Distribution and Risk Assessment of Potential Invasiveness of Australoheros facetus (Jenyns, 1842) in Portugal

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
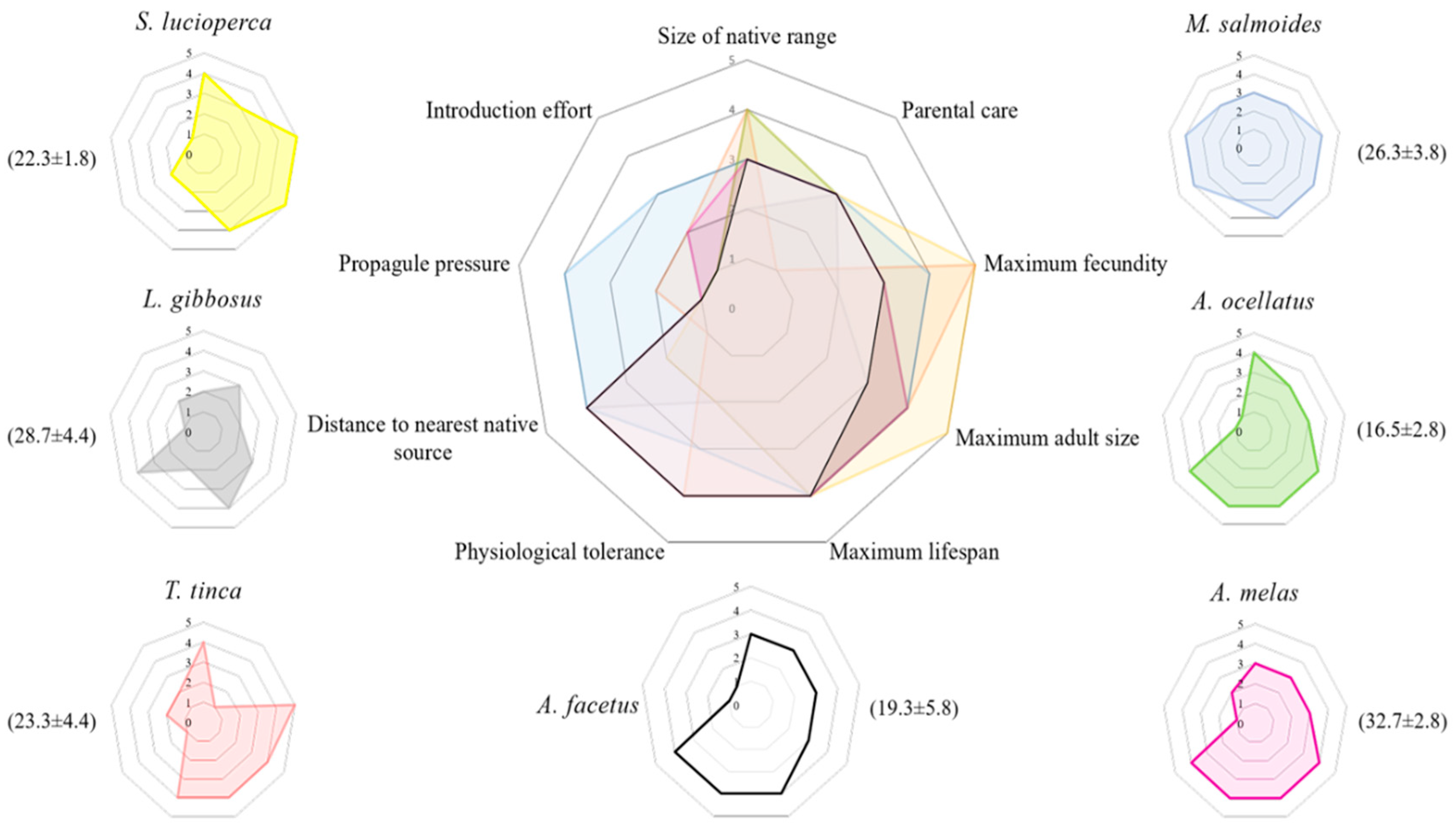
2. Results
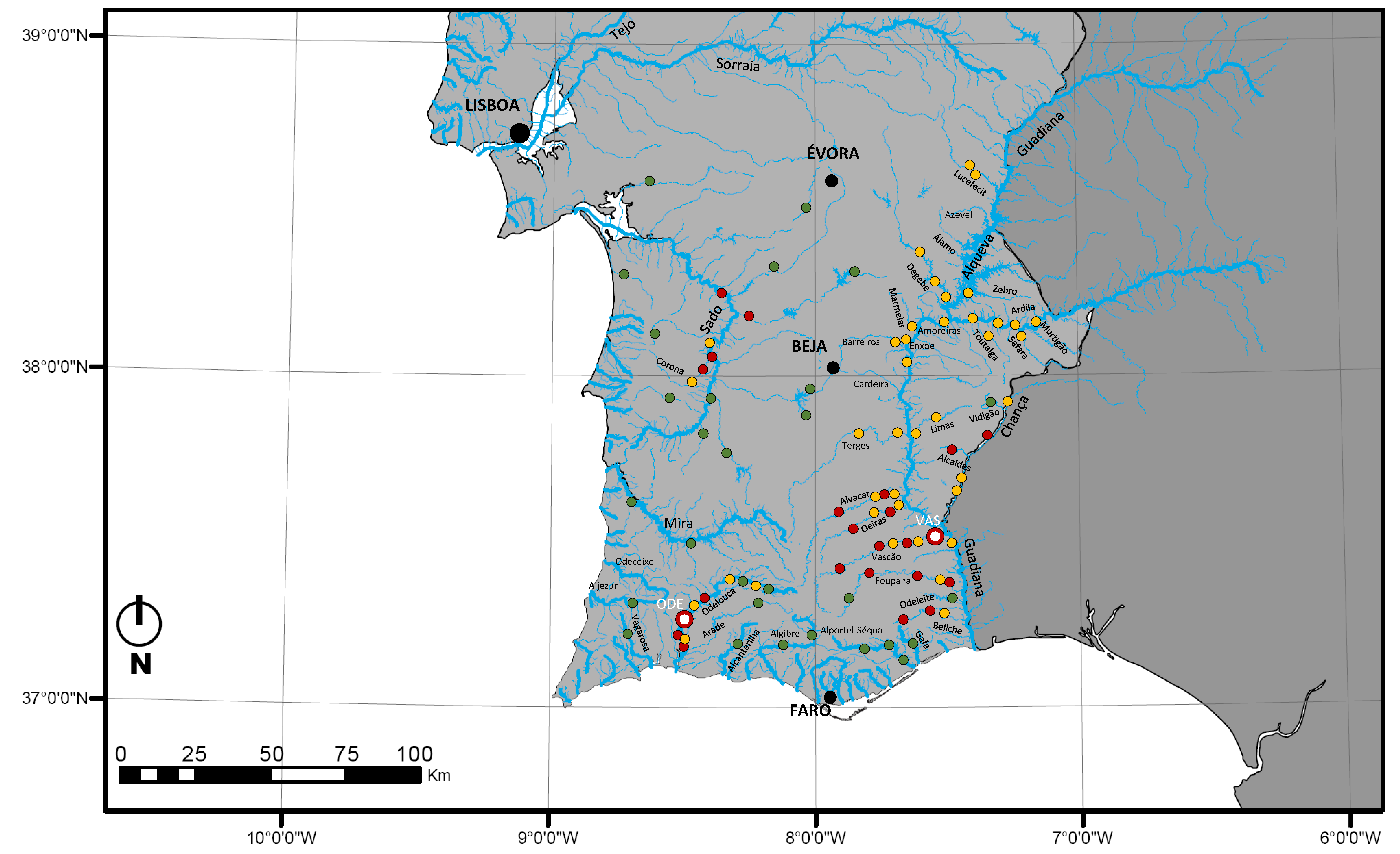
2.1. Australoheros Facetus Distribution in Southern Portugal
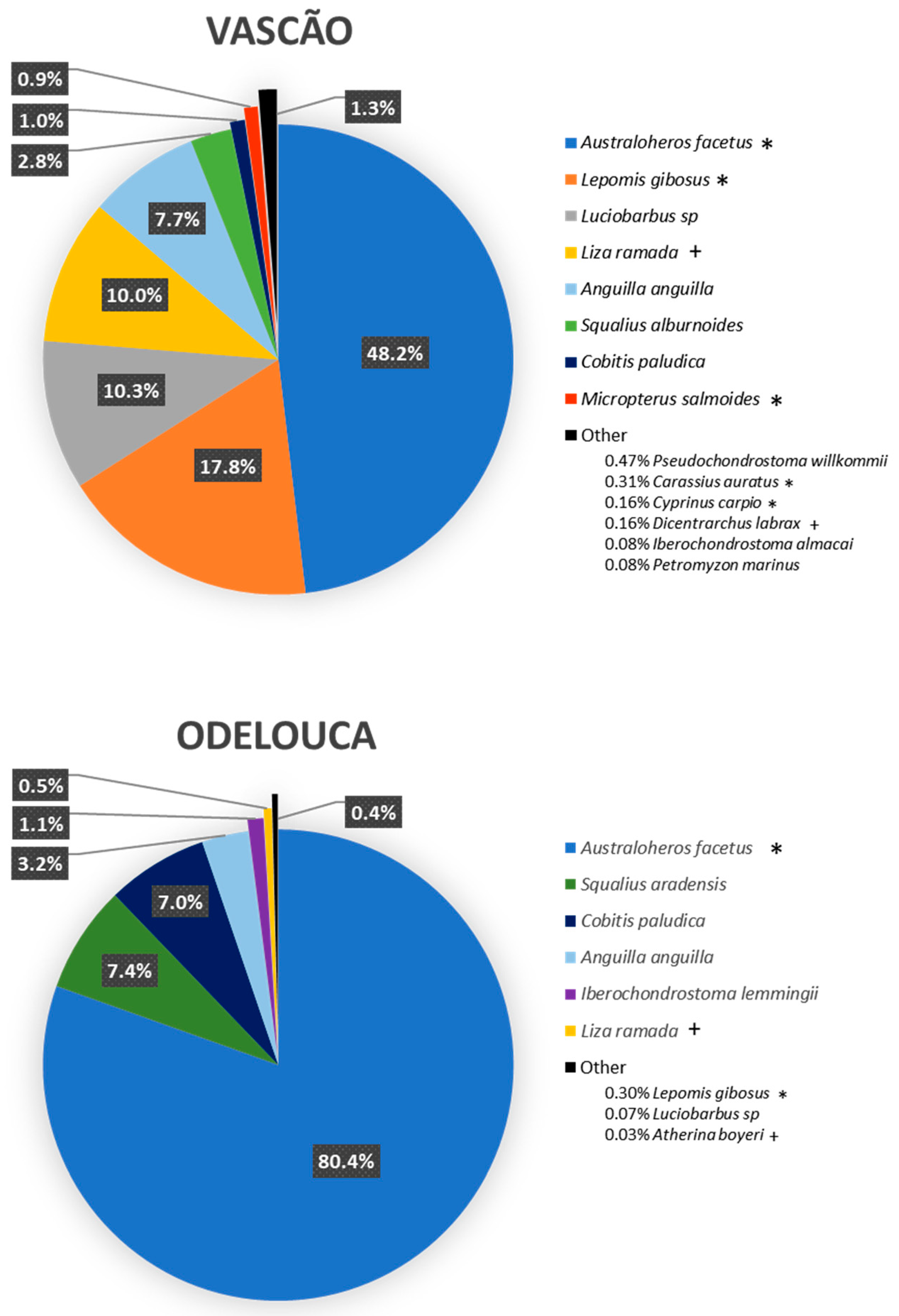
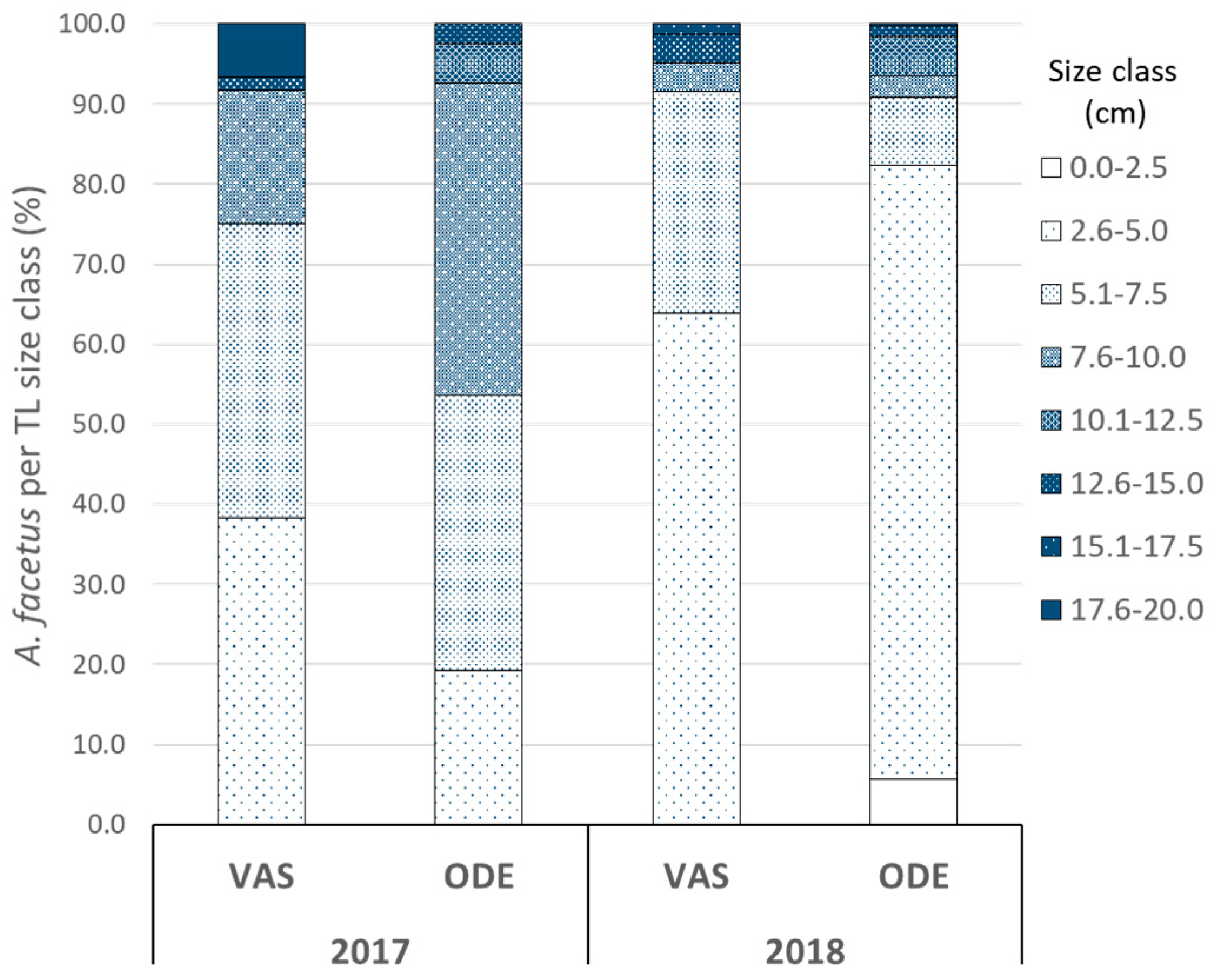
2.2. Relative Abundance of A. facetus in Fish Assemblages in Two Selected Sites
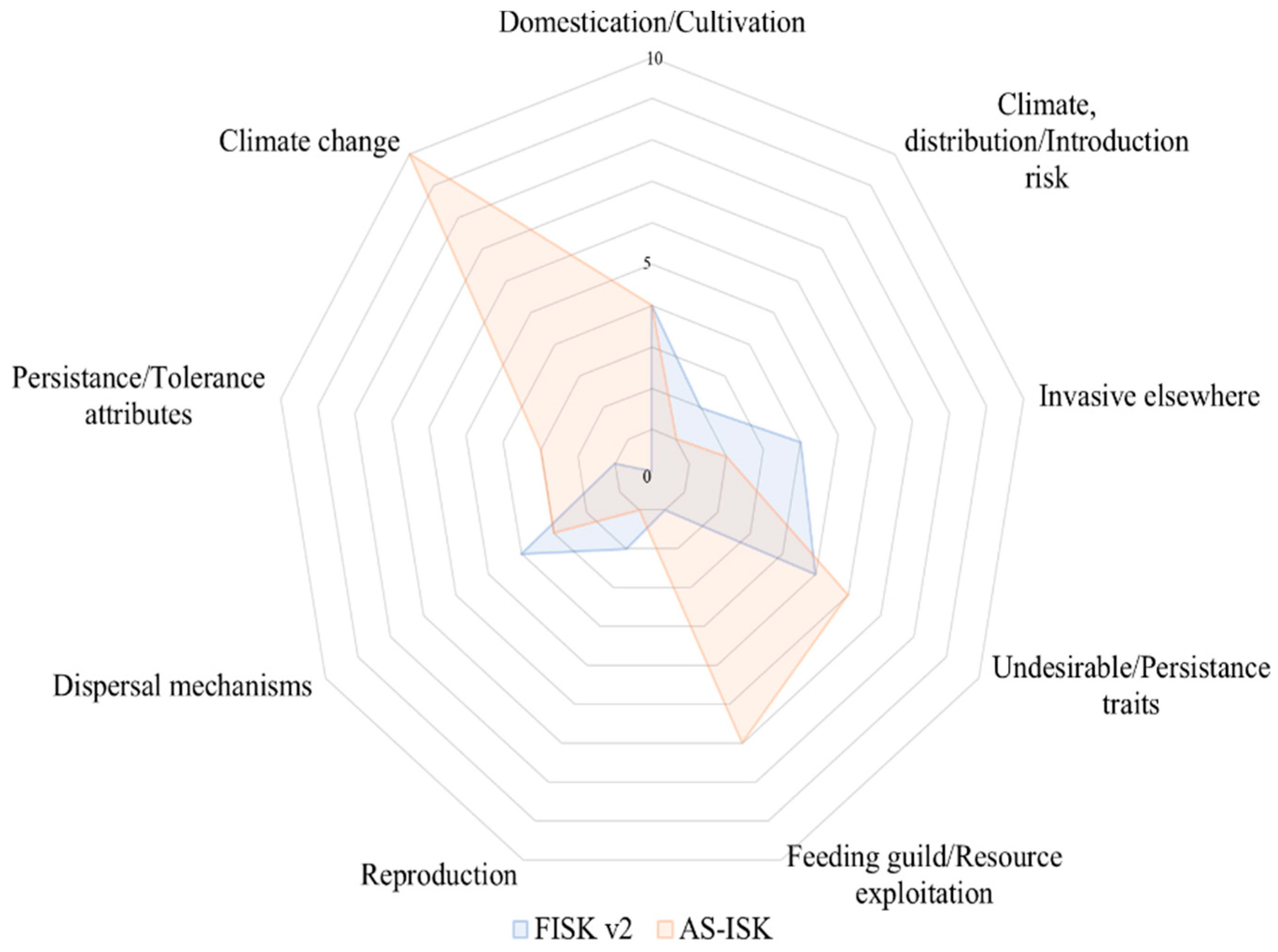
2.3. Risk Assessment Toolkits
3. Discussion
3.1. Distribution of A. facetus in Southern Portugal
3.2. Evaluation of the Toolkits and Reasons for Increased Invasive Potential
3.3. Gaps in Previous Legislation and the Recent Framework
4. Materials and Methods
4.1. Geographical Distribution and Community Surveys
4.2. Application of Risk Assessment Toolkits
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Casal, C.M.V. Global documentation of fish introductions: The growing crisis and recommendations for action. Biol. Invasions 2006, 8, 3–11. [Google Scholar] [CrossRef]
- Clavero, M.; Garcia-Berthou, E. Invasive species are a leading cause of animal extinctions. Trends Ecol. Evol. 2005, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, B.; Clavero, M.; Sanchez, M.I.; Vila, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef] [PubMed]
- International Union for Conservation of Nature. Guidelines for the Prevention of Biodiversity Loss Due to Biological Invasion; IUCN: Gland, Switzerland, 2000; p. 24. [Google Scholar]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2002, 10, 689–710. [Google Scholar] [CrossRef]
- Epanchin-Niell, R.S. Economics of invasive species policy and management. Biol. Invasions 2017. [Google Scholar] [CrossRef]
- Epanchin-Niell, R.S.; Hastings, A. Controlling established invaders: Integrating economics and spread dynamics to determine optimal management. Ecol. Lett. 2010, 13, 528–541. [Google Scholar] [CrossRef]
- Mandrak, N.E.; Cudmore, B. Risk assessment: Cornerstone of an aquatic invasive species program. Aquat. Ecosyst. Health Manag. 2015, 18, 312–320. [Google Scholar] [CrossRef]
- Drake, J.A. Biological Invasions: A Global Perspective; Scientific Committee on Problems of the Environment: Chichester, UK; New York, NY, USA, 1989. [Google Scholar]
- Clavero, M. Assessing the risk of freshwater fish introductions into the Iberian Peninsula. Freshw. Biol. 2011, 56, 2145–2155. [Google Scholar] [CrossRef]
- Vila-Gispert, A.; Alcaraz, C.; Garcia-Berthou, E. Life-history traits of invasive fish in small Mediterranean streams. Biol. Invasions 2005, 7, 107–116. [Google Scholar] [CrossRef]
- Heger, T.; Pahl, A.T.; Botta-Dukát, Z.; Gherardi, F.; Hoppe, C.; Hoste, I.; Jax, K.; Lindstrom, L.; Boets, P.; Haider, S.; et al. Conceptual frameworks and methods for advancing invasion ecology. Ambio 2013, 42, 527–540. [Google Scholar] [CrossRef]
- Pheloung, P.C.; Williams, P.A.; Halloy, S.R. A weed risk assessment model for use as a biosecurity tool evaluating plant introductions. J. Environ. Manag. 1999, 57, 239–251. [Google Scholar] [CrossRef]
- Copp, G.H.; Garthwaite, R.; Gozlan, R.E. Risk identification and assessment of non-native freshwater fishes: A summary of concepts and perspectives on protocols for the UK. J. Appl. Ichthyol. 2005, 21, 371–373. [Google Scholar] [CrossRef]
- Copp, G.H.; Britton, J.R.; Jeney, G.; Joly, J.-P.; Gherardi, F.; Gollasch, S.; Gozlan, R.E.; Jones, G.; MacLeod, A.; Midtlyng, P.J.; et al. Risk Assessment Protocols and Decision Making Tools for Use of Alien Species in Aquaculture and Stock Enhancement. In IMPASSE—Environmental Impacts of Alien Species in Aquaculture for Coordination Action Priority FP6 2005-SSP-5A, Sustainable Management of Europe’s Natural Resources; Project No.: 044142; European Commission: Brussels, Belgium, 2008. [Google Scholar]
- Copp, G.H. The Fish Invasiveness Screening Kit (FISK) for non-native freshwater fishes—A summary of current applications. Risk Anal. 2013, 33, 1394–1396. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.L.; Hill, J.E.; Vilizzi, L.; Hardin, S.; Copp, G.H. Revisions of the Fish Invasiveness Screening Kit (FISK) for its application in warmer climatic zones, with particular reference to peninsular Florida. Risk Anal. 2013, 33, 1414–1431. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.; Ribeiro, F.; Leunda, P.M.; Vilizzi, L.; Copp, G.H. Effectiveness of FISK, an invasiveness screening tool for non-native freshwater fishes, to perform risk identification assessments in the Iberian Peninsula. Risk Anal. 2013, 33, 1404–1413. [Google Scholar] [CrossRef]
- Copp, G.H.; Vilizzi, L.; Tidbury, H.; Stebbing, P.; Tarkan, A.S.; Miossec, L.; Goulletquer, P. Development of a generic decision-support tool for identifying potentially invasive aquatic taxa: AS-ISK. Manag. Biol. Invasions 2016, 7, 343–350. [Google Scholar] [CrossRef]
- Glamuzina, B.; Tutman, P.; Nikolić, V.; Vidović, Z.; Pavličević, J.; Vilizzi, L.; Copp, G.H.; Simonović, P. Comparison of taxon-specific and taxon-generic risk screening tools to identify potentially invasive non-native fishes in the river Neretva catchment (Bosnia and Herzegovina and Croatia). River Res. Appl. 2017. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Sarı, H.M.; İlhan, A.; Kurtul, I.; Vilizzi, L. Risk screening of non-native and translocated freshwater fish species in a Mediterranean-type shallow lake: Lake Marmara (West Anatolia). Zool. Middle East 2016, 63, 48–57. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Vilizzi, L.; Top, N.; Ekmekçi, F.G.; Stebbing, P.D.; Copp, G.H. Identification of potentially invasive freshwater fishes, including translocated species, in Turkey using the Aquatic Species Invasiveness Screening Kit (AS-ISK). Int. Rev. Hydrobiol. 2017. [Google Scholar] [CrossRef]
- FishBase. Available online: www.fishbase.org (accessed on 19 December 2019).
- Regulamentação da Introdução na Natureza de Espécies não Indígenas em Portugal; Diário da República: Lisbon, Portuguesa, 1999. Decreto-Lei (nº 565/1999), I Séria-A, nº 295. Available online: https://data.dre.pt/eli/dec-lei/565/1999/12/21/p/dre/pt/html (accessed on 22 December 2019).
- Regulamentação da Introdução na Natureza de Espécies não Indígenas em Portugal; Diário da República: Lisbon, Portuguesa, 2019. Decreto-Lei (nº 92/2019), 1ª Séria, nº 130. Available online: https://data.dre.pt/eli/dec-lei/92/2019/07/10/p/dre (accessed on 22 December 2019).
- Rícan, O.; Kullander, S.O. Character- and tree-based delimitation of species in the ‘Cichlasoma’ facetum group (Teleostei, Cichlidae) with the description of a new genus. J. Zool. Syst. Evol. Res. 2006, 44, 136–152. [Google Scholar] [CrossRef]
- Carecho, J.; Baduy, F.; Guerreiro, P.M.; Saraiva, J.L.; Ribeiro, F.; Verissímo, A. Taxonomic re-evaluation of the non-native cichlid in Portuguese drainages. Fishes Mediterr. Environ. 2018, 1, 12. [Google Scholar] [CrossRef]
- Baduy, F.; Guerreiro, P.M.; Canário, A.V.; Saraiva, J.L. Social organization and endocrine profiles of Australoheros facetus, an exotic freshwater fish in southern Portugal. Acta Ethologica 2017. [Google Scholar] [CrossRef]
- Ribeiro, F.; Orjuela, R.L.; Magalhães, M.F.; Collares-Pereira, M.J. Variability in feeding ecology of a South American cichlid: A reason for successful invasion in mediterranean-type rivers? Ecol. Freshw. Fish 2008, 16, 559–569. [Google Scholar] [CrossRef]
- Ruiz, V.H.R.; Moyano, H.G.; Marchant, M.S.M. Aspectos biologicos del pez exotico Cichlasoma facetum (Jenyns, 1842) (Pisces, Cichlidae) en aguas dulces de Concepcion. Bol. Soc. Biol. Concepc. 1992, 63, 193–201. [Google Scholar]
- Leunda, P.M. Impacts of non-native fishes on Iberian freshwater ichthyofauna: Current knowledge and gaps. Aquat. Invasions 2010, 5, 239–262. [Google Scholar] [CrossRef]
- Sáez-Gómez, P.; Prenda, J. Updating the distribution data of recently introduced freshwater fish in the Guadalquivir River Basin (Spain). Bioinvasions Rec. 2019, 8. [Google Scholar] [CrossRef]
- BioDiversit4All. Available online: www.biodiversity4all.org (accessed on 28 October 2019).
- Pyke, G.H. Plague Minnow or Mosquito Fish? A Review of the Biology and Impacts of Introduced Gambusia Species. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 171–191. [Google Scholar] [CrossRef]
- Ruiz-Navarro, A.; Verdiell-Cubedo, D.; Torralva, M.; Oliva-Paterna, F.J. Removal control of the highly invasive fish Gambusia holbrooki and effects on its population biology: Learning by doing. Wildl. Res. 2013, 40, 82–89. [Google Scholar] [CrossRef]
- Cano-Rocabayera, O.; de Sostoa, A.; Coll, L.; Maceda-Veiga, A. Managing small, highly prolific invasive aquatic species: Exploring anecosystem approach for the eastern mosquitofish (Gambusia holbrooki). Sci. Total Environ. 2019, 673, 594–604. [Google Scholar] [CrossRef]
- Hermoso, V.; Clavero, M.; Blanco-Garrido, F.; Prenda, J. Invasive species and habitat degradation in Iberian streams: An analysis of their role in freshwater fish diversity loss. Ecol. Appl. 2011, 21, 175–188. [Google Scholar] [CrossRef]
- Ribeiro, F.; Chaves, M.L.; Marques, T.A.; Da Costa, L.M. First record of Ameiurus melas (Siluriformes, Ictaluridae) in the Alqueva reservoir, Guadiana basin (Portugal). Cybium 2006, 30, 283–284. [Google Scholar]
- Kodde, A.; Gkenas, C.; Cheoo, G.; Ribeiro, F.; Magalhães, M.F. Univited dinner guests: The effect of invasive fish and temperature on the foraging efficiency of Southern Iberian Chub. Fishes Mediterr. Environ. 2016, 11, 3. [Google Scholar]
- Ribeiro, F.; Collares-Pereira, M.J.; Moyle, P.B. Non-native fish in the fresh waters of Portugal, Azores and Madeira Islands: A growing threat to aquatic biodiversity. Fish. Manag. Ecol. 2009, 16, 255–2637. [Google Scholar] [CrossRef]
- Pires, A.M.; Da Costa, L.M.; Alves, M.J.; Coelho, M.M. Fish assemblage structure across the Arade basin (southern Portugal). Cybium 2004, 28, 357–365. [Google Scholar]
- Pires, D.F.; Pires, A.M.; Collares-Pereira, M.J.; Magalhaes, M.F. Variation in fish assemblages across dry-season pools in a Mediterranean stream: Effects of pool morphology, physicochemical factors and spatial context. Ecol. Freshw. Fish 2010, 19, 74–86. [Google Scholar] [CrossRef]
- Ribeiro, F.; Magalhães, M.F.; Collares-Pereira, M.J. Spatial and temporal variation in assemblage structure of fish larvae in mediterranean-type streams: Contrasts between native and non-native species. Environ. Biol. Fishes 2013, 96, 467–480. [Google Scholar] [CrossRef]
- García-Berthou, E. The characteristics of invasive fishes: What has been learned so far? J. Fish Biol. 2007, 71, 33–55. [Google Scholar] [CrossRef]
- Anastácio, P.M.; Ribeiro, F.; Capinha, C.; Banha, F.; Gama, M.; Filipe, A.F.; Rebelo, R.; Sousa, R. Non-native freshwater fauna in Portugal: A review. Sci. Total Environ. 2019, 650, 1923–1934. [Google Scholar] [CrossRef]
- Carveth, C.J.; Widmer, A.M.; Bonar, S.A. Comparison of upper thermal tolerances of native and non-native fish species in Arizona. Trans. Am. Fish. Soc. 2001, 135, 1433–1440. [Google Scholar] [CrossRef]
- Gutierre, S.M.M.; Schofield, P.J.; Prodocimo, V. Salinity and temperature tolerance of an emergent alien species, the Amazon fish Astronotus ocellatus. Hydrobiologia 2016, 777, 21–31. [Google Scholar] [CrossRef]
- Pires, A.M.; Cowx, I.G.; Coelho, M.M. Seasonal changes in fish community structure of intermittent streams in the middle reaches of the Guadiana basin, Portugal. J. Fish Biol. 1999, 54, 235–249. [Google Scholar] [CrossRef]
- Magalhaes, M.F.; Beja, P.; Canas, C.; Collares-Pereira, M.J. Functional heterogeneity of dry-season fish refugia across a Mediterranean catchment: The role of habitat and predation. Freshw. Biol. 2002, 47, 1919–1934. [Google Scholar] [CrossRef]
- Baduy, F.; Soares, J.; Silva, M.; Canario, A.V.M.; Saraiva, J.L.; Guerreiro, P.M. Critical thermal maximum and minimum in Australoheros facetus, a neotropical invader in the Iberian Peninsula. Fishes Mediterr. Environ. 2016, 12, 3. [Google Scholar] [CrossRef]
- Baduy, F.; Saraiva, J.L.; Vargas, M.; Silva, M.; Soares, J.; Canário, A.V.M.; Guerreiro, P.M. Plastic physiology, social behaviour and endocrine profiles of an invasive cichlid in Southern Portugal. Adv. Comp. Endocrinol. 2016. [Google Scholar] [CrossRef]
- Baduy, F.; Teixeira, D.; Soares, D.; Saraiva, J.L.; Canário, A.V.M.; Guerreiro, P.M. Dispersion of the invasive Australoheros facetus into upper estuaries may be deterred by salinity-related effects on both physiology and behaviour. In Proceedings of the VI Iberian Congress of Ichthyology, Murcia, Spain, 21 June 2016. [Google Scholar]
- Matono, P.; Bernardo, J.M.; Oberdorff, T.; Ilheu, M. Effects of natural hydrological variability on fish assemblages in small Mediterranean streams: Implications for ecological assessment. Ecol. Indic. 2012, 23, 467–481. [Google Scholar] [CrossRef]
- Alexandre, C.M.; Ferreira, T.F.; Almeida, P.R. Fish assemblages in non-regulated and regulated rivers from permanent and temporary Iberian systems. River Res. Appl. 2012. [Google Scholar] [CrossRef]
- Bernardo, J.M.; Ilheu, M.; Matono, P.; Costa, A.M. Interannual variation of fish assemblage structure in a Mediterranean River: Implications of streamflow on the dominance of native or exotic species. River Res. Appl. 2003, 19, 521–532. [Google Scholar] [CrossRef]
- Godinho, F.N.; Ferreira, M.T.; Cortes, R.V. Composition and spatial organization of fish assemblages in the lower Guadiana basin, southern Iberia. Ecol. Freshw. Fish 1997, 6, 134–143. [Google Scholar] [CrossRef]
- Ribeiro, F.; Collares-Pereira, M.J. Life-history variability of non-native centrarchids in regulated river systems of the lower River Guadiana drainage (south-west Iberian Peninsula). J. Fish Biol. 2010, 76, 522–537. [Google Scholar] [CrossRef]
- Ribeiro, F.; Elvira, B.; Collares-Pereira, M.J.; Moyle, P.B. Life-history traits of non-native fishes in Iberian watersheds across several invasion stages: A first approach. Biol. Invasions 2008, 10, 89–102. [Google Scholar] [CrossRef]
- Range, I.L. Applicability of Fish Risk Assessment (FISK) to Ornamental Species. Master’s Thesis, University of Lisbon, Lisbon, Portugal, 2013. [Google Scholar]
- Oliveira, J.M.; Segurado, P.; Santos, J.M.; Teixeira, A.; Ferreira, M.T.; Cortes, R.V. Modelling stream-fish functional traits in reference conditions: Regional and local environmental correlates. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Gasith, A.; Resh, V.H. Streams in Mediterranean climate regions: Abiotic influences and biotic responses to predictable seasonal events. Annu. Rev. Ecol. Evol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]
- Juette, T.; Cucherousset, J.; Cote, J. Animal personality and the ecological impacts of freshwater non-native species. Curr. Zool. 2014, 60, 417–427. [Google Scholar] [CrossRef]
- Marchetti, M.P.; Moyle, P.B.; Levine, R. Alien fishes in California watersheds: Characteristics of successful and failed invaders. Ecol. Appl. 2004, 14, 587–596. [Google Scholar] [CrossRef]
- Grabowska, J.; Przybylski, M. Life-history traits of non-native freshwater fish invaders differentiate them from natives in the Central European bioregion. Rev. Fish Biol. Fish. 2014, 25, 165–178. [Google Scholar] [CrossRef]
- Colgan, P.W.; Brown, J.A. Dynamics of nest defense by male centrarchid fish. Behav. Process. 1998, 17, 17–26. [Google Scholar] [CrossRef]
- Vilizzi, L.; Copp, G.H.; Adamovich, B.; Almeida, D.; Chan, J.; Dembski, S.; Koutsikos, N.; Magalhães, F.; Mendoza, R.; Onikura, N.; et al. A global review and meta-analysis of applications of the freshwater Fish Invasiveness Screening Kit. Rev. Fish Biol. Fish. 2019, 29, 529–568. [Google Scholar] [CrossRef]
- Britton, J.R.; Cucherousset, J.; Davies, G.D.; Godard, M.J.; Copp, G.H. Non-nativefishes and climate change: Predicting species responses towarming temperatures in a temperate region. Freshw. Biol. 2010, 55, 1130–1141. [Google Scholar] [CrossRef]
- Real Decreto 630/2013 del Reyno de España. Available online: https://www.boe.es/eli/es/rd/2013/08/02/630 (accessed on 1 November 2019).
- Define as Bases da Política de Ambiente; Diário da República Lisbon, Portuguesa; Lei (nº 19/2014), I Série, nº 73. Available online: https://data.dre.pt/eli/lei/19/2014/04/14/p/dre/pt/html (accessed on 22 December 2019).
- Hirsch, P.E.; N’Guyen, A.; Adrian-Kalchhauser, I.; Burkhardt-Holm, P. What do we really know about the impacts of one of the 100 worst invaders in Europe? A reality check. Ambio 2016, 45, 267–279. [Google Scholar] [CrossRef]
- Ribeiro, F.; Leunda, P.M. Non-native fish impacts on Mediterranean freshwater ecosystems: Current knowledge and research needs. Fish. Manag. Ecol. 2012, 19, 142–156. [Google Scholar] [CrossRef]
- Cefas—Centre for Environment, Fisheries and Aquaculture Science. Available online: www.cefas.co.uk (accessed on 5 February 2018).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YEAR | SITE | S | N | d | J’ | F | H’ | 1-λ | Af n% |
|---|---|---|---|---|---|---|---|---|---|
| 2014 | VAS | 9 | 321 | 1.39 | 0.54 | 1.72 | 1.20 | 0.59 | 0.57 |
| ODE | 8 | 588 | 1.10 | 0.38 | 1.31 | 0.79 | 0.34 | 0.81 | |
| 2015 | VAS | 7 | 448 | 0.98 | 0.58 | 1.18 | 1.14 | 0.55 | 0.64 |
| ODE | 6 | 633 | 0.78 | 0.46 | 0.92 | 0.83 | 0.39 | 0.77 | |
| 2017 | VAS | 11 | 249 | 1.81 | 0.70 | 2.36 | 1.69 | 0.79 | 0.24 |
| ODE | 6 | 1193 | 0.71 | 0.39 | 0.82 | 0.71 | 0.36 | 0.79 | |
| 2018 | VAS | 6 | 264 | 0.90 | 0.82 | 1.09 | 1.47 | 0.74 | 0.31 |
| ODE | 3 | 596 | 0.31 | 0.41 | 0.41 | 0.45 | 0.22 | 0.88 |
| Scores | FISK v2 | AS-ISK |
|---|---|---|
| Total score | 23 | 37 |
| Basic risk assessment (BRA) score | 23 | 27 |
| Climate-change assessment (CCA) score | - | 10 |
| Outcome | High | High |
| Score Partition | ||
| A. Biogeography/Historical | ||
| 1. Domestication/cultivation | 4 | 4 |
| 2. FISK v2: climate, distribution; AS-ISK: climate, distribution, and introduction risk | 2 | 1 |
| 3. Invasive elsewhere | 4 | 2 |
| B. Biology/Ecology | ||
| 4. FISK v2: undesirable traits; AS-ISK: undesirable or persistence traits | 5 | 6 |
| 5. FISK v2: feeding guild; AS-ISK: resource exploitation | 1 | 7 |
| 6. Reproduction | 2 | 1 |
| 7. Dispersal mechanisms | 4 | 3 |
| 8. FISK v2: persistence attributes; AS-ISK: tolerance attributes | 1 | 3 |
| C. Climate Change | ||
| 9. Climate change | - | 10 |
| Questions | ||
| Answered | 46 | 53 |
| Not applicable | 3 | 2 |
| A. Biogeography/Historical | ||
| 1. Domestication/cultivation | 3 | 3 |
| 2. FISK v2: climate, distribution; AS-ISK: climate, distribution, and introduction risk | 5 | 4 |
| 3. Invasive elsewhere | 5 | 4 |
| B. Biology/Ecology | ||
| 4. FISK v2: undesirable traits; AS-ISK: undesirable or persistence traits | 12 | 12 |
| 5. FISK v2: feeding guild; AS-ISK: resource exploitation | 1 | 2 |
| 6. Reproduction | 7 | 7 |
| 7. Dispersal mechanisms | 8 | 9 |
| 8. FISK v2: persistence attributes; AS-ISK: tolerance attributes | 5 | 6 |
| C. Climate Change | ||
| 9. Climate change | - | 6 |
| Sectors Affected | ||
| Aquaculture (FISK v2)/commercial (AS-ISK) | 18 | 8 |
| Environmental | 21 | 11 |
| Nuisance | 1 | 22 |
| Thresholds | ||
| Medium | 19 | 19 |
| High | 20 | 20 |
| Confidence | 0.78 | 0.78 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baduy, F.; Saraiva, J.L.; Ribeiro, F.; Canario, A.V.M.; Guerreiro, P.M. Distribution and Risk Assessment of Potential Invasiveness of Australoheros facetus (Jenyns, 1842) in Portugal. Fishes 2020, 5, 3. https://doi.org/10.3390/fishes5010003
Baduy F, Saraiva JL, Ribeiro F, Canario AVM, Guerreiro PM. Distribution and Risk Assessment of Potential Invasiveness of Australoheros facetus (Jenyns, 1842) in Portugal. Fishes. 2020; 5(1):3. https://doi.org/10.3390/fishes5010003
Chicago/Turabian StyleBaduy, Flavia, João L. Saraiva, Filipe Ribeiro, Adelino V. M. Canario, and Pedro M. Guerreiro. 2020. "Distribution and Risk Assessment of Potential Invasiveness of Australoheros facetus (Jenyns, 1842) in Portugal" Fishes 5, no. 1: 3. https://doi.org/10.3390/fishes5010003
APA StyleBaduy, F., Saraiva, J. L., Ribeiro, F., Canario, A. V. M., & Guerreiro, P. M. (2020). Distribution and Risk Assessment of Potential Invasiveness of Australoheros facetus (Jenyns, 1842) in Portugal. Fishes, 5(1), 3. https://doi.org/10.3390/fishes5010003