
Hidden Giants: The Story of Bolbometopon muricatum at Ningaloo Reef

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
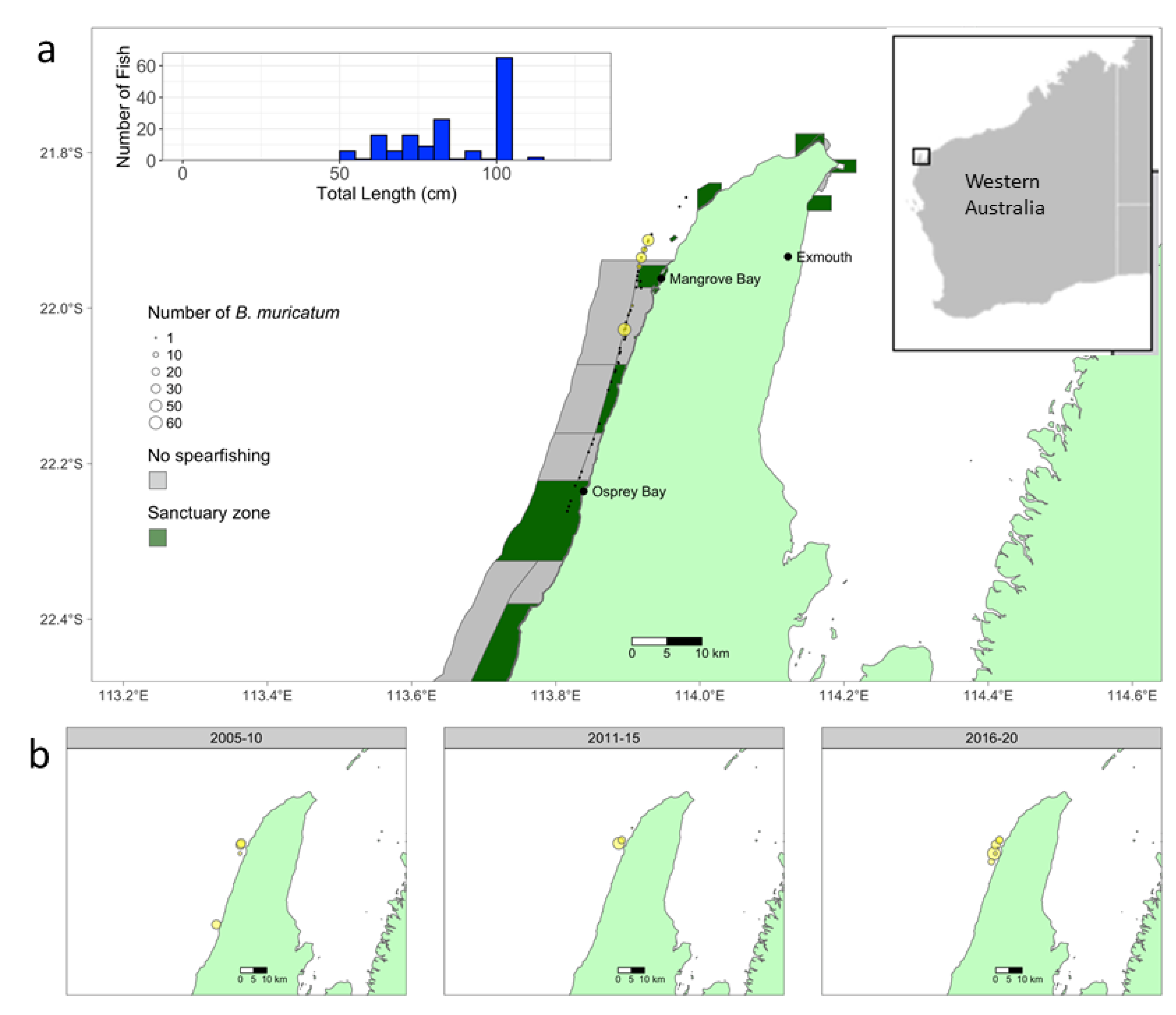
2.1. Study Location
2.2. Historic Reef Fish Surveys (1987–2019)
2.3. Qualitative Observations of B. muricatum at Northern Ningaloo (2006–2020) Solicited from Expert Witnesses
2.4. Observations of B. muricatum at Northern Ningaloo (2021) from Reef Crest Timed UVC Surveys
3. Results
3.1. Historic Reef Fish Surveys (1987–2019)
3.2. Qualitative Observations of B. muricatum at Northern Ningaloo (2006–2020) Solicited from Expert Witnesses
3.3. Observations of B. muricatum at Northern Ningaloo (2021) from Reef Crest Timed UVC Surveys
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as Ecosystem Engineers. In Ecosystem Management: Selected Readings; Springer: New York, NY, USA, 1996; pp. 130–147. [Google Scholar] [CrossRef]
- Coleman, F.C.; Williams, S.L. Overexploiting marine ecosystem engineers: Potential consequences for biodiversity. Trends Ecol. Evol. 2002, 17, 40–44. [Google Scholar] [CrossRef]
- Pogoda, B.; Brown, J.; Hancock, B.; Preston, J.; Pouvreau, S.; Kamermans, P.; Sanderson, W.; von Nordheim, H. The Native Oyster Restoration Alliance (NORA) and the Berlin Oyster Recommendation: Bringing back a key ecosystem engineer by developing and supporting best practice in Europe. Aquat. Living Resour. 2019, 32, 13. [Google Scholar] [CrossRef] [Green Version]
- Beger, M.; Babcock, R.; Booth, D.J.; Bucher, D.; Condie, S.A.; Creese, B.; Cvitanovic, C.; Dalton, S.J.; Harrison, P.; Hoey, A. Research challenges to improve the management and conservation of subtropical reefs to tackle climate change threats: (Findings of a workshop conducted in Coffs Harbour, Australia on 13 September 2010). Ecol. Manag. Restor. 2011, 12, e7–e10. [Google Scholar] [CrossRef]
- Darling, E.S.; McClanahan, T.R.; Maina, J.; Gurney, G.G.; Graham, N.A.J.; Januchowski-Hartley, F.; Cinner, J.E.; Mora, C.; Hicks, C.C.; Maire, E.; et al. Social–environmental drivers inform strategic management of coral reefs in the Anthropocene. Nat. Ecol. Evol. 2019, 3, 1341–1350. [Google Scholar] [CrossRef]
- Jenkins, A.P.; Jupiter, S.D.; Qauqau, I.; Atherton, J. The importance of ecosystem-based management for conserving aquatic migratory pathways on tropical high islands: A case study from Fiji. Aquat. Conserv. -Mar. Freshw. Ecosyst. 2010, 20, 224–238. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Graham, N.A.; MacNeil, M.A.; Muthiga, N.A.; Cinner, J.E.; Bruggemann, J.H.; Wilson, S.K. Critical thresholds and tangible targets for ecosystem-based management of coral reef fisheries. Proc. Natl. Acad. Sci. USA 2011, 108, 17230–17233. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, R.J.; Almany, G.R.; Stevens, D.; Bode, M.; Pita, J.; Peterson, N.A.; Choat, J.H. Hyperstability masks declines in bumphead parrotfish (Bolbometopon muricatum) populations. Coral Reefs 2016, 35, 751–763. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hoey, A.S.; Choat, J.H. Limited functional redundancy in high diversity systems: Resilience and ecosystem function on coral reefs. Ecol. Lett. 2003, 6, 281–285. [Google Scholar] [CrossRef]
- Hoey, A.; Bellwood, D. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2008, 27, 37–47. [Google Scholar] [CrossRef]
- Bonaldo, R.M.; Hoey, A.S.; Bellwood, D.R. The ecosystem roles of parrotfishes on tropical reefs. Oceanogr. Mar. Biol. Annu. Rev. 2014, 52, 81–132. [Google Scholar]
- Bellwood, D.R.; Hoey, A.S.; Hughes, T.P. Human activity selectively impacts the ecosystem roles of parrotfishes on coral reefs. Proc. R. Soc. B Biol. Sci. 2012, 279, 1621–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choat, J.H.; Robertson, D.R. Age-based studies. Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem; Academic Press: San Diego, CA, USA, 2002; pp. 57–80. [Google Scholar]
- Choat, J.; Axe, L.; Lou, D. Growth and longevity in fishes of the family Scaridae. Mar. Ecol. Prog. Ser. 1996, 145, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Choat, J.H.; Klanten, O.S.; Van Herwerden, L.; Robertson, D.R.; Clements, K.D. Patterns and processes in the evolutionary history of parrotfishes (Family Labridae). Biol. J. Linn. Soc. 2012, 107, 529–557. [Google Scholar] [CrossRef] [Green Version]
- Dulvy, N.K.; Polunin, N.V. Using informal knowledge to infer human-induced rarity of a conspicuous reef fish. Anim. Conserv. 2004, 7, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Johannes, R.E. Working with fishermen to improve coastal tropical fisheries and resource management. Bull. Mar. Sci. 1981, 31, 673–680. [Google Scholar]
- Donaldson, T.J.; Dulvy, N.K. Threatened fishes of the world: Bolbometopon muricatum (Valenciennes 1840)(Scaridae). Environ. Biol. Fishes 2004, 70, 373. [Google Scholar] [CrossRef]
- Kobayashi, D.R.; Friedlander, A.M.; Grimes, C.B.; Nichols, R.S.; Zgliczynski, B. Bumphead parrotfish (Bolbometopon muricatum) status review. In NOAA Technical Memorandum NMFS-PIFSC; Pacific Islands Fisheries Science Center: Honolulu, HI, USA, 2011. [Google Scholar]
- Polloi, K.; Golbuu, Y.; Merep, G.; Koshiba, S.; Friedlander, A.; Koike, H. An assessment of Maml and Kemedukl in Palau and management recommendations. A Report to The Nature Conservancy Micronesia Program Technical Report No. 14-07. In The Nature Conservancy Micronesia Program Technical Report; Palau International Coral Reef Center: Koror, Palau, 2014; p. 37. [Google Scholar]
- Environment Protection and Biodiversity Conservation Act 1999. No. 91, 1999, Volume 1: Sections 1–266; The Office of the Parliamentary Counsel: Canberra, Australia, 1999.
- Management plan for the Ningaloo Marine Park and Muiron Islands Marine Management Area 2005–2015. In Perth, Western Australia: Conservation and Land Management and Marine Parks and Reserves Authority; Government of Western Australia: Perth, Australia, 2005; p. 111.
- Deloitte Access Economics Pty Ltd. Economic contribution of Ningaloo: One of Australia’s best kept secrets. In Report Commissioned by DBCA; DBCA: Perth, Australia, 2020; p. 58. [Google Scholar]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Jenkins, D.; Morrison, S. Fishes of Ningaloo; Blue Ocean Publications: Exmouth, Australia, 2018. [Google Scholar]
- Vanderklift, M.A.; Babcock, R.C.; Barnes, P.B.; Cresswell, A.K.; Feng, M.; Haywood, M.D.; Holmes, T.H.; Lavery, P.S.; Pillans, R.D.; Smallwood, C.B. The Oceanography and Marine Ecology of Ningaloo, A World Heritage Area. In Oceanography and Marine Biology: An Annual Review; Hawkins, S.J., Allcock, A.L., Bates, A.E., Evans, A.J., Firth, L.B., McQuaid, C.D., Russel, B.D., Smith, I.P., Swearer, S.E., Todd, P.A., Eds.; Taylor & Francis: London, UK, 2020. [Google Scholar]
- Thomson, D.P.; Babcock, R.C.; Haywood, M.D.; Vanderklift, M.A.; Pillans, R.D.; Bessey, C.; Cresswell, A.; Orr, M.; Boschetti, F.; Wilson, S.K. Zone specific trends in coral cover, genera and growth-forms in the World-Heritage listed Ningaloo Reef. Mar. Environ. Res. 2020, 160, 105020. [Google Scholar] [CrossRef] [PubMed]
- Cresswell, A.; Langlois, T.; Wilson, S.; Claudet, J.; Thomson, D.P.; Renton, M.; Fulton, C.J.; Fisher, R.; Vanderklift, M.; Babcock, R. Disentangling the response of fishes to recreational fishing over 30 years within a fringing coral reef reserve network. Biol. Conserv. 2019, 237, 514–524. [Google Scholar] [CrossRef]
- Langlois, T.J.; Harvey, E.S.; Fitzpatrick, B.; Meeuwig, J.J.; Shedrawi, G.; Watson, D.L. Cost-efficient sampling of fish assemblages: Comparison of baited video stations and diver video transects. Aquat. Biol. 2010, 9, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Murphy, H.M.; Jenkins, G.P. Observational methods used in marine spatial monitoring of fishes and associated habitats: A review. Mar. Freshw. Res. 2010, 61, 236–252. [Google Scholar] [CrossRef]
- Perry, C.T.; Alvarez-Filip, L.; Graham, N.A.; Mumby, P.J.; Wilson, S.K.; Kench, P.S.; Manzello, D.P.; Morgan, K.M.; Slangen, A.B.; Thomson, D.P. Loss of coral reef growth capacity to track future increases in sea level. Nature 2018, 558, 396–400. [Google Scholar] [CrossRef]
- Hamilton, R.; Adams, S.; Choat, J. Sexual development and reproductive demography of the green humphead parrotfish (Bolbometopon muricatum) in the Solomon Islands. Coral Reefs 2008, 27, 153–163. [Google Scholar] [CrossRef]
- Bellwood, D.; Choat, J. Dangerous demographics: The lack of juvenile humphead parrotfishes Bolbometopon muricatum on the Great Barrier Reef. Coral Reefs 2011, 30, 549–554. [Google Scholar] [CrossRef]
- Roff, G.; Doropoulos, C.; Mereb, G.; Mumby, P.J. Mass spawning aggregation of the giant bumphead parrotfish Bolbometopon muricatum. J. Fish Biol. 2017, 91, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.; Meeuwig, J.; Feng, M.; Harvey, E.; Lam, V.; Langlois, T.; Slawinski, D.; Sun, C.; Pauly, D. Climate-change induced tropicalisation of marine communities in Western Australia. Mar. Freshw. Res. 2012, 63, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Domeier, M.L.; Colin, P.L. Tropical reef fish spawning aggregations: Defined and reviewed. Bull. Mar. Sci. 1997, 60, 698–726. [Google Scholar]
- MacKenzie, D.I.; Royle, J.A.; Brown, J.A.; Nichols, J.D.; Thompson, W. Occupancy estimation and modeling for rare and elusive populations. In Sampling Rare or Elusive Species: Concepts, Designs, and Techniques for Estimating Population Parameters; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; pp. 149–171. [Google Scholar]
- Colvocoresses, J.; Acosta, A. A large-scale field comparison of strip transect and stationary point count methods for conducting length-based underwater visual surveys of reef fish populations. Fish. Res. 2007, 85, 130–141. [Google Scholar] [CrossRef]

{kind=link}
| Reef Zone | B. muricatum | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Survey Method | Years | Lagoon | Back Reef | Reef Flat | Reef Crest | Reef Slope | Total Time/Area | Number | DENSITY Ind/ha |
| BRUV (hours) | 2000–2015 | 351.5 | 34.0 | 66.0 | 0 | 426.0 | 877.5 h | 0 | 0 |
| DOV (ha) | 2006–2016 | 13.6 | 4.0 | 1.0 | 0 | 3.0 | 21.6 ha | 0 | 0 |
| UVC (ha) | 1987–2019 | 49.9 | 30.2 | 28.6 | 0 | 35.7 | 144.4 ha | 0 | 0 |
| This study Timed UVC swim (ha) | 2021 | 0.0 | 0.0 | 0.0 | 63.2 | 0 | 63.2 ha | 155 | 2.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomson, D.P.; Cresswell, A.K.; Doropoulos, C.; Haywood, M.D.E.; Orr, M.; Hoey, A.S. Hidden Giants: The Story of Bolbometopon muricatum at Ningaloo Reef. Fishes 2021, 6, 73. https://doi.org/10.3390/fishes6040073
Thomson DP, Cresswell AK, Doropoulos C, Haywood MDE, Orr M, Hoey AS. Hidden Giants: The Story of Bolbometopon muricatum at Ningaloo Reef. Fishes. 2021; 6(4):73. https://doi.org/10.3390/fishes6040073
Chicago/Turabian StyleThomson, Damian P., Anna K. Cresswell, Christopher Doropoulos, Michael D. E. Haywood, Melanie Orr, and Andrew S. Hoey. 2021. "Hidden Giants: The Story of Bolbometopon muricatum at Ningaloo Reef" Fishes 6, no. 4: 73. https://doi.org/10.3390/fishes6040073
APA StyleThomson, D. P., Cresswell, A. K., Doropoulos, C., Haywood, M. D. E., Orr, M., & Hoey, A. S. (2021). Hidden Giants: The Story of Bolbometopon muricatum at Ningaloo Reef. Fishes, 6(4), 73. https://doi.org/10.3390/fishes6040073