
Preliminary Study on Some Blood Parameters of White Snook (Centropomus viridis) Broodstock Reared in Aquaculture Recirculating System (RAS)

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Aquaculture System and Fish Husbandry
2.2. Proximal Analysis
2.3. Blood Sample Collection and Biometrics
2.4. Hematological Analysis
2.5. Blood Biochemistry Analysis
2.6. Statistical Analysis
3. Results
3.1. Fish Biometrics

{kind=link}
| Parameters | Male (n = 20) | Max–Min | Female (n = 14) | Max–Min |
|---|---|---|---|---|
| Wet weight (K) | 2.200 ± 0.963 | 5.300–1.400 | 7.132 ± 1.610 * | 10.700–4.100 |
| Total length (M) | 0.632 ± 6 | 0.86–0.57 | 0.94 ± 0.04 * | 1.03–0.84 |
| K | 0.84 ± 0.15 | 1.10–0.54 | 0.83 ± 0.14 | 1.02–0.54 |
3.2. Proximate Analysis of Feeds
3.3. Hematology
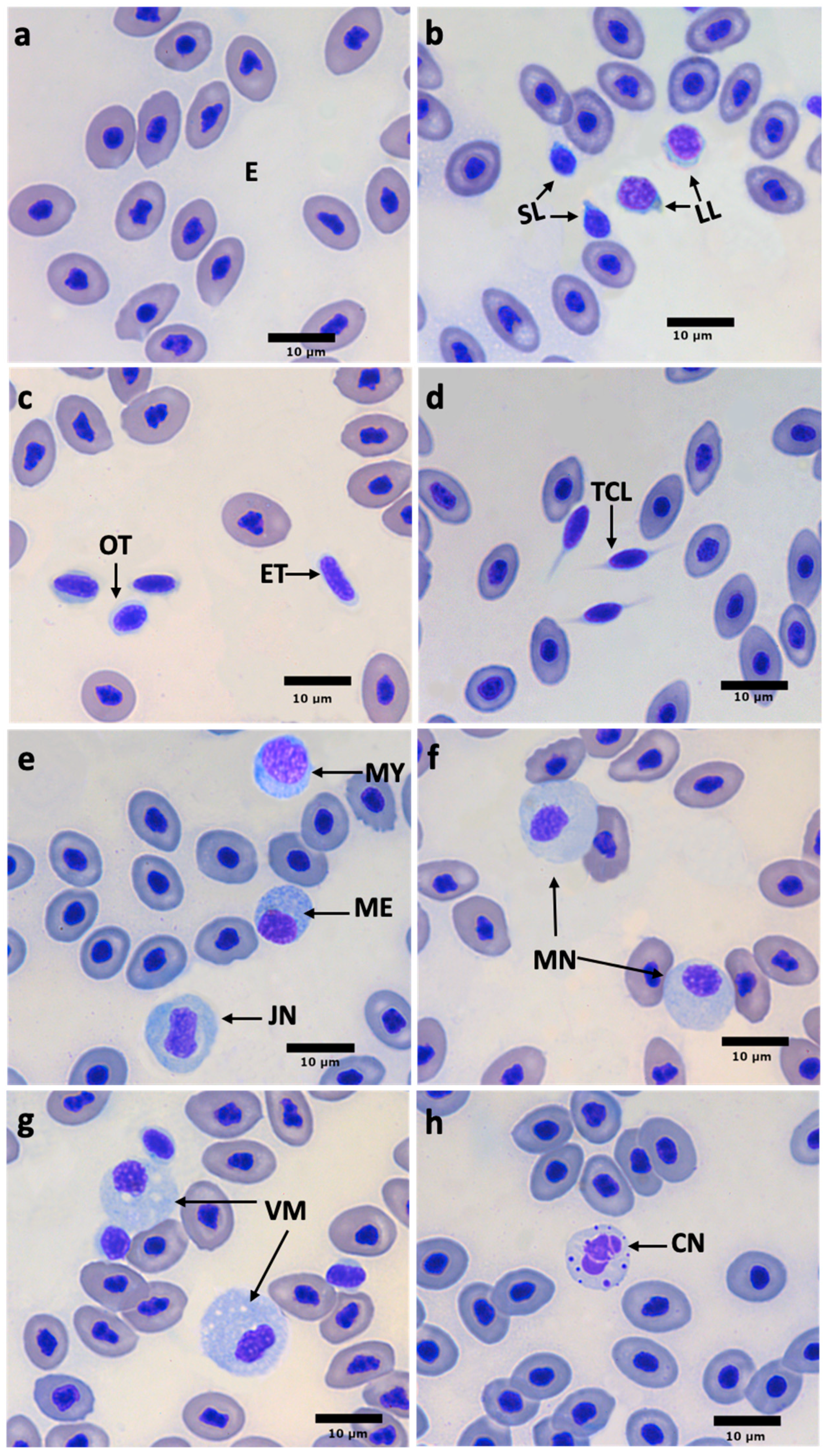
3.4. Blood Cell Size and Morphology
3.5. Blood Biochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IIO | Instituto de Investigaciones Oceanológicas |
| RBCs | Red Blood Cells |
| MCV | Mean Corpuscular Volume |
| MHC | Mean Corpuscular Hemoglobin |
| MCHC | Mean Corpuscular Hemoglobin Concentration |
| WBCs | White Blood Cells |
| AOAC | Association of Official Analytical Collaboration |
| UDK | Kjeldahl Distillation Unit |
| NFE | Nitrogen-free extract |
| CIAD | Centro de Investigación en Alimentación y Desarrollo S.A. |
| HGB | Hemoglobin |
| CMOS | Complementary Metal–Oxide Semiconductor |
| HCT | Hematocrit |
| DE | Standard Deviation |
| SL | Small Lymphocyte |
| LL | Large Lymphocyte |
| OT | Oval Thrombocyte |
| ET | Elongate Thrombocyte |
| TCL | Thrombocytes With Cytoplasmic Lengthening |
| MY | Myelocyte |
| ME | Metamyelocyte |
| JN | Juvenile Neutrophil |
| MN | Mature Neutrophil |
| VM | Vacuolated Monocyte |
| CN | Neutrophil-like cell |
| PSU | Practical Salinity Unit |
| RAS | Recirculating aquaculture systems |
| Nd | No determinate, cytoplasm almost null. |
References
- Gonzales, A.; Curto, G.; Fernández-Mendez, C. Hematological parameters of Brycon amazonicus (Bryconidae) breeders in culture. Rev. Investig. Vet. Perú 2019, 30, 133–142. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Wedemeyer, G.A. Biological Interactions. In Physiology of Fish in Intensive Culture Systems; Springer: Boston, MA, USA, 1996; pp. 166–202. [Google Scholar]
- Gayatri, A.; Mohanty, P.K. Effect of sex on haemocyte biochemical profiling of silver tiger fish (Datnioides polota Hamilton, 1822). Comp. Clin. Pathol. 2018, 27, 1335–1342. [Google Scholar] [CrossRef]
- Ruiz-González, L.E.; Vega-Villasante, F.; Tintos-Gómez, A.; Del Río-Zaragoza, O.B.; Hernández-Rodríguez, M.; Patiño-Barragán, M.; Badillo-Zapata, D. Some hematology and blood chemistry parameters of the Pacific fat sleeper Dormitator latifrons (Richardson, 1844). Lat. Am. J. Aquat. Res. 2020, 48, 131–135. [Google Scholar] [CrossRef]
- Ahmed, I.; Reshi, Q.M.; Fazio, F. The influence of the endogenous and exogenous factors on hematological parameters in different fish species: A review. Aquacult. Int. 2020, 28, 869–899. [Google Scholar] [CrossRef]
- Clauss, T.M.; Dove, A.D.M.; Arnold, J.E. Hematologic disorders of fish. Fishes 2008, 11, 445–462. [Google Scholar] [CrossRef]
- Cnaani, A.; Tinman, S.; Avidar, Y.; Ron, M.; Hulata, G. Comparative study of biochemical parameters in response to stress in Oreochromis aureus, O. mossambicus and two strains of O. niloticus. Aquac. Res. 2004, 35, 1434–1440. [Google Scholar] [CrossRef]
- Parrino, V.; Cappello, T.; Costa, G.; Cannavà, C.; Sanfilippo, M.; Fazio, F.; Fasulo, S. Comparative study of haematology of two teleost fish (Mugil cephalus and Carassius auratus) from different environments and feeding habits. Eur. Zool. J. 2018, 85, 194–200. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Aragona, F.; Habib, S.S.; Fazio, F.; Zumbo, A.; Costa, A.; Riolo, K.; Giannetto, A.; Parrino, V. Morphometric, Nutritional, and Blood Analyses in Hybrid Striped Bass (Morone chrysops x Morone saxatilis, Walbaum 1972) Reared in a Recirculating Aquaculture System (RAS) Implant in Sicily, Italy. Fishes 2025, 10, 278. [Google Scholar] [CrossRef]
- Del Rio-Zaragoza, O.B.; Araújo, B.C.; Viana, M.T. Health status evaluation of striped bass (Morone saxatilis) exposed to low temperature in sea cage culture during the grow-out. Aquac. Res. 2021, 52, 2435–2445. [Google Scholar] [CrossRef]
- Ruiz-González, L.E.; Tafoya-Sánchez, D.J.; Tintos-Gómez, A.; Del Río-Zaragoza, O.B.; Vargas-Ceballos, M.A.; Vega-Villasante, F. Some blood parameters of Pacific fat sleepers, Dormitator latifrons (Richardson, 1844): A comparative study between male and female in two growth stages. Lat. Am. J. Aquat. Res. 2023, 51, 592–597. [Google Scholar] [CrossRef]
- Correa Negrete, J.C.; Garrido Correa, A.A.; Prieto Guevara, M.J.; Atencio García, V.J.; Pardo Carrasco, S.C. Caracterización de células sanguíneas y parámetros hematológicos en blanquillo Sorubim cuspicaudus. Zootec. Trop. 2009, 27, 393–405. [Google Scholar]
- Fazio, F.; Lanteri, G.; Saoca, C.; Laria, C.; Piccione, G.; Orefice, T.; Calabrese, E.; Vazzana, I. Individual variability of blood parameters in striped bass Morone saxatilis: Possible differences related to weight and length. Aquac. Int. 2020, 28, 1665–1673. [Google Scholar] [CrossRef]
- Sáez, G.; Chero, J.; Cruces, C.; Minaya, D.; Rodriguez, C.; Suyo, B.; Romero, S.; Guabloche, A.; Tuesta, E.; Alvariño, L.; et al. Hematological and blood biochemistry parameters in ten species of marine fish captured by artisanal fisheries in the Bay of Callao, Peru. Rev. Investig. Vet. Perú 2018, 29, 1161–1177. [Google Scholar] [CrossRef]
- Labastida-Che, A.; Núñez-Orozco, A.L.; Oviedo-Piamonte, J.A. Aspectos biológicos del robalo hocicudo Centropomus viridis, en el sistema lagunar Chantuto-Panzacola, Chiapas, México. Cienc. Pesq. 2013, 21, 21–28. [Google Scholar]
- Navarro-Flores, J.; Martínez-Brown, J.M.; Zavala-Leal, I.; Rojo-Cebreros, A.H.; Ibarra-Castro, L. Assessing the feasibility of exogenous 17β-estradiol for inducing sex change in white snook, C. viridis: From growth, resting and maturation studies. Aqua. Rep. 2023, 33, 101767. [Google Scholar] [CrossRef]
- Ibarra-Castro, L.; Navarro-Flores, J.; Sánchez-Téllez, J.L.; Martínez-Brown, J.M.; Ochoa- Bojórquez, L.A.; Rojo-Cebreros, A.H. Hatchery Production of Pacific White Snook at CIAD-Unity Mazatlan. Mex. World Aquac. 2017, 48, 25–29. [Google Scholar]
- Baldini, G.; Santamaría-Miranda, A.; Martínez-Brown, J.M.; Ibarra-Castro, L. Technical-economic viability of white snook Centropomus viridis culture in floating cages in a coastal lagoon in northwestern Mexico. Aquac. Rep. 2022, 23, 101048. [Google Scholar] [CrossRef]
- Montoya Ponce, C.O.; Santamaría Miranda, A.; Trigueros Salmerón, J.Á.; Apún Molina, J.P.; Valenzuela Orduño, F.G.; Lugo Gamboa, R.R. Bioeconomic Analysis of Snook Centropomus viridis, C. nigrescens, and C. medius for the Development of Mariculture in Northern Sinaloa. Fishes 2024, 9, 39. [Google Scholar] [CrossRef]
- Çelik, E.Ş.; Kaya, H.; Yılmaz, S.; Çakıcı, H. Effect of water temperature, salinity, season, reproduction, sex, size, and age on hematological parameters of horse mackerel (Trachurus trachurus). Kafkas Univ. Vet. Fak. Derg. 2012, 18, 551–558. [Google Scholar] [CrossRef]
- Suljević, D.; Mitrašinović-Brulić, M. The first record of brook trout (Salvelinus fontinalis, Salmonidae) blood cell characteristics and hematological profile: The influence of fish sex on leukocyte count. Aquac. Int. 2020, 28, 2505–2516. [Google Scholar] [CrossRef]
- Abdo-de la Parra, M.I.; Rodríguez-Ibarra, L.E.; Ibarra-Castro, L.; Martínez-Brown, J.M.; Álvarez-González, C.A.; Peña, E.; Velasco-Blanco, G.; Domínguez-Jiménez, P.; Rodríguez-Montes de Oca, G. Evaluation of different levels of dietary protein and lipids on the growth, feed efficiency, and biometric and hematological indexes of juvenile white snooks, Centropomus viridis. Cienc. Mar. 2023, 49, e3368. [Google Scholar] [CrossRef]
- [AOAC] Association of Official Analytical Chemists. Official Methods of Analysis of the Association of Official Analytical Chemists, 17th ed.; AOAC: Arlington, MA, USA, 2000; 684p. [Google Scholar]
- Abdo-de la Parra, M.I.; Rodríguez-Ibarra, L.E.; Ibarra-Castro, L.; Martínez-Brown, J.M.; Velasco-Blanco, G. Effects of frequency and feeding time on growth, food utilization, somatic indexes and survival of juvenile white snook Centropomus viridis. Cienc. Marinas. 2020, 46, 155–163. [Google Scholar] [CrossRef]
- Del Rio-Zaragoza, O.B.; Fajer-Ávila, E.J.; Almazán-Rueda, P.; Abdo de la Parra, M.I. Hematological characteristics of the spotted rose snapper Lutjanus guttatus (Steindachner, 1869) healthy and naturally infected by dactylogyrid monogeneans. Tiss. Cell 2011, 43, 137–142. [Google Scholar] [CrossRef]
- Del Rio-Zaragoza, O.B.; Hernández-Rodríguez, M.; Bückle-Ramirez, L.F. Thermal stress effect on tilapia Oreochromis mossambicus (Pisces: Cichlidae) blood parameters. Mar. Freshw. Behav. Physiol. 2008, 41, 135–145. [Google Scholar] [CrossRef]
- Natt, M.P.; Herrick, C.A. A new blood diluent for counting erythrocytes and leukocytes of the chicken. Poult. Sci. 1952, 31, 735–738. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Fernández-Palacios, H.; Tacon, A.G.J. Effect of broodstock nutrition on reproductive performance of fish. Aquaculture 2001, 197, 25–42. [Google Scholar] [CrossRef]
- Nurjanah, N.; Suseno, S.H.; Hidayat, T.; Arifianto, T.B. Changes in nutritional composition of skipjack (Katsuwonus pelamis) due to frying process. Int. Food Res. J. 2015, 22, 2093–2102. [Google Scholar]
- Dayakar, B.; Xavier, K.A.M.; Ngasotter, S.; Layana, P.; Balange, A.K.; Priyadarshini, B.; Nayak, B.B. Characterization of spray-dried carotenoprotein powder from Pacific white shrimp (Litopenaeus vannamei) shells and head waste extracted using papain: Antioxidant, spectroscopic, and microstructural properties. LWT 2022, 159, 113188. [Google Scholar] [CrossRef]
- Ranzani-Paiva, M.J.; Ishikawa, C.M. Haematological characteristics of freshwater-reared and wild mullet, Mugil platanus Günther (Osteichthyes, Mugilidae). Rev. Bras. Zool. 1996, 13, 561–568. [Google Scholar] [CrossRef]
- Chen, H.; Luo, D. Application of haematology parameters for health management in fish farms. Rev. Aquac. 2022, 15, 704–737. [Google Scholar] [CrossRef]
- Kim, S.G.; Kang, J.C. Effect of dietary copper exposure on accumulation, growth and hematological parameters of the juvenile rockfish, Sebastes schlegeli. Mar. Environ. Res. 2004, 58, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.M.; Baldwin, J.; Seymour, R.S.; Christian, K.; Brittain, T. Red blood cell function and haematology in two tropical freshwater fishes from Australia. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 141, 87–93. [Google Scholar] [CrossRef]
- Elahee, K.B.; Bhagwant, S. Hematological and gill histopathological parameters of three tropical fish species from a polluted lagoon on the west coast of Mauritius. Ecotoxicol. Environ. Saf. 2007, 68, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Kochinian, P.; Salati, A.P. The effect of sexuality on some haematological parameters of the yellowfin seabream, Acanthopagrus latus in Persian Gulf. Iran. J. Vet. Res. 2013, 14, 273–278. [Google Scholar] [CrossRef]
- Witeska, M. Erythrocytes in teleost fishes: A review. Zool. Ecol. 2013, 23, 275–281. [Google Scholar] [CrossRef]
- Sayed, A.E.H.; Mahmoud, U.M.; Muhammad, O.I. Comparative study of two carnivorous fish (Parupeneus forsskali and Thalassoma klunzingeri) from the Red Sea: Hemato-biochemical parameters and cellular characterization. Tissue Cell 2020, 63, 101316. [Google Scholar] [CrossRef]
- Habiba, M.M.; Hussein, E.E.; Ashry, A.M.; El-Zayat, A.M.; Hassan, A.M.; El-Shehawi, A.M.; Sewilam, H.; Van Doan, H.; Dawood, M.A.O. Dietary cinnamon successfully enhanced the growth performance, growth hormone, antibacterial capacity, and immunity of European Sea bass (Dicentrarchus labrax). Animals 2021, 11, 2128. [Google Scholar] [CrossRef]
- Satheeshkumar, P.; Ananthan, G.; Senthilkumar, D.; Khan, A.B.; Jeevanantham, K. Comparative investigation on haematological and biochemical studies on wild marine teleost fishes from Vellar estuary, southeast coast of India. Comp. Clin. Pathol. 2012, 21, 275–281. [Google Scholar] [CrossRef]
- Zhu, P.; Tang, Y.; Fan, J.; Fang, J.; Peng, X.; Cui, H. Hematological parameters and blood cell morphology of male and female Schizothorax (Racoma) davidi (Sauvage). J. World Aquac. Soc. 2017, 48, 821–830. [Google Scholar] [CrossRef]
- Nabi, N.; Ahmed, I.; Wani, G.B. Hematological and serum biochemical reference intervals of rainbow trout, Oncorhynchus mykiss cultured in Himalayan aquaculture: Morphology, morphometrics and quantification of peripheral blood cells. Saudi J. Biol. Sci. 2022, 29, 2942–2957. [Google Scholar] [CrossRef]
- Santos, A.A.; Ranzani-Paivad, M.J.T.; Leite da Veigae, M.; Faustinof, L.; Egami, M.I. Hematological parameters and phagocytic activity in fat snook (Centropomus parallelus) bred in captivity. Fish Shellfish Immunol. 2012, 30, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Yao, H.; Qiang, L.; Chen, X.; Gao, Y. Comparative study of peripheral blood cells in two marine fishes (Synechogobius hasta and Sebastes schlegelii): Morphological and cytochemical characterization. Tissue Cell 2021, 73, 101633. [Google Scholar] [CrossRef]
- Imagawa, T.; Hashimoto, Y.; Kitagawa, H.; Kon, Y.; Kudo, N.; Sugimura, M. Morphology of blood cells in carp (Cyprinus carpio L.). Nihon Juigaku Zasshi 1989, 51, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Qiang, L.; Wu, N.; Wang, H.; Hao, Y. Morphological and cytochemical characterization of the peripheral blood cells in Paralichthys olivaceus. Heliyon 2024, 10, e37011. [Google Scholar] [CrossRef]
- Zhu, W.; Su, J. Immune functions of phagocytic blood cells in teleost. Rev. Aquac. 2021, 14, 630–646. [Google Scholar] [CrossRef]
- Havixbeck, J.J.; Barreda, D.R. Neutrophil development, migration, and function in teleost fish. Biology 2015, 4, 715–734. [Google Scholar] [CrossRef]
- Buchmann, K. Neutrophils and aquatic pathogens. Parasite Immunol. 2022, 44, e12915. [Google Scholar] [CrossRef]
- Megarani, D.V.; Hardian, A.B.; Arifianto, D.; Santosa, C.M.; Salasia, S.I.O. Comparative morphology and morphometry of blood cells in zebrafish (Danio rerio), common carp (Cyprinus carpio carpio), and tilapia (Oreochromis niloticus). J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 673–680. [Google Scholar] [CrossRef]
- Rowley, A.F.; Hunt, T.C.; Page, M.; Mainwaring, G. Fish. In Vertebrate Blood Cells; Rowley, A.F., Ratcliffe, N.A., Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 19–127. [Google Scholar]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef]
- Tavares-Dias, M.; Barcellos, J.F.M.; Marcon, J.L.; Menezes, G.C.; Ono, E.A.; Affonso, E.G. Hematological and biochemical parameters for the pirarucu Arapaima gigas Schinz, 1822 (Osteoglossiformes, Arapaimatidae) in net cage culture. Electron. J. Ichthyol. 2007, 2, 61–68. [Google Scholar] [CrossRef]
- Hrubec, T.C.; Cardinale, J.L.; Smith, S.A. Haematology and plasma chemistry reference intervals for cultured tilapia (Oreochromis hybrid). Vet. Clin. Pathol. 2000, 29, 7–12. [Google Scholar] [CrossRef]
- Fajer-Ávila, E.J.; Guzman-Beltran, L.; Zárate-Rodríguez, W.C.; Del Río-Zaragoza, O.B.; Almazan-Rueda, P. Pathology caused by adult Pseudochondracanthus diceraus (Copepoda: Chondracanthidae), a parasite of bullseye puffer fish Sphoeroides annulatus. Rev. Biol. Mar. Oceanogr. 2011, 46, 293–302. [Google Scholar] [CrossRef]
- Nieto-Vera, M.T.; Rodríguez-Pulido, J.A.; Góngora-Orjuela, A. ¿Qué sabemos de los esteroides sexuales y las gonadotropinas en la reproducción de teleósteos neotropicales? Orinoquia 2020, 24, 52–63. [Google Scholar] [CrossRef]
- Lubzens, E.; Young, G.; Bobe, J.; Cerdà, J. Oogenesis in teleosts: How fish eggs are formed. Gen. Comp. Endocrinol. 2010, 165, 367–389. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, S.; Fiocchi, E.; Toomey, L.; Boscarato, M.; Manfrin, A.; Dimitroglou, A.; Papaharisis, L.; Passabi, E.; Stefani, A.; Lembo, G.; et al. Comparative analysis of blood protein fractions in two Mediterranean farmed fish: Dicentrarchus labrax and Sparus aurata. BMC Vet. Res. 2024, 20, 322. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose metabolism in fish: A review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Petillo, E.C.; Ferreira, A.d.C.; Oliveira CPFd Brandão, L.V.; Marinho-Pereira, T.; Cavero, B.A.S. Tambaqui (Colossoma macropomum) in RAS Technology: Zootechnical, Hematological, Biochemical and Kn Profiles at Different Stocking Densities During the Initial Grow-Out Phase. Aquac. J. 2025, 5, 1. [Google Scholar] [CrossRef]
- Barbacariu, C.-A.; Rimbu, C.M.; Burducea, M.; Dirvariu, L.; Miron, L.-D.; Boiangiu, R.S.; Dumitru, G.; Todirascu-Ciornea, E. Comparative Study of Flesh Quality, Blood Profile, Antioxidant Status, and Intestinal Microbiota of European Catfish (Silurus glanis) Cultivated in a Recirculating Aquaculture System (RAS) and Earthen Pond System. Life 2023, 13, 1282. [Google Scholar] [CrossRef]
- Stallbohm, R.; Owatari, M.S.; Zaniboni-Filho, E.; Martins, M.L. Recirculating aquaculture systems affects hematological parameters and increases ectoparasite susceptibility in Nile tilapia Oreochromis niloticus. Mar. Fish. Sci. (MAFIS) 2024, 37, 609–618. [Google Scholar] [CrossRef]
| RAS | Temperature (°C) | Salinity (PSU) | pH | Oxygen (mg mL−1) | TAN (NH4+ + NH3) (ppm) | Nitrate (NO3−) (ppm) | Nitrite (NO2−) (ppm) |
|---|---|---|---|---|---|---|---|
| 1 | 20.8 ± 0.6 | 34.8 ± 0.6 | 7.83 ± 0.08 | 6.9 ± 0.9 | 0.25 | <0.25 | <0.25 |
| 2 | 21.7 ± 0.6 | 34.7 ± 0.4 | 7.80 ± 0.13 | 6.8 ± 0.9 | <0.25 | <0.25 | <0.25 |
| 3 | 21.3 ± 0.7 | 34.5 ± 0.5 | 7.45 ± 0.23 | 6.7 ± 0.9 | <0.25 | <0.25 | <0.25 |
| 4 | 21.5 ± 0.7 | 34.7 ± 0.3 | 7.62 ± 0.12 | 6.7 ± 0.9 | <0.25 | <0.25 | <0.25 |
| Crude Protein | Crude Lipids | Ash | Moisture | |
|---|---|---|---|---|
| Semi-moist feed | 70.15 ± 0.84 b | 6.97 ± 0.55 b | 14.5 ± 0.09 b | 58.75 ± 0.35 a |
| Shrimp cephalothorax | 56.05 ± 2.71 a | 9.51 ± 0.44 c | 18.52 ± 0.05 c | 69.86 ± 0.70 b |
| Skipjack muscle | 87.43 ± 0.51 c | 1.41 ± 0.02 a | 6.33 ± 0.08 a | 74.8 ± 0.14 c |
| Parameters | Male | Max.–Min. | Female | Max.–Min. |
|---|---|---|---|---|
| HCT (%) | 28.94 ± 6.46 | 36–12 | 33.71 ± 6.11 * | 48–28 |
| RBC (×106 mm3) | 1.66 ± 0.52 | 2–0.4 | 1.62 ± 0.36 | 2–0.7 |
| HGB (g/dL) | 9.88 ± 4.46 | 18–2 | 14.03 ± 6.92 | 22–3 |
| MCV (fl) | 157.98 ± 36.21 | 42–105 | 222.91 ± 85.68 * | 462–149 |
| MCH (pg) | 69.33 ± 43.75 | 177–14 | 101.46 ± 64.66 | 277–45 |
| MCHC (g/dL) | 41.53 ± 24.50 | 112–8 | 43.66 ± 16.90 | 70–9 |
| WBC (×103 mm3) | 13.25 ± 0.91 | 48–12 | 13.76 ± 0.92 | 15–12 |
| Leukocyte (%) | 54.99 ± 1.76 | 58–52 | 55.06 ± 2.77 | 61–51 |
| Thrombocyte (%) | 45 ± 1.76 | 47–41 | 44.93 ± 2.77 | 48–38 |
| Lymphocyte (%) | 81.27 ± 7.33 | 95–73 | 82.42 ± 9.15 | 96–74 |
| Neutrophil (%) | 15.07 ± 5.94 | 22–5 | 12.40 ± 6.76 | 20–0.87 |
| Monocyte (%) | 1.98 ± 1.44 | 4–0.54 | 2.93 ± 1.87 | 5–1.03 |
| Male | Female | |||
|---|---|---|---|---|
| Cell Type | Nucleus Length (μm) | Whole-Cell Length (μm) | Nucleus Length (μm) | Whole-Cell Length (μm) |
| Erythrocytes (n = 100) | 3.82 ± 0.34 | 9.82 ± 0.78 | 3.94 ± 0.43 | 10.20 ± 0.63 * |
| Small lymphocyte (n = 100) | 4.66 ± 0.58 | Nd | 4.78 ± 0.56 | Nd |
| Large lymphocyte (n = 50) | 6.70 ± 0.43 | Nd | 6.71 ± 0.53 | Nd |
| Oval thrombocyte (n = 50) | 5.17 ± 0.49 | Nd | 5.35 ± 0.44 | Nd |
| Spindle-shaped thrombocyte (n = 50) | 6.50 ± 0.37 | Nd | 6.87 ± 0.74 | Nd |
| Neutrophil (n = 50) | 5.32 ± 0.73 | 8.50 ± 0.67 | 5.99 ± 0.94 | 8.93 ± 1.08 |
| Monocytes (n = 50) | 5.81 ± 1.58 | 10.86 ± 0.80 | 5.40 ± 0.63 | 10.69 ± 0.91 |
| Sex | Protein (g/L) | Max.–Min. | Glucose (mg/dL) | Max.–Min. | Triglycerides (mg/dL) | Max.–Min. | Cholesterol (mg/dL) | Max.–Min. |
|---|---|---|---|---|---|---|---|---|
| Male | 47.82 ± 6.8 | 41–60 | 62.40 ± 12.07 | 89–45 | 202.20 ± 100.52 | 375–28 | 224.76 ± 76.32 | 308–36 |
| Female | 49.53 ± 9.5 | 27–60 | 78.22 ± 18.12 * | 114–38 | 349.94 ± 156.23 * | 605–78 | 311.06 ± 63.80 * | 477–230 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-López, I.A.; Domínguez-Jiménez, V.P.; Medina-Guerrero, R.M.; Lozano-Olvera, R.; Del Rio-Zaragoza, O.B.; Ibarra-Castro, L.; Martínez-Brown, J.M.; Peña-Marín, E.S. Preliminary Study on Some Blood Parameters of White Snook (Centropomus viridis) Broodstock Reared in Aquaculture Recirculating System (RAS). Fishes 2025, 10, 347. https://doi.org/10.3390/fishes10070347
Hernández-López IA, Domínguez-Jiménez VP, Medina-Guerrero RM, Lozano-Olvera R, Del Rio-Zaragoza OB, Ibarra-Castro L, Martínez-Brown JM, Peña-Marín ES. Preliminary Study on Some Blood Parameters of White Snook (Centropomus viridis) Broodstock Reared in Aquaculture Recirculating System (RAS). Fishes. 2025; 10(7):347. https://doi.org/10.3390/fishes10070347
Chicago/Turabian StyleHernández-López, Iris Adriana, Virginia Patricia Domínguez-Jiménez, Rosa María Medina-Guerrero, Rodolfo Lozano-Olvera, Oscar Basilio Del Rio-Zaragoza, Leonardo Ibarra-Castro, Juan Manuel Martínez-Brown, and Emyr Saúl Peña-Marín. 2025. "Preliminary Study on Some Blood Parameters of White Snook (Centropomus viridis) Broodstock Reared in Aquaculture Recirculating System (RAS)" Fishes 10, no. 7: 347. https://doi.org/10.3390/fishes10070347
APA StyleHernández-López, I. A., Domínguez-Jiménez, V. P., Medina-Guerrero, R. M., Lozano-Olvera, R., Del Rio-Zaragoza, O. B., Ibarra-Castro, L., Martínez-Brown, J. M., & Peña-Marín, E. S. (2025). Preliminary Study on Some Blood Parameters of White Snook (Centropomus viridis) Broodstock Reared in Aquaculture Recirculating System (RAS). Fishes, 10(7), 347. https://doi.org/10.3390/fishes10070347