
Modelling the Spatial Distribution of Dosidicus gigas in the Southeast Pacific Ocean at Multiple Temporal Scales Based on Deep Learning



 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Data Preprocessing
2.2.1. Experimental Cases Design
2.2.2. Normalization and Invalid Value Handling
2.3. Model Architecture
2.3.1. Generalized Additive Model (GAM)
2.3.2. Extreme Gradient Boosting (XGBoost)
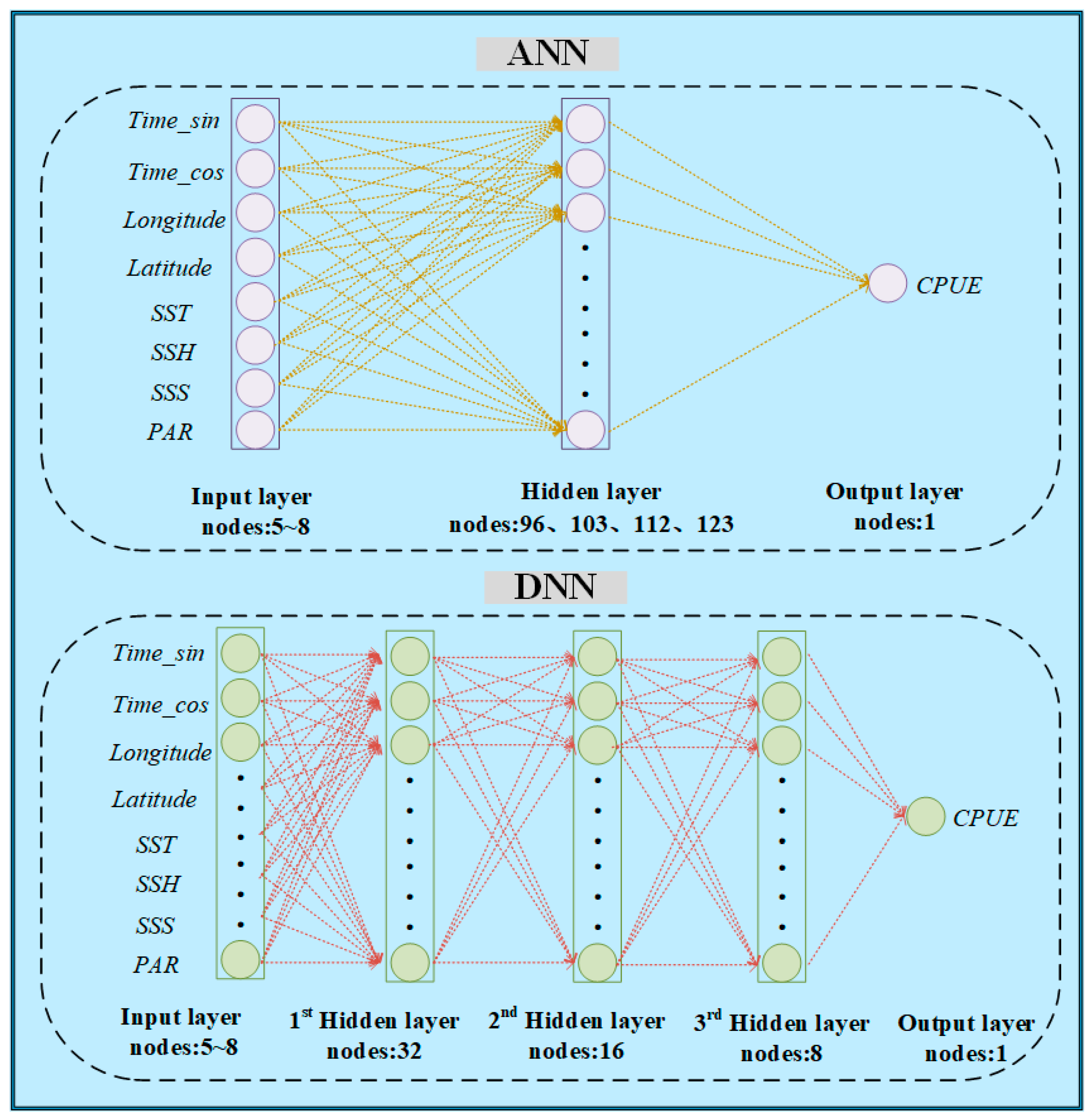
2.3.3. Artificial Neural Network (ANN) and Deep Neural Network (DNN)
2.4. Model Evaluation Parameters
2.5. Model Implementation
2.6. Interpretability of Model Input Factors
3. Results
3.1. MSE and MAE of Different Models
3.2. AUC Evaluation of P-R Curves
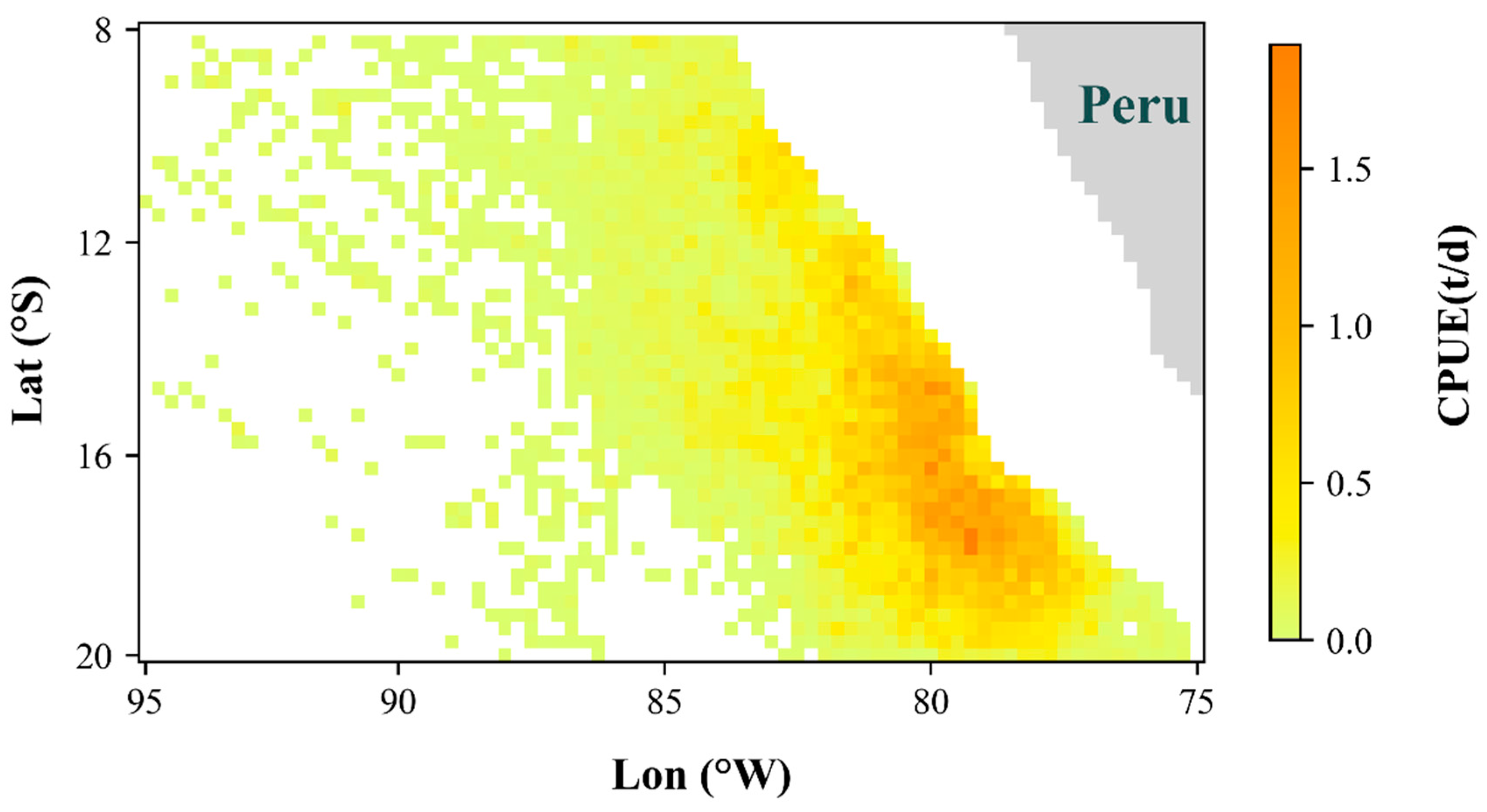
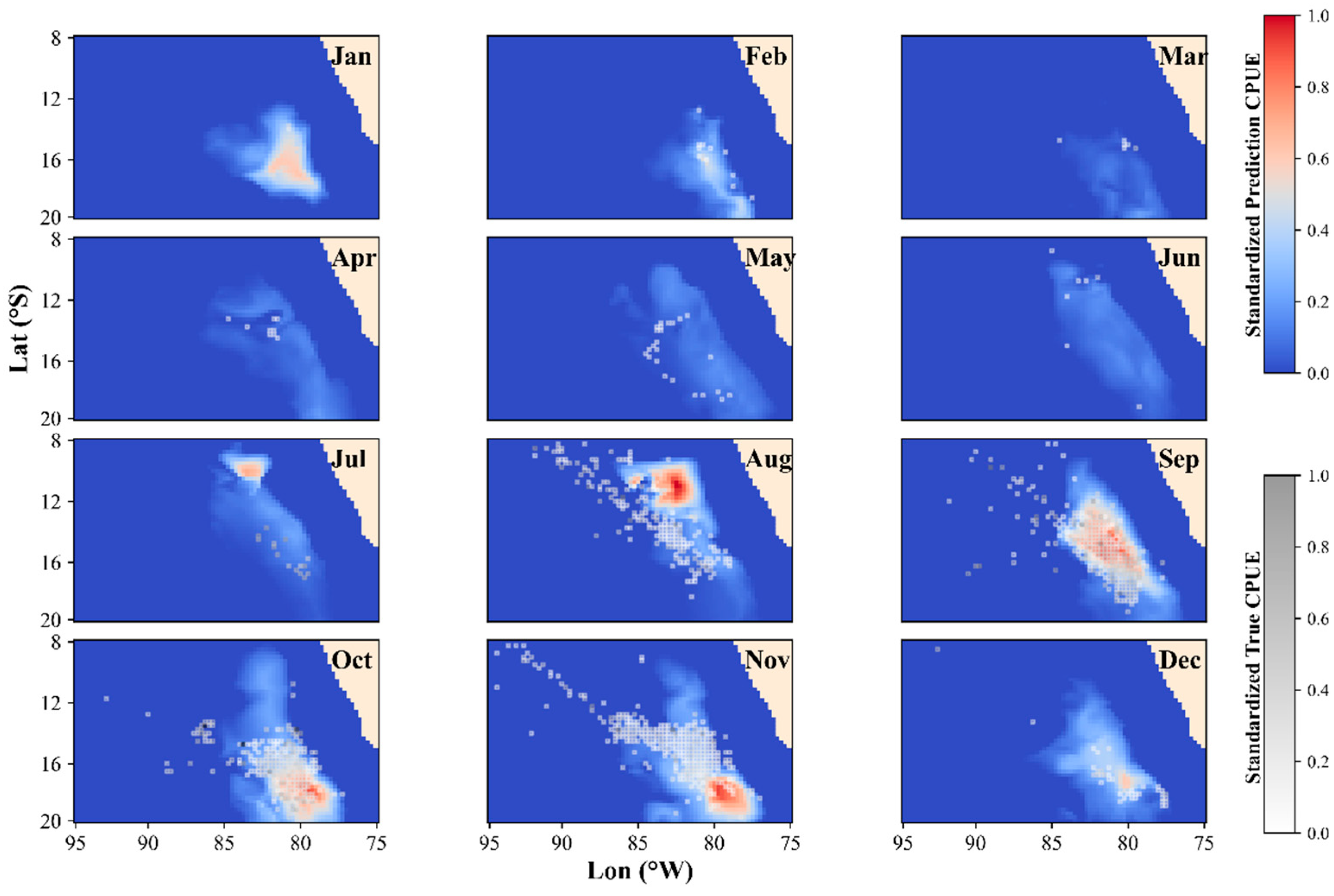
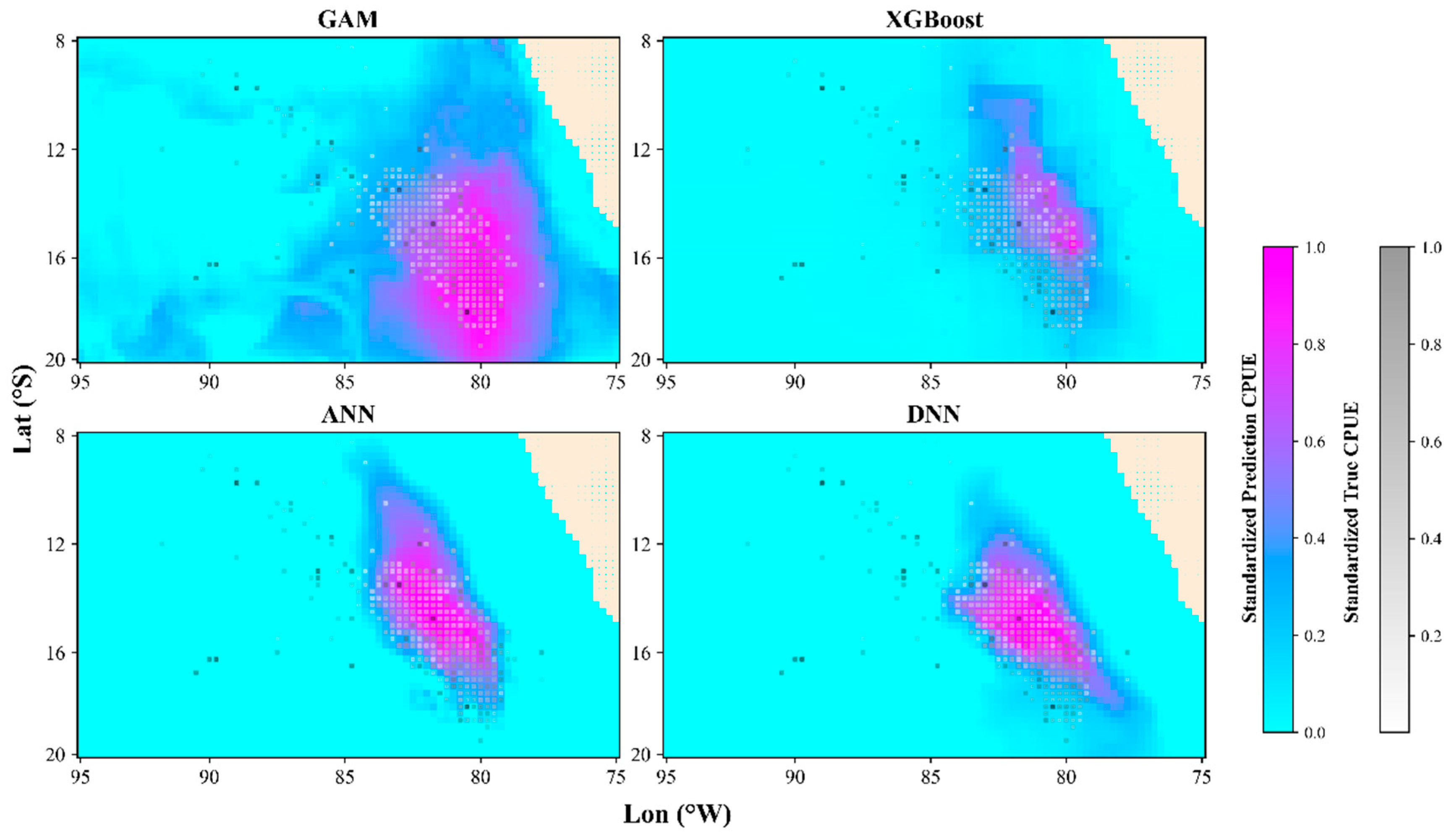
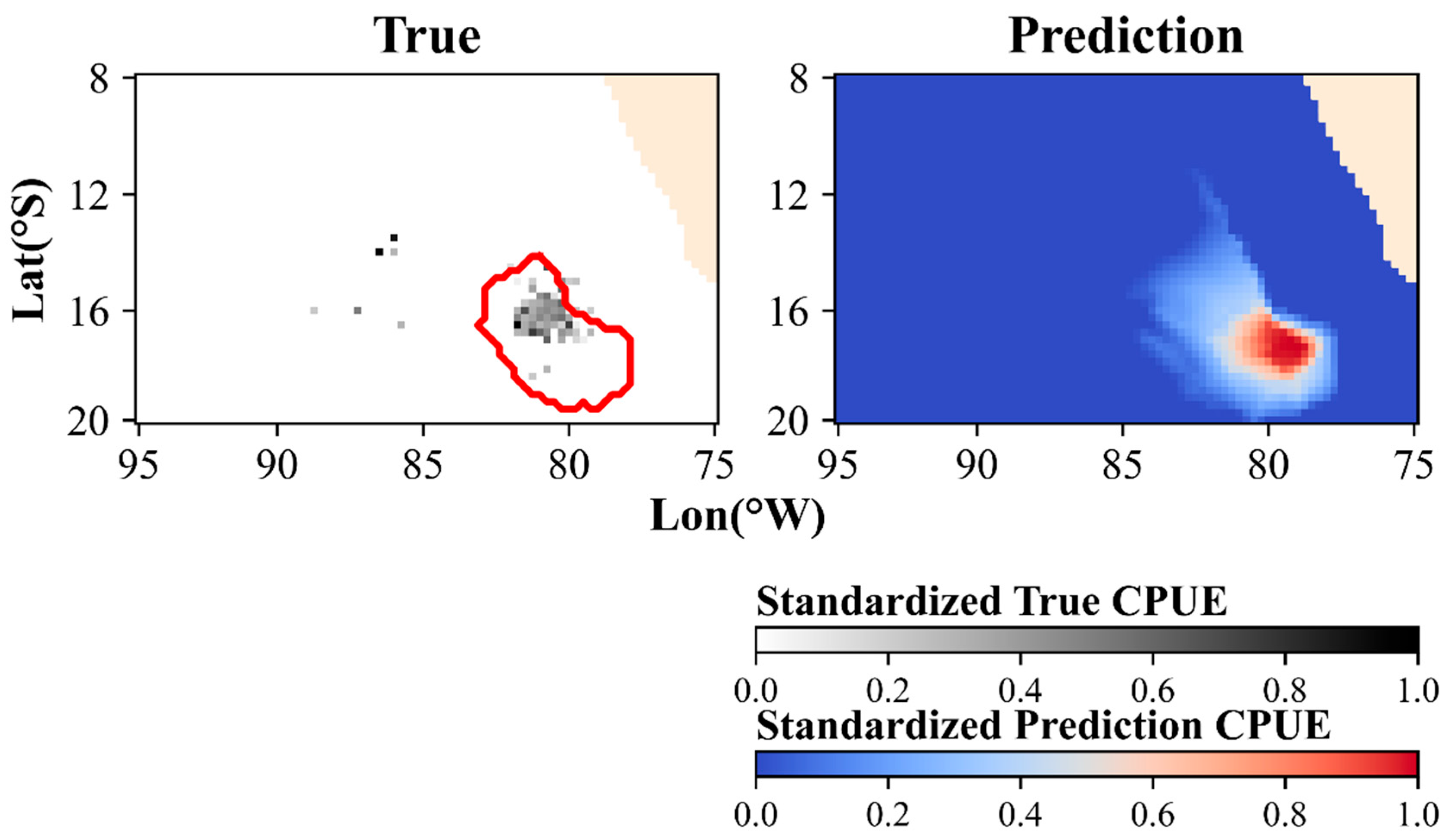
3.3. Spatiotemporal Distribution in the Optimal Model
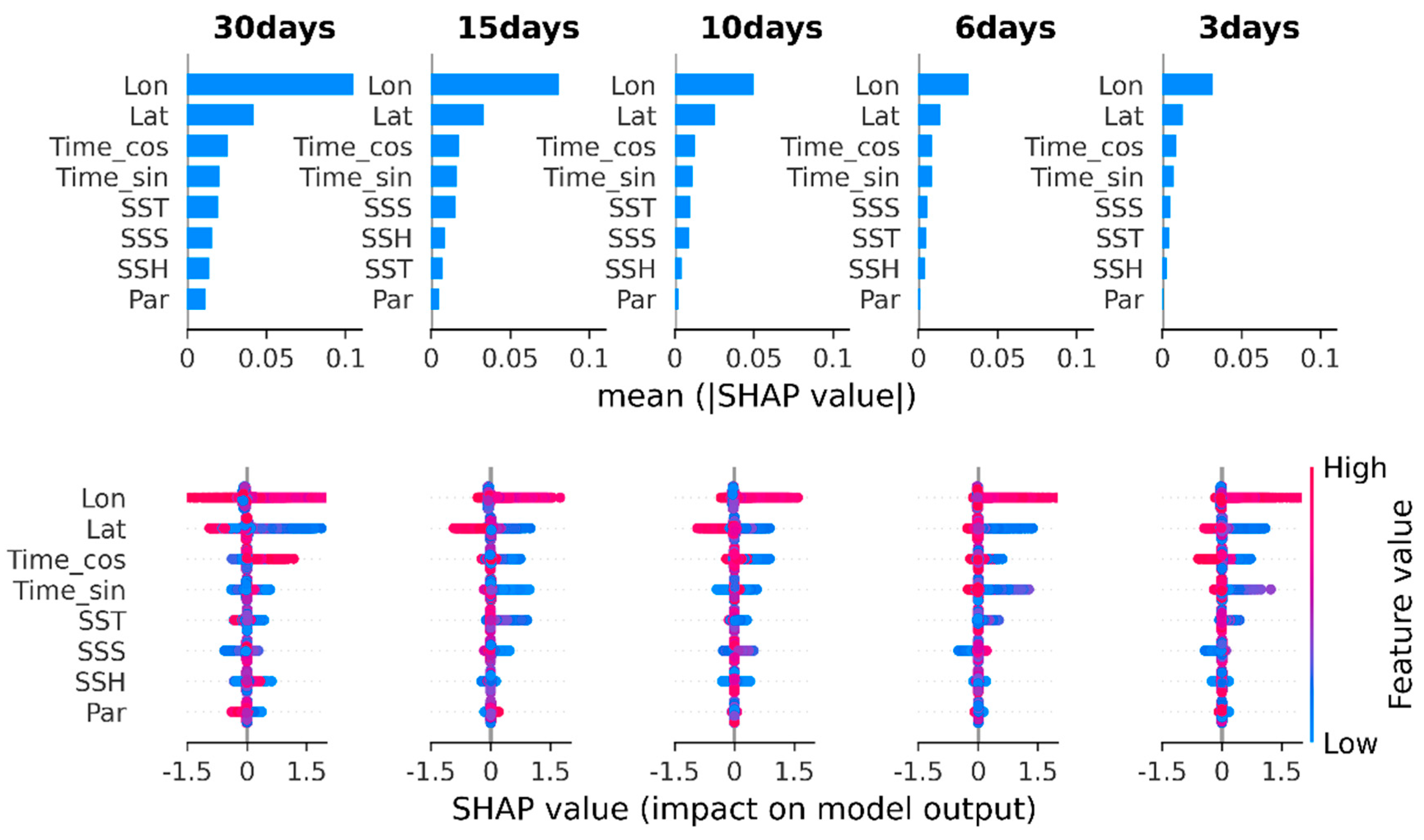
3.4. Shapley Additive Explanation of Model Predictions
4. Discussion
4.1. Performance Comparison of Different Models
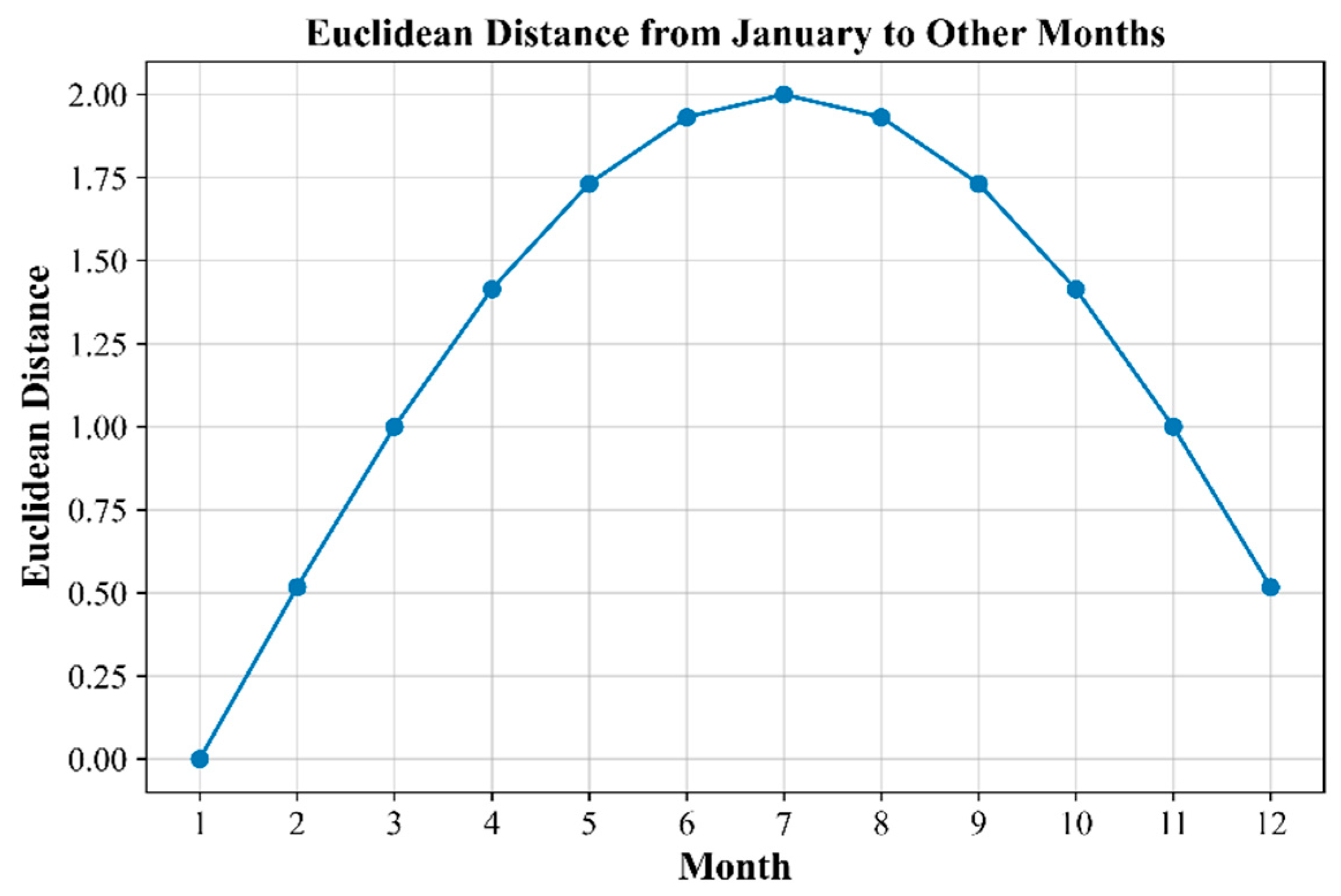
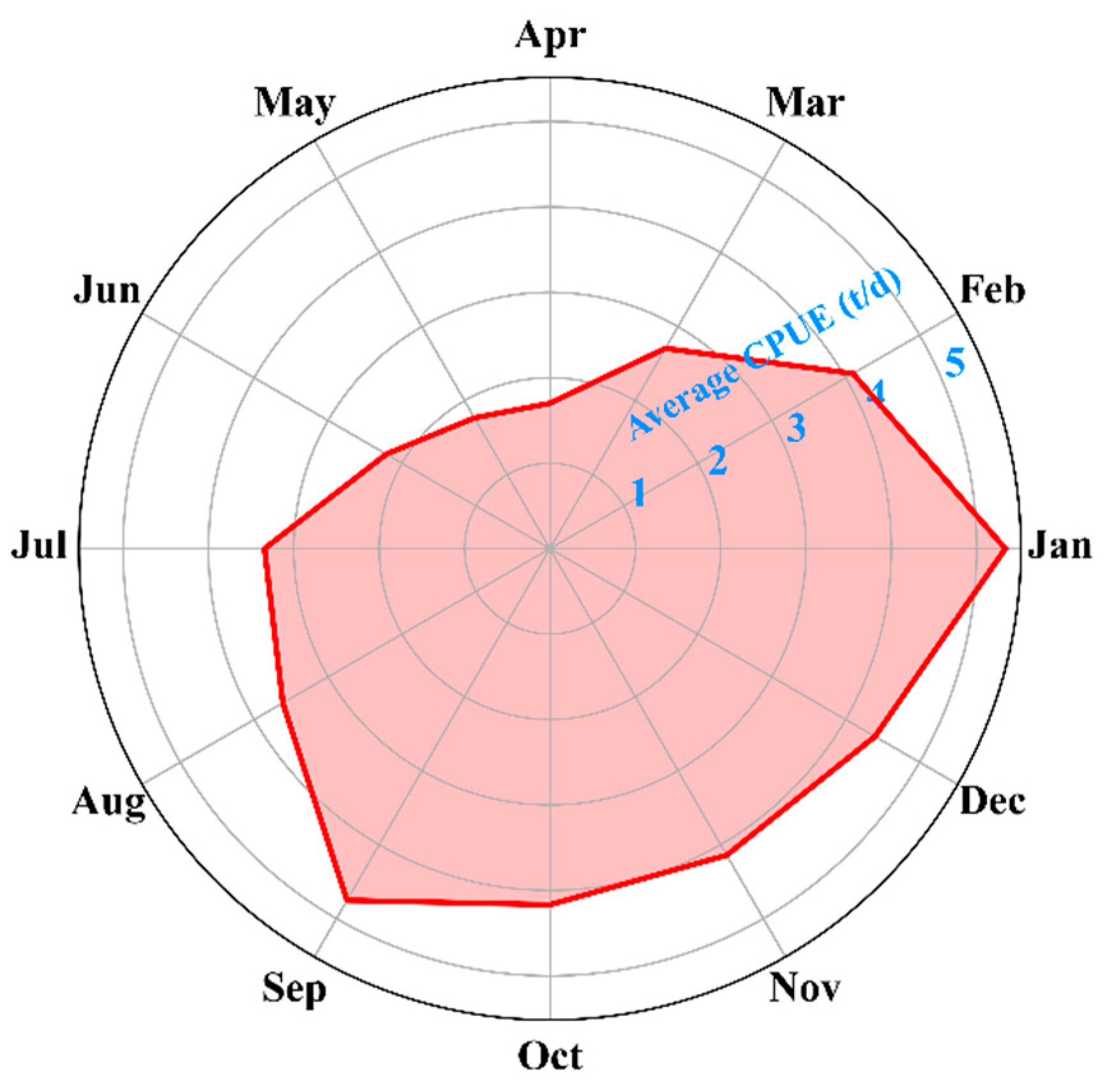
4.2. Differences Across Temporal Scales
4.3. Interpretation of Input Factors Effect
4.4. The Challenge of Presence-Only Data in SDM
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, X.; Liu, B.; Chen, Y. A review of the development of Chinese distant-water squid jigging fisheries. Fish. Res. 2008, 89, 211–221. [Google Scholar] [CrossRef]
- Rocha, F.; Vega, M.A. Overview of cephalopod fisheries in Chilean waters. Fish. Res. 2003, 60, 151–159. [Google Scholar] [CrossRef]
- Chen, X. Theory and Method of Fisheries Forecasting; Springer Nature: Singapore, 2022. [Google Scholar]
- Xiang, D.; Li, Y.; Jiang, K.; Han, H.; Wang, Y.; Yang, S.; Zhang, H.; Sun, Y. Environmental influences on Illex argentinus trawling grounds in the southwest Atlantic high seas. Fishes 2024, 9, 209. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X. Ocean warming-induced range-shifting of potential habitat for jumbo flying squid Dosidicus gigas in the Southeast Pacific Ocean off Peru. Fish. Res. 2018, 204, 137–146. [Google Scholar] [CrossRef]
- Tafur, R.; Keyl, F.; Argüelles, J. Reproductive biology of jumbo squid Dosidicus gigas in relation to environmental variability of the northern Humboldt Current System. Mar. Ecol. Prog. Ser. 2010, 400, 127–141. [Google Scholar] [CrossRef]
- Keyl, F.; Argüelles, J.U.A.N.; Mariategui, L.; Tafur, R.; Wolff, M.; Yamashiro, C. A hypothesis on range expansion and spatio-temporal shifts in size-at-maturity of jumbo squid (Dosidicus gigas) in the Eastern Pacific Ocean. CalCOFI Rep. 2008, 49, 119–128. [Google Scholar]
- Fang, X.; Zhang, Y.; Yu, W.; Chen, X. Geographical distribution variations of Humboldt squid habitat in the Eastern Pacific Ocean. Ecosyst. Health Sustain. 2023, 9, 10. [Google Scholar] [CrossRef]
- Wu, X.; Jin, P.; Zhang, Y.; Yu, W. Spatial Distribution and Abundance of a Pelagic Squid during the Evolution of Eddies in the Southeast Pacific Ocean. J. Mar. Sci. Eng. 2024, 12, 1015. [Google Scholar] [CrossRef]
- Yu, W.; Yi, Q.; Chen, X.; Chen, Y. Modelling the effects of climate variability on habitat suitability of jumbo flying squid, Dosidicus gigas, in the Southeast Pacific Ocean off Peru. ICES J. Mar. Sci. 2016, 73, 239–249. [Google Scholar] [CrossRef]
- Jia, S.; Bei, L.; Li, Y.; Zhao, Q. Spatiotemporal analysis of ocean primary productivity in Bohai Sea estimated using improved DINEOF reconstructed MODIS data. Ecol. Inform. 2024, 84, 102920. [Google Scholar] [CrossRef]
- Medellín-Ortiz, A.; Cadena-Cárdenas, L.; Santana-Morales, O. Environmental effects on the jumbo squid fishery along Baja California’s west coast. Fish. Sci. 2016, 82, 851–861. [Google Scholar] [CrossRef]
- Castillo, R.; Dalla Rosa, L.; García Diaz, W.; Madureira, L.; Gutierrez, M.; Vásquez, L.; Koppelmann, R. Anchovy distribution off Peru in relation to abiotic parameters: A 32-year time series from 1985 to 2017. Fish. Oceanogr. 2019, 28, 389–401. [Google Scholar] [CrossRef]
- Armas, E.; Arancibia, H.; Neira, S.; Marín, M.C. Neural network approach for detecting spatial changes in catch probability of Engraulis ringens during El Niño-Southern Oscillation events in northern Chile. Fish. Oceanogr. 2024, 33, e12672. [Google Scholar] [CrossRef]
- Poisson, F.; Ellis, J.R.; McCully Phillips, S.R. Preliminary Insights on the Habitat Use and Vertical Movements of the Pelagic Stingray (Pteroplatytrygon violacea) in the Western Mediterranean Sea. Fishes 2024, 9, 238. [Google Scholar] [CrossRef]
- Xie, M.; Liu, B.; Chen, X. Deep learning-based fishing ground prediction with multiple environmental factors. Mar. Life Sci. Technol. 2024, 6, 736–749. [Google Scholar] [CrossRef]
- Catalano, G.A.; D’Urso, P.R.; Arcidiacono, C. Predicting potential biomass production by geospatial modelling: The case study of citrus in a Mediterranean area. Ecol. Inform. 2024, 83, 102848. [Google Scholar] [CrossRef]
- Ribeiro, M.C.; Pinho, P.; Llop, E.; Branquinho, C.; Sousa, A.J.; Pereira, M.J. Multivariate geostatistical methods for analysis of relationships between ecological indicators and environmental factors at multiple spatial scales. Ecol. Indic. 2013, 29, 339–347. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Li, Y.; Boenish, R. The effects of climate-induced environmental variability on Pacific Ocean squids. ICES J. Mar. Sci. 2023, 80, 878–888. [Google Scholar] [CrossRef]
- Li, X.; Liu, B.; Zheng, G.; Ren, Y.; Zhang, S.; Liu, Y.; Gao, L.; Liu, Y.; Zhang, B.; Wang, F. Deep-learning-based information mining from ocean remote-sensing imagery. Natl. Sci. Rev. 2020, 7, 1584–1605. [Google Scholar] [CrossRef]
- Rubbens, P.; Brodie, S.; Cordier, T.; Destro Barcellos, D.; Devos, P.; Fernandes-Salvador, J.A.; Fincharm, J.I.; Gomes, A.; Handegard, N.O.; Howell, K.; et al. Machine learning in marine ecology: An overview of techniques and applications. ICES J. Mar. Sci. 2023, 80, 1829–1853. [Google Scholar] [CrossRef]
- Landy, J.C.; Dawson, G.J.; Tsamados, M.; Bushuk, M.; Stroeve, J.C.; Howell, S.E.; Krumpen, T.; Babb, D.G.; Komarov, A.S.; Heorton, H.D.; et al. A year-round satellite sea-ice thickness record from CryoSat-2. Nature 2022, 609, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Tupan, J.M.; Rieuwpassa, F.; Setha, B.; Latuny, W.; Goesniady, S. A Deep Learning Approach to Automated Treatment Classification in Tuna Processing: Enhancing Quality Control in Indonesian Fisheries. Fishes 2025, 10, 75. [Google Scholar] [CrossRef]
- Kroodsma, D.A.; Mayorga, J.; Hochberg, T.; Miller, N.A.; Boerder, K.; Ferretti, F.; Wilson, A.; Bergman, B.; White, T.D.; Block, B.A.; et al. Tracking the global footprint of fisheries. Science 2018, 359, 904–908. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, S.; Tang, F.; Shi, Y.; Wu, Y.; He, J.; Chen, Y.; Li, L. Behavior Recognition of Squid Jigger Based on Deep Learning. Fishes 2023, 8, 502. [Google Scholar] [CrossRef]
- Wu, X.; Jin, P.; Zhang, Y.; Yu, W. Changing Humboldt Squid Abundance and Distribution at Different Stages of Oceanic Mesoscale Eddies. J. Mar. Sci. Eng. 2024, 12, 626. [Google Scholar] [CrossRef]
- Yu, W.; Feng, X.; Wen, J.; Wu, X.; Fang, X.; Cui, J.; Feng, Z.; Sheng, Y.; Zhao, Z.; Liu, B.; et al. The potential impacts of climate change on the life history and habitat of jumbo flying squid in the southeast Pacific Ocean: Overview and implications for fisheries management. Rev. Fish Biol. Fisher. 2025, 35, 707–731. [Google Scholar] [CrossRef]
- Tian, S.; Chen, X.; Chen, Y.; Xu, L.; Dai, X. Standardizing CPUE of Ommastrephes bartramii for Chinese squid-jigging fishery in Northwest Pacific Ocean. Chin. J. Oceanol. Limn. 2009, 27, 729–739. [Google Scholar] [CrossRef]
- Paulino, C.; Segura, M.; Chacón, G. Spatial variability of jumbo flying squid (Dosidicus gigas) fishery related to remotely sensed SST and chlorophyll-a concentration (2004–2012). Fish. Res. 2016, 173, 122–127. [Google Scholar] [CrossRef]
- Xie, Y.; Zhong, Z.; Li, B.; Xie, Y.; Chen, L.; Chen, H. An ARM-FPGA Hybrid Acceleration and Fault Tolerant Technique for Phase Factor Calculation in Spaceborne Synthetic Aperture Radar Imaging. IEEE J. Sel. Top. Appl. Earth Obs. Remote. Sens. 2024, 17, 5059–5072. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, X.; Yang, S.; Dai, Y.; Cui, X.; Wu, Y.; Zhang, S.; Fan, W.; Han, H.; Zhang, H.; et al. Construction of CPUE standardization model and its simulation testing for chub mackerel (Scomber japonicus) in the Northwest Pacific Ocean. Ecol. Indic. 2023, 155, 111022. [Google Scholar] [CrossRef]
- Uzer, U. Influence of Environmental Parameters on the Abundance of Tub Gurnard, Chelidonichthys lucerna, in the Eastern Sea of Marmara. Fishes 2025, 10, 127. [Google Scholar] [CrossRef]
- Wang, L.; Yang, C.; Shan, B.; Liu, Y.; Zou, J.; Sun, D.; Guo, T. Spatiotemporal Variation and Predictors of the Purpleback Flying Squid (Sthenoteuthis oualaniensis) Distribution Surrounding the Xisha and Zhongsha Islands during a Fishing Moratorium. Fishes 2024, 9, 253. [Google Scholar] [CrossRef]
- Hamzaoui, M.; Aoueileyine, M.O.E.; Romdhani, L.; Bouallegue, R. Optimizing XGBoost performance for fish weight prediction through parameter pre-selection. Fishes 2023, 8, 505. [Google Scholar] [CrossRef]
- Xing, B.; Zhang, L.; Liu, Z.; Sheng, H.; Bi, F.; Xu, J. The study of fishing vessel behavior identification based on ais data: A case study of the east China sea. J. Mar. Sci. Eng. 2023, 11, 1093. [Google Scholar] [CrossRef]
- Xu, S.; Wang, J.; Chen, X.; Zhu, J. Identifying optimal variables for machine-learning-based fish distribution modeling. Can. J. Fish. Aquat. Sci. 2024, 81, 687–698. [Google Scholar] [CrossRef]
- Chen, B.; Mu, X.; Chen, P.; Wang, B.; Choi, J.; Park, H.; Xu, S.; Wu, Y.; Yang, H. Machine learning-based inversion of water quality parameters in typical reach of the urban river by UAV multispectral data. Ecol. Indic. 2021, 133, 108434. [Google Scholar] [CrossRef]
- Yang, W.; Fu, B.; Li, S.; Lao, Z.; Deng, T.; He, W.; He, H.; Chen, Z. Monitoring multi-water quality of internationally important karst wetland through deep learning, multi-sensor and multi-platform remote sensing images: A case study of Guilin, China. Ecol. Indic. 2023, 154, 110755. [Google Scholar] [CrossRef]
- Lin, H.; Wang, J.; Zhu, J.; Chen, X. Evaluating the impacts of environmental and fishery variability on the distribution of bigeye tuna in the Pacific Ocean. ICES J. Mar. Sci. 2023, 80, 2642–2656. [Google Scholar] [CrossRef]
- Kunimatsu, S.; Kurota, H.; Muko, S.; Ohshimo, S.; Tomiyama, T. Predicting unseen chub mackerel densities through spatiotemporal machine learning: Indications of potential hyperdepletion in catch-per-unit-effort due to fishing ground contraction. Ecol. Inform. 2024, 85, 102944. [Google Scholar] [CrossRef]
- Zou, S.; Zhang, L.; Huang, X.; Osei, F.B.; Ou, G. Early ecological security warning of cultivated lands using RF-MLP integration model: A case study on China’s main grain-producing areas. Ecol. Indic. 2022, 141, 109059. [Google Scholar] [CrossRef]
- Burton, M.L.; Potts, J.C.; Ostrowski, A.D. Preliminary estimates of age, growth and natural mortality of margate, Haemulon album, and black margate, Anisotremus surinamensis, from the southeastern United States. Fishes 2019, 4, 44. [Google Scholar] [CrossRef]
- Gu, K.; Chen, Y. YOLOv3-MSSA based hot spot defect detection for photovoltaic power stations. J. Meas. Eng. 2024, 12, 23–39. [Google Scholar] [CrossRef]
- Štrumbelj, E.; Kononenko, I. Explaining prediction models and individual predictions with feature contributions. Knowl. Inf. Syst. 2014, 41, 647–665. [Google Scholar] [CrossRef]
- Shrikumar, A.; Greenside, P.; Kundaje, A. Learning important features through propagating activation differences. In Proceedings of the International Conference on Machine Learning, Sydney, Australia, 6–11 August 2017; pp. 3145–3153. [Google Scholar]
- Jamei, M.; Ali, M.; Malik, A.; Rai, P.; Karbasi, M.; Farooque, A.A.; Yaseen, Z.M. Designing a decomposition-based multi-phase pre-processing strategy coupled with EDBi-LSTM deep learning approach for sediment load forecasting. Ecol. Indic. 2023, 153, 110478. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, M.; Liu, K. Deep neural networks for global wildfire susceptibility modelling. Ecol. Indic. 2021, 127, 107735. [Google Scholar] [CrossRef]
- Han, H.; Yang, C.; Jiang, B.; Shang, C.; Sun, Y.; Zhao, X.; Xiang, D.; Zhang, H.; Shi, Y. Construction of chub mackerel (Scomber japonicus) fishing ground prediction model in the northwestern Pacific Ocean based on deep learning and marine environmental variables. Mar. Pollut. Bull. 2023, 193, 115158. [Google Scholar] [CrossRef]
- Moyano, G.; Plaza, G.; Cerna, F.; Muñoz, A.A. Local and global environmental drivers of growth chronologies in a demersal fish in the south-eastern pacific ocean. Ecol. Indic. 2021, 131, 108151. [Google Scholar] [CrossRef]
- Reichstein, M.; Camps-Valls, G.; Stevens, B.; Jung, M.; Denzler, J.; Carvalhais, N.; Prabhat, F. Deep learning and process understanding for data-driven Earth system science. Nature 2019, 566, 195–204. [Google Scholar] [CrossRef]
- Anderson, C.I.; Rodhouse, P.G. Life cycles, oceanography and variability: Ommastrephid squid in variable oceanographic environments. Fish. Res. 2001, 54, 133–143. [Google Scholar] [CrossRef]
- Tafur, R.; Villegas, P.; Rabí, M.; Yamashiro, C. Dynamics of maturation, seasonality of reproduction and spawning grounds of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) in Peruvian waters. Fish. Res. 2001, 54, 33–50. [Google Scholar] [CrossRef]
- Martínez-Minaya, J.; Cameletti, M.; Conesa, D.; Pennino, M.G. Species distribution modeling: A statistical review with focus in spatio-temporal issues. Stoch. Environ. Res. Risk A 2018, 32, 3227–3244. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Iturbide, M.; Bedia, J.; Herrera, S.; del Hierro, O.; Pinto, M.; Gutiérrez, J.M. A framework for species distribution modelling with improved pseudo-absence generation. Ecol. Model. 2015, 312, 166–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temporal Scale | 3 Days | 6 Days | 10 Days | 15 Days | 30 Days |
|---|---|---|---|---|---|
| Numbers of periods from 2012 to 2021 | 1200 | 600 | 360 | 240 | 120 |
| Data volume (×103) | 4608 | 2304 | 1382.4 | 921.6 | 460.8 |
| Cases Types | SST | SSH | SSS | PAR |
|---|---|---|---|---|
| Case 1 | √ | |||
| Case 2 | √ | √ | ||
| Case 3 | √ | √ | ||
| Case 4 | √ | √ | ||
| Case 5 | √ | √ | √ | |
| Case 6 | √ | √ | √ | |
| Case 7 | √ | √ | √ | |
| Case 8 | √ | √ | √ | √ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, M.; Liu, B.; Chen, X.; Yu, W.; Wang, J.; Xu, J. Modelling the Spatial Distribution of Dosidicus gigas in the Southeast Pacific Ocean at Multiple Temporal Scales Based on Deep Learning. Fishes 2025, 10, 273. https://doi.org/10.3390/fishes10060273
Xie M, Liu B, Chen X, Yu W, Wang J, Xu J. Modelling the Spatial Distribution of Dosidicus gigas in the Southeast Pacific Ocean at Multiple Temporal Scales Based on Deep Learning. Fishes. 2025; 10(6):273. https://doi.org/10.3390/fishes10060273
Chicago/Turabian StyleXie, Mingyang, Bin Liu, Xinjun Chen, Wei Yu, Jintao Wang, and Jiawen Xu. 2025. "Modelling the Spatial Distribution of Dosidicus gigas in the Southeast Pacific Ocean at Multiple Temporal Scales Based on Deep Learning" Fishes 10, no. 6: 273. https://doi.org/10.3390/fishes10060273
APA StyleXie, M., Liu, B., Chen, X., Yu, W., Wang, J., & Xu, J. (2025). Modelling the Spatial Distribution of Dosidicus gigas in the Southeast Pacific Ocean at Multiple Temporal Scales Based on Deep Learning. Fishes, 10(6), 273. https://doi.org/10.3390/fishes10060273