
Behavioral Interactions between a Native and an Invasive Fish Species in a Thermally Heterogeneous Experimental Chamber


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Fish
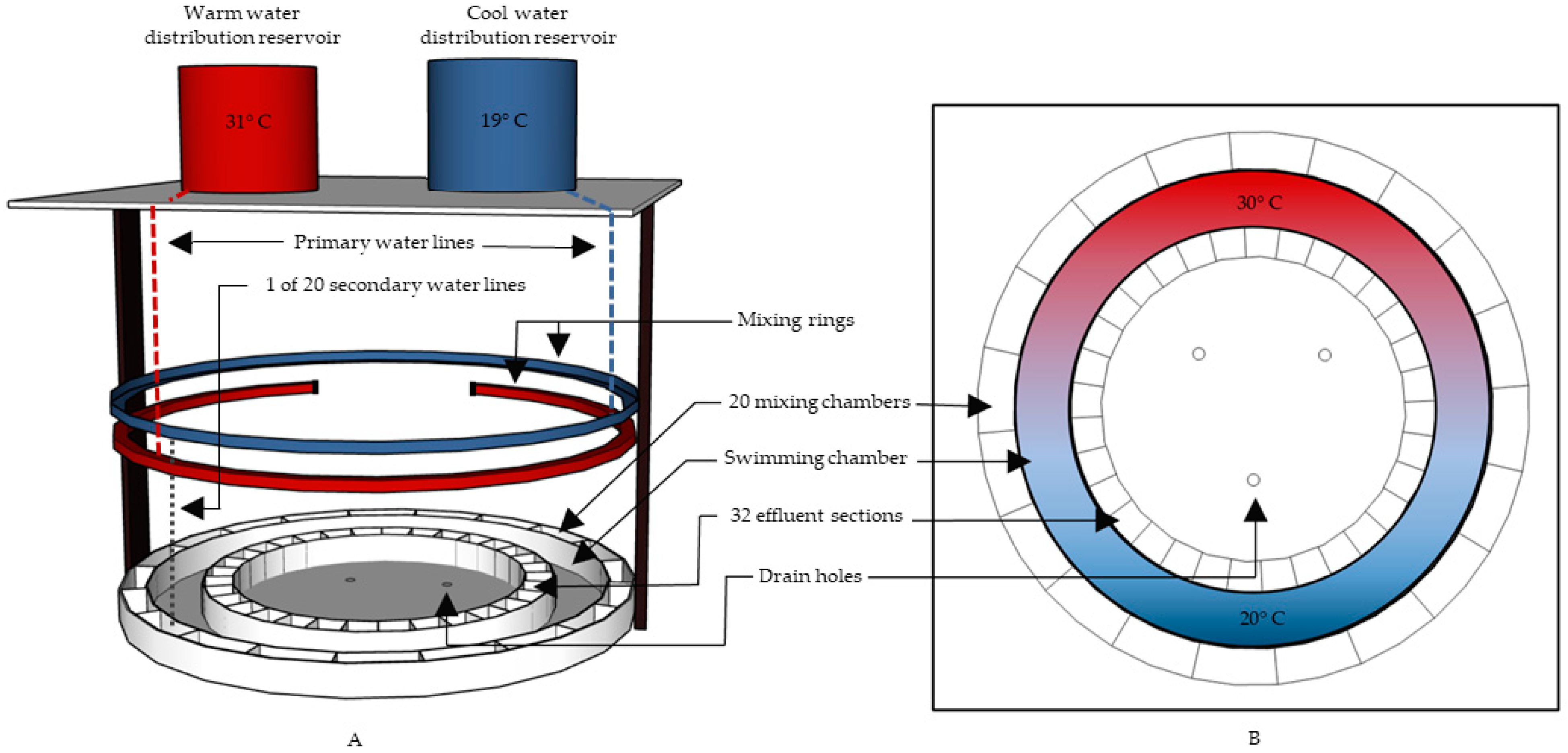
2.3. Experimental Apparatus
2.4. Experimental Fish Trials
2.5. Statistical Methods
3. Results
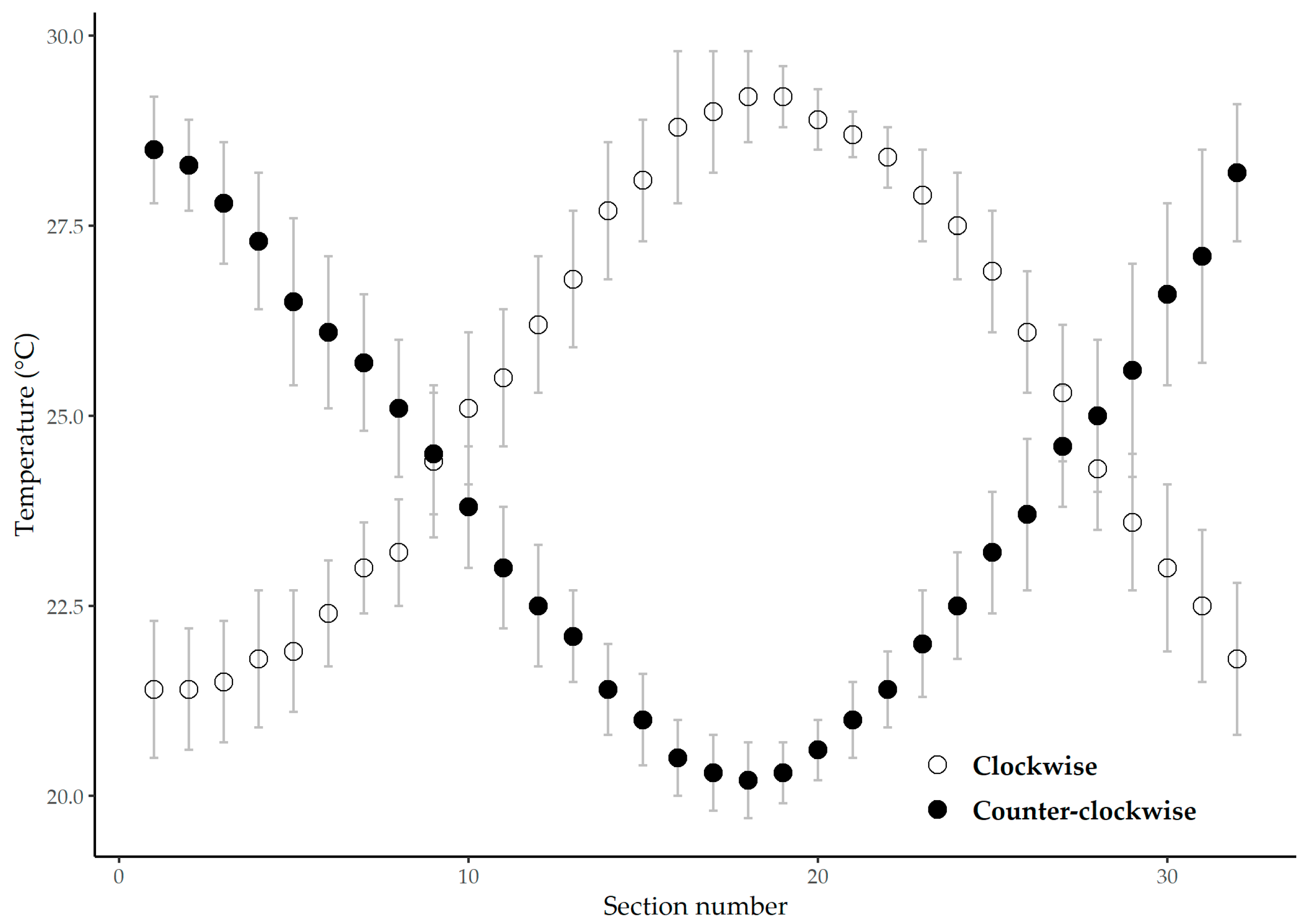
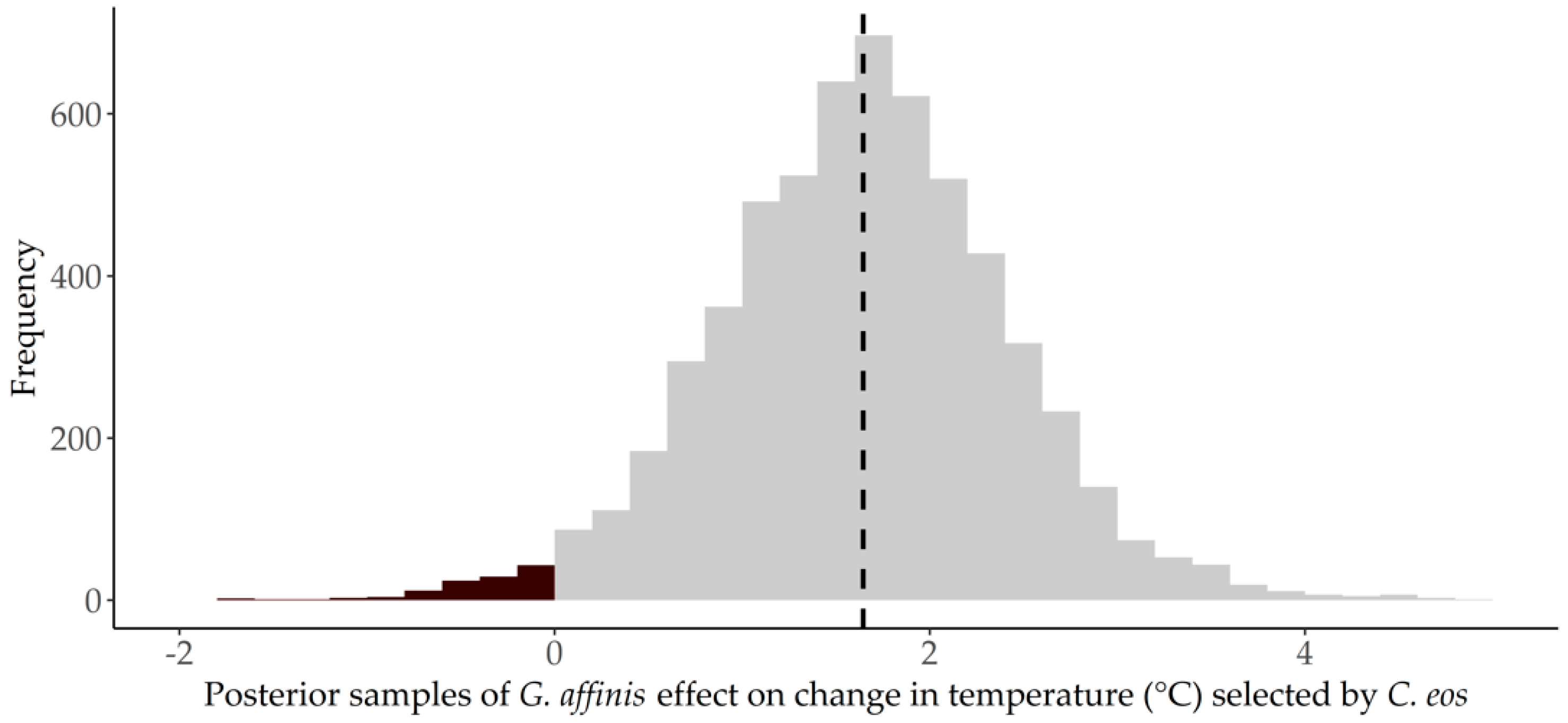
3.1. Thermal Preference
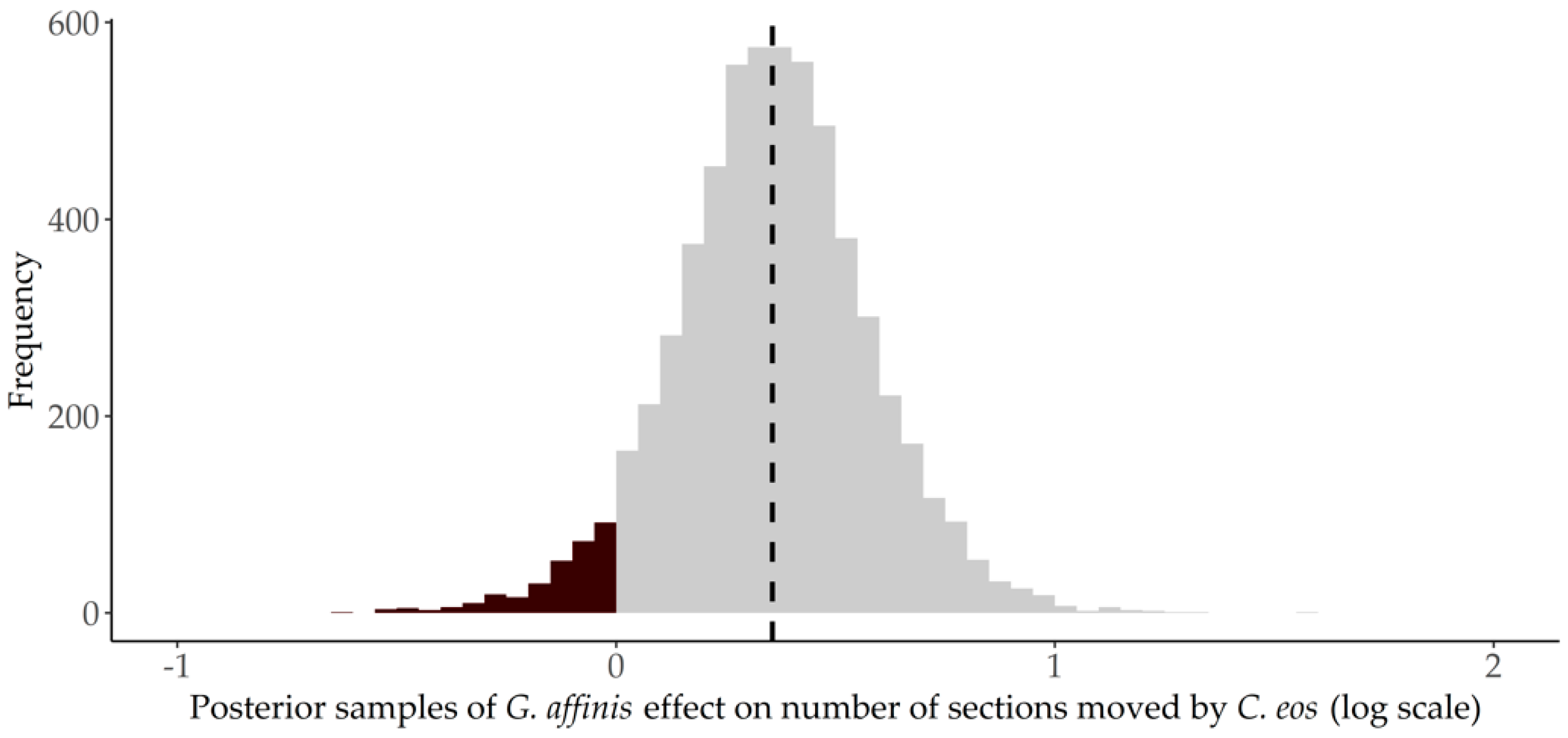
3.2. Movement
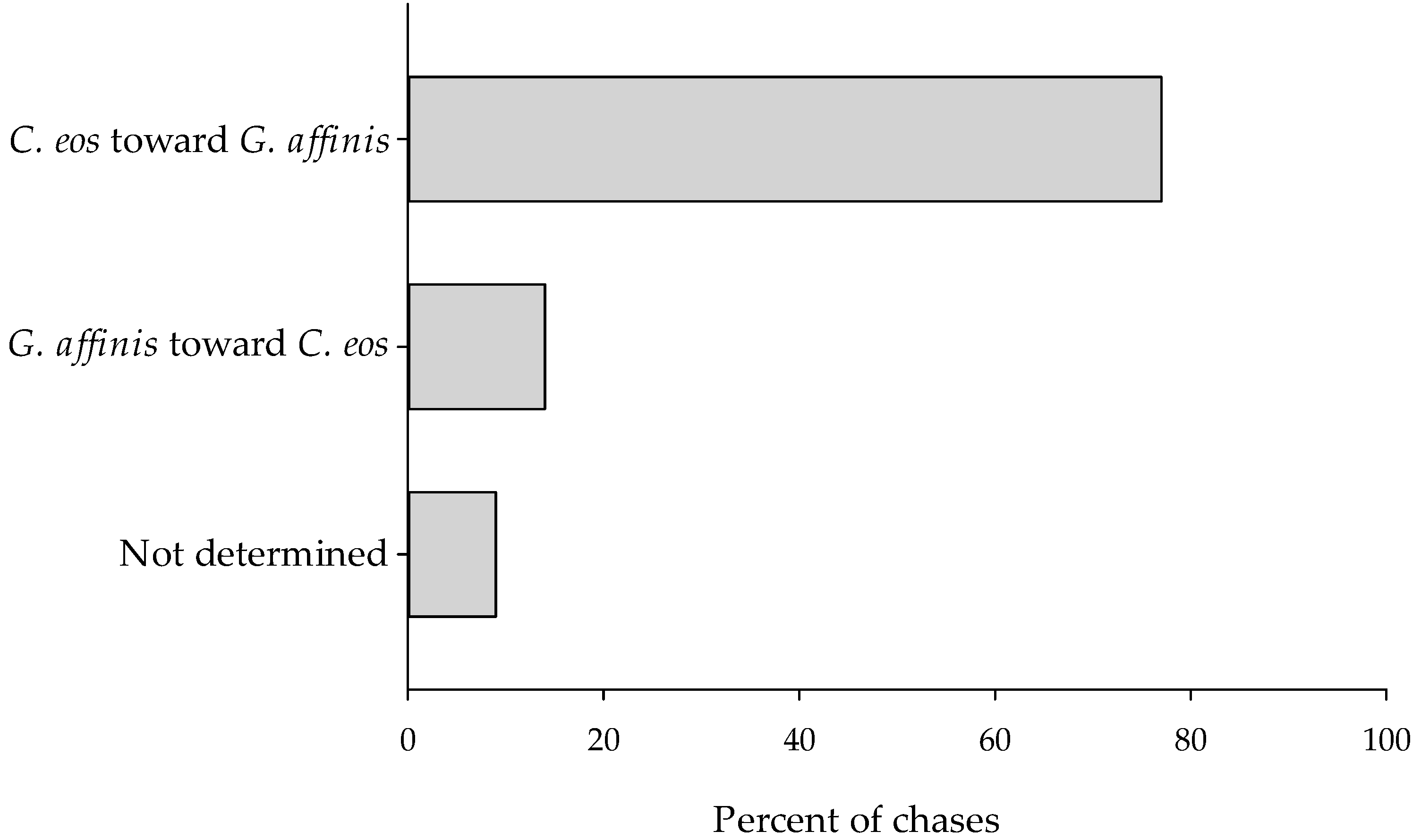
3.3. Agonistic Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olden, J.D.; Poff, N.L. Toward a mechanistic understanding and prediction of biotic homogenization. Am. Nat. 2003, 162, 442–460. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilá, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Lapointe, N.W.R.; Marchetti, M.P. Non-indigenous fishes and their role in freshwater imperilment. In Conservation of Freshwater Fishes; Closs, G.P., Krkosek, M., Olden, J.D., Eds.; Cambridge University Press: Cambridge, UK, 2016; Volume 1, pp. 238–269. [Google Scholar]
- Van Snik Gray, E.; Stauffer, J.R., Jr. Substrate choice by three species of darters (Teleostei: Percidae) in an artificial stream: Effects of a nonnative species. Copeia 2001, 2001, 254–261. [Google Scholar] [CrossRef]
- Hasegawa, K.; Yamamoto, T.; Murakami, M.; Maekawa, K. Comparison of competitive ability between native and introduced salmonids: Evidence from pairwise contests. Ichthyol. Res. 2004, 51, 191–194. [Google Scholar] [CrossRef]
- McMahon, T.E.; Zale, A.V.; Barrows, F.T.; Selong, J.H.; Danehy, R.J. Temperature and competition between Bull Trout and Brook Trout: A test of the elevation refuge hypothesis. Trans. Am. Fish. Soc. 2007, 136, 1313–1326. [Google Scholar] [CrossRef]
- Courtenay, W.R., Jr.; Meffe, G.K. Small fishes in strange places: A review of introduced poeciliids. In Ecology and Evolution of Livebearing Fishes (Poeciliidae); Meffe, G.K., Snelson, F.F., Jr., Eds.; Prentice Hall: Englewood Cliffs, NJ, USA, 1989; pp. 319–331. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species. A Selection from the Global Invasive Species Database. Published by the Invasive Species Specialist Group. Available online: https://rewilding.org/wp-content/uploads/2012/04/IUCN-GISP.pdf (accessed on 15 September 2021).
- Schumann, D.A.; Hoback, W.W.; Koupal, K.D. Complex interactions between native and invasive species: Investigating the differential displacement of two topminnows native to Nebraska. Aquat. Invasions 2015, 10, 339–346. [Google Scholar] [CrossRef]
- Segev, O.; Mangel, M.; Blaustein, L. Deleterious effects by mosquitofish (Gambusia affinis) on the endangered fire salamander (Salamandra infraimmaculata). Anim. Conserv. 2008, 12, 29–37. [Google Scholar] [CrossRef]
- Schulse, C.D.; Semlitsch, R.D.; Trauth, K.M. Mosquitofish dominate amphibian and invertebrate community development in experimental wetlands. J. Appl. Ecol. 2013, 50, 1244–1256. [Google Scholar] [CrossRef]
- Peck, G.W.; Walton, W.R. Effects of mosquitofish (Gambusia affinis) and sestonic food abundance on the invertebrate community within a constructed treatment wetland. Freshw. Biol. 2008, 53, 2220–2233. [Google Scholar] [CrossRef]
- Preston, D.L.; Hedman, H.D.; Esfahani, E.R.; Pena, E.M.; Boland, C.E.; Lunde, K.B.; Johnson, P.J.T. Responses of a wetland ecosystem to the controlled introduction of invasive fish. Freshw. Biol. 2017, 62, 767–778. [Google Scholar] [CrossRef]
- Harmon, J.J.; Smith, G.R. Invasive fish (Gambusia affinis) as an ecological filter for macroinvertebrate colonization of experimental ponds. Freshw. Sci. 2020, 40, 151–161. [Google Scholar] [CrossRef]
- Otto, R.G. Temperature tolerance of the mosquitofish, Gambusia affinis (Baird and Girard). J. Fish Biol. 1973, 5, 575–585. [Google Scholar] [CrossRef]
- Pyke, G.H. A review of the biology of Gambusia affinis and G. holbrooki. Rev. Fish Biol. Fisher. 2005, 15, 339–365. [Google Scholar] [CrossRef]
- Mills, M.D.; Rader, R.B.; Belk, M.C. Complex interactions between native and invasive fish: The simultaneous effects of multiple negative interactions. Oecologia 2004, 141, 713–721. [Google Scholar] [CrossRef]
- Laha, M.; Mattingly, H.T. Ex Situ evaluation of impacts of invasive mosquitofish on the imperiled Barrens topminnow. Environ. Biol. Fish. 2007, 78, 1–11. [Google Scholar] [CrossRef]
- Sutton, T.M.; Zeiber, R.A.; Fisher, B.E. Agonistic behavioral interactions between introduced western mosquitofish and native topminnows. J. Freshw. Ecol. 2013, 28, 1–16. [Google Scholar] [CrossRef][Green Version]
- Boisclair, D.; Leggett, W.C. The importance of activity in bioenergetics models applied to actively foraging fishes. Can. J. Fish. Aquat. Sci. 1989, 46, 1859–1867. [Google Scholar] [CrossRef]
- Hulthén, K.; Chappman, B.B.; Nilsson, P.A.; Hansson, L.; Skov, C.; Brodersen, J.; Vinterstare, J.; Brönmark, C. A predation cost to bold fish in the wild. Sci. Rep. 2017, 7, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rennie, M.D.; Collins, N.C.; Shuter, B.J.; Rajotte, J.W.; Couture, P. A comparison of methods for estimating activity costs of wild fish populations: More active fish observed to grow slower. Can. J. Fish. Aquat. Sci. 2004, 62, 767–780. [Google Scholar] [CrossRef]
- Stasiak, R. Northern Redbelly Dace (Phoxinus eos): A Technical Conservation Assessment. [Online]. USDA Forest Service, Rocky Mountain Region. Available online: http://www.fs.fed.us/r2/projects/scp/assessments/northernredbellydace.pdf (accessed on 15 September 2021).
- Bestgen, K.R. Distribution and notes on the biology of Phoxinus eos (Cyprinidae) in Colorado. Southwest. Nat. 1989, 34, 225–231. [Google Scholar] [CrossRef]
- Stasiak, R.H. Phoxinus eos (Cope), northern redbelly dace. In Atlas of North American Freshwater Fishes; Lee, D.S., Gilbert, C.R., Hocutt, C.H., Jenkins, R.E., McAllister, D.E., Stauffer, J.R., Jr., Eds.; North Carolina State Museum of Natural History: Raleigh, NC, USA, 1980; p. 336. [Google Scholar]
- Carveth, C.J.; Widmer, A.M.; Bonar, S.A. Comparison of upper thermal tolerances of native and nonnative fish species in Arizona. T. Am. Fish. Soc. 2006, 135, 1433–1440. [Google Scholar] [CrossRef]
- Winkler, P. Thermal preference of Gambusia affinis affinis as determined under field and laboratory conditions. Copeia 1979, 60–64. [Google Scholar] [CrossRef]
- Brett, J.R. Energetic response of salmon to temperature. A study of some thermal relations in the physiology and freshwater ecology of sockeye salmon (Oncorhynchus nerka). Am. Zool. 1971, 11, 99–113. [Google Scholar] [CrossRef]
- Coutant, C.C. Thermal preference: When does an asset become a liability? Environ. Biol. Fish. 1987, 18, 161–172. [Google Scholar] [CrossRef]
- Beauchamp, D. Bioenergetic Ontogeny: Linking climate and mass-specific feeding to life-cycle growth and survival of salmon. Am. Fish. Soc. Symp. 2009, 70, 53–72. [Google Scholar]
- Pankhurst, N.W.; Munday, P.L. Effects of climate change on fish reproduction and early life history stages. Mar. Freshw. Res. 2011, 62, 1015–1026. [Google Scholar] [CrossRef]
- Colchen, T.; Teletchea, F.; Fontaine, P.; Pasquet, A. Temperature modifies activity, inter-individual relationships and group structure in a fish. Curr. Zool. 2017, 63, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Kessler, K.; Lampert, W. Fitness optimization of Daphnia in a trade-off between food and temperature. Oecologia 2004, 140, 381–387. [Google Scholar] [CrossRef]
- Morris, W. Adaptation and habitat selection in the eco-evolutionary process. Proc. R. Soc. B. 2011, 278, 2401–2411. [Google Scholar] [CrossRef]
- Brewitt, K.S.; Danner, E.M.; Moore, J.W. Hot eats and cool creeks. Hot eats and cool creeks: Juvenile Pacific salmonids use mainstem prey while in thermal refuges. Can. J. Fish. Aquat. Sci. 2017, 74, 1588–1602. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Rahel, F.J.; Novinger, D.C.; Gerow, K.G. Temperature mediation of competitive interactions among three fish species that replace each other along longitudinal stream gradients. Can. J. Fish. Aquat. Sci. 1998, 55, 1894–1901. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Nakano, S. Condition-specific competition: Implications for the altitudinal distribution of stream fishes. Ecology 2000, 81, 2027–2039. [Google Scholar] [CrossRef]
- Yamada, T.; Koizumi, I.; Urabe, H.; Nakamura, F. Temperature-dependent swimming performance differs by species: Implications for condition-specific competition between stream salmonids. Zool. Sci. 2020, 37, 429–433. [Google Scholar] [CrossRef]
- Carmona-Catot, G.; Magellan, G.K.; García-Berthou, E. Temperature specific competition between invasive Mosquitofish and endangered Cyprinodontid fish. PLoS ONE 2013, 8, e54734. [Google Scholar] [CrossRef] [PubMed]
- Majerova, M.; Neilson, B.T.; Roper, B.B. Beaver dam influences on streamflow hydraulic properties and thermal regimes. Sci. Total Environ. 2020, 718, 134853. [Google Scholar] [CrossRef] [PubMed]
- Nijman, V.; Heuts, B.A. Effect of environmental enrichment upon resource holding power in fish in prior residence situations. Behav. Process. 2000, 49, 77–83. [Google Scholar] [CrossRef]
- Gilmour, K.M.; DiBattista, J.D.; Thomas, J.B. Physiological causes and consequences of social status in salmonid fish. Integr. Comp. Biol. 2005, 45, 263–273. [Google Scholar] [CrossRef]
- Kua, Z.X.; Hamilton, I.M.; McLaughlin, A.L.; Brodnik, R.M.; Keitzer, S.C.; Gilliland, J.; Hoskins, E.A.; Ludsin, S.A. Water warming increases aggression in a tropical fish. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Kelly, N.I.; Burness, G.; McDermid, J.L.; Wilson, C.C. Ice age fish in a warming world: Minimal variation in thermal acclimation capacity among lake trout (Salvelinus namaycush) populations. Conserv. Physiol. 2014, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Myrick, C.A.; Kurtis, F.D.; Cech, J.J. An annular chamber for aquatic animal preference studies. Trans. Am. Fish. Soc. 2004, 133, 427–433. [Google Scholar] [CrossRef]
- Caballero, C.; Castro, J.J. Effect of residence and size asymmetries upon the agonistic interactions between juvenile white-seabream (Diplodus sargus cadenati de la Paz, Bauchot and Daget, 1974). Aggress. Behav. 1999, 25, 297–303. [Google Scholar] [CrossRef]
- Domínguez-Castanedo, O. Agonistic interactions with asymmetric body size in two adult-age groups of the annual killifish Millerichthys robustus (Miller & Hubbs, 1974). J. Fish Biol. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Slivko, V.M.; Zhokhov, A.E.; Gopko, M.V.; Mikheev, V.N. Agonistic behavior of young perch Perca fluviatilis: The effects of fish size and macroparasite load. J. Ichthyol. 2021, 61, 476–481. [Google Scholar] [CrossRef]
- Plummer, M. JAGS Version 4.3.0 User Manual [online]. Available online: http://mcmc-jags.sourceforge.net (accessed on 12 May 2019).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 29 September 2021).
- Kellner, K. jagsUI: A Wrapper around ‘rjags’ to Streamline “JAGS” Analyses; R Package Version 1.5.2. Available online: https://cran.r-project.org/web/packages/jagsUI/index.html (accessed on 30 October 2021).
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Stauffer, J.R., Jr.; Melisky, E.L.; Hocutt, C.H. Temperature preference of the northern redbelly dace Phoxinus eos (Cope). Arch. Hydrobiol. 1980, 90, 121–126. [Google Scholar]
- Kellogg, R.L.; Gift, J.J. Relationships between optimum temperature for growth and preferred temperatures for the young of four species. Trans. Am. Fish. Soc. 1983, 112, 424–430. [Google Scholar] [CrossRef]
- Boltaña, S.; Sanhueza, N.; Aguilar, A.; Gallardo-Escarate, C.; Arriagada, G.; Valdes, J.A.; Soto, D.; Quiñones, R.A. Influences of thermal environment on fish growth. Ecol. Evol. 2017, 7, 6814–6825. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.; Wieser, W. Partitioning of energy in fish: Can reduction of swimming activity compensate for the cost of production? J. Exp. Biol. 1983, 107, 141–146. [Google Scholar] [CrossRef]
- Trudel, M.; Boisclair, D. Estimation of fish activity costs using underwater video cameras. J. Fish Biol. 1996, 48, 40–53. [Google Scholar] [CrossRef]
- Meffe, G.K. Predation and species replacement in American Southwestern Fishes: A case study. Southwest. Nat. 1985, 30, 173–187. [Google Scholar] [CrossRef]
- Deverill, J.I.; Adams, C.E.; Bean, C.W. Prior residence, aggression and territory acquisition in hatchery-reared and wild brown trout. J. Fish Biol. 1999, 55, 868–875. [Google Scholar] [CrossRef]
- Mesa, M.G. Variation in feeding, aggression, and position choice between hatchery and wild cutthroat trout in an artificial stream. Trans. Am. Fish. Soc. 1991, 120, 723–727. [Google Scholar] [CrossRef]
- Harwood, A.J.; Griffiths, S.W.; Metcalfe, N.B.; Armstrong, J.D. The relative influence of prior residency and dominance on the early feeding behavior of juvenile Atlantic salmon. Anim. Behav. 2003, 65, 1141–1149. [Google Scholar] [CrossRef][Green Version]
- Nijman, V.; Heuts, B.A. Aggression and dominance in cichlids in resident-intruder tests: The role of environmental enrichment. Neotrop. Ichthyol. 2011, 9, 543–545. [Google Scholar] [CrossRef][Green Version]
- Kvingedal, E.; Einum, S. Prior residency advantage for Atlantic salmon in the wild: Effects of habitat quality. Behav. Ecol. Sociobiol. 2011, 65, 1295–1303. [Google Scholar] [CrossRef]
- Conte, F.S. Stress and the welfare of cultured fish. Appl. Anim. Behav. Sci. 2004, 86, 205–223. [Google Scholar] [CrossRef]
- Harper, C.; Wolf, J.C. Morphological effects of the stress response in fish. ILAR J. 2009, 50, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Monirian, J.; Sutphin, Z.; Myrick, C. Effects of holding temperature and handling stress on the upper thermal tolerance of threadfin shad Dorosoma petenense. J. Fish Biol. 2010, 76, 1329–1342. [Google Scholar] [CrossRef]
- Schreck, C.B.; Olla, B.L.; Davis, M.W. Behavior responses to stress. In Fish Stress and Health in Aquaculture, Society for Experimental Biology Seminar Series 62; Iwama, G.K., Pickering, A.D., Sumpter, J.P., Schreck, C.B., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 145–170. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciepiela, L.R.; Fitzpatrick, R.M.; Lewis, S.T.; Kanno, Y. Behavioral Interactions between a Native and an Invasive Fish Species in a Thermally Heterogeneous Experimental Chamber. Fishes 2021, 6, 75. https://doi.org/10.3390/fishes6040075
Ciepiela LR, Fitzpatrick RM, Lewis ST, Kanno Y. Behavioral Interactions between a Native and an Invasive Fish Species in a Thermally Heterogeneous Experimental Chamber. Fishes. 2021; 6(4):75. https://doi.org/10.3390/fishes6040075
Chicago/Turabian StyleCiepiela, Lindsy R., Ryan M. Fitzpatrick, Samuel T. Lewis, and Yoichiro Kanno. 2021. "Behavioral Interactions between a Native and an Invasive Fish Species in a Thermally Heterogeneous Experimental Chamber" Fishes 6, no. 4: 75. https://doi.org/10.3390/fishes6040075
APA StyleCiepiela, L. R., Fitzpatrick, R. M., Lewis, S. T., & Kanno, Y. (2021). Behavioral Interactions between a Native and an Invasive Fish Species in a Thermally Heterogeneous Experimental Chamber. Fishes, 6(4), 75. https://doi.org/10.3390/fishes6040075