 (0.5) | 0.94 (0.14) | |
| REF-3 | M | 137 | 179.6 | 1.1 | 0.62 | 0.9 | 0.52 |
| M | 139 | 165.1 | 0.8 | 0.47 | 1.0 | 0.63 | |
| M | 145 | 205.9 | 1.4 | 0.68 | 1.3 | 0.63 | |
| M | 163 | 257.3 | 1.7 | 0.67 | 3.3 | 1.26 | |
| M | 151 | 222.4 | 1.4 | 0.61 | 1.6 | 0.71 | |
| M | 148 | 220.8 | 1.9 | 0.86 | 2.0 | 0.92 | |
| Mean (SD) | 147.2 (9.4) | 208.5 (33.0) | 1.4 (0.4) | 0.65 (0.13) | 1.7 (0.9) | 0.78 (0.27) | |
| HO | F | 144 | 208.5 | 10.8 | 5.17 | 2.2 | 1.04 |
| M | 145 | 179.7 | 1.3 | 0.71 | 1.7 | 0.95 | |
| F | 145 | 200.8 | 7.8 | 3.86 | 1.8 | 0.88 | |
| F | 146 | 208.7 | 2.2 | 1.05 | 1.9 | 0.93 | |
| F | 150 | 234.1 | 3.3 | 1.40 | 1.6 | 0.67 | |
| F | 137 | 197.0 | 10.1 | 5.13 | 1.3 | 0.68 | |
| Mean (SD) | 144.5 (4.2) | 204.8 (17.9) | 5.9 (4.2) | 2.89 (2.07) | 1.8 (0.3) | 0.86 (0.15) |
| Site | N | SL (mm) | Weight (g) | Sex (M/F) | Lipid (%) | PCBs (ng/g) | |
|---|---|---|---|---|---|---|---|
| 1996 | |||||||
| PCB-1 | 6 | 152 (4.7) | 230.4 (16.8) | 5/1 | 4.35 (1.6) | 1591.1 a (1122.2) | |
| REF-1 | 7 | 137 (8.9) | 170.8 (32.6) | 5/2 | 4.4 (2.5) | 68.2 b (77.2) | |
| 1999 | |||||||
| PCB-1 | 6 | 156 (12.6) | 235.5 (41.8) | 3/3 | 4.2 (1.5) | 259.9 b (308.0) | |
| PCB-2 | 8 | 147 (39.3) | 179.7 (11.7) | 5/3 | 2.6 (0.5) | 3586.8 a (1957.9) | |
| REF-1 | 6 | 135 (12.7) | 147.4 (40.3) | 5/1 | 2.0 (0.7) | 18.5 b (20.3) | |
| REF-3 | 6 | 146 (10.2) | 204.4 (40.9) | 2/4 | 3.6 (2.4) | 9.4 b (5.1) | |
| 2001 | |||||||
| PCB-1 | 6 | 144 (11.2) | 184.3 (32.4) | 4/2 | 3.1 (0.9) | 409.4 a (333.1) | |
| REF-1 | 6 | 146 (7.7) | 194.1 (27.2) | 6/0 | 4.1 (1.6) | 17.8 b (7.7) | |
| REF-3 | 6 | 147 (0.4) | 208.5 (33.0) | 6/0 | 4.3 (1.9) | 16.1 b (11.6) | |
| HO | 6 | 144 (4.2) | 204.8 (17.8) | 1/5 | 3.8 (1.5) | 35.0 b (27.6) |
| Site | Nests | N # | Mean % Abnormal | CV | Range | Mean PCBs (ng/g Lipid) | |
|---|---|---|---|---|---|---|---|
| 1996 | |||||||
| PCB-1 | 46 | 145 | 5.8 (15.6) a | 271 | 0–100 | 39,853.4 a (32,035.0) | |
| REF-1 | 26 | 71 | 1.4 (1.1) b | 77 | 0–5.9 | 1819.7 b (2387.7) | |
| 1998 | |||||||
| PCB-1 | 41 | 385 | 6.0 (18.2) a | 302 | 0–100 | NA | |
| REF-1 | 28 | 30 | 1.4 (2.1) b | 153 | 0–9.9 | NA | |
| 1999 | |||||||
| PCB-1 | 30 | 58 | 2.8 (2.4) a | 87 | 0–8.3 | 10,419.2 b (14,347.3) | |
| PCB-2 | 5 | 10 | 6.2 (10.4) a | 174 | 0–34.7 | 138,032.5 a (65,773.7) | |
| REF-1 | 16 | 23 | 1.8 (1.4) a | 79 | 0–5.7 | 976.4 b (902.2) | |
| REF-2 | 8 | 16 | 2.5 (1.9) a | 78 | 0.5–7.8 | NA | |
| REF-3 | 10 | 13 | 1.7 (1.3) a | 75 | 0–3.9 | 350.3 b (301.75) | |
| 2001 | |||||||
| PCB-1 | 35 | 144 | 3.2 (5.8) a | 183 | 0–35.9 | 14,643.3 a (13,545.8) | |
| PCB-2 | 7 | 27 | 2.0 (2.5) a | 129 | 0–13.0 | NA | |
| REF-1 | 31 | 88 | 1.7 (3.9) b | 175 | 0–22.1 | 434.7 b (45.0) | |
| REF-2 | 13 | 24 | 1.3 (1.3) b | 98 | 0–4.8 | NA | |
| REF-3 | 12 | 36 | 1.6 (1.6) b | 99 | 0–7.4 | 364.6 b (155.4) | |
| HO | 2 | 5 | 2.9 (4.8) a | 175 | 0–11.5 | 894.3 b (520.3) |
| Sample | Site | Gonad PCB (ng/g) | Gonad Lipid (%) | Gonad PCBs (ng/g Lipid) | Body PCB (ng/g) * | Body Lipid (%) | Body PCBs (ng/g Lipid) |
|---|---|---|---|---|---|---|---|
| 2 | PCB-1 | 76.8 | 5.4 | 1430.2 | 783.2 | 4.1 | 17,145.6 |
| 3 | PCB-1 | 282.0 | 8.2 | 3451.7 | 2234 | 3.0 | 64,210.5 |
| 4 | PCB-1 | 170.0 | 4.8 | 3519.7 | 2079 | 3.8 | 49,713.5 |
| 7 | PCB-1 | 26.2 | 10.2 | 256.9 | 463.2 | 2.7 | 16,245.4 |
| 1 | REF-1 | 8.3 | 4.2 | 198.8 | 18.14 | 3.7 | 264.4 |
| 6 | REF-1 | 48.1 | 6.9 | 689.1 | 251.3 | 9.3 | 2175.6 |
| Site | Nest | PCBs (ng/g Lipid) | % Abnormal |
|---|---|---|---|
| PCB-1 | AE | 414.7 | 4.3 |
| PCB-1 | J | 1223.1 | 3.4 |
| PCB-1 | I | 13,000.0 | 8.3 |
| PCB-2 | Navy | 7117.6 | 34.7 |
| REF-1 | B14 | 790.5 | 0.5 |
| REF-1 | B14-2 | 3713.0 | 1.0 |
| PCB Congener Concentration (ng/g Lipid) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PCB | PCB-1 | PCB-2 | REF-1 | REF-3 | HO | |||||
| 1996 | 1999 | 2001 | 1999 | 1996 | 1999 | 2001 | 1999 | 2001 | 2001 | |
| Mono-ortho PCBs | ||||||||||
| 118 | NA | 341.6 (626.0) | 1548.2 (1644.2) | 2294.0 (1418.1) | 51.1 (56.3) | 32.4 (16.9) | 22.8 (10.0) | 36.5 (35.0) | 31.9 (6.6) | 20.2 (9.6) |
| 105 | NA | 325.5 (689.3) | 901.0 (861.8) | 245.0 (105.0) | 35.9 (36.4) | 16.9 (8.9) | 13.5 (2.6) | 22.0 (24.4) | 20.3 (3.6) | 9.5 (4.2) |
| 167 | NA | NA | 170.6 (132.9) | 2658.6 (1051.3) | NA | NA | 7.0 (1.5) | NA | 4.7 (2.7) | 8.9 (5.5) |
| 156 | NA | 477.5 (827.4) | 388.3 (346.2) | 1923.8 (801.3) | 30.1 (32.2) | 17.1 (11.2) | 10.4 (2.2) | 10.1 (10.7) | 7.8 (2.1) | 8.8 (4.7) |
| 157 | NA | NA | 90.2 (83.2) | 228.5 (90.4) | NA | NA | 2.4 (0.4) | NA | 1.9 (0.6) | 2.3 (1.2) |
| 189 | NA | NA | 20.4 (15.9) | 808.8 (379.6) | 8.2 (12.0) | NA | 1.8 (0.3) | NA | 0.7 (0.2) | 3.8 (2.3) |
| Non-ortho PCBs | ||||||||||
| 77 | NA | 3.7 (4.3) | ND (5.9) | 195 (36.6) | 3.0 (2.8) | 0.3 (0.1) | ND (0.1) | 0.3 (0.4) | ND (0.2) | ND (0.2) |
| 81 | NA | 2.0 (2.6) | ND (2.6) | 195 (36.6) | 0.8 (1.4) | 0.1 (0.1) | ND (0.03) | 0.003 (0.005) | ND (0.05) | ND (0.04) |
| 126 | NA | 10.6 (17.2) | 8.6 (5.8) | 431.6 (219.7) | 0.8 (0.9) | 0.5 (0.5) | 0.6 (0.1) | 0.4 (0.4) | 0.5 (0.2) | 1.4 (0.7) |
| 169 | NA | 2.2 (4.5) | 0.4 (0.3) | 133.8 (78.9) | ND (0.0009) | 0.05 (0.1) | 0.08 (0.01) | 0.05 (0.1) | 0.08 (0.05) | 0.4 (0.2) |
| Sum | NA | 1163.1 | 3127.7 | 9114.1 | 129.9 | 67.3 | 58.6 | 69.4 | 67.9 | 55.3 |
| TEQ a | NA | 0.0071 | 0.0043 | 0.3617 | 0.0013 | 0.00035 | 0.0003 | 0.0003 | 0.0003 | 0.00072 |
| TEQ b | NA | NA | 0.0463 | NA | NA | NA | 0 | NA | 0.0001 | 0.4922 |
| Site | As | Ba | Cd | Cr | Cu | Hg | Pb | Sb | Zn |
|---|---|---|---|---|---|---|---|---|---|
| PCB-1 | 2.17 (0.52) | 0.80 bc (0.52) | 0.04 (0.01) | 0.45 (0.07) | 0.73 (0.17) | 0.08 a (0.04) | 0.73 b (2.14) | ND | 20.68 (5.08) |
| REF-1 | 2.05 (0.77) | 1.35 a (0.57) | 0.08 (0.05) | 0.34 (0.07) | 0.59 (0.07) | 0.02 a (0.00) | 0.59 a (0.26) | ND | 19.07 (3.43) |
| REF-3 | 2.63 (0.21) | 0.49 b (0.17) | 0.09 (0.05) | 0.53 (0.28) | 0.69 (0.05) | 0.14 b (0.14) | 0.69 a (0.05) | ND | 18.05 (2.63) |
| HO | 2.03 (0.19) | 0.95 ac (0.29) | 0.04 (0.02) | 0.61 (0.46) | 0.61 (0.07) | 0.03 a (0.01) | 0.61 a (0.25) | 0.01 (0) | 17.40 (4.24) |
| WCS * | 2.10 | 0.95 | 0.29 | 0.45 | 0.65 | 0.02 | 0.07 | ND | 11.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobel, L.K.; Lobel, P.S. Damselfish Embryos as a Bioindicator for Military Contamination on Coral Reefs at Johnston Atoll, Pacific Ocean. Fishes 2022, 7, 55. https://doi.org/10.3390/fishes7020055
Lobel LK, Lobel PS. Damselfish Embryos as a Bioindicator for Military Contamination on Coral Reefs at Johnston Atoll, Pacific Ocean. Fishes. 2022; 7(2):55. https://doi.org/10.3390/fishes7020055
Chicago/Turabian StyleLobel, Lisa Kerr, and Phillip S. Lobel. 2022. "Damselfish Embryos as a Bioindicator for Military Contamination on Coral Reefs at Johnston Atoll, Pacific Ocean" Fishes 7, no. 2: 55. https://doi.org/10.3390/fishes7020055
APA StyleLobel, L. K., & Lobel, P. S. (2022). Damselfish Embryos as a Bioindicator for Military Contamination on Coral Reefs at Johnston Atoll, Pacific Ocean. Fishes, 7(2), 55. https://doi.org/10.3390/fishes7020055