Survival, Growth, and Development in the Early Stages of the Tropical Gar Atractosteus tropicus: Developmental Critical Windows and the Influence of Temperature, Salinity, and Oxygen Availability

 , ,
, , 

Abstract
1. Introduction
2. Results
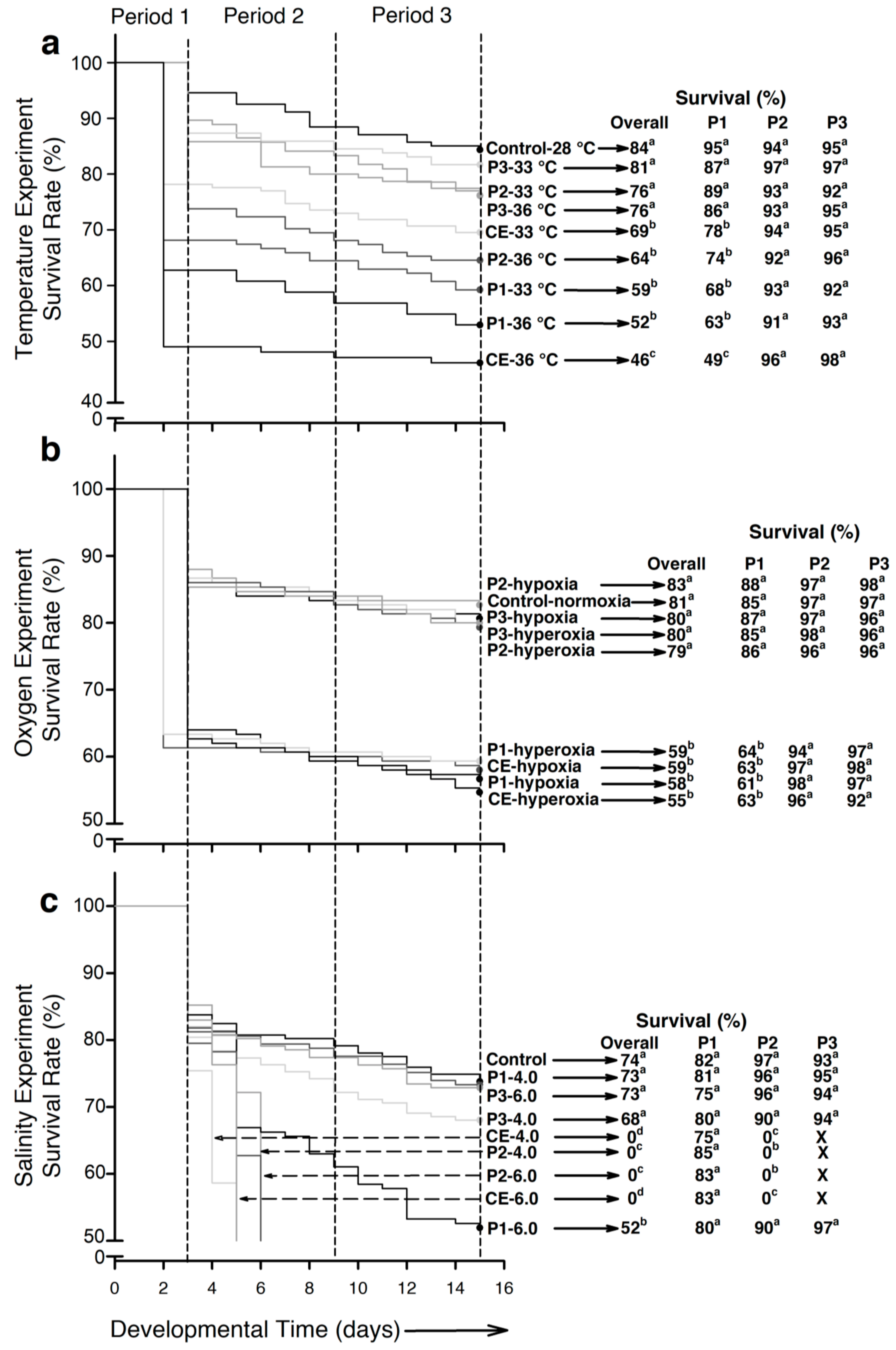
2.1. Survival as a Function of Stressor
2.1.1. Temperature
2.1.2. Air Saturation Level
2.1.3. Salinity
2.2. Survival as a Function of Development Stage
2.2.1. Period 1 (P1)—Fertilization to Hatch
- Temperature. Control larvae exhibited the highest survival (95%; p < 0.001) followed by larvae from experimental groups P2-33 °C, P3-33 °C, P2-36 °C, and P3-36 °C (74% to 89%). Lower survival (p < 0.001) was registered for fish from experimental groups P1-33 °C and P1-36 °C at 68% and 63%, respectively. The lowest survival (p < 0.001) occurred in larvae from experimental group CE-36 °C (49%; Figure 1a).
- Air saturation. Control larvae and those from experimental groups P2-hypoxia, P3-hypoxia, P2-hypeoxia, and P3-hyperoxia exhibited survival of 85–88%, which significantly differed (p < 0.001) from experimental groups P1-hypoxia, P1-hyperoxia, CE-hypoxia, and CE-hyperoxia (61–64%; Figure 1b).
- Salinity. No significant differences occurred in survival (p > 0.05), which ranged from 75% to 85% in the different groups (Figure 1c).
2.2.2. Period 2 (P2)—From 1 to 6 dph
- Temperature and air saturation. No significant differences were found (p > 0.05). Survival was >90% in both experiments (Figure 1a,b).
- Salinity. The highest salinity-induced mortality events occurred in this intermediate period (p < 0.001). By 2 days post hatch (dph), all larvae from experimental groups CE-4.0 and CE-6.0 had died. By 3 dph, the fish from experimental groups P2-4.0 and P2-6.0 had also died. The rest of the experimental groups showed no significant differences (p > 0.05; survival 90% to 97%; Figure 1c).
2.2.3. Period 3 (P3)—From 7 to 12 dph
- Developmental period 3 was the most resilient to stressors. No significant differences were found between experimental groups in any experiment (p > 0.05): temperature 91–98% survival, hypoxia-hyperoxia 92–98%, and salinity 93–97% (Figure 1a–c).
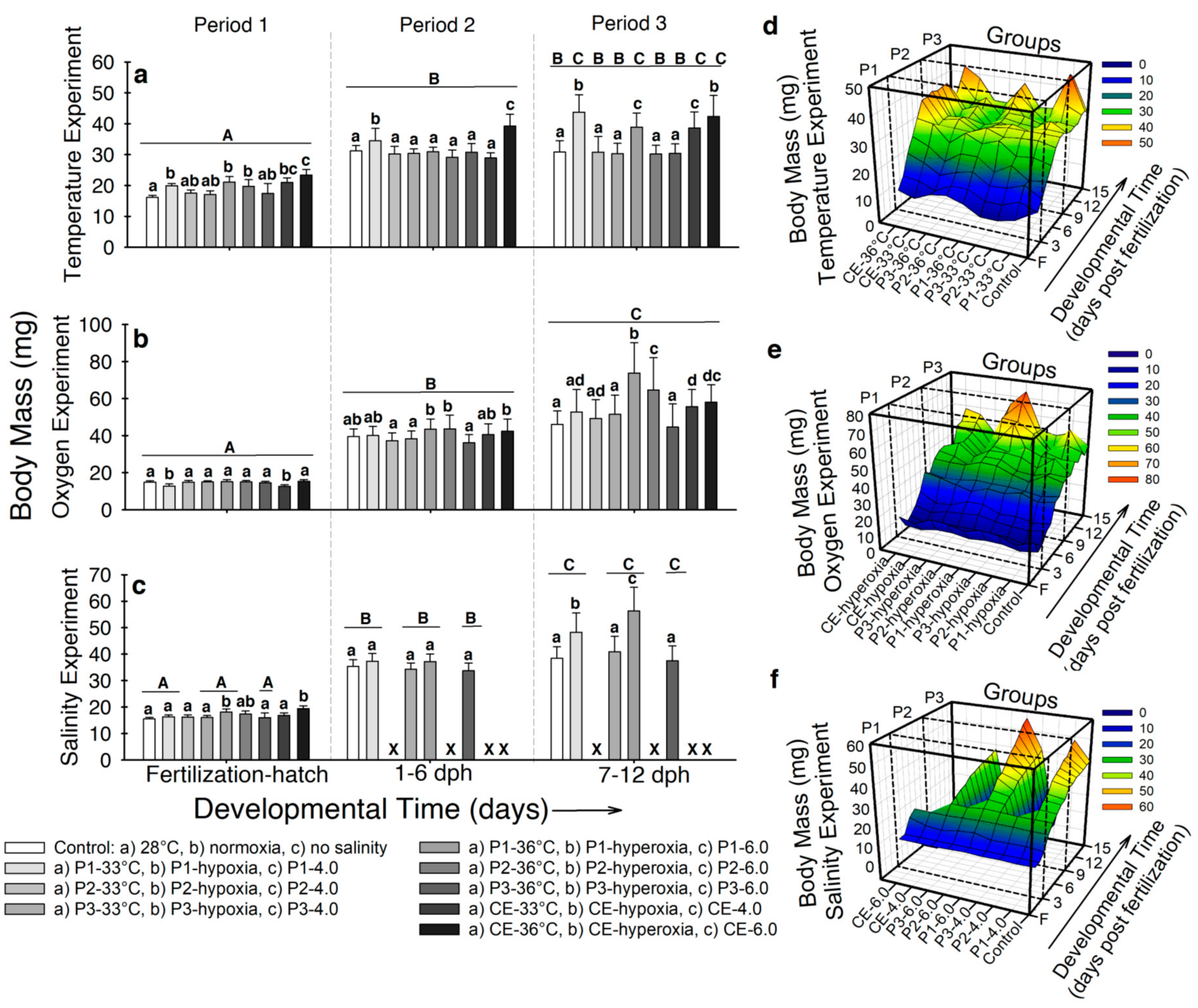
2.3. Body Morphology and Growth as a Function of Stressors and Stage of Exposure
2.3.1. Body Mass (BM)
- Temperature. At the end of developmental period 1, the control larvae displayed the lowest body mass (16.1 ± 0.6 mg), while larvae hatched in 33 °C and 36 °C exhibited the highest (BM ~19–23 mg; p < 0.001; Figure 2a). For developmental period 2, the highest BM was registered for larvae from experimental groups P1-33 °C (34.9 ± 3.9 mg) and CE-36 °C (39.2 ± 3,7 mg; p < 0.001; Figure 2a). By the end of developmental period 3, larvae from experimental groups P1-33 °C, P1-36 °C, CE-33 °C, and CE-33° showed higher BM (~38–43 mg) compared to the rest of the experimental groups (BM ~30 mg; p < 0.001; Figure 2a). Figure 2d illustrates how BM increased immediately after hatching in all the experimental groups, especially in CE-33 °C and CE-36 °C. At the beginning of developmental period 3, a slight increase in BM occurred in larvae from experimental groups P3-33 °C and P3-36 °C. However, after 2–3 days BM decreased to control values. The groups that exhibited the highest BM at the end of the experiment started increasing BM at ~9 dph.
- Air saturation. Larvae from experimental groups P1-hypoxia and CE-hypoxia displayed the lowest BM (~12 mg) at hatching (p < 0.001; Figure 2b). By developmental period 2, larvae from experimental groups P1-hyperoxia, P2-hyperoxia, and CE-hyperoxia exhibited the highest BM (~42 mg; p < 0.001; Figure 2b). By developmental period 3, fish from the control and P3-hyperoxia exhibited the lowest BM (~45 mg); conversely, individuals from experimental groups P1-hyperoxia and P2-hyperoxia showed the highest (BM ~64–73 mg; p < 0.001; Figure 2b). Figure 2e shows high variation in BM for all groups since the beginning of developmental period 2. Fish from experimental groups with the highest BM at the end of the experiment started showing differences at ~9 dph (p < 0.001). Body mass of larvae from experimental group P3-hyperoxia showed little variation through developmental period 3.
- Salinity. In developmental period 1, fish from experimental groups P1-6.0 and CE-6.0 showed the highest BM (18.12 ± 1.3 mg and 19.4 ± 1.6 mg respectively; p < 0.001; Figure 2c). At the end of developmental period 2, no significant differences in BM occurred between any of the five surviving groups (BM ~32–37 mg; p > 0.05). By developmental period 3, larvae from experimental groups P1-4.0 and P1-6.0 exhibited higher BM (48.2 ± 4.1 mg and 56.3 ± 6.1 mg, respectively) than the control and the rest of the groups (~37–40 mg; p < 0.001; Figure 2c). Figure 2f shows the BM to increase in P1-4.0 and P1-6.0 at ~7 dph and continued by the end of the experiment, while fish from the control and experimental groups P3-4.0 and P3-6.0 remained constant.
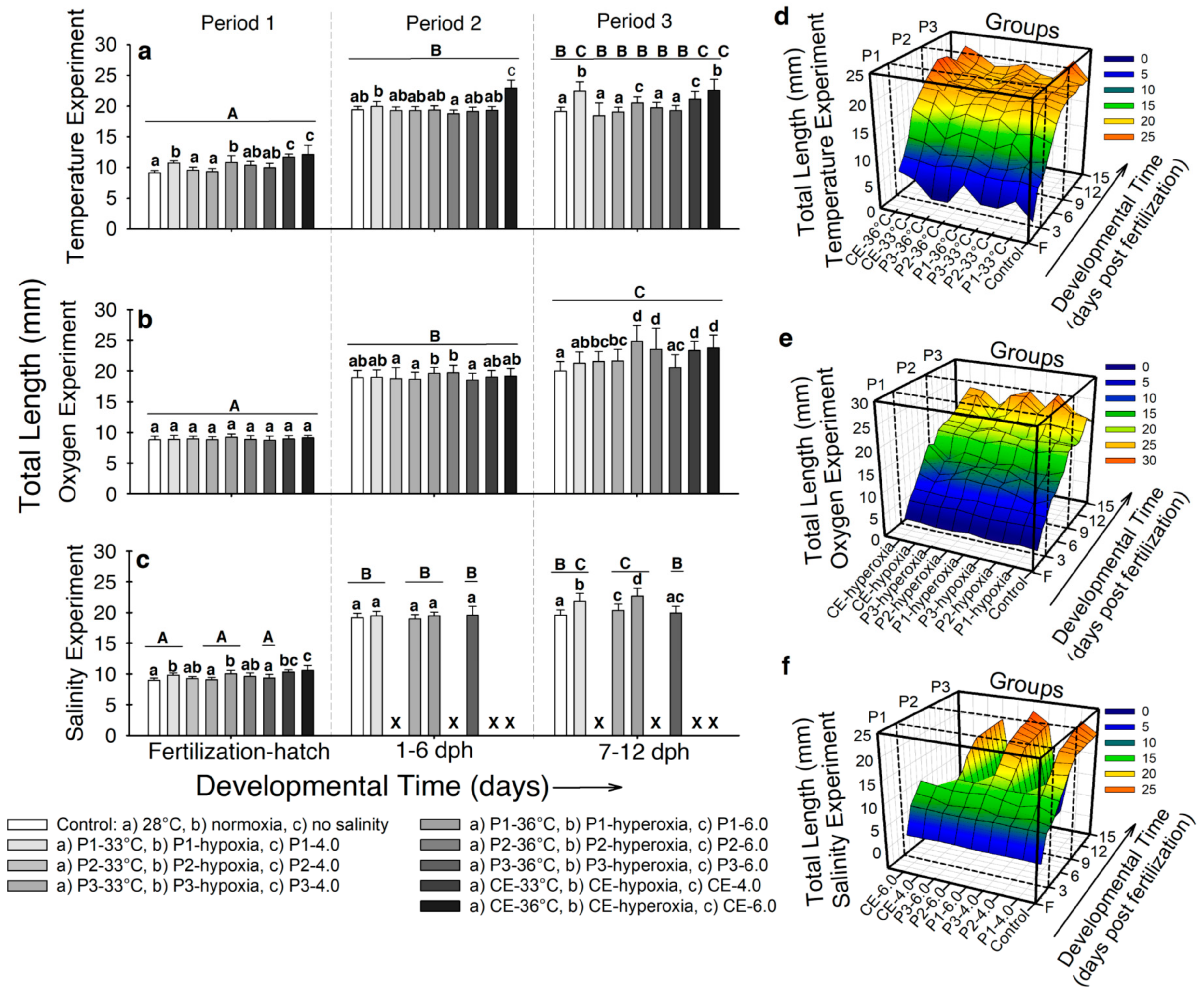
2.3.2. Total Length (LT)
- Temperature. Larvae exposed to either 33 °C or 36 °C during incubation exhibited higher total length (LT) (~10–12 mm) compared to the controls (9.1 ± 0.36 mm; p < 0.001; Figure 3a). For developmental period 2, the highest LT was observed in larvae from experimental groups CE-36 °C (22.9 ± 1.3 mm; p < 0.001; Figure 3a). By developmental period 3, larvae from experimental groups P1-33 °C, P1-36 °C, CE-33 °C, and CE-36 °C showed higher LT (~20–22 mm) than the control larvae (19.1 ± 0.7 mm; p < 0.001; Figure 3a). The interaction of all variables during the complete experiment is presented in Figure 3d. Total length from fish from experimental groups P1-33 °C and CE-36 °C significantly increased from incubation through 12 dph, compared to the rest of the groups (p < 0.001). However, larvae from experimental group P1-36 °C showed increased LT by hatching, but during developmental period 2, the values were closer to the control larvae (~19 mm).
- Air saturation. No significant differences were observed in fish LT by developmental period 1 (LT ~9 mm; p > 0.05; Figure 3b). In developmental period 2, larvae from experimental groups P1-hyperoxia and P2-hyperoxia showed higher LT (19.6 ± 0.9 mm; 19.7 ± 1.2 mm, respectively) compared to the rest (LT ~18.5 mm; p < 0.001; Figure 3b). For developmental period 3, LT was higher in larvae from experimental groups P1-hyperoxia (24.8 ± 1.6 mm), P2-hyperoxia (23.6 ± 2.3 mm), CE-hyperoxia (23.8 ± 2.2 mm), and CE-hypoxia (23.3 ± 1.3 mm; p < 0.001) compared to the control (20 ± 1.8 mm; Figure 3b). Figure 3e shows the interaction of development, LT, and time of exposure to hypoxia and hyperoxia throughout the experiment. All treatments showed a similar tendency in the LT increase from incubation to ~9 dph. However, at this point, LT of fishes from experimental group P1-hyperoxia, P2-hyperoxia, CE-hyperoxia, and CE-hypoxia showed major increase in LT than the fish in the rest of the groups.
- Salinity. By the end of developmental period 1, larvae from experimental groups P1-4.0, CE-4.0, P1-6.0, and CE-6.0 showed higher LT (~9.6-10 mm) compared to fish from the control and experimental groups P2-4.0, P3-4.0, P2-6.0, and P3-6.0 (LT ~9.2 mm; p < 0.001; Figure 3c). No significant differences in LT were observed at developmental period 2 between experimental groups (LT ~19.2 mm; p > 0.05; Figure 3c). At the end of the experiment, only larvae from experimental groups P1-4.0 and P1-6.0 showed significantly higher LT values (~20.3–21-8 mm) compared to the control and larvae from experimental groups P3-4.0 and P3-6.0 (~19.7 mm; p < 0.001; Figure 3c). The interaction of LT, development, and the groups during the whole experiment is presented in Figure 3f. From incubation to ~9 dph, all groups showed similar patterns of increase LT. Right after this point, fish from experimental groups P1-4.0 and P1-6.0 showed higher values of LT (~21 mm) than the control (19.2 ± 1.1 mm; p < 0.001), which continued through the experiment.
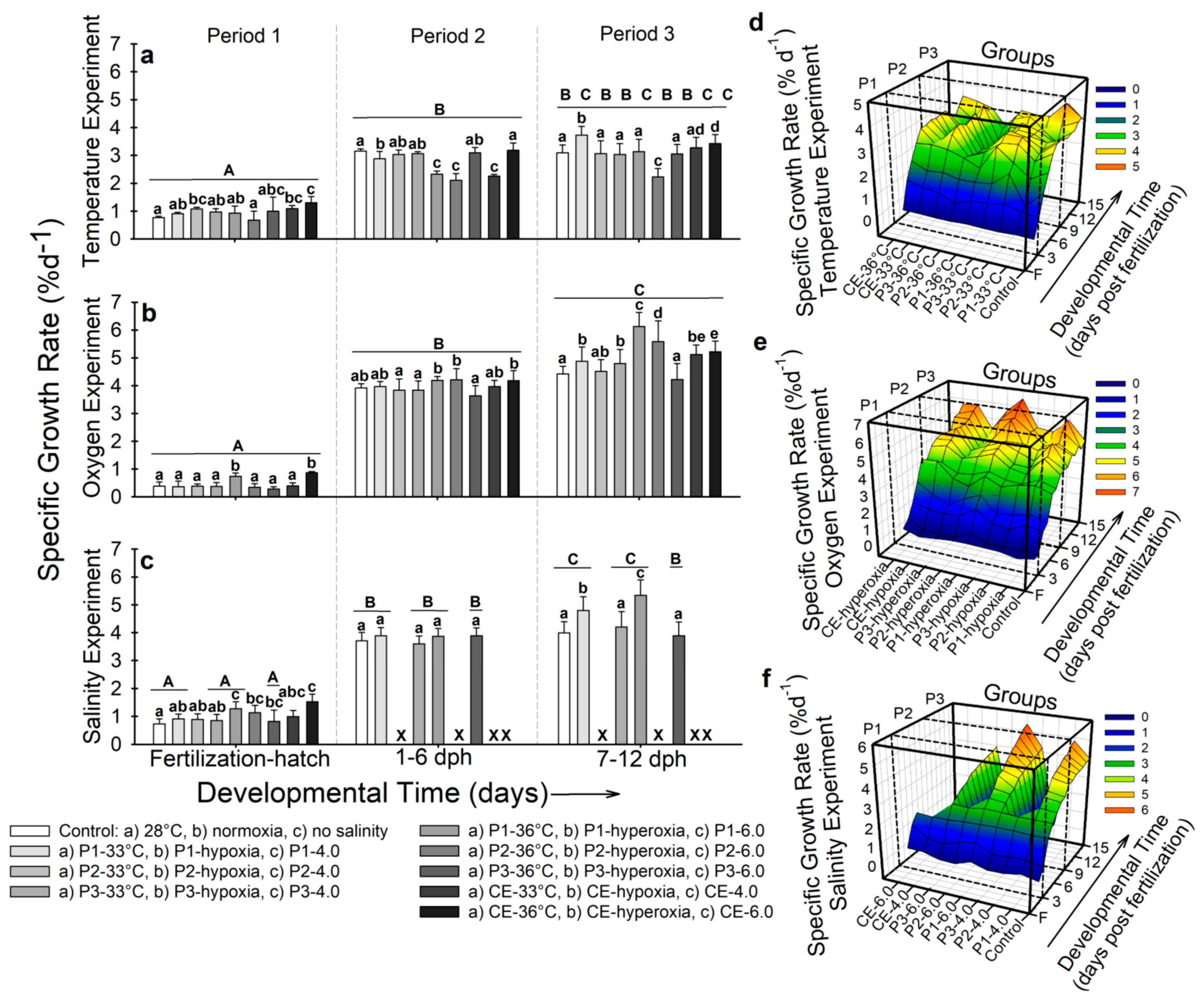
2.3.3. Specific Growth Rate (SGR)
- Temperature. During developmental period 1, fish from the control and experimental group P2-33 °C showed the lowest specific growth rate (SGR) (~0.7% d−1), while the highest SGR was observed in larvae from experimental group CE-36 °C (1.3 ± 0.2% d−1; p < 0.001; Figure 4a).
- Air saturation. At the end of developmental period 1, fish from experimental groups P1-hyperoxia and CE-hyperoxia showed the highest SGR (0.74–0.86% d−1; p < 0.001; Figure 4b). By developmental period 2, fish from experimental groups P1-hyperoxia, P2-hyperoxia, and CE-hyperoxia exhibited the highest SGR (~4.5% d−1) and the lowest was registered in larvae from P3-hyperoxia (3.5 ± 0.4% d−1), which did not present significant differences compared to the control (3.9 ± 0.2% d−1; p > 0.05; Figure 4b). At the end of the experiment, fishes from experimental groups P1-hyperoxia and P2-hyperoxia showed the highest SGR values (6.1 ± 0.4% d−1 and 5.4 ± 0.8% d−1 respectively; Figure 4b). Figure 4e shows the overall interactions in this experiment. From ~6 dph, larvae from experimental group P3-hyperoxia showed no increase in SGR throughout 12 dph. Fish from groups with the highest SGR (P1-hyperoxia and P2-hyperoxia) increased their SGR at ~9 dph. For the rest of the groups, a continuous but lower increase than P1-hyperoxia and P2-hyperoxia occurred (p < 0.001).
- Salinity. By developmental period 1, SGR in experimental groups P1-6.0 (1.28 ± 0.1% d−1) and CE-6.0 (1.53 ± 0.2% d−1) was significantly higher than larvae from the control (0.73 ± 0.1% d−1; p < 0.001; Figure 4c). During developmental period 2, no significant differences (p > 0.05) were observed (SGR ~3.4–3.9% d−1; Figure 4c). By developmental period 3, higher SGR was observed in experimental groups P1-4.0 (4.8 ± 0.6% d−1) and P1-6.0 (5.3 ± 0.8% d−1) compared to the rest of the fish (SGR ~3.9–4.2% d−1; p < 0.001; Figure 4c). Figure 4f shows similar values of SGR in all the groups by hatching and 1 dph for the seven surviving populations. The control group and the four surviving groups show a similar pattern in SGR along the experiment. However, by 8 dph, fish from P1-4.0 and P1-6.0 started to show higher SGR values (p < 0.001).
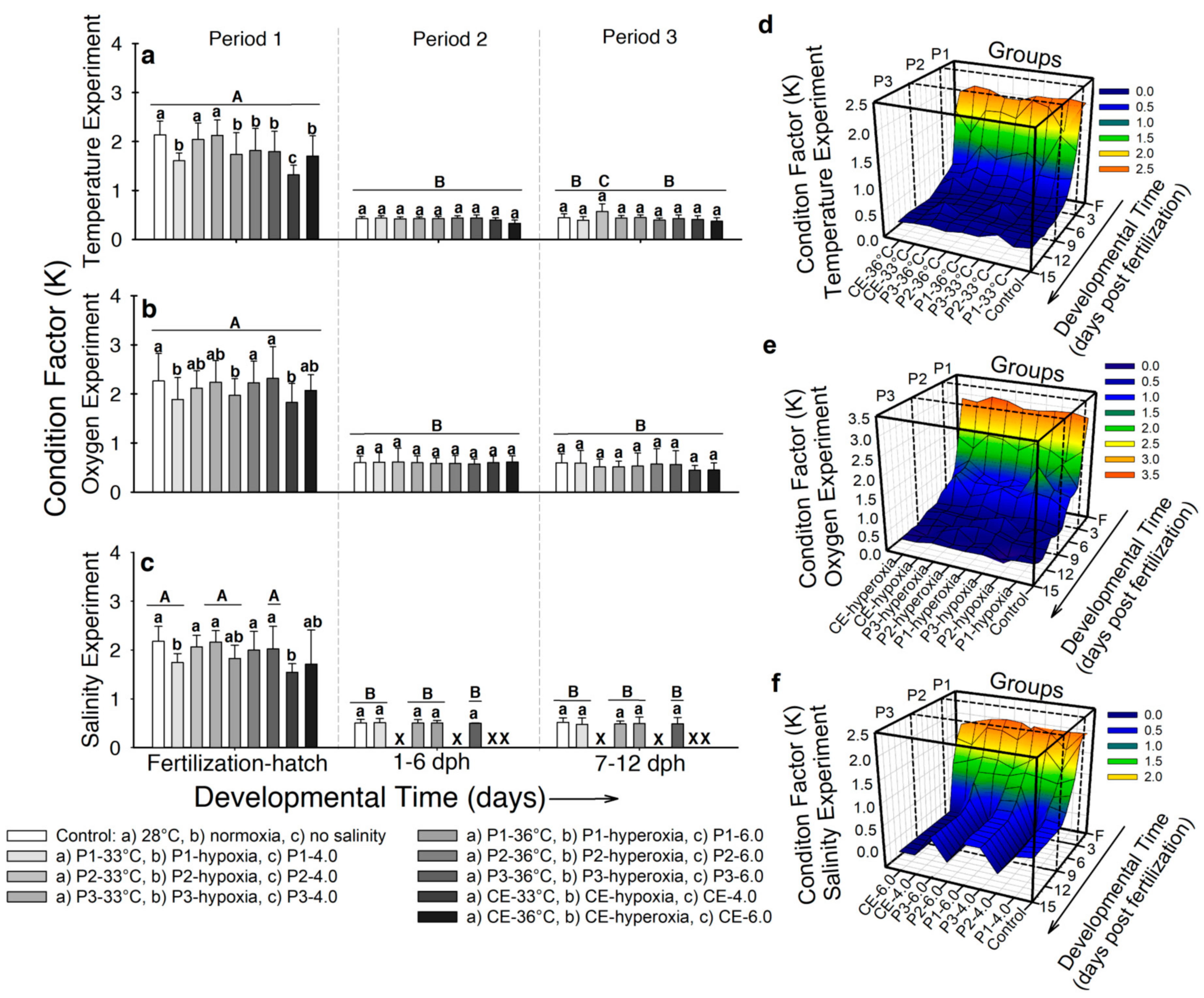
2.3.4. Fulton’s Condition Factor (K)
- Temperature. In developmental period 1, larvae from the control group and experimental groups P2-33 °C and P3-33 °C showed the highest Fulton’s condition factor (K) values (2.13 ± 0.3) and the larvae from experimental group CE-33 °C presented the lowest (1.32 ± 0.2; p < 0.001; Figure 5a). No significant differences were exhibited by developmental periods 2 and 3 (p > 0.05; Figure 5a). Figure 5d shows the interaction of K values, temperature, and time of exposure. Condition factor decreases in all the groups as development progresses. Peaks in experimental groups P3-33 °C, P3-36 °C, and CE-33 °C occurred at different times of development (7 dph, 9 dph, and 4 dph respectively). However, by the end of developmental periods 2 and 3, no difference was observed (p > 0.05).
- Air saturation. In developmental period 1, larvae from the control and experimental groups P2-hypoxia, P3-hypoxia, P2-hyperoxia, and P3-hyperoxia showed higher K values (K~2.1) than the larvae incubated in hypoxia and hyperoxia (K~1.8; p < 0.001; Figure 5b). No significant differences in K occurred in developmental periods 2 and 3 and K values occurred as ~0.57 (p > 0.05; Figure 5b). Figure 5e shows how K values decreased in most of the groups from hatch to 5 dph. Two peaks were registered at 2 dph and 3 dph for experimental groups P1-hypoxia and P2-hypoxia, respectively. However, one day later K decreased to values close to the control group (K = 1.2 ± 0.1).
- Salinity. At the end of developmental period 1, the control and larvae from experimental groups P2-4.0, P3-4.0, P2-6.0, and P3-6.0 showed higher K values (~2.1) than larvae incubated in higher salinity (K ~1.7–1.9; p < 0.001; Figure 5c). No differences in K occurred by developmental periods 2 and 3 (K ~0.5; p > 0.05; Figure 5c). Figure 5f shows how K values decreased from hatching to 5 dph and remained constant for the rest of the experiment in the five surviving groups.
2.4. Timing of Developmental Events as a Function of Stressors and Exposure Stage
2.4.1. Time to Hatching
2.4.2. Time to Exogenous Feeding
2.4.3. Time to Yolk Depletion
2.4.4. Time to Free Swimming
2.4.5. Time to Snout Shape Change
3. Discussion
3.1. Critical Windows for Survival
3.2. Growth
3.3. Developmental Events
3.3.1. Time to Hatching
3.3.2. Time to Exogenous Feeding
3.3.3. Time to Yolk Depletion
3.3.4. Time to Free Swimming
3.3.5. Time to Snout Shape Change
4. Materials and Methods
4.1. Ethical Statement
4.2. Fish Acquisition and Maintenance
4.3. Developmental Stages and Experimental Design
4.4. Treatment Protocols
4.4.1. Temperature
4.4.2. Air Saturation
4.4.3. Salinity
4.5. Survival
4.6. Morphological Variables
4.7. Developmental Events
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Burggren, W.W.; Reyna, K.S. Developmental trajectories, critical windows and phenotypic alteration during cardio-respiratory development. Respir. Physiol. Neurobiol. 2011, 178, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, J.A. Old wine in new bottles: Reaction norms in salmonid fishes. Heredity 2011, 106, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Burggren, W.W. Phenotypic switching resulting from developmental plasticity: Fixed or reversible? Front. Physiol. 2020, 10, 1634. [Google Scholar] [CrossRef] [PubMed]
- Vehaskari, V.M.; Aviles, D.H.; Manning, J. Prenatal programming of adult hypertension in the rat. Kidney Int. 2001, 59, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.M.; Huryn, V.M.; Downes, S.R.; Mercer, A.R. The effects of queenlessness on the maturation of the honeybee olfactory system. Behav. Brain Res. 1998, 91, 115–126. [Google Scholar] [CrossRef]
- Sallout, B.; Walker, M. The fetal origin of adult diseases. J. Obstet. Gynaecol. 2003, 23, 555–560. [Google Scholar] [CrossRef]
- Pinkerton, K.E.; Joad, J.P. The mammalian respiratory system and critical windows of exposure for children’s health. Environ. Health Perspect. 2000, 108 (Suppl. S3), 457–462. [Google Scholar] [PubMed]
- Hogan, N.S.; Duarte, P.; Wade, M.G.; Lean, D.R.; Trudeau, V.L. Estrogenic exposure affects metamorphosis and alters sex ratios in the northern leopard frog (Rana pipiens): Identifying critically vulnerable periods of development. Gen. Comp. Endocrinol. 2008, 156, 515–523. [Google Scholar] [CrossRef]
- Burggren, W.W.; Mueller, C.A. Developmental critical windows and sensitive periods as three-dimensional constructs in time and space. Physiol. Biochem. Zool. 2015, 88, 91–102. [Google Scholar] [CrossRef]
- Dzialowski, E.M.; von Plettenberg, D.; Elmonoufy, N.A.; Burggren, W.W. Chronic hypoxia alters the physiological and morphological trajectories of developing chicken embryos. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 131, 713–724. [Google Scholar] [CrossRef]
- Chan, T.; Burggren, W.W. Hypoxic incubation creates differential morphological effects during specific developmental critical windows in the embryo of the chicken (Gallus gallus). Respir. Physiol. Neurobiol. 2005, 145, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wong-Riley, M.T. Postnatal changes in the expressions of serotonin 1A, 1B, and 2A receptors in ten brain stem nuclei of the rat: Implication for a sensitive period. Neuroscience 2010, 165, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Tate, K.B.; Kohl, Z.F.; Eme, J.; Rhen, T.; Crossley, D.A. Critical windows of cardiovascular susceptibility to developmental hypoxia in common snapping turtle (Chelydra serpentina) embryos. Physiol. Biochem. Zool. 2015, 88, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.A.; Willis, E.; Burggren, W.W. Salt sensitivity of the morphometry of Artemia franciscana during development: A demonstration of 3D critical windows. J. Exp. Biol. 2016, 219 Pt 4, 571–581. [Google Scholar] [CrossRef]
- Burggren, W.W.; Elmonoufy, N.A. Critical developmental windows for morphology and hematology revealed by intermittent and continuous hypoxic incubation in embryos of quail (Coturnix coturnix). PLoS ONE 2017, 12, e0183649. [Google Scholar] [CrossRef]
- Mueller, C.A.; Eme, J.; Manzon, R.G.; Somers, C.M.; Boreham, D.R.; Wilson, J.Y. Embryonic critical windows: Changes in incubation temperature alter survival, hatching phenotype, and cost of development in lake whitefish (Coregonus clupeaformis). J. Comp. Physiol. B 2015, 185, 315–331. [Google Scholar] [CrossRef]
- Lee Pow, C.S.D.; Tilahun, K.; Creech, K.; Law, J.M.; Cope, W.G.; Kwak, T.J.; Kullman, S.W. Windows of susceptibility and consequences of early life exposures to 17β–estradiol on Medaka (Oryzias latipes) reproductive success. Environ. Sci. Technol. 2017, 51, 5296–5305. [Google Scholar] [CrossRef]
- Kupsco, A.; Schlenk, D. Stage susceptibility of Japanese medaka (Oryzias latipes) to selenomethionine and hypersaline developmental toxicity. Environ. Toxicol. Chem. 2016, 35, 1247–1256. [Google Scholar] [CrossRef]
- Spicer, J.I.; Burggren, W.W. Development of physiological regulatory systems: Altering the timing of crucial events. Zoology 2003, 106, 91–99. [Google Scholar] [CrossRef]
- Spicer, J.I.; Rundle, S.D. Plasticity in the timing of physiological development: Physiological heterokairy—What is it, how frequent is it, and does it matter? Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 712–719. [Google Scholar] [CrossRef]
- Spicer, J.I.; Rundle, S.D.; Tills, O. Studying the altered timing of physiological events during development: It’s about time… or is it? Respir. Physiol. Neurobiol. 2011, 178, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Rundle, S.D.; Spicer, J.I. Heterokairy: A significant form of developmental plasticity? Biol. Lett. 2016, 12, 20160509. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, S.; Bayley, M.; McKenzie, D.J. Measuring oxygen uptake in fishes with bimodal respiration. J. Fish Biol. 2016, 88, 206–231. [Google Scholar] [CrossRef] [PubMed]
- Little, C. The Colonisation of Land: Origins and Adaptations of Terrestrial Animals; Cambridge University Press: Cambridge, UK, 1983; p. 290. [Google Scholar]
- Milsom, W.K. New insights into gill chemoreception: Receptor distribution and roles in water and air breathing fish. Respir. Physiol. Neurobiol. 2012, 184, 326–339. [Google Scholar] [CrossRef]
- Perry, S.F.; Wilson, R.J.; Straus, C.; Harris, M.B.; Remmers, J.E. Which came first, the lung or the breath? Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 37–47. [Google Scholar] [CrossRef]
- Randall, D.J.; Ip, Y.K. Ammonia as a respiratory gas in water and air-breathing fishes. Respir. Physiol. Neurobiol. 2006, 154, 216–225. [Google Scholar] [CrossRef]
- Shartau, R.B.; Brauner, C.J. Acid–base and ion balance in fishes with bimodal respiration. J. Fish Biol. 2014, 84, 682–704. [Google Scholar] [CrossRef]
- Johnson, D.W.; Christie, M.R.; Moye, J. Quantifying evolutionary potential of marine fish larvae: Heritability, selection, and evolutionary constraints. Evolution 2010, 64, 2614–2628. [Google Scholar] [CrossRef]
- Blank, T.; Burggren, W.W. Hypoxia-induced developmental plasticity of the gills and air-breathing organ of Trichopodus trichopterus. J. Fish Biol. 2014, 84, 808–826. [Google Scholar] [CrossRef]
- Gonzalez, R.J.; Brauner, C.J.; Wang, Y.X.; Richards, J.G.; Patrick, M.L.; Xi, W.; Matey, V.; Val, A.L. Impact of ontogenetic changes in branchial morphology on gill function in Arapaima gigas. Physiol. Biochem. Zool. 2010, 83, 322–332. [Google Scholar] [CrossRef]
- Joss, J.M. Lungfish evolution and development. Gen. Comp. Endocrinol. 2006, 148, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Sanchez, J.F.; Burggren, W.W. Environmental modulation of the onset of air breathing and survival of Betta splendens and Trichopodus trichopterus. J. Fish Biol. 2014, 84, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.A.; Joss, J.M.; Seymour, R.S. Effects of environmental oxygen on development and respiration of Australian lungfish (Neoceratodus forsteri) embryos. J. Comp. Physiol. B 2011, 181, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M. The pattern of histogenesis and growth of tooth plates in larval stages of extant lungfish. J. Anat. 1985, 140 Pt 4, 627–643. [Google Scholar]
- Aguilera, C.; Mendoza, R.; Rodríguez, G.; Márquez, G. Morphological description of alligator gar and tropical gar larvae, with an emphasis on growth indicators. Trans. Am. Fish. Soc. 2002, 131, 899–909. [Google Scholar] [CrossRef]
- Burggren, W.W.; Bautista, G.M.; Coop, S.C.; Couturier, G.M.; Delgadillo, S.P.; García, R.M.; González, C.A.A. Developmental cardiorespiratory physiology of the air-breathing tropical gar, Atractosteus tropicus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R689–R701. [Google Scholar] [CrossRef]
- Comabella, Y.; Azanza, J.; Hurtado, A.; Canabal, J.; García-Galano, T. Allometric growth in Cuban gar (Atractosteus tristoechus) larvae. Univ. Cienc. 2013, 29, 2227–2690. [Google Scholar]
- Dean, B. The early development of garpike and sturgeon. J. Morphol. 1895, 11, 1–62. [Google Scholar] [CrossRef]
- Echelle, A.A.; Riggs, C.D. Aspects of the early life history of gars (Lepisosteus) in Lake Texoma. Trans. Am. Fish. Soc. 1972, 101, 106–112. [Google Scholar] [CrossRef]
- Long, W.L.; Ballard, W.W. Normal embryonic stages of the longnose gar, Lepisosteus osseus. BMC Dev. Biol. 2001, 1, 6. [Google Scholar] [CrossRef]
- Mendoza, R.; Aguilera, C.; Rodríguez, G.; González, M.; Castro, R. Morphophysiological studies on alligator gar (Atractosteus spatula) larval development as a basis for their culture and repopulation of their natural habitats. Rev. Fish Biol. Fish. 2002, 12, 133–142. [Google Scholar] [CrossRef]
- Barrientos-Villalobos, J.; Espinosa de los Monteros, A. Genetic variation and recent population history of the tropical gar Atractosteus tropicus Gill (Pisces: Lepisosteidae). J. Fish Biol. 2008, 73, 1919–1936. [Google Scholar] [CrossRef]
- Bussing, W.A. Freshwater Fishes of Costa Rica; Editorial Universidad de Costa Rica: San José, Costa Rica, 1998; Volume 46, p. 468. [Google Scholar]
- Miller, R.R.; Minckley, W.L.; Norris, S.M. Freshwater Fishes of Mexico; University of Chicago Press: Chicago, IL, USA, 2005; p. 490. [Google Scholar]
- Mora, M.; Cabrera-Peña, J.; Galeano, G. Reproducción y alimentación del gaspar Atractosteus tropicus (Pisces: Lepisosteidae) en el refugio nacional de vida silvestre Caño Negro, Costa Rica. Rev. Biol. Trop. 1997, 45, 861–866. [Google Scholar]
- Burggren, W.W.; Bagatto, B. Cardiovascular anatomy and physiology. In Fish Larval Physiology; Finn, R.N., Kapoor, B.J., Eds.; Science Publishers: Enfield, NH, USA, 2008; pp. 119–161. [Google Scholar]
- Rudneva, I. Biomarkers for Stress in Fish Embryos and Larvae; CRS Press: Boca Raton, FL, USA, 2014; p. 206. [Google Scholar]
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.J. Overview of hypoxia around the world. J. Environ. Qual. 2001, 30, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Comabella, Y.; Hurtado, A.; Canabal, J.; García-Galano, T. Effect of temperature on hatching and growth of Cuban gar (Atractosteus tristoechus) larvae. ERA 2014, 1, 19–32. [Google Scholar]
- Del Río, A.M.; Davis, B.E.; Fangue, N.A.; Todgham, A.E. Combined effects of warming and hypoxia on early life stage Chinook salmon physiology and development. Conserv. Physiol. 2019, 7, coy078. [Google Scholar] [CrossRef]
- Hassell, K.L.; Coutin, P.C.; Nugegoda, D. Hypoxia impairs embryo development and survival in black bream (Acanthopagrus butcheri). Mar. Pollut. Bull. 2008, 57, 302–306. [Google Scholar] [CrossRef]
- Hou, Z.S.; Wen, H.S.; Li, J.F.; He, F.; Li, Y.; Qi, X. Environmental hypoxia causes growth retardation, osteoclast differentiation and calcium dyshomeostasis in juvenile rainbow trout (Oncorhynchus mykiss). Sci. Total Environ. 2020, 705, 135272. [Google Scholar] [CrossRef]
- Levesque, K.D.; Wright, P.A.; Bernier, N.J. Cross talk without cross tolerance: Effect of rearing temperature on the hypoxia response of embryonic zebrafish. Physiol. Biochem. Zool. 2019, 92, 349–364. [Google Scholar] [CrossRef]
- Fry, J. The effect of environmental factors on the physiology of fish. In Fish Physiology 5; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA, 1974; pp. 1–98. [Google Scholar]
- Matthews, M.D.; Prangnell, D.; Glenewinkel, H. Sensitivity of Guadalupe bass swim-up fry to hyperoxia. N. Am. J. Aquac. 2017, 79, 289–298. [Google Scholar] [CrossRef]
- Pleizier, N.; Nelson, C.; Cooke, S.; Brauner, C.J. Understanding gas bubble trauma in an era of hydropower expansion: How do fish compensate at depth? Can. J. Fish. Aquat. Sci. 2019, 77, 556–563. [Google Scholar] [CrossRef]
- Beamish, F.W.H. Osmoregulation in juvenile and adult lampreys. Can. J. Fish. Aquat. Sci. 1980, 37, 1739–1750. [Google Scholar] [CrossRef]
- Richards, J.E.; Beamish, F.W.H. Initiation of feeding and salinity tolerance in the pacific lamprey Lampetra tridentata. Mar. Biol. 1981, 63, 73–77. [Google Scholar] [CrossRef]
- McDowall, R.M. Diadromy in Fishes: Migrations between Freshwater and Marine Environments; Timber Press: Portland, OR, USA, 1988; p. 308. [Google Scholar]
- Allen, P.J.; Cech, J.J. Age/size effects on juvenile green sturgeon, Acipenser medirostris, oxygen consumption, growth, and osmoregulation in saline environments. Environ. Biol. Fishes 2007, 79, 211–229. [Google Scholar] [CrossRef]
- Allen, P.J.; McEnroe, M.; Forostyan, T.; Cole, S.; Nicholl, M.M.; Hodge, B.; Cech, J.J. Ontogeny of salinity tolerance and evidence for seawater-entry preparation in juvenile green sturgeon, Acipenser medirostris. J. Comp. Physiol. B 2011, 181, 1045–1062. [Google Scholar] [CrossRef]
- Schwarz, D.E.; Allen, P.J. Effects of salinity on growth and ion regulation of juvenile alligator gar Atractosteus spatula. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2014, 169, 44–50. [Google Scholar] [CrossRef]
- Brauner, C.J.; Rombough, P.J. Ontogeny and paleophysiology of the gill: New insights from larval and air-breathing fish. Respir. Physiol. Neurobiol. 2012, 184, 293–300. [Google Scholar] [CrossRef]
- Kwan, G.T.; Wexler, J.B.; Wegner, N.C.; Tresguerres, M. Ontogenetic changes in cutaneous and branchial ionocytes and morphology in yellowfin tuna (Thunnus albacares) larvae. J. Comp. Physiol. B 2019, 189, 81–95. [Google Scholar] [CrossRef]
- Melo, L.H.; Martins, Y.S.; Melo, R.M.C.; Prado, P.S.; Luz, R.K.; Bazzoli, N.; Rizzo, E. Low salinity negatively affects early larval development of Nile tilapia, Oreochromis niloticus: Insights from skeletal muscle and molecular biomarkers. Zygote 2019, 27, 375–381. [Google Scholar] [CrossRef]
- Tresguerres, M.; Clifford, A.M.; Harter, T.S.; Roa, J.N.; Thies, A.B.; Yee, D.P.; Brauner, C.J. Evolutionary links between intra-and extracellular acid–base regulation in fish and other aquatic animals. J. Exp. Zool. A Ecol. Integr. Physiol. 2020, 333, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.D.; Thomas, G.A.; Clark, A.L. Early piscivory and timing of the critical period in postlarval longnose gar at mile 571 of the Ohio River. Trans. Ky. Acad. Sci. 1979, 40, 122–128. [Google Scholar]
- Simon, T.P.; Tyberghein, E.J. Contributions to the early life history of the spotted gar, Lepisosteus oculatus Winchell, from Hatchet Creek, Alabama. J. Ky. Acad. Sci. 1991, 52, 124–131. [Google Scholar]
- Hjort, J. Fluctuations in the Great Fisheries of Northern Europe Viewed in the Light of Biological Research; Andr. Fred. Høst & Søn: Copenhagen, Denmark, 1914; Volume 20, pp. 1–228. [Google Scholar]
- Jonassen, T.M.; Imsland, A.K.; Stefansson, S.O. The interaction of temperature and fish size on growth of juvenile halibut. J. Fish Biol. 1999, 54, 556–572. [Google Scholar] [CrossRef]
- Keckeis, H.; Kamler, E.; Bauer-Nemeschkal, E.; Schneeweiss, K. Survival, development and food energy partitioning of nase larvae and early juveniles at different temperatures. J. Fish Biol. 2001, 59, 45–61. [Google Scholar] [CrossRef]
- Mendiola, D.; Alvarez, P.; Cotano, U.; Martínez de Murguía, A. Early development and growth of the laboratory reared north-east Atlantic mackerel Scomber scombrus L. J. Fish Biol. 2007, 70, 911–933. [Google Scholar] [CrossRef]
- Person-Le Ruyet, J.; Buchet, V.; Vincent, B.; Le Delliou, H.; Quéméner, L. Effects of temperature on the growth of pollack (Pollachius pollachius) juveniles. Aquaculture 2006, 251, 340–345. [Google Scholar] [CrossRef]
- Boeuf, G.; Payan, P. How should salinity influence fish growth? Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 411–423. [Google Scholar] [CrossRef]
- Smatresk, N.J.; Cameron, J.N. Respiration and acid-base physiology of the spotted gar, a bimodal breather: III. Response to a transfer from fresh water to 50% sea water, and control of ventilation. J. Exp. Biol. 1982, 96, 295–306. [Google Scholar]
- Malik, A.; Abbas, G.; Jabbar, A.; Sajjad Shah, S.; Ali Muhammad, A. Effect of different salinity level on spawning, fertilization, hatching and survival of common carp, Cyprinus carpio (Linnaeus, 1758) in semi-artificial environment. Iran. J. Fish. Sci. 2018, 17, 790–804. [Google Scholar]
- Vetemaa, M.; Saat, T. Effects of salinity on the development of fresh-water and brackish-water ruffe Gymnocephalus cernuus (L.) embryos. Ann. Zool. Fenn. 1996, 33, 687–691. [Google Scholar]
- Hernández-Rubio, M.C.; Figueroa-Lucero, G. Effects of temperature and salinity during the embryonic period of Chirostoma humboldtianum and Chirostoma riojai (Atherinopsidae) until hatching. Hidrobiológica 2013, 23, 365–373. [Google Scholar]
- Holliday, F.G.T. The effect of salinity on the eggs and larvae of teleost. Fish Physiol. 1969, 1, 293–311. [Google Scholar]
- Eddy, F.B.; Handy, R.D. Ecological and Environmental Physiology of Fishes; Oxford University Press: Oxford, UK, 2012; p. 252. [Google Scholar]
- Shumway, D.L.; Warren, C.E.; Doudoroff, P. Influence of oxygen concentration and water movement on the growth of steelhead trout and coho salmon embryos. Trans. Am. Fish. Soc. 1964, 93, 342–356. [Google Scholar] [CrossRef]
- Marks, C.; Kaut, K.P.; Moore, F.B.; Bagatto, B. Ontogenetic oxygen changes alter zebra fish size, behavior, and blood glucose. Physiol. Biochem. Zool. 2012, 85, 635–644. [Google Scholar] [CrossRef]
- Rombough, P.J. Growth, aerobic metabolism, and dissolved oxygen requirements of embryos and alevins of steelhead, Salmo gairdneri. Can. J. Zool. 1988, 66, 651–660. [Google Scholar] [CrossRef]
- Mason, J.C. Hypoxial stress prior to emergence and competition among coho salmon fry. J. Fish. Res. Board Can. 1969, 26, 63–91. [Google Scholar] [CrossRef]
- Pepin, P. Effect of temperature and size on development, mortality, and survival rates of the pelagic early life history stages of marine fish. Can. J. Fish. Aquat. Sci. 1991, 48, 503–518. [Google Scholar] [CrossRef]
- Tills, O.; Spicer, J.I.; Rundle, S.D. Salinity-induced heterokairy in an upper-estuarine population of the snail Radix balthica (Mollusca: Pulmonata). Aquat. Biol. 2010, 9, 95–105. [Google Scholar] [CrossRef]
- Márquez-Couturier, G.; Vázquez-Navarrete, C.J.; Contreras-Sánchez, W.M.; Álvarez-González, C.A. Acuicultura Tropical Sustentable: Una Estrategia para la Producción y Conservación del Pejelagarto (Atractosteus tropicus) en Tabasco, México, 2nd ed.; Colección José Narciso Rovirosa; Universidad Juárez Autónoma de Tabasco: Tabasco, México, 2015; Volume 3, p. 223. [Google Scholar]
- Frías-Quintana, C.A.; Álvarez-González, C.A.; Márquez-Couturier, G. Diseño de microdietas para el larvicultivo de pejelagarto Atractosteus tropicus, Gill 1863. Univ. Cienc. 2010, 26, 265–282. [Google Scholar]
- Morales, G. Reproducción y Desarrollo Embriológico del Catán (Lepisosteus spatula Lacepede): Primeros Resultados; SEPES. Dirección General de Acuacultura: Ciudad de Mexico, Mexico, 1987; p. 70. [Google Scholar]
- Morales, G. Especies nativas de México. Acuavisión 1999, 12, 7–9. [Google Scholar]
- Rao, T.R. Influence of salinity on the eggs and larvae of the California killifish Fundulus parvipinnis. Mar. Biol. 1974, 24, 155–162. [Google Scholar] [CrossRef]
- Czerkies, P.; Brzuzan, P.; Kordalski, K.; Luczynski, M. Critical partial pressures of oxygen causing precocious hatching in Coregonus lavaretus and C. albula embryos. Aquaculture 2001, 196, 151–158. [Google Scholar] [CrossRef]
- Garside, E.T. Some effects of oxygen in relation to temperature on the development of lake trout embryos. Can. J. Zool. 1959, 37, 689–698. [Google Scholar] [CrossRef]
- Warkentin, K.M. Oxygen, gills, and embryo behavior: Mechanisms of adaptive plasticity in hatching. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Yúfera, M.; Darias, M.J. The onset of exogenous feeding in marine fish larvae. Aquaculture 2007, 268, 53–63. [Google Scholar] [CrossRef]
- Rønnestad, I.; Yúfera, M.; Ueberschär, B.; Ribeiro, L.; Sæle, Ø.; Boglione, C. Feeding behaviour and digestive physiology in larval fish: Current knowledge, and gaps and bottlenecks in research. Rev. Aquacult. 2013, 5, S59–S98. [Google Scholar] [CrossRef]
- Camacho, S.; Carmona, R.; Llorente, J.I.; Sanz, A.; García-Gallego, M.; Domezain, A.; Dominguez, N.; Ostos-Garrido, M.V. Stomach development in the sturgeon Acipenser naccarii: Histoenzymatic and ultrastructural analysis. J. Appl. Ichthyol. 2011, 27, 693–700. [Google Scholar] [CrossRef]
- Dou, S.; Masuda, R.; Tanaka, M.; Tsukamoto, K. Feeding resumption, morphological changes and mortality during starvation in Japanese flounder larvae. J. Fish Biol. 2002, 60, 1363–1380. [Google Scholar] [CrossRef]
- Comabella Soto, Y. Desarrollo Larval del Manjuarí Atractosteus tristoechus (Bloch y Schneider, 1801) (Pisces: Lepisosteidae) en Condiciones de Cultivo: Aspectos Morfo-Fisiológicos. Ph.D. Thesis, Universidad de la Habana, La Habana, Cuba, 2011. [Google Scholar]
- Moteki, M.; Yoseda, K.; Sahin, T.; Üstündağ, C.; Kohno, H. Transition from endogenous to exogenous nutritional sources in larval black sea turbot Psetta maxima. Fish. Sci. 2001, 67, 571–578. [Google Scholar] [CrossRef][Green Version]
- Williams, K.; Papanikos, N.; Phelps, R.P.; Shardo, J.D. Development, growth, and yolk utilization of hatchery-reared red snapper Lutjanus campechanus larvae. Mar. Ecol. Prog. Ser. 2004, 275, 231–239. [Google Scholar] [CrossRef]
- Coughlin, D.J. Ontogeny of feeding behavior of first-feeding Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 1991, 48, 1896–1904. [Google Scholar] [CrossRef]
- Makrakis, M.C.; Nakatani, K.; Bialetzki, A.; Sanches, P.V.; Baumgartner, G.; Gomes, L.C. Ontogenetic shifts in digestive tract morphology and diet of fish larvae of the Itaipu Reservoir, Brazil. Environ. Biol. Fishes 2005, 72, 99–107. [Google Scholar] [CrossRef]
- Collins, L.A.; Nelson, S.G. Effects of temperature on oxygen consumption, growth, and development of embryos and yolk-sac larvae of Siganus randalli (Pisces: Siganidae). Mar. Biol. 1993, 117, 195–204. [Google Scholar] [CrossRef]
- Hart, P.R.; Purser, G.J. Effects of salinity and temperature on eggs and yolk sac larvae of the greenback flounder (Rhombosolea tapirina Günther, 1862). Aquaculture 1995, 136, 221–230. [Google Scholar] [CrossRef]
- May, R.C. Effects of temperature and salinity on yolk utilization in Bairdiella icistia (Jordan & Gilbert) (Pisces: Sciaenidae). J. Exp. Mar. Biol. Ecol. 1974, 16, 213–225. [Google Scholar]
- Khan, U. Effect of salinity on yolk utilization and growth of brook trout alevins (Salvelinus fontinalis). J. Anatol. Environ. Animal Sci. 2019, 4, 93–96. [Google Scholar]
- Massa, F.; Delorme, C.; Baglinière, J.L.; Prunet, P.; Grimaldi, C. Expositions d’oeufs de truite commune (Salmo trutta) à des hypoxies temporaires ou continue: Effets sur la branchie, la résorption de la vésicule vitelline et les caractéristiques morphométriques des alevins. Bull. Français. Pêche Piscic. 1999, 355, 421–440. [Google Scholar] [CrossRef]
- Polymeropoulos, E.T.; Elliott, N.G.; Frappell, P.B. Hypoxic acclimation leads to metabolic compensation after reoxygenation in Atlantic salmon yolk-sac alevins. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 213, 28–35. [Google Scholar] [CrossRef]
- Roussel, J.M. Carry-over effects in brown trout (Salmo trutta): Hypoxia on embryos impairs predator avoidance by alevins in experimental channels. Can. J. Fish. Aquat. Sci. 2007, 64, 786–792. [Google Scholar] [CrossRef]
- Voesenek, C.J.; Muijres, F.T.; van Leeuwen, J.L. Biomechanics of swimming in developing larval fish. J. Exp. Biol. 2018, 221, jeb149583. [Google Scholar] [CrossRef] [PubMed]
- Batty, R.S. Development of swimming movements and musculature of larval herring (Clupea harengus). J. Exp. Biol. 1984, 110, 217–229. [Google Scholar]
- Vieira, V.I.A.; Johnston, I.A. Influence of temperature on muscle-fibre development in larvae of the herring Clupea harengus. Mar. Biol. 1992, 112, 333–341. [Google Scholar] [CrossRef]
- Domenici, P.; Herbert, N.A.; Lefrançois, C.; Steffensen, J.F.; McKenzie, D.J. The effect of hypoxia on fish swimming performance and behaviour. In Swimming Physiology of Fish; Springer: Berlin/Heidelberg, Germany, 2013; pp. 129–159. [Google Scholar]
- Kaufmann, R.; Wieser, W. Influence of temperature and ambient oxygen on the swimming energetics of cyprinid larvae and juveniles. In Environmental Biology of European Cyprinids; Weiser, W., Scheimer, F., Goldschmidt, A., Kotrschal, K., Eds.; Springer Science & Business Media: Luxemburg, 1992; pp. 87–96. [Google Scholar]
- Márquez-Couturier, G.; Álvarez-González, C.A.; Contreras-Sánchez, W.M.; Hernández-Vidal, U.; Hernández-Franyutti, A.A.; Mendoza-Alfaro, R.E.; Aguilera-González, C.; García-Galano, T.; Civera-Cerecedo, R.; Goytortua-Bores, E. Avances en la alimentación y nutrición del pejelagarto Atractosteus tropicus. In Avances en Nutrición Acuícola VIII, Proceedings of the VIII Simposium Internacional de Nutrición Acuícola, Mazatlán, SI, México, 15–17 November 2006; Cruz-Suárez, E., Ricque, D.M., Tapia-Salazar, M., Nieto-López, M.G., Villarreal-Cavazos, D.A., Puello-Cruz, A.C., García-Ortega, A., Eds.; Universidad Autónoma de Nuevo León: Monterrey, NL, Mexico, 2006; pp. 446–523. [Google Scholar]
- Busch, A. Transition from endogenous to exogenous nutrition: Larval size parameters determining the start of external feeding and size of prey ingested by Ruegen spring herring Clupea harengus. Mar. Ecol. Prog. Ser. 1996, 130, 39–45. [Google Scholar] [CrossRef]
- Dabrowski, K.; Bardega, R. Mouth size and predicted food size preferences of larvae of three cyprinid fish species. Aquaculture 1984, 40, 41–46. [Google Scholar] [CrossRef]
- Hecht, T.; Pienaar, A.G. A review of cannibalism and its implications in fish larviculture. J. World Aquac. Soc. 1993, 24, 246–261. [Google Scholar] [CrossRef]
- Hill, L.; Renfro, J.; Reynolds, R. Effects of dissolved oxygen tensions upon the rate of young spotted gar, Lepisosteus oculatus (Lepisosteidae). Southwest. Nat. 1972, 17, 273–278. [Google Scholar] [CrossRef]
- Rimoldi, S.; Terova, G.; Zaccone, G.; Parker, T.; Kuciel, M.; Dabrowski, K. The effect of hypoxia and hyperoxia on growth and expression of hypoxia-related genes and proteins in spotted gar Lepisosteus oculatus larvae and juveniles. J. Exp. Zool. B Mol. Dev. Evol. 2016, 326, 250–267. [Google Scholar] [CrossRef]
- Timmons, M.B.; Ebeling, J.M. Recirculating Aquaculture; Cayuga Aqua Ventures: Ithaca, NY, USA, 2010; p. 976. [Google Scholar]
- Brett, J.R.; Groves, D.D. Physiological energetics. In Volume VIII—Fish Physiology. Bioenergetics and Growth; Hoar, W.S., Randall, D.J., Brett, J.R., Eds.; Academic Press: New York, NY, USA, 1979; p. 344. [Google Scholar]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 1975, 191, 1–382. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event | Experiment | Control | Treatment 1 (33 °C, Hypoxia, or Salinity = 4 ppt) | Treatment 2 (36 °C, Hyperoxia, or Salinity = 6 ppt) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P1-Treatment 1 | P2- Treatment 1 | P3-Treatmemt 1 | CE- Treatment 1 | P1- Treatment 2 | P2- Treatment 2 | P3- Treatment 2 | CE- Treatment 2 | |||
| Hatching | Temperature | 63 ± 4 a | 48 ± 2 b | 63 ± 3 a | 62 ± 3 a | 48 ± 1 b | 36 ± 1 c | 62 ± 2 a | 61 ± 2 a | 36 ± 1 c |
| Oxygen | 60 ± 1 a | 50 ± 1 b | 61 ± 1 a | 61 ± 1 a | 49 ± 1 b | 67 ± 2 c | 60 ± 1 a | 61 ± 1 a | 67 ± 2 c | |
| Salinity | 64 ± 1 a | 72 ± 3 b | 63 ± 1 a | 62 ± 1 a | 72 ± 3 b | 78 ± 3 b | 64 ± 1 a | 63 ± 1 a | 78 ± 3 b | |
| Exogenous Feeding | Temperature | 116 ± 5 a | 98 ± 3 b | 111 ± 6 a | 109 ± 5 a | 88 ± 2 b | 92 ± 2 b | 94 ± 4 b | 112 ± 5 a | 82 ± 1 c |
| Oxygen | 122 ± 3 a | 122 ± 3 a | 121 ± 3 a | 121 ± 3 a | 123 ± 2 a | 123 ± 2 a | 121 ± 3 a | 122 ± 3 a | 123 ± 3 a | |
| Salinity | 122 ± 3 a | 122 ± 1 a | - | 121 ± 1 a | - | 122 ± 2 a | - | 123 ± 1 a | - | |
| Yolk Depletion | Temperature | 124 ± 1 a | 118 ± 1 b | 120 ± 2 b | 125 ± 2 a | 96 ± 1 c | 96 ± 1 c | 115 ± 2 b | 124 ± 3 a | 88 ± 1 d |
| Oxygen | 126 ± 2 a | 132 ± 2 b | 126 ± 3 a | 126 ± 3 a | 125 ± 2 a | 125 ± 2 a | 125 ± 3 a | 127 ± 1 a | 127 ± 1 a | |
| Salinity | 125 ± 2 a | 126 ± 3 a | - | 127 ± 2 a | - | 127 ± 3 a | - | 126 ± 3 a | - | |
| Free Swimming | Temperature | 140 ± 2 a | 125 ± 2 b | 125 ± 1 b | 138 ± 2 a | 105 ± 1 c | 102 ± 1 c | 126 ± 1 b | 140 ± 2 a | 96 ± 1 d |
| Oxygen | 130 ± 3 a | 136 ± 3 a | 132 ± 2 a | 132 ± 2 a | 130 ± 3 a | 136 ± 2 b | 131 ± 3 a | 132 ± 4 a | 131 ± 2 a | |
| Salinity | 142 ± 3 a | 152 ± 3 a | - | 143 ± 2 a | - | 154 ± 3 b | - | 145 ± 3 a | - | |
| Snout Shape Change | Temperature | 168 ± 3 a | 156 ± 2 b | 140 ± 2 c | 168 ± 2 a | 126 ± 1 d | 155 ± 4 b | 152 ± 2 b | 168 ± 4 a | 118 ± 1 d |
| Oxygen | 172 ± 3 a | 183 ± 1 b | 172 ± 3 a | 171 ± 1 a | 173 ± 3 a | 171 ± 3 a | 172 ± 2 a | 171 ± 2 a | 172 ± 3 a | |
| Salinity | 174 ± 4 a | 174 ± 3 a | - | 174 ± 4 a | - | 174 ± 4 a | - | 173 ± 4 a | - | |
| Event | Temperature Intervals | ||
|---|---|---|---|
| Q10 (28–33 °C) | Q10 (33–36 °C) | Q10 (28–36 °C) | |
| Hatching | 1.5 ± 0.2 a | 3.1 ± 0.3 b | 0. 9 ± 0.1 c |
| Exogenous Feeding | 1.5 ± 0.3 a | 3.1 ± 0.4 b | 0.9 ± 0.1 c |
| Yolk Depletion | 1.6 ± 0.2 a | 3.1 ± 0.3 b | 0.9 ± 0.1 c |
| Free Swimming | 1.5 ± 0.4 a | 3.1 ± 0.6 b | 0.9 ± 0.1 c |
| Snout Shape Change | 1.5 ± 0.3 a | 3.1 ± 0.6 b | 0.9 ± 0.1 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, G.; Peña, E.; Martínez, R.; Camarillo, S.; Burggren, W.; Álvarez, A. Survival, Growth, and Development in the Early Stages of the Tropical Gar Atractosteus tropicus: Developmental Critical Windows and the Influence of Temperature, Salinity, and Oxygen Availability. Fishes 2021, 6, 5. https://doi.org/10.3390/fishes6010005
Martínez G, Peña E, Martínez R, Camarillo S, Burggren W, Álvarez A. Survival, Growth, and Development in the Early Stages of the Tropical Gar Atractosteus tropicus: Developmental Critical Windows and the Influence of Temperature, Salinity, and Oxygen Availability. Fishes. 2021; 6(1):5. https://doi.org/10.3390/fishes6010005
Chicago/Turabian StyleMartínez, Gil, Emyr Peña, Rafael Martínez, Susana Camarillo, Warren Burggren, and Alfonso Álvarez. 2021. "Survival, Growth, and Development in the Early Stages of the Tropical Gar Atractosteus tropicus: Developmental Critical Windows and the Influence of Temperature, Salinity, and Oxygen Availability" Fishes 6, no. 1: 5. https://doi.org/10.3390/fishes6010005
APA StyleMartínez, G., Peña, E., Martínez, R., Camarillo, S., Burggren, W., & Álvarez, A. (2021). Survival, Growth, and Development in the Early Stages of the Tropical Gar Atractosteus tropicus: Developmental Critical Windows and the Influence of Temperature, Salinity, and Oxygen Availability. Fishes, 6(1), 5. https://doi.org/10.3390/fishes6010005