Consumption of Atlantic Salmon Smolt by Striped Bass: A Review of the Predator-Prey Encounter Literature and Implications for the Design of Effective Sampling Strategies

Abstract
1. Introduction
2. Materials and Methods
2.1. Striped Bass Prey Size Selection, Gape Size Limitations, and Smolt Vulnerability
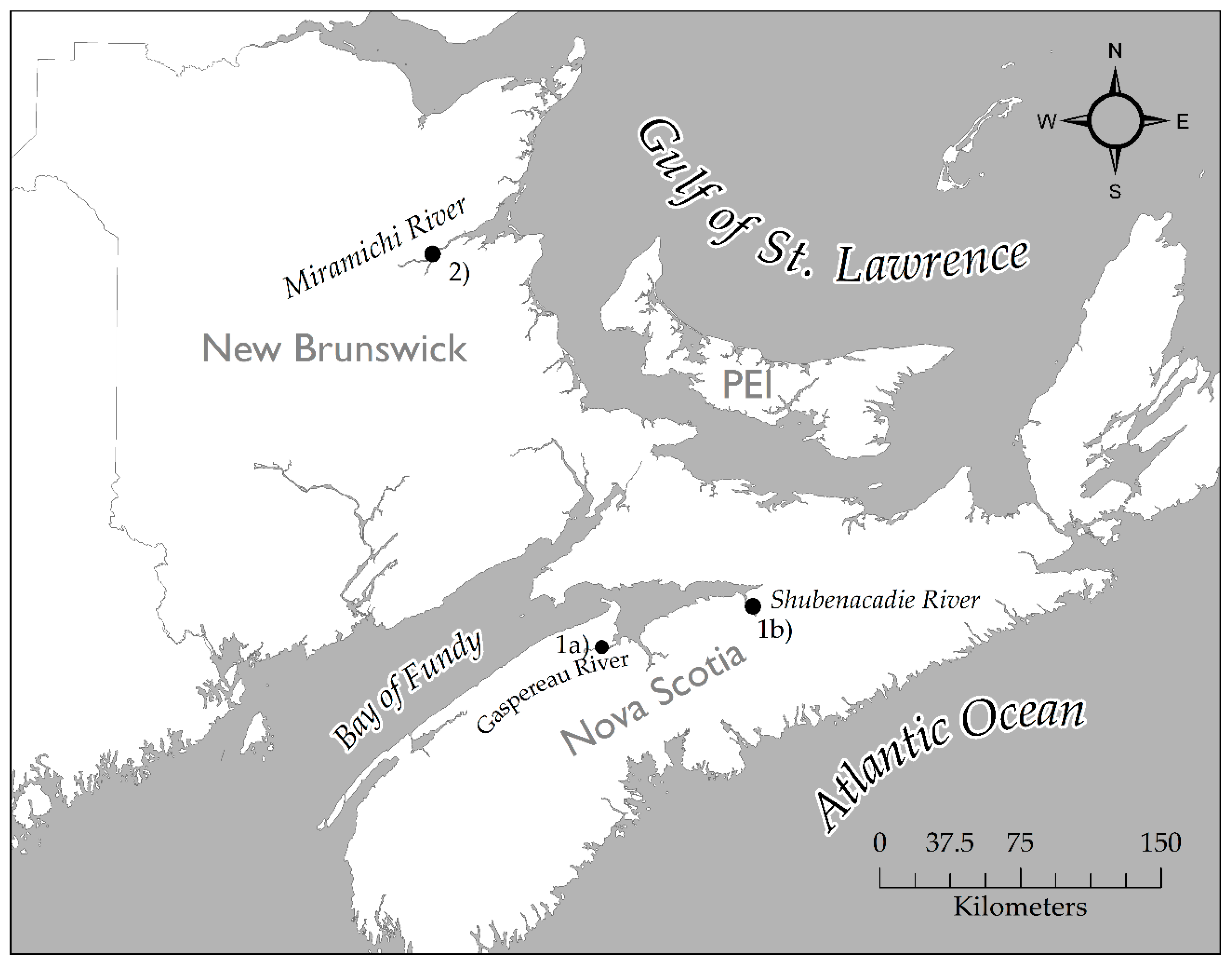
2.2. Feeding Behavior of Striped Bass During the Pre-Spawn, Spawn, and Post-Spawn Periods
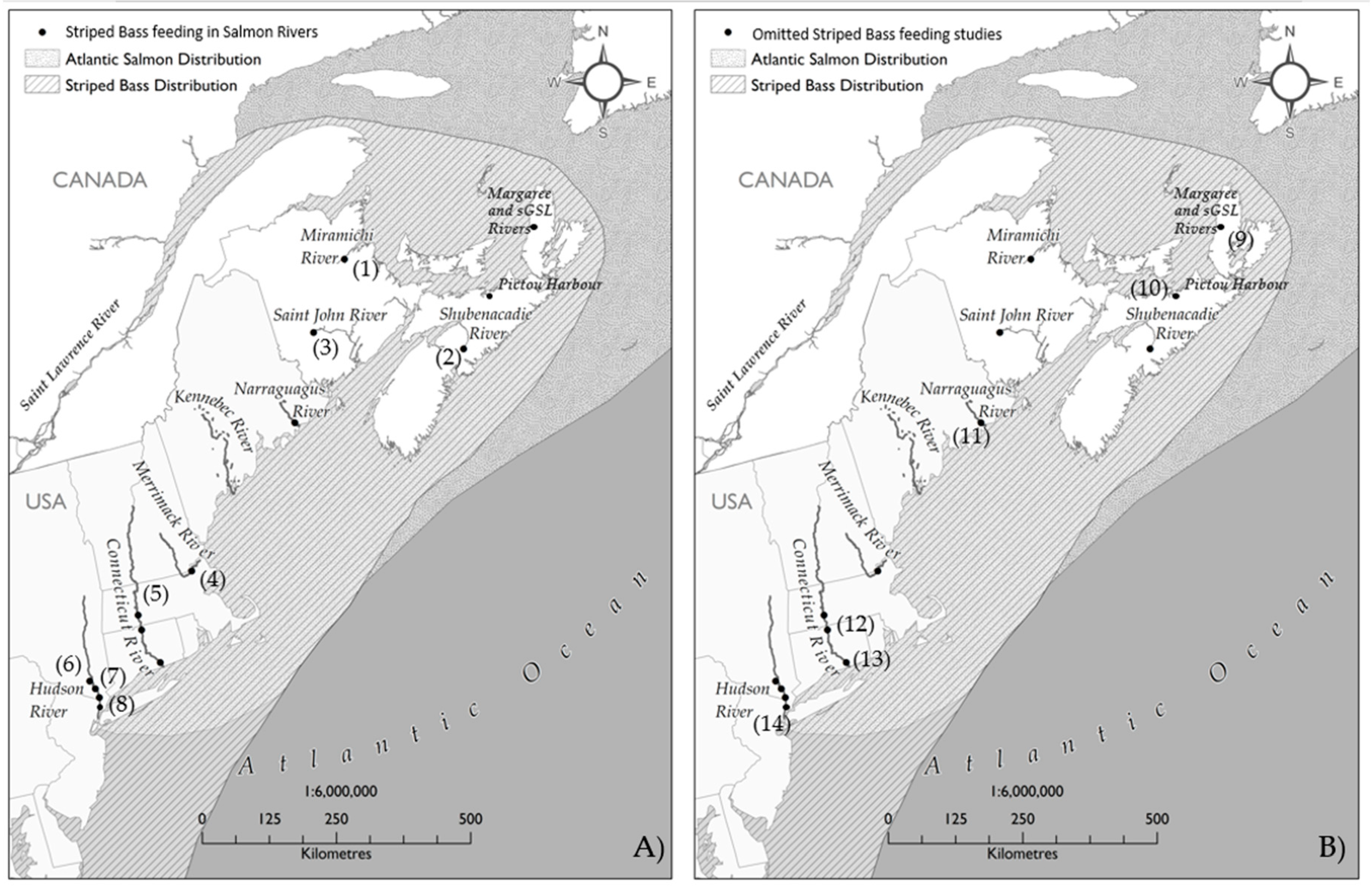
2.3. Documented Predation by Striped Bass on Atlantic Salmon Smolts
2.3.1. Blackwell and Juanes 1998: Predation on Atlantic Salmon Smolts by Striped Bass after Dam Passage [45]
2.3.2. Beland et al. 2001: Striped Bass Predation upon Atlantic Salmon Smolt in Maine [36]
2.3.3. DFO 2015: Spawner Abundance and Biological Characteristics of Striped Bass (Morone Saxatilis) in the Southern Gulf of St. Lawrence in 2015 [22]
2.4. Assessment of Smolt Predation by Acoustic Tagging
2.4.1. Gibson et al. 2015: Effects of Predation on Telemetry-based Estimates: Insights from a Study on Endangered Atlantic Salmon Smolts [80]
2.4.2. Daniels et al. 2018: Estimating Consumption Rate of Atlantic Salmon Smolts (Salmo salar) by Striped Bass (Morone saxatilis) in the Miramichi River Estuary Using Acoustic Telemetry [20]
2.4.3. Considerations for Acoustic Predation Assessment
3. Discussion
4. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
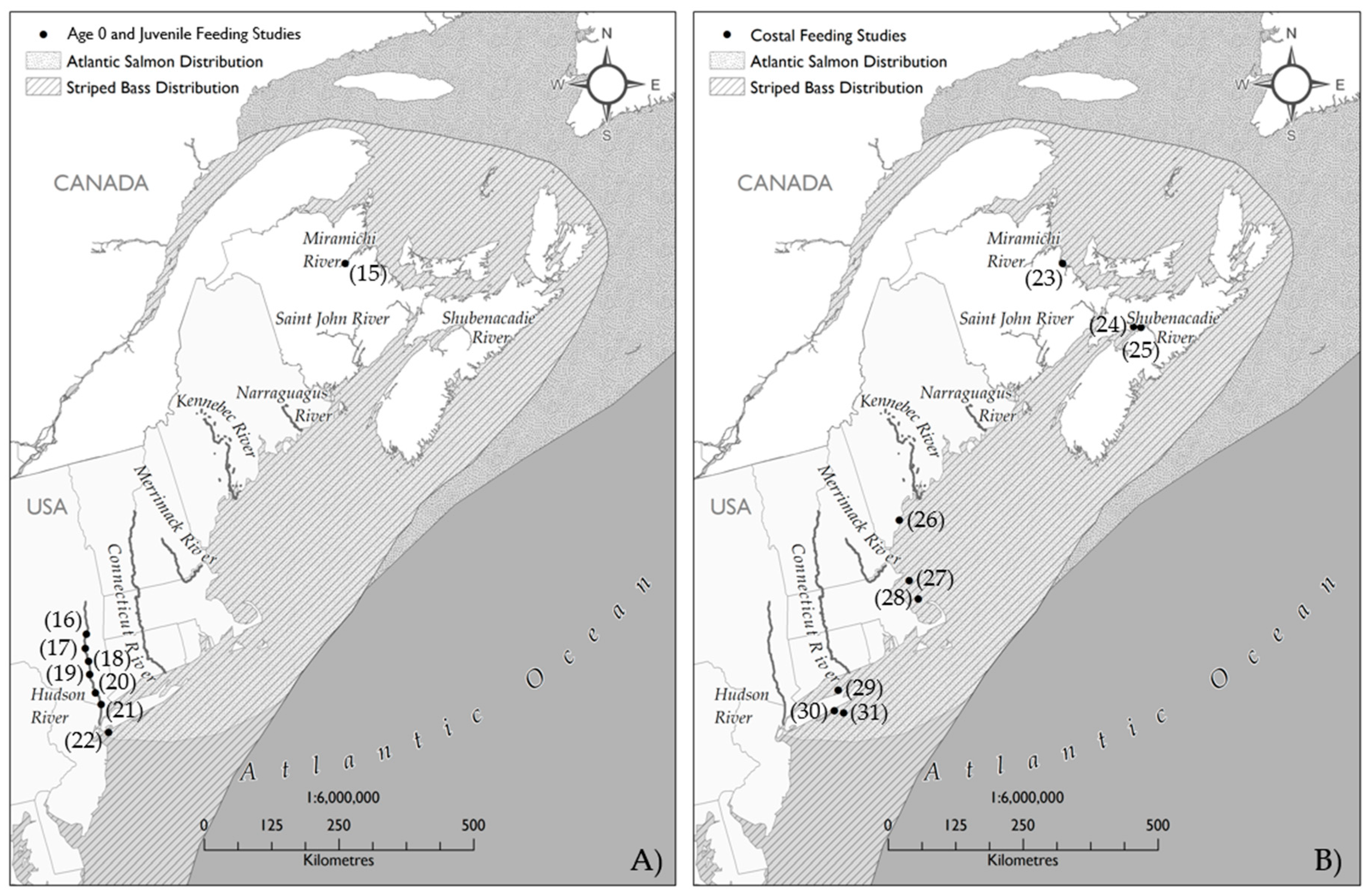
| Source | Map # | Location | Time of Year/Years | Size of Fish Sampled (mm)/Age | Stomachs Sampled | Full Stomachs | Proportion Full | Primary Food | |
|---|---|---|---|---|---|---|---|---|---|
| Age 0 and Juvenile Striped Bass Diet Sampling | |||||||||
| Robichaud-Leblanc et al. [94] | 15 | Miramichi River | June–November 1992 | 2.9–153.3 mm | 2928 | 2247 | 0.76 | Rotifers, Copepods, Mysids, Crangon | |
| Smircich et al. [95] | 16 | Hudson River (near 96 km) | Early June–mid-September 1988–2012 | 6–60 mm | 840 | 642 | 0.76 | Copepoda, Amphiods (Gammardiae, Corophiidae) | |
| Gardinier and Hoff [40] | 17 | Hudson River (53–74 km) | April–November 1974 | <76 mm | 501 | 422 | 0.82 | Amphipods (Gammarus), Copepods and Cladocerans | |
| - | - | - | - | 76–150 mm | - | - | Amphipods (Gammarus), Bay Anchovy, Atlantic Tomcod, Banded Killifish | ||
| - | - | - | - | 151–200 mm | - | - | Blueback Herring, Bay Anchovy, Atlantic Tomcod, Banded Killifish, Mummichog | ||
| - | - | - | - | 201–275 mm | 13* | - | Atlantic Tomcod, alosines, White Perch, Striped Bass | ||
| Jordan and Juanes [96] | 18 | Hudson River (35–70 km) | July–November 1994–1997 | <50 to >100 mm | 695 | 576 | 0.82 | Amphipods | |
| Howe and Juanes [97] | 19 | Hudson River (32–72 km) | Mid-July to early November 1998 | 57.3 ± 10.9 mm | 254 | 231 | 0.91 | Amphipods | |
| Jordan et al. [98] | 20 | Hudson River (35–70 km) | July–November 1994–1997 | <50 to >100 mm | 950 ** | 798 | 0.84 | Amphipods | |
| Curran and Ries [99] | 21 | Hudson River (Dennings Point) | 23 July–13 August | 30–110 mm | 105 | - | Amphipods (Gammarus), Chronomids and Copepods | ||
| Buckel and Mckown [100] | 22 | New York Bight | May–November 1997–1998 | 39–296 mm | 602 | 439 | 0.72 | Sand Shrimp (sp.) | |
| Total | 2.9–296 mm | 6193 | 4779 | 0.77 | 77% full stomachs | ||||
| Marine Striped Bass Diet Sampling | |||||||||
| Melvin [101] | 23 | Kouchibuguac River | 15 June–1 November 1978 | 27.4–54.6 cm | 255 | 179 | 0.70 | Sand Shrimp, Fundulus | |
| Rulifson and McKenna [102] | 24 | Minas Basin, Cobequid Bay | 1 June–18 October 1985 | 69–94 mm | 80 | 78 | 0.98 | Crangon septemspinosa | |
| Liem [103] | 25 | Bass River, Nova Scotia | NA | ~20 cm | NA | NA | Crangon | ||
| Wilkinson [104] | 26 | Saco Bay, Maine | May–October 2011 | 43.4–59.9 cm | 57 | 43 | 0.75 | Sand Lance | |
| - | - | - | - | 60.0–74.9 cm | - | - | Sand Lance | ||
| - | - | - | - | 75.0–109.2 cm | - | - | Sand Lance, American lobster | ||
| Ferry and Mather [67] | 27 | Massachusetts estuaries | May–October 1999 | 37.5–47.5 cm | 797 | 677 | 0.85 | Alosines | |
| Nelson et al. [105] | 28 | Coastal Massachusetts | June–September 1997–2000 | 29.0–116.2 cm | 3006 | 1720 | 0.57 | Crustaceans and Atlantic Menhaden | |
| Sagarese et al. [106] | 29 | Long Island, New York | May–October 2007–2008 | 39.6–95.9 cm | 23 | 15 | 0.65 | Sand Shrimp, Summer Flounder | |
| Schaefer et al. [75] | 30 | Long Island, New York | 27 April–24 November 1964 | 27.5–39.9 cm | 61 | 49 | 0.80 | Amphipods and mysids | |
| - | - | - | - | 40.0–59.9 cm | 183 | 145 | 0.79 | Amphipods and Bay Anchovy | |
| - | - | - | - | 60.0–94.0 cm | 123 | 28 | 0.22 | Amphipods, Bay Anchovy, and Squirrel Hake (Urophycis chuss) | |
| - | - | - | - | 14.1–24.0 cm | - | - | Crangon septemspinosa | ||
| - | - | - | - | 27.1–36.0 cm | - | - | Crangon septemspinos, Unidentified fish | ||
| - | - | - | - | 38.1–44.0 cm | - | - | Crangon septemspinos, Unidentified fish | ||
| - | - | - | - | 48.1–52.0 cm | - | - | Unidentified fish | ||
| Merriman [107] 1941 | 31 | New Jersey, Massachusetts | April–November 1936 and 1937 | 6.5–115 cm | 550 | 264 | 0.48 | Atlantic Menhaden, Atlantic Silverside | |
| Total | 6.5–116.2 cm | 5135 | 3198 | 0.62 | 62% full stomachs | ||||

References
- MacCrimmon, H.R.; Gots, B.L. World distribution of Atlantic Salmon, Salmo Salar. J. Fish. Res. Board Can. 1979, 36, 422–457. [Google Scholar] [CrossRef]
- Scott, W.B.; Scott, M.G. Family Percichthyidae/Temperate Basses. In Atlantic Fishes of Canada; Canadian Bulletin of Fisheries and Aquatic Sciences 219; University of Toronto Press: Toronto, ON, Canada, 1988; pp. 356–360. [Google Scholar]
- Tremblay, S.; Caron, F.; Groleau, C.; Deschamps, D. Bilan de L’exploitation du Saumon au Québec en 1998; Faune et Parcs Québec: Québec, QC, Canada, 1999. [Google Scholar]
- World Wildlife Federation (WWF). The Status of Wild Atlantic Salmon: A River by River Assessment; WWF: Washington, DC, USA, 2001. [Google Scholar]
- Rulifson, R.A.; Dadswell, M.J. Life history and population characteristics of Striped Bass in Atlantic Canada. Trans. Am. Fish. Soc. 1995, 124, 477–507. [Google Scholar] [CrossRef]
- Greene, K.E.; Zimmerman, J.L.; Laney, R.W.; Thomas-Blate, J.C. Atlantic Coast Diadromous Fish Habitat: A Review of Utilization, Threats, Recommendations for Conservation, and Research Needs; Atlantic States Marine Fisheries Commission: Washington, DC, USA, 2009. [Google Scholar]
- Leblanc, N.M.; Andrews, S.N.; Avery, T.S.; Puncher, G.N.; Gahagan, B.I.; Whiteley, A.R.; Curry, R.A.; Pavey, S.A. Evidence of a Genetically Distinct population of Striped Bass in the Saint John River, New Brunswick, Canada. N. Am. J. Fish. Manag. 2018, 38, 1339–1349. [Google Scholar] [CrossRef]
- Stolte, L. The Forgotten Salmon of the Merrimack; Department of the Interior: Washington, DC, USA, 1981.
- Stolte, L.W. Atlantic Salmon restoration in the Merrimack River Basin. In A Hard Look at Some Tough Issues: New England Atlantic Salmon Management Conference, Proceedings of the New England Atlantic Salmon Management Conference, Danvers, MA, USA, 22–23 April 1994; Calibi, S., Stout, A., Eds.; New England Salmon Association: Newburyport, MA, USA, 1994. [Google Scholar]
- Moring, J.R. Restoration of Atlantic Salmon to Maine: Overcoming physical and biological problems in the estuary. In Proceedings of the Coastal Zone ‘87: Fifth Symposium on Coastal and Ocean Management, Seattle, WA, USA, 26–29 May 1987; Magoon, O.T., Converse, H., Miner, D., Eds.; American Society of Civil Engineers: Seattle, WA, USA, 1987; pp. 3129–3140. [Google Scholar]
- Meyers, T. The program to restore Atlantic salmon in the Connecticut River. In A Hard Look at Some Tough Issues: New England Atlantic Salmon Management Conference, Proceedings of the New England Atlantic Salmon Management Conference, Danvers, MA, USA, 22–23 April 1994; Calibi, S., Stout, A., Eds.; New England Salmon Association: Newburyport, MA, USA, 1994; pp. 11–21. [Google Scholar]
- USASAC. Annual Report of the U. S. Atlantic Salmon Assessment Committee; Report No. 1–29; USASAC: Huntsville, AL, USA, 1989–2017.
- Izzo, L.K.; Maynard, G.A.; Zydlewski, J. Upstream Movements of Atlantic Salmon in the lower Penobscot River, Maine Following Two Dam removals and Fish Passage Modifications. Mar. Coast. Fish. 2016, 8, 448–461. [Google Scholar] [CrossRef]
- ICES. Report of the Working Group for North Atlantic Salmon (WGNAS); International Council for the Exploration of the Sea. Advisory Committee on Fisheries Management: Woods Hole, MA, USA, 2018. [Google Scholar]
- COSEWIC. COSEWIC Assessment and Status Report on the Atlantic Salmon Salmo Salar (Nunavik Population, Labrador Population, Northeast Newfoundland Population, South Newfoundland Population, Southwest Newfoundland Population, Northwest Newfoundland Population, Quebec Eastern North Shore Population, Quebec Western North Shore Population, Anticosti Island Population, Inner St. Lawrence Population, Lake Ontario Population, Gaspé-Southern Gulf of St. Lawrence Population, Eastern Cape Breton Population, Nova Scotia Southern Upland Population, Inner Bay of Fundy Population, Outer Bay of Fundy Population) in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2010. [Google Scholar]
- Fay, C.; Bartron, M.; Craig, S.; Hecht, A.; Pruden, J.; Saunders, R.; Sheehan, T.; Trial, J. Status Review for Anadromous Atlantic SALMON (Salmo Salar) in the United States; United States Fish and Wildlife Service: Washington, DC, USA, 2006.
- DFO. Update of Indicators of Atlantic Salmon (Salmo Salar) in DFO Gulf Region Salmon Fishing Areas 15-18 for 2016; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2017.
- McGinnity, P.; Prodöhl, P.; Ferguson, A.; Hynes, R.; Maoiléidigh, N.Ó.; Baker, N.; Cotter, D.; O’Hea, B.; Cooke, D.; Rogan, G.; et al. Fitness reduction and potential extinction of wild populations of Atlantic Salmon, Salmo salar, as a result of interactions with escaped farm salmon. Proc. R. Soc. Biol. Sci. 2003, 270, 2443–2450. [Google Scholar] [CrossRef]
- Breau, C.; Cunjak, R.A.; Peake, S.J. Behaviour during elevated water temperatures: Can physiology explain movement of juvenile Atlantic salmon to cool water? J. Anim. Ecol. 2011, 80, 844–853. [Google Scholar] [CrossRef]
- Daniels, J.M.; Chaput, G.J.; Carr, J. Estimating consumption rate of Atlantic Salmon smolts (Salmo salar) by Striped Bass (Morone saxatilis) in the Miramichi River estuary using acoustic telemetry. Can. J. Fish. Aquat. Sci. 2018, 75, 1811–1822. [Google Scholar] [CrossRef]
- Chaput, G.; Douglas, S.G.; Hayward, J. Biological Characteristics and Population Dynamics of Atlantic Salmon (Salmo Salar) from the Miramichi River, New Brunswick, Canada; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2016.
- DFO. Spawner Abundance and Biological Characteristics of Striped Bass (Morone Saxatilis) in the Southern Gulf of St. Lawrence in 2015; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2016.
- Boreman, J.; Lewis, R.R. Atlantic coastal migration of striped bass. In Common Strategies of Anadromous and Catadromous Fishes; Dadswell, M.J., Klauda, J.R., Moffit, C.M., Rulifson, R.A., Saunders, R.L., Eds.; American Fisheries Society Symposium: Bethesda, MD, USA, 1987; pp. 331–339. [Google Scholar]
- Andrews, S.N.; Dadswell, M.J.; Buhariwalla, C.F.; Linnansaari, T.; Curry, R.A. Looking for Striped Bass in Atlantic Canada: The Reconciliation of Local, Scientific, and Historical Knowledge. Natl. Emerg. Number Assoc. 2019, 26, 1–30. [Google Scholar] [CrossRef]
- Setzler, E.M.; Boynton, W.R.; Wood, K.V.; Zion, H.H.; Lubbers, L.; Mountford, N.K.; Frere, P.; Tucker, L.; Mihursky, J.A. Synopsis of Biological Data on Striped Bass, Morone Saxatilis (Walbaum); NOAA Technical Report, NMFS Circular 433: Seattle, WA, USA, 1980.
- Field, J.D. Atlantic Striped Bass management: Where did we go right? Fisheries 1997, 22, 6–8. [Google Scholar]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Coutant, C.C. Temperature-oxygen habitat for freshwater and coastal Striped Bass in a changing climate. Trans. Am. Fish. Soc. 1990, 119, 240–253. [Google Scholar] [CrossRef]
- NunatuKavut Community Council (NCC). NunatuKavut Annual Report 2016–2017; NunatuKavut Community Council: Happy Valley-Goose Bay, NL, Canada, 2017. [Google Scholar]
- DFO. Update of Indicators to 2017 of Adult Atlantic Salmon for the Miramichi River (NB), Salmon Fishing Area 16, DFO Gulf Region; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2018.
- Bradford, R.G.; Chaput, G.; Douglas, S.; Hayward, J. Status of Striped Bass (Morone saxatilis) in the Gulf of St. Lawrence in 1999; Canadian Stock Assessment Secretariat Research Document 2001/006; Department of Fisheries and Oceans: Ottawa, ON, Canada, 2001.
- DFO. Spawner Abundance and Biological Characteristics of Striped Bass (Morone Saxatilis) in the Southern Gulf of St. Lawrence in 2017; DFO Canadian Science Advisory Secretariat Science Response 2018/016; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2018.
- Manooch, C.S. Food Habits and yearling and Adult Striped Bass, Morone saxatilis Walbaum, from Albemarle Sound, North Carolina. Cheasapeake Sci. 1973, 14, 73–86. [Google Scholar] [CrossRef]
- Chaput, G.; Hardie, P.; Hayward, J.; Moore, D.; Shaesgreen, J.; Northumberland Salmon Protection Association. Migrations and Biological Characteristics of Atlantic Salmon (Salmo Salar) Smolts from the Northwest Miramichi River, 1998 to 2000; Canadian Technical Report of Fisheries and Aquatic Science 2415; Department of Fisheries and Oceans: Ottawa, ON, Canada, 2002.
- Schulze, M.B. Using a Field Survey to Assess Potential Temporal and Spatial Overlap between Piscivores and Their Prey, and a Bioenergetics Model to Examine Potential Consumption of Prey, Especially Juvenile Anadromous Fish, in the Connecticut River Estuary. Master’s Thesis, University of Massachusetts, Amherst, MA, USA, 1996. [Google Scholar]
- Beland, F.B.; Kocik, J.F.; VandeSande, J.; Sheehan, T.F. Striped Bass predation upon Atlantic Salmon smolt in Maine. Natl. Emerg. Number Assoc. 2001, 8, 267–274. [Google Scholar]
- Davis, J.P.; Schultz, E.T.; Vokoun, J.C. Striped Bass consumption of Blueback Herring during vernal riverine migrations: Does relaxing harvest restrictions on predators help conserve a prey species of concern? Mar. Coast. Fish. 2012, 4, 239–251. [Google Scholar] [CrossRef]
- Dunning, D.J.; Waldman, J.R.; Ross, Q.E.; Mattson, M.T. Use of Atlantic Tomcod and other prey by Striped Bass in the lower Hudson River estuary during winter. Trans. Am. Fish. Soc. 1997, 126, 857–861. [Google Scholar] [CrossRef]
- Buhariwalla, C.F.; MacMillan, J.L.; Gregoire, M.J.; Dadswell, M.J.; Stokesbury, M.J.W. Population characteristics of Striped Bass killed by cold shock during winter shutdown of a power plant in Nova Scotia. Natl. Emerg. Number Assoc. 2016, 23, 163–173. [Google Scholar] [CrossRef]
- Gardinier, M.N.; Hoff, T.B. Diet of Striped Bass in the Hudson River Estuary. N. Y. Fish Game J. 1982, 29, 152–165. [Google Scholar]
- Hyslop, E.J. Stomach analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Baker, R.; Buckland, A.; Sheaves, M. Fish gut content analysis: Robust measures of diet composition. Fish Fish. 2014, 15, 170–177. [Google Scholar] [CrossRef]
- Davidson, V.M. Fish (Salmon) Management; Fisheries Research Board of Canada No. 427, Manuscript Reports of the Biological Stations; Fisheries Research Board of Canada: Toronto, ON, Canada, 1950. [Google Scholar]
- Andrews, S.N.; Zelman, K.; Ellis, T.; Linnansaari, T.; Curry, R.A. Diet of Striped Bass and Muskellunge downstream of a large hydroelectric dam: A preliminary investigation into suspected Atlantic Salmon Smolt predation. N. Am. J. Fish. Manag. 2018, 38, 734–746. [Google Scholar] [CrossRef]
- Blackwell, F.B.; Juanes, F. Predation on Atlantic Salmon smolts by Striped Bass after dam passage. N. Am. J. Fish. Manag. 1998, 18, 936–939. [Google Scholar] [CrossRef]
- Warner, J.; Kynard, B. Scavenger Feeding by Subadult Striped Bass, Morone saxatilis below a Low-Head hydroelectric Dam. Fish. Bull. 1986, 84, 220–222. [Google Scholar]
- Kahnle, A.W.; Hattala, K.A. Striped Bass Predation on Adult American Shad: Occurrence and Observed Effects on American Shad Abundance in Atlantic Coastal Rivers and Estuaries; Supplement to American Shad Stock Assessment Report for Peer Review; Atlantic States Marine Fisheries Commission: Washington, DC, USA, 2007; Volume 1, pp. 182–220. [Google Scholar]
- Dew, C.B. Stomach contents of commercially caught Hudson River Striped Bass, Morone saxatilis, 1973–1975. Fish. Biol. NOAA 1988, 86, 397–401. [Google Scholar]
- Juanes, F. A length-based approach to predator-prey relationships in marine predators. Can. J. Fish. Aquat. Sci. 2016, 73, 677–684. [Google Scholar] [CrossRef]
- Mihalitsis, M.; Bellwood, D.R. A morphological and functional basis for maximum prey size in piscivorous fishes. PLoS ONE 2017, 12, e0184679. [Google Scholar] [CrossRef] [PubMed]
- Rudershausen, P.J.; Tuomikoski, J.E.; Buckel, J.A.; Hightower, J.E. Prey Selectivity and Diet of Striped Bass in western Albermarle Sound, North Carolina. Trans. Am. Fish. Soc. 2005, 134, 1059–1074. [Google Scholar] [CrossRef]
- Chaput, G.; Douglas, S.G.; Hayward, J. Biological Characteristics and Population Dynamics of Atlantic Salmon (Salmo salar) from the Miramichi River, New Brunswick, Canada; Canadian Science Advisory Secretariat Research Document 2016/029; Department of Fisheries and Oceans: Gulf Region, Ottawa, ON, Canada, 2016.
- Robichaud-Leblanc, K.A.; Courtenay, S.C.; Locke, A. Spawning and early life history of a northern population of Striped Bass (Morone saxatilis) in the Miramichi River estuary, Gulf of St. Lawrence. Can. J. Zoo 1996, 74, 1645–1655. [Google Scholar] [CrossRef]
- McCoy, E.G. Quantitative Sampling of Striped Bass, Roccus Saxatilis (Walbaum), Eggs in the Roanoke River. Master’s Thesis, North Carolina State University, Raleigh North, NC, USA, 1959. [Google Scholar]
- Talbot, G.B. Estuarine Environmental Requirements and Limiting Factors for Striped Bass; Bureau of Sport Fisheries and Wildlife: United States Department of the Interior, Tiburon, CA, USA, 1966. [Google Scholar]
- Rulifson, R.A.; Tull, K.A. Striped Bass spawning in a tidal bore river: The Shubenacadie Estuary, Atlantic Canada. Trans. Am. Fish. Soc. 1999, 128, 613–624. [Google Scholar] [CrossRef]
- Douglas, S.G.; Chaput, G.; Hayward, J.; Sheasgreen, J. Pre-spawning, spawning, and post-spawning behavior of Striped Bass in the Miramichi River. Trans. Am. Fish. Soc. 2009, 138, 121–134. [Google Scholar] [CrossRef]
- Walters, J.F., III; Austin, H.M. Diet composition of large Striped Bass (Morone saxatilis) in Chesapeake Bay. Fish. Bull. 2003, 101, 414–423. [Google Scholar]
- DFO. Assessment of the Habitat Quality and Habitat Use by the Striped Bass Population of the St. Lawrence Estuary, Quebec; Canadian Science Advisory Secretariat Science Advisory Report 2010/069; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2011.
- Trent, L.; Hassler, W.W. Feeding behavior of adult Striped Bass, Roccus saxatilis, in relation to stages of sexual maturity. Chesapeake Sci. 1966, 7, 189–192. [Google Scholar] [CrossRef]
- Scofield, E.C. The Striped Bass of California (Roccus Lineatus); Division of Fish and Game of California Fisheries, Bulletin No. 29: Sacramento, CA, USA, 1931. [Google Scholar]
- Hollis, E.H. Variations in the feeding habits of the Striped Bass, Roccus saxatilis (Walbaum), in the Chesapeake Bay. Bull. Bingham Oceanogr. Collect. 1952, 14, 111–131. [Google Scholar]
- Stevens, D.E. Food habits of Striped Bass, Roccus saxatilis, in the Sacramento-San Joaquin Delta. In Ecological Studies of the Sacramento-San Joaquin Delta, Part II: Fishes of the Delta; Turner, J.L., Kelly, D.W., Eds.; California Department of Fish and Game, Fisheries Bulletin: Sacramento, CA, USA, 1966; Volume 136, pp. 68–96. [Google Scholar]
- Raney, E.C. The life history of the Striped Bass, Roccus saxatilis (Walbaum). Bull. Bingham Oceanogr. Collect. 1952, 14, 1–97. [Google Scholar]
- Morgan, A.R.; Gerlach, A.R. Striped Bass Studies on Coos Bay, Oregon in 1949 and 1950: A Report to the Forty-Dixth Legislature; Oregon Fish Commission: Portland, OR, USA, 1950. [Google Scholar]
- McLaren, J.B.; Cooper, J.C.; Hoff, T.B.; Lander, V. Movements of Hudson River Striped Bass. Trans. Am. Fish. Soc. 1981, 110, 158–167. [Google Scholar] [CrossRef]
- Ferry, K.H.; Mather, M.E. Spatial and temporal diet patterns of subadult and small adult Striped Bass in Massachusetts estuaries: Data, a synthesis, and trends across scales. Mar. Coast. Fish. 2012, 4, 30–45. [Google Scholar] [CrossRef]
- Beukers-Stewart, B.D.; Jones, G.P. The influence of prey abundance on the feeding ecology of two piscivorous species of coral reef fish. J. Exp. Mar. Biol. Ecol. 2004, 299, 155–184. [Google Scholar] [CrossRef]
- Stevens, R.E. Landlocked Striped Bass. Annu. Northeast Fish Wildl. Conf. 1969, 25, 2–11. [Google Scholar]
- Mather, M.E.; Finn, J.T.; Ferry, K.H.; Deegan, L.A.; Nelson, G.A. Use of non-natal estuaries by migratory Striped Bass (Morone saxatilis) in summer. Fish. Bull. 2009, 107, 329–333. [Google Scholar]
- Walter, J.F., III; Overton, A.S.; Ferry, K.H.; Mather, M.E. Atlantic coast feeding habits of Striped Bass: A synthesis supporting a coast-wide understanding of trophic biology. Fish. Manag. Ecol. 2003, 10, 349–360. [Google Scholar] [CrossRef]
- Grossman, G.D.; Essington, T.; Johnson, B.; Miller, J.; Monsen, N.E.; Pearsons, T.N. Effects of Fish Predation on Salmonids in the Sacramento River-San Joaquin Delta and Associated Ecosystems; Delta Stewardship Council: Sacramento, CA, USA, 2013. [Google Scholar]
- Sabal, M.; Hayes, S.; Merz, J.; Setka, J. Habitat alterations and a non-native predator, the Striped Bass, increase native Chinook salmon mortality in the Central Valley, California. N. Am. J. Fish. Manag. 2016, 36, 309–320. [Google Scholar] [CrossRef]
- Schultz, A.A.; Kumagai, K.K.; Bridges, B.B. Methods to evaluate gut evacuation rates and predation using acoustic telemetry in the Tracy Fish Collection Facility primary channel. Anim. Biotelem. 2015, 3, 13. [Google Scholar] [CrossRef]
- Schaefer, R.H. Feeding habits of Striped Bass from the surf waters of Long Island. N. Y. Fish Game J. 1970, 17, 1–17. [Google Scholar]
- Jobling, M.; Breiby, A. The use and abuse of fish otoliths in studies of feeding habits of marine piscivores. Sarsia 1986, 71, 265–274. [Google Scholar] [CrossRef]
- Campana, S.E. Photographic Atlas of Fish Otoliths of the Northwest Atlantic Ocean; NRC Research Press: Ottawa, ON, Canada, 2004. [Google Scholar]
- Altin, A.; Ayyildiz, H. Relationships between total length and otolith measurements for 36 fish species from Gökçeada Island, Turkey. J. Appl. Ichthyol. 2017, 34, 136–141. [Google Scholar] [CrossRef]
- González, A.F.; López, A.; Guerra, A.; Barreiro, A. Diets of marine mammals stranded on the northwestern Spanish Atlantic coast with special reference to Cephalopoda. Fish. Res. 1994, 21, 179–191. [Google Scholar] [CrossRef]
- Gibson, J.F.; Halfyard, E.A.; Bradford, R.G.; Stokesbury, M.J.W.; Redden, A.M. Effects of predation on telemetry-based estimates: Insights from a study on endangered Atlantic Salmon smolts. Can. J. Fish. Aquat. Sci. 2015, 72, 728–741. [Google Scholar] [CrossRef]
- Bradford, R.G.; Halfyard, E.A.; Hayman, T.; Leblanc, P. Overview of 2013 Bay of Fundy Striped Bass Biology and General Status; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2015.
- Lacroix, G.L.; Knox, D.; McCurdy, P. Effects of implanted dummy acoustic transmitters on juvenile Atlantic Salmon. Trans. Am. Fish. Soc. 2004, 133, 211–220. [Google Scholar] [CrossRef]
- Wagner, G.N.; Cooke, S.J.; Brown, R.S.; Deters, K.A. Surgical implantation techniques for electronic tags in fish. Rev. Fish Biol. Fish. 2011, 21, 71–81. [Google Scholar] [CrossRef]
- Collins, A.L.; Scott, S.G.; Welch, D.W.; Cooke, S.J.; Clarke, T.D. Intracoelomic acoustic tagging of juvenile sockeye salmon: Swimming performance, survival, and postsurgical wound healing in freshwater and during a transition to seawater. Trans. Am. Fish. Soc. 2013, 142, 515–523. [Google Scholar] [CrossRef]
- Adams, N.S.; Rondorf, D.W.; Evans, S.D.; Kelly, J.E.; Perry, R.W. Effects of surgical and gastrically implanted radio transmitters on swimming performance and predator avoidance of juvenile Chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 1998, 55, 781–787. [Google Scholar] [CrossRef]
- Voegeli, F.A.; Lacroix, G.L.; Anderson, J.M. Development of miniature pingers for tracking Atlantic Salmon smolts at sea. Hydrobiologica 1998, 371, 35–46. [Google Scholar] [CrossRef]
- Melnychuk, M.C. Detection efficiency in telemetry studies: Definitions and evaluation methods. In Telemetry Techniques: A User Guide for Fisheries Research; Adams, N.S., Beeman, J.W., Eiler, J.H., Eds.; American Fisheries Society: Bethesda, MD, USA, 2012; pp. 339–357. [Google Scholar]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Evol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities-the roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar]
- Collie, J.S.; Botsford, L.W.; Hastings, A.; Kaplan, I.C.; Largier, J.L.; Livingston, P.A.; Plagányi, É.; Rose, K.A.; Wells, B.K.; Werner, F.E. Ecosystem models for fisheries management: Finding the sweet spot. Fish Fish. 2016, 17, 101–125. [Google Scholar] [CrossRef]
- Walters, C.; Christensen, V.; Fulton, B.; Smith, A.D.; Hilborn, R. Predictions from simple predator-prey theory about impacts of harvesting forage fishes. Ecol. Model. 2016, 337, 272–280. [Google Scholar] [CrossRef]
- Gjelland, K.Ø.; Hedger, R.D. Environmental influence on transmitter detection probability in biotelemetry: Developing a general model of acoustic transmission. Multistate Essay Exam. 2013, 4, 665–674. [Google Scholar] [CrossRef]
- Kessel, S.T.; Cooke, S.J.; Heupel, M.R.; Hussey, N.E.; Simpfendorfer, C.A.; Vagle, S.; Fisk, A.T. A review of detection range testing in aquatic passive acoustic telemetry studies. Rev. Fish Biol. Fish. 2014, 24, 199–218. [Google Scholar] [CrossRef]
- Robichaud-LeBlanc, K.A.; Courtenay, S.C.; Hanson, J.M. Ontogenetic diet shifts in age-0 Striped Bass, Morone saxatilis, from the Miramichi River estuary, Gulf of St. Lawrence. Can. J. Zoo 1997, 75, 1300–1309. [Google Scholar] [CrossRef]
- Smircich, M.G. Long-Term Feeding Ecology of Early-Stage Striped Bass (Morone Saxatilis) and American Shad (Alosa Sapidissima) in the Hudson River Estuary. Master’s Thesis, University of Connecticut, Mannsfield, CT, USA, 2016. [Google Scholar]
- Jordan, R.C.; Juanes, F. Ontogenetic shifts in feeding habits of juvenile Striped Bass (Morone saxatilis) in the mid-Hudson River estuary. In Final Reports of the Tibor T. Polgar Fellowship Program 1998; Nieder, W.C., Waldman, J.R., Eds.; Hudson River Foundation: New York, NY, USA, 1999. [Google Scholar]
- Howe, D.V.; Juanes, F. Habitat variation and the diet, growth, and conditions of juvenile Striped Bass (Morone saxatilis) in the mid-Hudson River Estuary. In Final Reports of the Tibor T. Polgar Fellowship Program 1999; Nieder, W.C., Waldman, J.R., Eds.; Hudson River Foundation: New York, NY, USA, 2000. [Google Scholar]
- Jordan, R.C.; Howe, D.V.; Hurst, T.P.; Juanes, F. Feeding Habits of Age-0 Striped Bass, Morone saxatilis, in the Mid-Hudson River Estuary: Temporal, Spatial, and ontogenetic Variation. Estuaries 2003, 26, 1486–1493. [Google Scholar] [CrossRef]
- Curran, H.W.; Ries, D.T. Fisheries investigations in the lower Hudson River. In A Biological Survey of the Lower Hudson Watershed, Supplement to the Twenty-Sixth Annual Report, 1936; Moore, E., Ed.; New York State Department of Conservation, Albany: New York, NY, USA, 1937; pp. 124–145. [Google Scholar]
- Buckel, J.A.; McKown, K.A. Competition between juvenile Striped Bass and Bluefish: Resource partitioning and growth rate. Mar. Ecol. Progress. Ser. 2002, 234, 191–204. [Google Scholar] [CrossRef]
- Melvin, G. A Survey of Striped Bass (Morone Saxatilis) in Kouchibouguac National Park, New Brunswick; Parks Canada, St.: Andrews, NB, Canada, 1979. [Google Scholar]
- Rulifson, R.A.; McKenna, S.A. Food of Striped Bass in the Upper Bay of Fundy, Canada. Trans. Am. Fish. Soc. 1987, 116, 119–122. [Google Scholar] [CrossRef]
- Leim, A.H. The fauna of Minas Channel, Minas Basin and the Shubenacadie River. Atl. Biol. Sta Orig. MS 1931, 278, 1–46. [Google Scholar]
- Wilkinson, E.B. Consumptive and Non-Consumptive Effects of Predatory Fishes on Lobster in Southern Maine. Master’s Thesis, University of New England, Biddeford, ME, USA, January 2013. [Google Scholar]
- Nelson, G.A.; Chase, B.C.; Stockwell, J. Food habits of Striped Bass (Morone saxatilis) in coastal waters of Massachusetts. J. Northwest Fish. Sci. 2003, 32, 1–25. [Google Scholar] [CrossRef]
- Sagarese, S.R.; Cerrato, R.M.; Frisk, M.G. Diet composition and feeding habits of common fishes in Long Island bays, New York. Natl. Emerg. Number Assoc. 2011, 18, 291–314. [Google Scholar] [CrossRef]
- Merriman, D. Studies on the Striped Bass (Roccus saxatilis) of the Atlantic coast. U.S. Fishery Bulletin. U. S. Fish Wildl. Serv. 1941, 50, 1–77. [Google Scholar]


| Source | Map # | Location | Time of Year/Years | Stomachs Sampled | Full Stomachs | Proportion Full | Collection Method | Evaluation Method | Bass Size Range | Primary Food |
|---|---|---|---|---|---|---|---|---|---|---|
| Department of Fisheries and Oceans (DFO) [2] | 1 | Miramichi River | May–June 2013–2015 | 1844 | 576 | 0.31 | Angling and trap net | Laboratory dissection | 19.2–86.2 cm FL | Rainbow Smelt |
| Davidson [43] | 2 | Shubenacadie Lake | 12 May–15 September 1950 | 32 | 19 | 0.59 | Angling | Angler observation | 0.1kg–7.25 kg | Unidentified Fish |
| Andrews et al. [44] | 3 | Saint John River | 25 April–3 November 2016 | 244 | 182 | 0.75 | Angling and fish lift | Gastric lavage | 40–117 cm TL | Alosines |
| Blackwell and Juanes [45] | 4 | Merrimack River | 6–28 May 1997 | 212 | 41 | 0.19 | Angling | Gastric lavage | 30–78 cm FL | Smolt |
| Warner and Kynard [46] | 5 | Connecticut River | 25 May–14 June 1982 | 78 | 65 | 0.83 | Fish lift | Laboratory dissection | 22–44 cm TL | Alosines |
| Kahnle and Hattala [47] | 6 | Hudson River | Spring 1990–2006 | 1859 | 304 | 0.16 | Electro-fishing, haul sein, gill net | Laboratory dissection | 35.6–116.5 cm TL | Unidentified Fish |
| Dew [48] | 7 | Hudson River | 28 March–20 May (1973–1975) | 510 | 201 | 0.39 | Commercial gill net | Laboratory dissection | >40 cm TL | Blueback Herring |
| Gardinier and Hoff [40] | 8 | Hudson River | April–May 1976–1977 | 380 | 102 | 0.27 | Haul sein | Laboratory dissection | 20–80 cm TL | Unidentified fish |
| Omitted Due to the Lack of Detailed Information (or Due to the Winter Sampling 2, 6) | ||||||||||
| DFO [2] 1 | 9 | SGLS* River and Coasts | 2013–2015 | 467 | 219 | 0.47 | Angling and trap net | NA | 21.3–73.1 cm FL | Shrimp spp. |
| Buhariwalla et al. [39] 2 | 10 | Pictou Harbour | 14 January 2013 | 98 | 89 | 0.91 | Thermal shock mortalities | Laboratory dissection | 11.8–60.2 cm TL | Fourspine Sticklebacks, Striped Bass |
| Beland [36] 3 | 11 | Narraguagus River | 24 May 2001 | 1 | 1 | 1 | Angling | Dissection | 34 cm FL | Smolt |
| Davis et al. [37] 4 | 12 | Connecticut River | Spring 2005–2007 | 642 | NA | NA | Electro-fishing | NA | ~75% >30 cm TL | Blueback Herring |
| Schulze [35] 5 | 13 | Connecticut River | Mid-March–mid-December 1994 | 646 | 401 | 0.62 | Gill netting | Gastric lavage | >35 cm | Invertebrates and fish |
| Dunning et al. [38] 6 | 14 | Hudson River | Winter 1986–1994 | 1703 | 814 | 0.48 | Trawl | Laboratory dissection | 8.8–66.8 cm TL | Crangon spp. |
| Gardinier and Hoff [40] 7 | 8 | Hudson River | April–November 1974 | 13 | NA | NA | Beach sein, bottom trawl | Laboratory dissection | 20.1–27.5 cm TL | Atlantic Tomcod, Clupeids, White Perch |
| Prey Items | Number of Stomachs Containing Prey/1490 | Frequency of Occurrence |
|---|---|---|
| Rainbow Smelt (Osmerus mordax) | 357 | 0.240 |
| Unidentified Alosines (A1) | 247 | 0.166 |
| Alewife (Alosa pseudoharengus) (A2) | 122 | 0.082 |
| White perch (Morone americanus) | 52 | 0.035 |
| Blueback Herring (Alosa aestivalis) (A3) | 51 | 0.034 |
| Atlantic salmon (Salmo salar) | 48 | 0.032 |
| American Eel (Anguilla rostrata) | 21 | 0.014 |
| Yellow Perch (Perca flavescens) | 18 | 0.012 |
| Atlantic Menhadden (Brevoortia tyrannus) | 17 | 0.011 |
| American Sandlance (Amodytes americanus) | 15 | 0.010 |
| Atlantic Tomcod (Microgadus tomcod) | 9 | 0.006 |
| Sea Lamprey (Petromyzon marinus) | 9 | 0.006 |
| Spottail Shiner (Notropis hudsonius) | 7 | 0.005 |
| Brown Bullhead (Ameiurus nebulosus) | 3 | 0.002 |
| Flatfish (Pleuronectidae sp.) | 3 | 0.002 |
| Needlefish (Belonidae sp.) | 3 | 0.002 |
| Anchovy (Anchoa mitchilli) | 2 | 0.001 |
| Northern Pipefish (Syngnathus fuscus) | 2 | 0.001 |
| Moronidae (sp.) | 2 | 0.001 |
| American Shad (Alosa sapidissima) (A4) | 2 | 0.001 |
| White Sucker (Catostomus commersonii) | 2 | 0.001 |
| White Bullhead (Ictalurus catus) | 1 | 0.001 |
| Cunner (Tautogolabrus adspersus) | 1 | 0.001 |
| Striped Bass (Morone saxatilis) | 1 | 0.001 |
| Atlantic Mackerel (Scomber scombrus) | 1 | 0.001 |
| Unidentified fish * | 280 | 0.188 |
| Non-salmonid Unidentified fish | 89 | 0.060 |
| Total Alosines (sum A1–A4) | 422 | 0.283 |
| Fishes (total) | 1365 | 0.916 |
| Unidentified invertebrates | 90 | 0.060 |
| Insects | 49 | 0.033 |
| Crustaceans | 44 | 0.030 |
| Blue Crab (Callinectes sapidus) | 41 | 0.028 |
| Crangon (sp.) | 38 | 0.026 |
| Isopods | 35 | 0.023 |
| Unidentified crabs | 10 | 0.007 |
| Nematodes | 11 | 0.007 |
| Amphipods | 7 | 0.005 |
| Gammarus (spp.) | 4 | 0.003 |
| Polychaetes | 5 | 0.003 |
| Squid | 3 | 0.002 |
| Mud Crab (Rhithropanopeus harrisii) | 2 | 0.001 |
| Total Invertebrates | 339 | 0.228 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrews, S.N.; Hirtle, S.V.; Linnansaari, T.; Curry, R.A. Consumption of Atlantic Salmon Smolt by Striped Bass: A Review of the Predator-Prey Encounter Literature and Implications for the Design of Effective Sampling Strategies. Fishes 2019, 4, 50. https://doi.org/10.3390/fishes4040050
Andrews SN, Hirtle SV, Linnansaari T, Curry RA. Consumption of Atlantic Salmon Smolt by Striped Bass: A Review of the Predator-Prey Encounter Literature and Implications for the Design of Effective Sampling Strategies. Fishes. 2019; 4(4):50. https://doi.org/10.3390/fishes4040050
Chicago/Turabian StyleAndrews, Samuel N., Sarah V. Hirtle, Tommi Linnansaari, and R. Allen Curry. 2019. "Consumption of Atlantic Salmon Smolt by Striped Bass: A Review of the Predator-Prey Encounter Literature and Implications for the Design of Effective Sampling Strategies" Fishes 4, no. 4: 50. https://doi.org/10.3390/fishes4040050
APA StyleAndrews, S. N., Hirtle, S. V., Linnansaari, T., & Curry, R. A. (2019). Consumption of Atlantic Salmon Smolt by Striped Bass: A Review of the Predator-Prey Encounter Literature and Implications for the Design of Effective Sampling Strategies. Fishes, 4(4), 50. https://doi.org/10.3390/fishes4040050