The Inhibitory Effects and Positive Contributions of Live Foods on Protease Activities of Meagre, Argyrosomus regius (Asso 1801), Larvae In Vitro Assay

Abstract
:1. Introduction
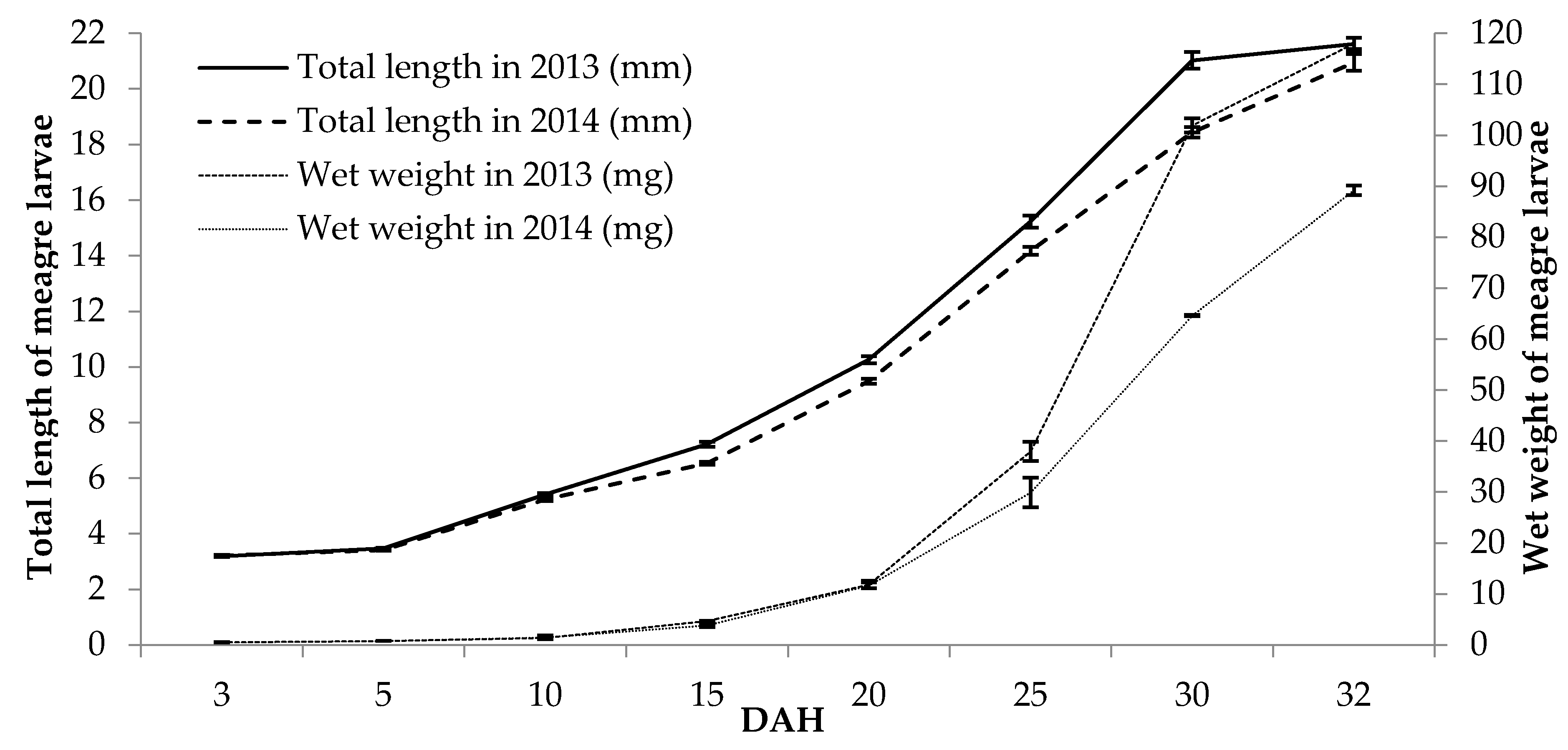
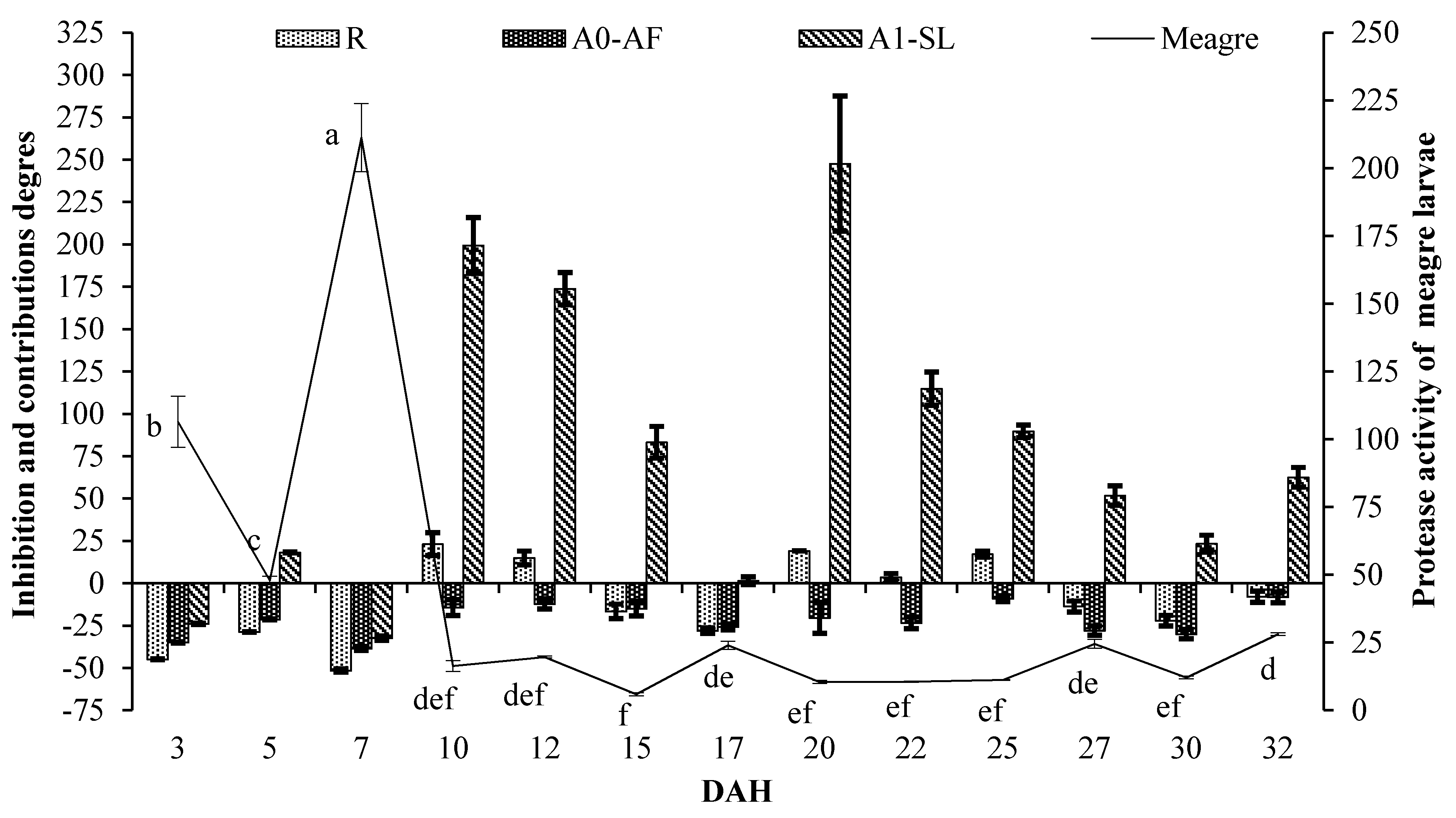
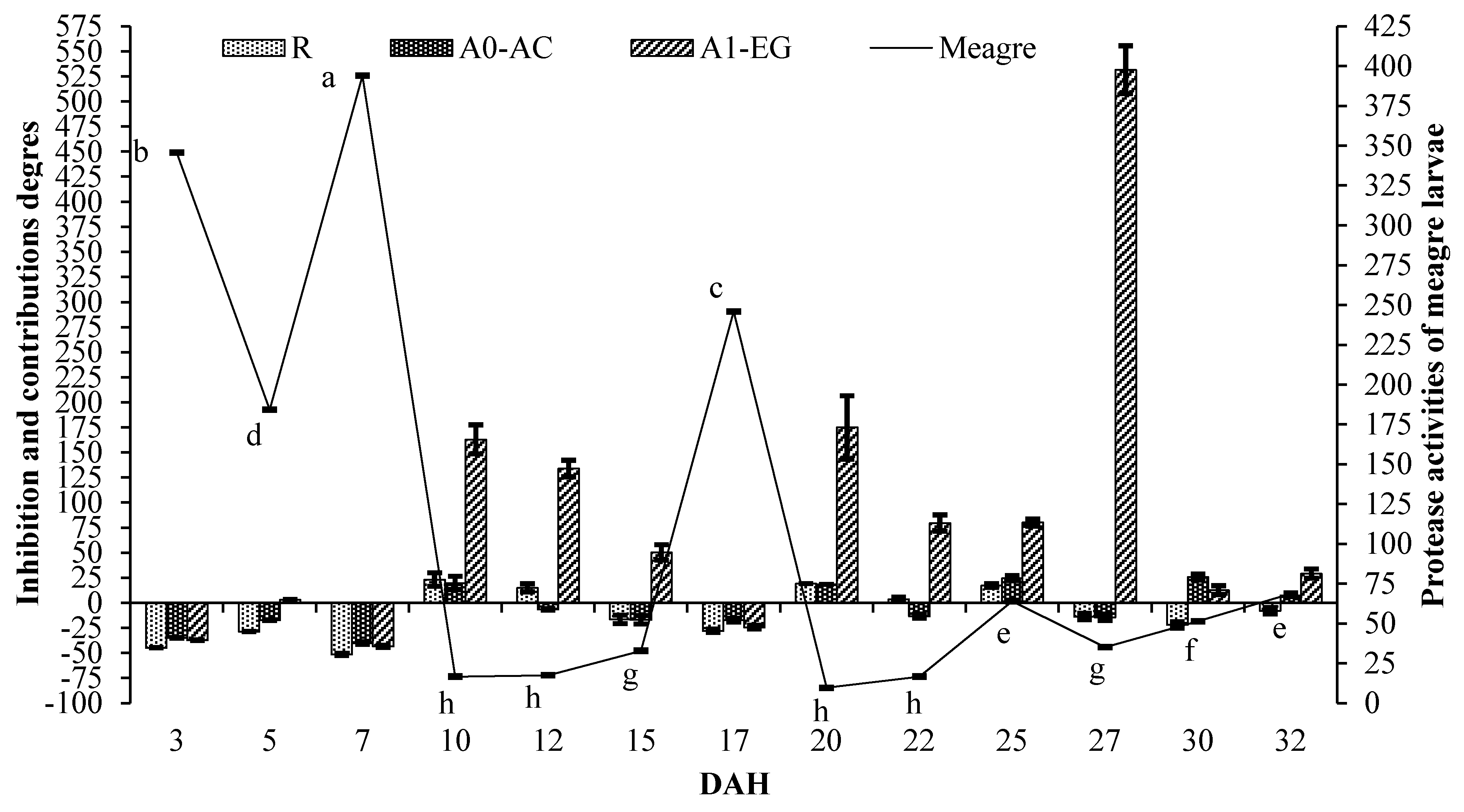
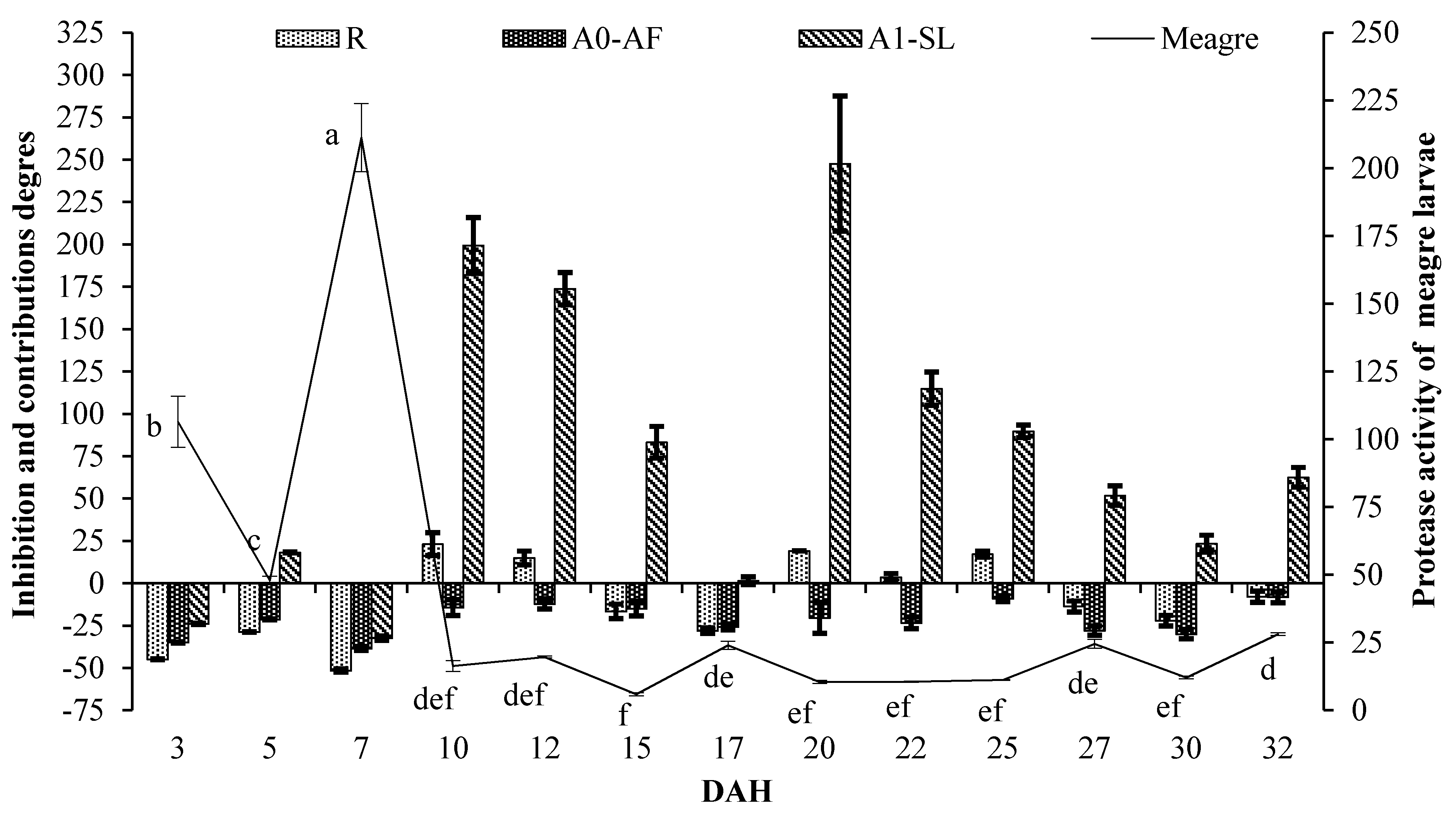
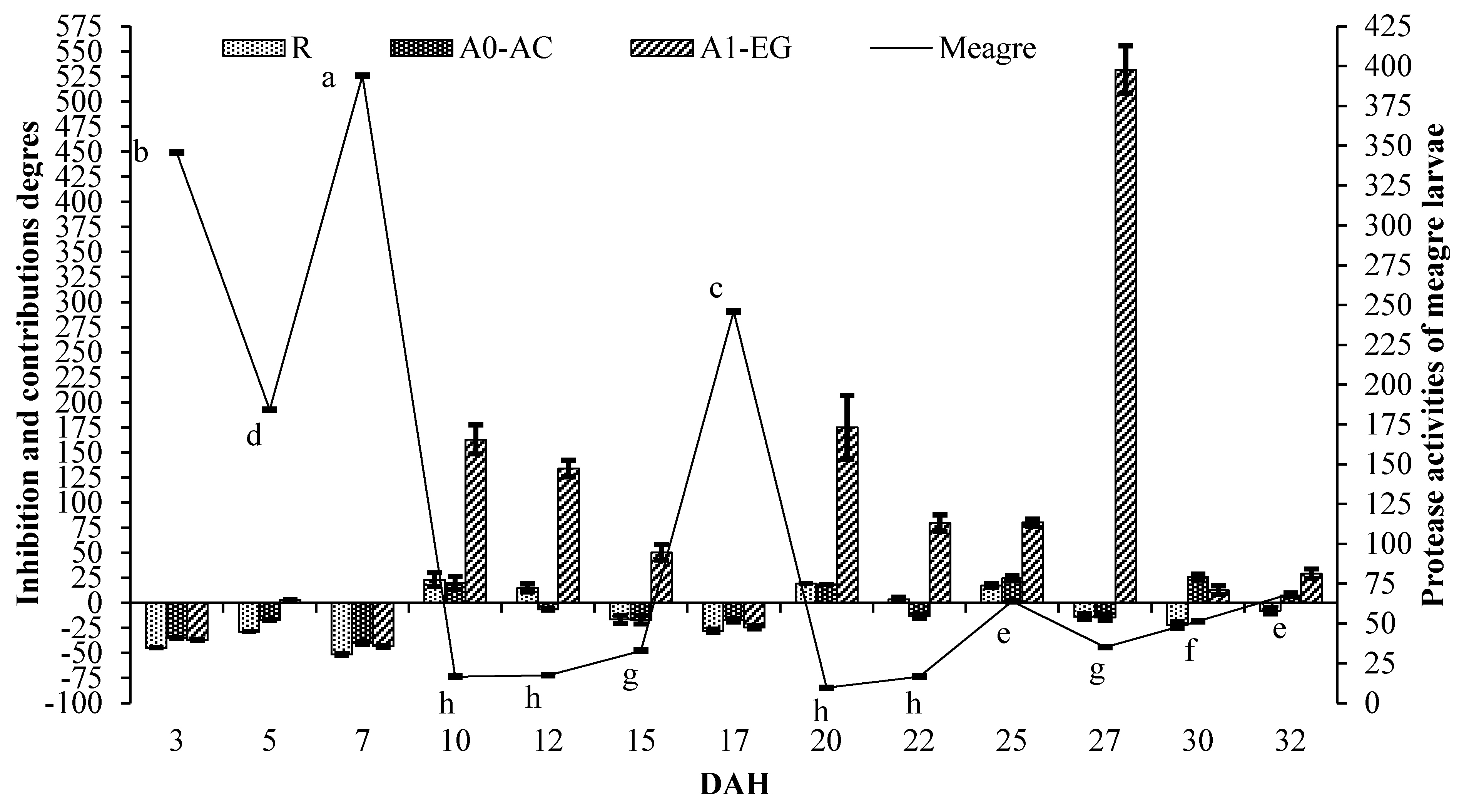
2. Results
3. Discussion
4. Materials and Methods
4.1. Larval Rearing and Sampling
4.2. Analytical Methods
4.2.1. Extracts of Larvae
4.2.2. Extracts of Live Foods
4.2.3. Determination of Protease Activities of Larvae and Live Foods
4.2.4. Effects of Live Foods on Protease Activities of Larvae
4.2.5. Statistical Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rainuzzo, J.R.; Retain, K.I.; Olsen, Y. The signifinance of lipids at early stages of marine fish: A Review. Aquaculture 1997, 155, 103–115. [Google Scholar] [CrossRef]
- Palmtag, M.R.; Faulk, C.K.; Holt, G.J. Highly unsaturated fatty acid composition of rotifers (Brachionus plicatilis) and Artemia fed various enrichments. J. World Aquacult. Soc. 2006, 37, 126–131. [Google Scholar] [CrossRef]
- Tucker, J.W., Jr. Marine Fish. In Aquaculture Farming Aquatic Animals and Plant, 2nd ed.; Lucas, J.S., Southgate, P.C., Eds.; Wiley-Blackwell: Chichester, UK, 2012; pp. 384–444. [Google Scholar]
- Uys, W.; Hecht, T. Evaluation and preparation of an optimal dry feed for the primary nursing of Clarias gariepinus larvae (Piscess: Claridae). Aquaculture 1985, 47, 173–183. [Google Scholar] [CrossRef]
- Appelbaum, S.; Van Damme, P. The feasibility of using exclusively artificial dry feed for rearing of Israeli Clarias gariepinus (Burchell, 1882) larvae and fry. J. Appl. Ichthyol. 1998, 4, 105–110. [Google Scholar] [CrossRef]
- Cahu, C.L.; Infante, J.Z. Early weaning of sea bass (Dicentrarchus labrax) larvae with a compound diet: Effect on digestive enzymes. Comp. Biochem. Physiol. Part A Physiol. 1994, 109, 213–222. [Google Scholar] [CrossRef]
- Munilla-Moran, R.; Starch, J.R.; Barbout, A. The role of exogenous enzymes in digestion in cultured turbot larvae (Scophthalmus maximus). Aquaculture 1990, 88, 337–350. [Google Scholar] [CrossRef]
- Kolkovski, S.; Tandler, A.; Kissil, W.G.; Gertlez, A. The effect of dietary exogenous digestive enzymes on ingestion, assimilation, growth and survival of gilthead seabream (Sparus aurata, L.) larvae. Fish Physiol. Biochem. 1993, 12, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Kolkovski, S.; Kowen, W.; Tandler, A. The mode of action of Artemia in enhancing utilisation of microdiet by gilthead seabream Sparus aurata larvae. Aquaculture 1997, 155, 193–205. [Google Scholar] [CrossRef]
- Kurokawa, T.; Shiraishi, M.; Suzuki, T. Quantification of exogenous protease derived from zooplankton in the intestine of Japanese sardine Sardinops melanotictus larvae. Aquaculture 1998, 161, 491–499. [Google Scholar] [CrossRef]
- García-Ortega, A.; Verreth, J.; Segner, H. Post–prandial protease activity in the digestive tract of African catfish Clarias gariepinus larvae fed decapsulated cysts of Artemia. Fish Physiol. Biochem. 2000, 22, 237–244. [Google Scholar] [CrossRef]
- Cahu, C.L.; Zambonino, I.J.L.; Le Gall, M.M.; Quazuguel, P. Early weaning of seabass: Are digestive enzymes limiting. In Larvi’95-Fish and Shellfish Larviculture Symposium; Lavens, P., Jaspers, E., Roelants, I., Eds.; European Aquaculture Society Special Publication: Gent, Belgium, 1995; pp. 268–271. [Google Scholar]
- García-Ortega, A.; Verreth, J.; Coutteau, P.; Segner, H.; Huisman, E.A.; Sorgeloos, P. Biochemical and enzymatic characterization of decapsulated cysts and nauplii of the brine shrimp Artemia at different developmental stages. Aquaculture 1998, 161, 501–514. [Google Scholar] [CrossRef]
- Naz, M. The changes in the biochemical compositions and enzymatic activities of rotifer (Branchionus plicatilis, Müller) and Artemia during the enrichment and starvation period. Fish Physiol. Biochem. 2008, 34, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-S.; O’Bryen, P.J.; Marcus, N.H. Copepods in Aquaculture, 1st ed.; Blackwell Publishing Ltd.: Oxford, UK, 2005; ISBN 978-0-8138-0066-0. [Google Scholar]
- Holt, G.J.; Webb, K.A.; Rust, M.B. Microparticule diets: Testing and evaluating success. In Larval Fish Nutrition, 1st ed.; Holt, G.J., Ed.; John Wiley & Sons Inc. Publication: Chichester, UK, 2011; pp. 353–372. ISBN 978-0-8138-1792-7. [Google Scholar]
- Øie, G.; Reitan, K.J.; Evjemo, J.O.; Støttrup, J.; Olsen, Y. Live feeds. In Larval Fish Nutrition, 1st ed.; Holt, G.J., Ed.; John Wiley & Sons Inc. Publication: Chichester, UK, 2011; pp. 307–334. ISBN 978-0-8138-1792-7. [Google Scholar]
- Naz, M.; Yılmaz, E.; Töre, Y.; Diken, G.; Tanrıverdi, Z.; Tekin, S. Canlı Yemlerin Proteaz Aktivitelerinin Belirlenmesi. In 16. Ulusal Su Ürünleri Sempozyumu; Mustafa, Ü., Deniz, Ç., Eds.; Akdeniz Üniversitesi: Antalya, Turkey, 2011; p. 146. [Google Scholar]
- Naz, M.; Yúfera, M. The Potential inhibitory effects of the commercial diets on protease activities of larvae and live foods. J. Fish. Sci. 2012, 6, 224–231. [Google Scholar] [CrossRef]
- Haközü, G. Çipura (Sparus aurata, Linneaus 1758) Larvalarının Kortizol Seviyeleri Üzerine Ticari Besleme Prosedürünün Etkileri. Master’s Thesis, Department of Aquaculture, Mustafa Kemal Üniversitesi, Hatay, Turkey, 2014. [Google Scholar]
- Kassim, Z.; John, A.; Chin, L.K.; Zakaria, N.F.; Asgnari, N.H. Sustainable technique for selected live feed culture. In Sustainable Aquaculture Techniques; Hernandez-Vergara, M.P., Perez-Rostro, C.I., Eds.; InTech: London, UK, 2014; pp. 105–133. [Google Scholar]
- Diken, G.; Demir, O.; Naz, M. The potential effects of commercial feeding protocol on protease activities and cortisol stress responses of meagre (Argyrosomus regius). Agric. Food 2016, 4, 460–472. [Google Scholar]
- Diken, G.; Demir, O.; Naz, M. Sörvaj dönemi çipura (Sparus aurata Linnaeus, 1758) ve levrek balığı (Dicentrarchus labrax Linnaeus, 1758) larvalarının proteaz aktiviteleri üzerine besinlerin etkileri ve fizyolojik durumlarının belirlenmesi. In IV. Balık Besleme ve Yem Teknolojileri Çalıştayı; Çukurova Üniversitesi: Adana, Turkey, 2016; p. 4. [Google Scholar]
- Dabrowski, K. The Feeding of Fish Larvae: Present «State of the Art» and Perspectives. Reprod. Nutr. Dév. 1984, 24, 807–833. [Google Scholar] [CrossRef]
- Koven, W.; Kolkovski, S.; Hadas, H.; Gamsız, K.; Tandler, A. Advances in the development of microdiets for gilthead seabream, Sparus aurata: A Review. Aquaculture 2001, 194, 107–121. [Google Scholar] [CrossRef]
- Lazo, J.P.; Darias, M.J.; Gisbert, E. Ontogeny of the digestive tract. In Larval Fish Nutrition, 1st ed.; Holt, J.G., Ed.; John Wiley & Sons Inc. Publication: Chichester, UK, 2011; pp. 5–46. ISBN 978-0-8138-1792-7. [Google Scholar]
- Hernández-Cruz, C.M.; Schuchardt, D.; Roo, J.; Borrero, C.; Fernández-Palacios, H. Optimización del protocolo de destete de corvina (Argyrosomus regius, Asso, 1801). In Actas XI Congreso Nacional de Acuicultura; Eiroa, A.C., Díaz, A.G., Acosta, C.P., Eds.; Centro De Investigacións Mariñas De Coron Consellería De Pesca E Asuntos Marítimos Xunta De Galicia: Vigo, Española, 2007; pp. 751–754. ISBN 978-84-611-9086-7. [Google Scholar]
- Roo, J.; Hernández-Cruz, C.M.; Borrero, C.; Schuchardt, D.; Fernández-Palacios, H. Effect of larval density and feeding sequence an meagre (Argyrosomus regius; Asso, 1801) larval rearing. Aquaculture 2010, 302, 82–88. [Google Scholar] [CrossRef]
- Arda, G. Sarıağız (Argyrosomus regius) Larvalarında Gastrointestinal Tüpün Histolojik Gelişimi. Master’s Thesis, Department of Aquaculture, Ege Üniversitesi, İzmir, Turkey, 2011. [Google Scholar]
- Gracia, E.P.; Jofre, A.G. Cultivo de esciénidos. I: La corvina. In Diversificación de Especies en la Piscicultura Marina Española Fundación; Martínez, E.A., Atarés, I.A., Eds.; Observatorio Español de Acuicultura Instituto Español de Oceanografía: Madrid, Española, 2013; pp. 117–153. [Google Scholar]
- Süzer, C.; Kamacı, H.O.; Çoban, D.; Yıldırım, Ş.; Fırat, K.; Saka, S. Functional changes in digestive enzyme activities of meagre (Argyrosomus regius; Asso, 1801) during early ontogeny. Fish Physiol. Biochem. 2013, 39, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.; Grade, A.; Candeias-Mendes, A.; Pereiraa, T.G.; Teixeiraa, B.; Yúfera, M.; Conceiçãoc, L.E.C.; Mendesa, R.; Pousão-Ferreira, P. Different dietary protein levels affect meagre (Argyrosomus regius) larval survival and muscle cellularity. Aquaculture 2016, 450, 89–94. [Google Scholar] [CrossRef]
- Diken, G.; Demir, O.; Naz, M. Determination of the inhibitory effects of microdiets used in routine commercial feeding protocols on protease activities of Argyrosomus regius (Asso, 1801) larva. Iran. J. Fish. Sci. 2017, 16, 96–107. [Google Scholar]
- Diken, G.; Demir, O.; Naz, M. The inhibitory effects of different diets on the protease activities of Argyrosomus regius (Pisces, Scianidae) larvae as a potential candidate species. J. Appl. Anim. Res. 2018, 46, 94–99. [Google Scholar] [CrossRef]
- Candeias-Mendes, A.; Castanho, S.; Ribeiro, L.; Conceição, L.E.C.; Dias, J.; Costa, S.; Bandarra, N.M.; Pousão-Ferreira, P. Melhoramento do cultivo larvar de corvina, Argyrosomus regius. In XV Congreso Nacional y I Congreso Ibérico de Acuicultura; Navas, J.I., Cordero, M.L., Cárdenas, S., Eds.; Año de Publicación: Huelva, España, 2015; pp. 536–537. [Google Scholar]
- Solovyev, M.M.; Campoverde, C.; Öztürk, S.; Moreira, C.; Diaz, M.; Moyano, F.J.; Estévez, A.; Gisbert, E. Morphological and functional description of the development of the digestive system in meagre (Argyrosomus regius): An integrative approach. Aquaculture 2016, 464, 381–391. [Google Scholar] [CrossRef]
- Zambonino, I.J.L.; Cahu, C.L. Ontogeny of the gastrointestinal tract of marine fish larvae. Comp. Biochem. Physiol. Part C 2001, 130, 477–487. [Google Scholar] [CrossRef]
- Warner, A.H.; Perz, M.J.; Osahan, J.K.; Zielinski, B.S. Potential role in development of the major cysteine protease in larvae of the brine shrimp Artemia franciscana. Cell Tissue Res. 1995, 282, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Yúfera, M.; Fernández-Diaz, C.; Pascual, E.; Sarasquete, M.C.; Moyano, F.J.; Diaz, M.; Alarcom, F.J.; Garcia-Gallego, M.; Para, G. Towards an inert diet for first feeeding gilthead seabream Sparus aurata L. larvae. Aquacult. Nutr. 2000, 6, 143–152. [Google Scholar] [CrossRef]
- Warner, A.H.; Shridhar, V. Purification and characterization of a cytosol protease from dormant cysts of the brine shrimp Artermia. J. Biol. Chem. 1985, 260, 7008–7014. [Google Scholar] [PubMed]
- Cahu, C.; Zambonino, I.J.L. Substitution of Live Food by Formulated Diets in Marine Fish Larvae. Aquaculture 2001, 200, 161–180. [Google Scholar] [CrossRef]
- Zambonino, I.J.L.; Cahu, C.L. Development and response to a diet change of some digestive enzymes in sea bass (Dicentrarchus labrax) larvae. Fish Physiol. Biochem. 1994, 12, 399–408. [Google Scholar]
- Cahu, C.L.; Zambonino, I.J.L. Effect of the molecular form of dietary nitrogen supply in sea bass larvae: Response of pancreatic enzymes and intestinal peptidases. Fish Physiol. Biochem. 1995, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Moyano, F.J.; Diaz, M.; Alarcon, F.J.; Sarasquete, M.C. Characterization of digestive enzyme activity during larval development of gilthead seabream Sparus aurata. Fish Physiol. Biochem. 1996, 15, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Walter, H.E. Proteinases: Methods with haemoglobin, casein and azocoll as substrates. In Methods of Enzymatic Analysis Volume V Enzymes 3: Peptidases, Proteinases and Their Inhibitors, 3rd ed.; Bergmeyer, H.U., Bergmeyer, J., Graßl, M., Eds.; Verlag Chemie: Weinheim, Germany, 1984; pp. 270–277. ISBN 3-527-26045-5. [Google Scholar]
- Bradford, M.M. A rapid sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- García-Carreño, F.L. Proteinase inhibitors. Trends Food Sci. Technol. 1996, 7, 197–204. [Google Scholar] [CrossRef]
- SPSS Inc. SPSS for IBM Version 23.0; SPSS Inc.: Chicago, IL, USA, 2015. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Rotifer | Artemia nauplii | Artemia metanauplii | |||
|---|---|---|---|---|---|
| R | A0–AF | A0–AC | A1–SL | A1–EG | |
| Protease Activity (U/mg Protein, Means ± SE) | |||||
| 21.76 ± 0.31 B | 36.00 ± 1.48 B | 29.3 3 ± 0.93 B | 416.44 ± 19.70 A | 403.53 ± 11.85 A | |
| Inhibition and Contributions Degres (%, Means ± SE) | |||||
| DAH | Larvae unit | ||||
| 3 | −44.87 ± 0.23 a | −35.02 ± 0.27 ab | −34.97 ± 0.27 a | −23.98 ± 0.32 a | −37.31 ± 0.26 a |
| 5 | −28.76 ± 0.18 b | −21.38 ± 0.20 cde | −17.49 ± 0.21 bc | 18.21 ± 0.30 bc | 2.91 ± 0.26bc |
| 7 | −51.60 ± 0.97 a | −38.46 ± 1.23 a | −40.29 ± 1.20 a | −32.46 ± 1.35 a | −43.57 ± 1.13 a |
| 10 | 23.06 ± 6.77f | −14.41 ± 4.71 def | 19.72 ± 6.59f | 199.31 ± 16.48g | 162.94 ± 14.48fg |
| 12 | 14.91 ± 4.01f | −12.18 ± 3.07 ef | −6.70 ± 0.65 d | 173.80 ± 9.57g | 133.87 ± 8.17f |
| 15 | −16.64 ± 4.27 cd | −14.99 ± 4.36 def | −17.02 ± 4.25 bc | 83.19 ± 9.39ef | 50.17 ± 7.70de |
| Weaning unit | |||||
| 17 | −28.10 ± 1.66 b | −25.80 ± 1.71 bcd | −17.49 ± 1.90 bc | 1.46 ± 2.34ab | −24.43 ± 1.74 ab |
| 20 | 19.05 ± 0.00f | −20.42 ± 9.14 cdef | 18.30 ± 0.00f | 247.55 ± 39.92h | 174.94 ± 31.58g |
| 22 | 3.50 ± 2.46e | −23.37 ± 3.50 bcde | −13.32 ± 1.90 cd | 114.80 ± 9.82f | 79.44 ± 8.20e |
| 25 | 17.14 ± 1.81f | −9.11 ± 1.78 f | 24.52 ± 2.44f | 89.64 ± 3.71ef | 80.07 ± 3.53e |
| 27 | −13.74 ± 3.26 cd | −27.98 ± 2.72 abc | −14.46 ± 3.24 cd | 51.77 ± 5.74cde | 531.53 ± 23.88h |
| 30 | −22.10 ± 3.11 bc | −30.05 ± 2.79 abc | −25.51 ± 2.97 b | 23.34 ± 4.92bcd | 12.54 ± 4.49bcd |
| 32 | −8.00 ± 3.30 d | −8.28 ± 3.29 f | 7.36 ± 1.12e | 62.46 ± 5.83de | 29.11 ± 4.63cd |
| Species | DAH | Species | DAH | Species | DAH | ||
|---|---|---|---|---|---|---|---|
| A. metanauplii (A1) | |||||||
| A. regius | 10–32 + | 15, 25 ~ | 30–32/17 | S. aurata1 | 34–81 | A. regius | 10, 12, 20 + |
| S. aurata1 | 34–81 | 34 ~ | 34 + | D. labrax1 | 35–67 + | D. labrax1 | 35, 45, 67 |
| A. regius | 20, 27 | ||||||
| S. aurata2 | 25 + | ||||||
| A. nauplii (A0) | Rotifer (B. plicatilis) | ||||||
| A. regius | 25 | A. regius | 10 | ||||
| S. aurata2 | 5 + | S. aurata2 | 25 + | ||||
| DAH * | Practice | ||
|---|---|---|---|
| Hormone injection (GnRH; 20 µg kg−1 ♀ and 10 µg kg−1 ♂) Eggs incubation (conical fiberglass tank; 23.6 ± 0.5 °C temperature in 2013 and 22.0 ± 0.2 at temperature in 2014, respectively) | |||
| 0–15 | Larva tank (7 m3 ellipsoidal fiberglass tanks with black walls; 75–80 larvae/L) | ||
| 15/16–32 | Weaning tank (raceway 27 and 15 m3 tank in 2013 and 2014, respectively and 10–12 larvae/L) | ||
| 0–32 | Sea water (sand, bag and UV filters; 20.8–24.1 °C temperature, 27.0–40.0 g/L salinity, 8.4–14.4 mg/L oxygen levels, 7.5–7.9 pH in 2013 and 20.8–22.2 °C temperature, 27.0–40.0 g/L salinity, 7.8–14.7 mg/L oxygen 7.7–8.1 pH levels in 2014, respectively and 18 h light and 6 h dark cycle) | ||
| Green water | |||
| 3–15 | (Sanolife GWS; INVE Aquaculture, NV Hoogveld, 91 9200, Dendermonde, Belgium or ω3 Algae®; Bernaqua, NV Hagelberg, 3 B-2250, Olen, Belgium in 2013 and Nannochloropsis occulata in 2014, respectively) | ||
| 16–26 | Sanolife GWS in 2013 and Nannochloropsis occulata in 2014. | ||
| Live food | |||
| 3–9 and 3–8 | Rotifer, Brachionus plicatilis | ||
| Culture 25 °C temperature and 28 g/L salinity (Algamac Protein Plus; Aquafaune Bio-Marine Inc. Hawthorne USA and Sparkle; INVE Aquaculture) | |||
| Enrich 26 °C temperature and 28 g/L salinity (Spresso; INVE Aquaculture) 10–15 prey/mL enriched rotifer (R) in 2013 and 2014 | |||
| 6–11 and 7–11 | Artemia nauplii (AF 480; INVE Aquaculture and Artemia Cysts in 2013 and Vinh Chau-Bac Lieu Artemia Co. in 2014, respectively) Culture 29 °C temperature and 28 g/L salinty 2–4 prey/mL in 2013 and 4–6 prey/mL in 2014 Artemia nauplii (A0), respectively | ||
| 10–32 | Artemia metanaupli (Artemia EG; Salt Lake Artemia Great Salt Lake Brine Shrimp Cooperative, Inc.) in 2013 and Artemia (SepArt EG; Aquaculture Salt Lake City Utah/USA in 2014). Culture 29 °C temperature and 28 g/L salinty Enrich 24 h, 26 °C temperature and 28 g/L salinity (Algamac 3050; Aquafaune Bio-Marine Inc., or Red Papper; Bernaqua, or Spresso; INVE Aquaculture in 2013 and Spresso; INVE Aquaculture in 2014). 1.5–6 prey/mL in 2013 and 1.5–5 prey/mL in 2014 enriched Artemia (A1), respectively | ||
| Microdiets | |||
| 16/17–32 | (Orange Start-S,100–200 µm, Orange Start-L, 200–300 µm, Orange Nurse-XS, 300–500 µm, Orange Grow-S, 300–500 µm, Orange Grow-L, 500–800 µm; INVE Aquaculture in 2013 and Gemma Micro 150, 100–200 µm; Skretting AS Sjøhagen 15, 4016 Stavanger/P.b. 319 Sentrum, 4002 Stavanger, Norway, Caviar 200–300 µm; BernAqua, Caviar 300–500 µm; BernAqua, Perla Larva Proactive 4.0 300–500 µm; Skretting AS in 2014, respectively) | ||
| Sampling | |||
| 3–32 | Before the morning feeding and stored in liquid nitrogen (−196 °C) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diken, G.; Demir, O.; Naz, M. The Inhibitory Effects and Positive Contributions of Live Foods on Protease Activities of Meagre, Argyrosomus regius (Asso 1801), Larvae In Vitro Assay. Fishes 2019, 4, 5. https://doi.org/10.3390/fishes4010005
Diken G, Demir O, Naz M. The Inhibitory Effects and Positive Contributions of Live Foods on Protease Activities of Meagre, Argyrosomus regius (Asso 1801), Larvae In Vitro Assay. Fishes. 2019; 4(1):5. https://doi.org/10.3390/fishes4010005
Chicago/Turabian StyleDiken, Gürkan, Orhan Demir, and Mehmet Naz. 2019. "The Inhibitory Effects and Positive Contributions of Live Foods on Protease Activities of Meagre, Argyrosomus regius (Asso 1801), Larvae In Vitro Assay" Fishes 4, no. 1: 5. https://doi.org/10.3390/fishes4010005
APA StyleDiken, G., Demir, O., & Naz, M. (2019). The Inhibitory Effects and Positive Contributions of Live Foods on Protease Activities of Meagre, Argyrosomus regius (Asso 1801), Larvae In Vitro Assay. Fishes, 4(1), 5. https://doi.org/10.3390/fishes4010005