


Ichthyofaunal Metabarcoding in the Southern Appalachians: Use of eDNA Metabarcoding in Fish Surveys in Lotic Systems of the Great Smoky Mountains National Park with Comparisons to Historic Electrofishing Data

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. eDNA Field Sampling
2.2. DNA Extraction
2.3. Inhibition Testing
2.4. Primer Selection and Evaluation
2.5. PCR Amplification and Sequencing
2.6. Bioinformatics
2.7. Data Analysis
2.8. Comparison of eDNA Metabarcoding Data with Traditional Monitoring Data
3. Results
3.1. Primer Validation
3.2. Sequencing Results
3.3. Species Richness
3.4. Assemblage Structure by Elevation and Stream Order
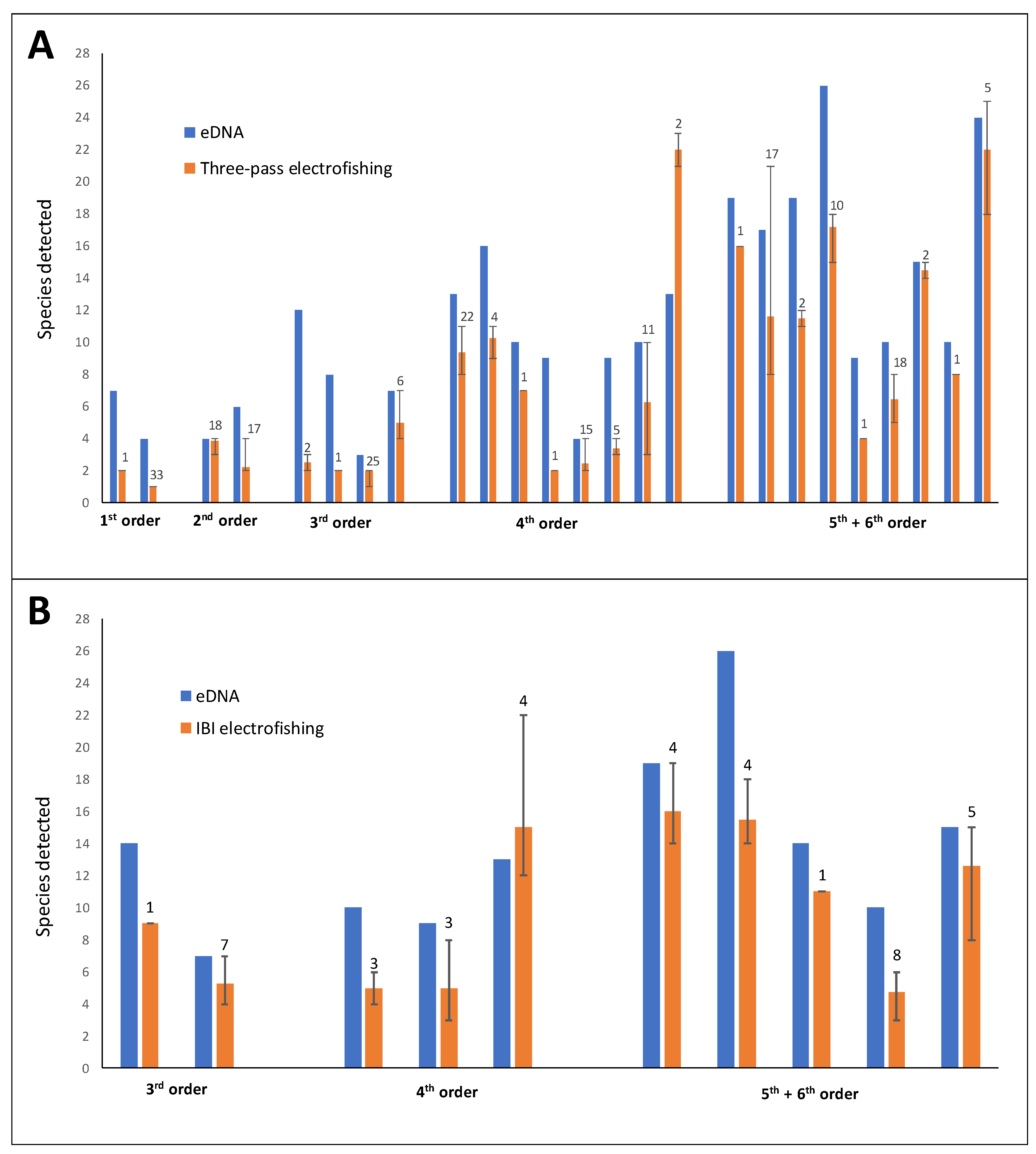
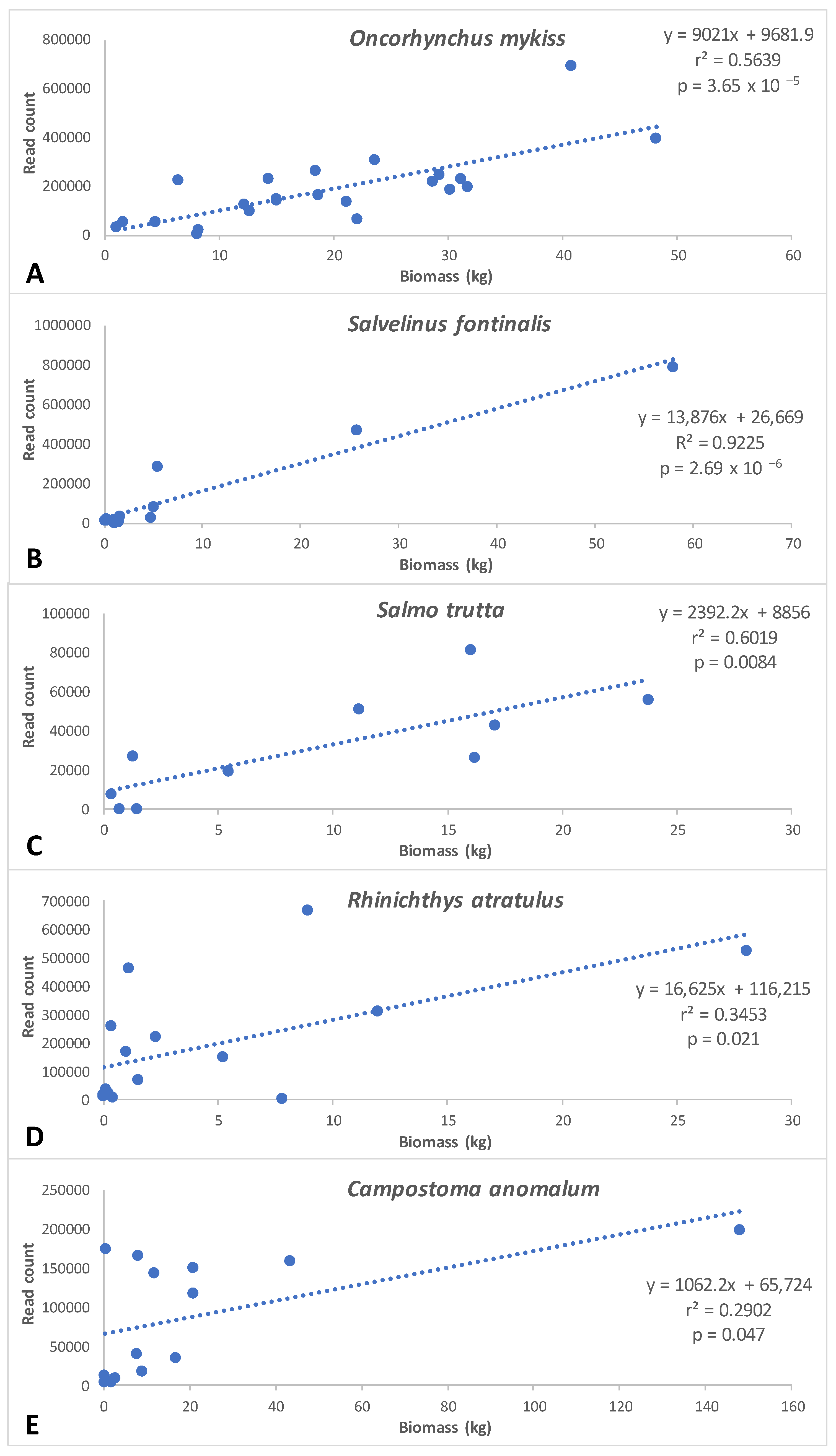
3.5. Comparison of eDNA Metabarcoding Data with Traditional Monitoring Data
4. Discussion
4.1. Primer Performance
4.2. Metabarcoding Critical Parameters
4.3. Comparison with Electrofishing Data
4.4. Detection of Unexpected Species
4.5. Community Composition Observations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species Detection Using Environmental DNA from Water Samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Saccò, M.; Kestel, J.H.; Nester, G.; Campbell, M.A.; van der Heyde, M.; Heydenrych, M.J.; Juszkiewicz, D.J.; Nevill, P.; Dawkins, K.L.; et al. Aquatic Environmental DNA: A Review of the Macro-Organismal Biomonitoring Revolution. Sci. Total Environ. 2023, 873, 162322. [Google Scholar] [CrossRef]
- Blackman, R.; Couton, M.; Keck, F.; Kirschner, D.; Carraro, L.; Cereghetti, E.; Perrelet, K.; Bossart, R.; Brantschen, J.; Zhang, Y.; et al. Environmental DNA: The next Chapter. Mol. Ecol. 2024, 33, e17355. [Google Scholar] [CrossRef]
- Bunce, M.; Freeth, A. Looking Further and Deeper into Environmental Protection, Regulation and Policy Using Environmental DNA (eDNA). Policy Q. 2022, 18, 33–39. [Google Scholar] [CrossRef]
- Lodge, D.M. Policy Action Needed to Unlock eDNA Potential. Front. Ecol. Environ. 2022, 20, 448–449. [Google Scholar] [CrossRef]
- Kelly, R.P.; Lodge, D.M.; Lee, K.N.; Theroux, S.; Sepulveda, A.J.; Scholin, C.A.; Craine, J.M.; Andruszkiewicz Allan, E.; Nichols, K.M.; Parsons, K.M.; et al. Toward a National eDNA Strategy for the United States. Environ. DNA 2024, 6, e432. [Google Scholar] [CrossRef]
- Buchner, D.; Sinclair, J.S.; Ayasse, M.; Beermann, A.J.; Buse, J.; Dziock, F.; Enss, J.; Frenzel, M.; Hörren, T.; Li, Y.; et al. Upscaling Biodiversity Monitoring: Metabarcoding Estimates 31,846 Insect Species from Malaise Traps across Germany. Mol. Ecol. Resour. 2025, 25, e14023. [Google Scholar] [CrossRef]
- Liu, M.; Clarke, L.J.; Baker, S.C.; Jordan, G.J.; Burridge, C.P. A Practical Guide to DNA Metabarcoding for Entomological Ecologists. Ecol. Entomol. 2020, 45, 373–385. [Google Scholar]
- Marquina, D.; Andersson, A.F.; Ronquist, F. New Mitochondrial Primers for Metabarcoding of Insects, Designed and Evaluated Using in Silico Methods. Mol. Ecol. Resour. 2019, 19, 90–104. [Google Scholar] [CrossRef]
- de Santana, C.D.; Parenti, L.R.; Dillman, C.B.; Coddington, J.A.; Bastos, D.A.; Baldwin, C.C.; Zuanon, J.; Torrente-Vilara, G.; Covain, R.; Menezes, N.A.; et al. The Critical Role of Natural History Museums in Advancing eDNA for Biodiversity Studies: A Case Study with Amazonian Fishes. Sci. Rep. 2021, 11, 18159. [Google Scholar] [CrossRef]
- Leduc, N.; Lacoursière-Roussel, A.; Howland, K.L.; Archambault, P.; Sevellec, M.; Normandeau, E.; Dispas, A.; Winkler, G.; McKindsey, C.W.; Simard, N.; et al. Comparing eDNA Metabarcoding and Species Collection for Documenting Arctic Metazoan Biodiversity. Environ. DNA 2019, 1, 342–358. [Google Scholar] [CrossRef]
- Shen, M.; Xiao, N.; Zhao, Z.; Guo, N.; Luo, Z.; Sun, G.; Li, J. eDNA Metabarcoding as a Promising Conservation Tool to Monitor Fish Diversity in Beijing Water Systems Compared with Ground Cages. Sci. Rep. 2022, 12, 11113. [Google Scholar] [CrossRef] [PubMed]
- Macher, T.H.; Schütz, R.; Yildiz, A.; Beermann, A.J.; Leese, F. Evaluating Five Primer Pairs for Environmental DNA Metabarcoding of Central European Fish Species Based on Mock Communities. Metabarcoding Metagenom. 2023, 7, e103856. [Google Scholar] [CrossRef]
- Beng, K.C.; Corlett, R.T. Applications of Environmental DNA (eDNA) in Ecology and Conservation: Opportunities, Challenges and Prospects. Biodivers. Conserv. 2020, 29, 2089–2121. [Google Scholar] [CrossRef]
- Wang, S.; Yan, Z.; Hänfling, B.; Zheng, X.; Wang, P.; Fan, J.; Li, J. Methodology of Fish eDNA and Its Applications in Ecology and Environment. Sci. Total Environ. 2021, 755, 142622. [Google Scholar] [CrossRef] [PubMed]
- Schenekar, T. The Current State of eDNA Research in Freshwater Ecosystems: Are We Shifting from the Developmental Phase to Standard Application in Biomonitoring? Hydrobiologia 2022, 850, 1263–1282. [Google Scholar] [CrossRef]
- Cortelezzi, A.; Paz, L.E. Macroinvertebrate Biomonitoring in Latin America: Progress and Challenges. Freshw. Sci. 2023, 42, 204–213. [Google Scholar] [CrossRef]
- De Brauwer, M.; Clarke, L.J.; Chariton, A.; Cooper, M.K.; de Bruyn, M.; Furlan, E.; MacDonald, A.J.; Rourke, M.L.; Sherman, C.D.H.; Suter, L.; et al. Best Practice Guidelines for Environmental DNA Biomonitoring in Australia and New Zealand. Environ. DNA 2023, 5, 417–423. [Google Scholar] [CrossRef]
- Jenkins, M.A. Vegetation Communities of Great Smoky Mountains National Park. Southeast. Nat. 2007, 6, 35–56. [Google Scholar] [CrossRef]
- Lennon, R.E. An Annotated List of the Fishes of the Great Smoky Mountains National Park. J. Tenn Acad. Sci. 1962, 37, 5–7. [Google Scholar]
- Simbeck, D.J. Distribution of the Fishes of The Great Smoky Mountains National Distribution of the Fishes of The Great Smoky Mountains National Park. Master Thesis, University of Tennessee, Knoxville, TN, USA, December 1990. [Google Scholar]
- Bates, P.C.; Miller, J.R.; Styers, D.M.; Langdon, K.; Burda, C.; Davis, R.; Martin, T.; Kloeppel, B.D.; McFarland, S. Natural Resource Condition Assessment Great Smoky Mountains National Park; Natural Resource Report NPS/GRSM/NRR; National Park Service: Fort Collins, CO, USA, 2018. [Google Scholar]
- Kulp, M.A.; Moore, S.E. A Case History in Fishing Regulations in Great Smoky Mountains National Park: 1934–2004. N. Am. J. Fish. Manag. 2005, 25, 510–524. [Google Scholar] [CrossRef]
- Baldigo, B.P.; Kulp, M.A.; Schwartz, J.S. Relationships between Indicators of Acid-Base Chemistry and Fish Assemblages in Streams of the Great Smoky Mountains National Park. Ecol. Indic. 2018, 88, 465–484. [Google Scholar] [CrossRef]
- Deyton, E.B.; Schwartz, J.S.; Robinson, R.B.; Neff, K.J.; Moore, S.E.; Kulp, M.A. Characterizing Episodic Stream Acidity during Stormflows in the Great Smoky Mountains National Park. Water Air Soil. Pollut. 2009, 196, 3–18. [Google Scholar] [CrossRef]
- Neff, K.J.; Schwartz, J.S.; Henry, T.B.; Bruce Robinson, R.; Moore, S.E.; Kulp, M.A. Physiological Stress in Native Southern Brook Trout during Episodic Stream Acidification in the Great Smoky Mountains National Park. Arch. Environ. Contam. Toxicol. 2009, 57, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Lennon, R.E.; Parker, P.S. The Stoneroller, Campostoma anomalum (Rafinesque), in Great Smoky Mountains National Park. Trans. Am. Fish. Soc. 1960, 89, 263–270. [Google Scholar] [CrossRef]
- King, W. A Program for the Management of Fish Resources in Great Smoky Mountains National Park. Trans. Am. Fish. Soc. 1939, 68, 86–95. [Google Scholar] [CrossRef]
- Fediajevaite, J.; Priestley, V.; Arnold, R.; Savolainen, V. Meta-Analysis Shows That Environmental DNA Outperforms Traditional Surveys, but Warrants Better Reporting Standards. Ecol. Evol. 2021, 11, 4803–4815. [Google Scholar] [CrossRef]
- McElroy, M.E.; Dressler, T.L.; Titcomb, G.C.; Wilson, E.A.; Deiner, K.; Dudley, T.L.; Eliason, E.J.; Evans, N.T.; Gaines, S.D.; Lafferty, K.D.; et al. Calibrating Environmental DNA Metabarcoding to Conventional Surveys for Measuring Fish Species Richness. Front. Ecol. Evol. 2020, 8, 276. [Google Scholar] [CrossRef]
- Yates, M.C.; Wilcox, T.M.; Kay, S.; Heath, D.D. A General Framework to Unify the Estimation of Numerical Abundance and Biomass from Quantitative EDNA Data. Environmental DNA. 2025, 7, e70073. [Google Scholar] [CrossRef]
- Jerde, C.L. Can We Manage Fisheries with the Inherent Uncertainty from eDNA? J. Fish. Biol. 2021, 98, 341–353. [Google Scholar]
- Keck, F.; Blackman, R.C.; Bossart, R.; Brantschen, J.; Couton, M.; Hürlemann, S.; Kirschner, D.; Locher, N.; Zhang, H.; Altermatt, F. Meta-Analysis Shows Both Congruence and Complementarity of DNA and eDNA Metabarcoding to Traditional Methods for Biological Community Assessment. Mol. Ecol. 2022, 31, 1820–1835. [Google Scholar] [CrossRef]
- Stoeckle, B.C.; Beggel, S.; Cerwenka, A.F.; Motivans, E.; Kuehn, R.; Geist, J. A Systematic Approach to Evaluate the Influence of Environmental Conditions on eDNA Detection Success in Aquatic Ecosystems. PLoS ONE 2017, 12, e0189119. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Hermans, S.M.; Lear, G.; Buckley, T.R.; Lee, K.C.; Buckley, H.L. A Systematic Review of Sources of Variability and Uncertainty in eDNA Data for Environmental Monitoring. Front. Ecol. Evol. 2020, 8, 135. [Google Scholar] [CrossRef]
- Baldigo, B.P.; Sporn, L.A.; George, S.D.; Ball, J.A. Efficacy of Environmental DNA to Detect and Quantify Brook Trout Populations in Headwater Streams of the Adirondack Mountains, New York. Trans. Am. Fish. Soc. 2017, 146, 99–111. [Google Scholar] [CrossRef]
- Yates, M.C.; Wilcox, T.M.; McKelvey, K.S.; Young, M.K.; Schwartz, M.K.; Derry, A.M. Allometric Scaling of eDNA Production in Stream-Dwelling Brook Trout (Salvelinus Fontinalis) Inferred from Population Size Structure. Environ. DNA 2021, 3, 553–560. [Google Scholar] [CrossRef]
- Urabe, H.; Mizumoto, H.; Tsuda-Yamaguchi, F.; Araki, H. Spatial Heterogeneity of eDNA Concentration as a Predictor of Small Biomass of Fish in a Mountain Stream. Limn 2024, 26, 223–233. [Google Scholar] [CrossRef]
- Aunins, A.W.; Eackles, M.S.; Super, P.E.; Kulp, M.A.; Nichols, B.J.; Lubinski, B.A.; Morrison, C.L.; King, T.L. Development of a DdPCR Assay for the Detection of the Smoky Madtom (Noturus baileyi) from eDNA in Stream Water Samples. Conserv. Genet. Resour. 2022, 14, 29–435. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, C.; You, W.; Li, S.; Wu, Y.; Liang, Y.; Chu, L.; Yan, Y.; Zhang, C. Comparison Between Environmental DNA Metabarcoding and Traditional Survey Method to Identify Community Composition and Assembly of Stream Fish. Ecol. Evol. 2024, 14, e70627. [Google Scholar] [CrossRef]
- Lee, V.M.; Berkman, L.K.; Geheber, A.D.; Landwer, B.; Ludwig, E.J.; Duvernell, D.D. Putting eDNA to the Test: A Field Comparison of eDNA Metabarcoding to Established Protocols for Assessing Biodiversity in Missouri’s Ozark Highland Streams. Environ. DNA 2024, 6, e510. [Google Scholar] [CrossRef]
- Strahler, A.N. Quantitative Analysis of Watershed Geomorphology. Eos Trans. Am. Geophys. Union 1957, 38, 913–920. [Google Scholar] [CrossRef]
- Majaneva, M.; Diserud, O.H.; Eagle, S.H.C.; Boström, E.; Hajibabaei, M.; Ekrem, T. Environmental DNA Filtration Techniques Affect Recovered Biodiversity. Sci. Rep. 2018, 8, 4682. [Google Scholar] [CrossRef]
- Hinlo, R.; Gleeson, D.; Lintermans, M.; Furlan, E. Methods to Maximize Recovery of Environmental DNA from Water Samples. PLoS ONE 2017, 12, e0179251. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Pilliod, D.S.; Arkle, R.S.; Waits, L.P. Molecular Detection of Vertebrates in Stream Water: A Demonstration Using Rocky Mountain Tailed Frogs and Idaho Giant Salamanders. PLoS ONE 2011, 6, e22746. [Google Scholar] [CrossRef]
- Carvalho, C.O.; Gromstad, W.; Dunthorn, M.; Karlsen, H.E.; Schrøder-Nielsen, A.; Ready, J.S.; Haugaasen, T.; Sørnes, G.; de Boer, H.; Mauvisseau, Q. Harnessing eDNA Metabarcoding to Investigate Fish Community Composition and Its Seasonal Changes in the Oslo Fjord. Sci. Rep. 2024, 14, 10154. [Google Scholar] [CrossRef]
- Taberlet, P.; Luikart, G.; Waits, L.P. Noninvasive Genetic Sampling: Look before You Leap. Trends Ecol. Evol. 1999, 14, 323–327. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Turner, C.R.; Deiner, K.; Klymus, K.E.; Thomsen, P.F.; Murphy, M.A.; Spear, S.F.; McKee, A.; Oyler-McCance, S.J.; Cornman, R.S.; et al. Critical Considerations for the Application of Environmental DNA Methods to Detect Aquatic Species. Methods Ecol. Evol. 2016, 7, 1299–1307. [Google Scholar] [CrossRef]
- Hartman, L.J.; Coyne, S.R.; Norwood, D.A. Development of a Novel Internal Positive Control for Taqman® Based Assays. Mol. Cell Probes 2005, 19, 51–59. [Google Scholar] [CrossRef]
- Pierson, T.W.; McKee, A.M.; Spear, S.F.; Maerz, J.C.; Camp, C.D.; Glenn, T.C. Detection of an Enigmatic Plethodontid Salamander Using Environmental DNA. Copeia 2016, 104, 78–82. [Google Scholar] [CrossRef]
- Collins, R.A.; Bakker, J.; Wangensteen, O.S.; Soto, A.Z.; Corrigan, L.; Sims, D.W.; Genner, M.J.; Mariani, S. Non-Specific Amplification Compromises Environmental DNA Metabarcoding with COI. Methods Ecol. Evol. 2019, 10, 1985–2001. [Google Scholar] [CrossRef]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a Set of Universal PCR Primers for Metabarcoding Environmental DNA from Fishes: Detection of More than 230 Subtropical Marine Species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef]
- Li, Y.; Tang, M.; Lu, S.; Zhang, X.; Fang, C.; Tan, L.; Xiong, F.; Zeng, H.; He, S. A Comparative Evaluation of eDNA Metabarcoding Primers in Fish Community Monitoring in the East Lake. Water 2024, 16, 631. [Google Scholar] [CrossRef]
- Ludwig, E.J.; Lee, V.M.; Berkman, L.K.; Geheber, A.D.; Duvernell, D.D. Biodiversity Assessment of a Mississippi River Backwater Complex Using eDNA Metabarcoding. Diversity 2024, 16, 495. [Google Scholar] [CrossRef]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-Generation Monitoring of Aquatic Biodiversity Using Environmental DNA Metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef]
- Elbrecht, V.; Leese, F. PrimerMiner: An r Package for Development and in Silico Validation of DNA Metabarcoding Primers. Methods Ecol. Evol. 2017, 8, 622–626. [Google Scholar] [CrossRef]
- Etnier, D.E.; Starnes, W.C. The Fishes of Tennessee; University of Tennessee Press: Knoxville, TN, USA, 1993. [Google Scholar]
- Lennon, R.E. The Fishes of the Great Smoky Mountains National Park; Bureau of Sport Fisheries and Wildlife: Fairfax, VA, USA, 1960. [Google Scholar]
- Glenn, T.C.; Pierson, T.W.; Bayona-Vásquez, N.J.; Kieran, T.J.; Hoffberg, S.L.; Thomas, J.C.; Lefever, D.E.; Finger, J.W.; Gao, B.; Bian, X.; et al. Adapterama II: Universal Amplicon Sequencing on Illumina Platforms (TaggiMatrix). PeerJ 2019, 7, e7786. [Google Scholar] [CrossRef] [PubMed]
- Van Den Bulcke, L.; De Backer, A.; Ampe, B.; Maes, S.; Wittoeck, J.; Waegeman, W.; Hostens, K.; Derycke, S. Towards Harmonization of DNA Metabarcoding for Monitoring Marine Macrobenthos: The Effect of Technical Replicates and Pooled DNA Extractions on Species Detection. Metabar. Metagenom. 2021, 5, 233–247. [Google Scholar] [CrossRef]
- Nichols, R.V.; Vollmers, C.; Newsom, L.A.; Wang, Y.; Heintzman, P.D.; Leighton, M.; Green, R.E.; Shapiro, B. Minimizing Polymerase Biases in Metabarcoding. Mol. Ecol. Resour. 2018, 18, 927–939. [Google Scholar] [CrossRef]
- Campbell, C.D.; MacDonald, A.J.; Sarre, S.D. Evaluation of Genetic Markers for the Metabarcoding of Australian Marsupials from Predator Scats. Wildl. Res. 2024, 51, WR23134. [Google Scholar] [CrossRef]
- Fernandes, K.; van der Heyde, M.; Coghlan, M.; Wardell-Johnson, G.; Bunce, M.; Harris, R.; Nevill, P. Invertebrate DNA Metabarcoding Reveals Changes in Communities across Mine Site Restoration Chronosequences. Restor. Ecol. 2019, 27, 1177–1186. [Google Scholar] [CrossRef]
- Schnell, I.B.; Bohmann, K.; Gilbert, M.T.P. Tag Jumps Illuminated—Reducing Sequence-to-Sample Misidentifications in Metabarcoding Studies. Mol. Ecol. Resour. 2015, 15, 1289–1303. [Google Scholar] [CrossRef]
- Klymus, K.E.; Baker, J.D.; Abbott, C.L.; Brown, R.J.; Craine, J.M.; Gold, Z.; Hunter, M.E.; Johnson, M.D.; Jones, D.N.; Jungbluth, M.J.; et al. The MIEM Guidelines: Minimum Information for Reporting of Environmental Metabarcoding Data. Metabar. Metagenom. 2024, 8, e128689. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Mathon, L.; Valentini, A.; Guérin, P.E.; Normandeau, E.; Noel, C.; Lionnet, C.; Boulanger, E.; Thuiller, W.; Bernatchez, L.; Mouillot, D.; et al. Benchmarking Bioinformatic Tools for Fast and Accurate eDNA Metabarcoding Species Identification. Mol. Ecol. Resour. 2021, 21, 2565–2579. [Google Scholar] [CrossRef] [PubMed]
- McColl-Gausden, E.F.; Weeks, A.R.; Coleman, R.A.; Robinson, K.L.; Song, S.; Raadik, T.A.; Tingley, R. Multispecies Models Reveal That eDNA Metabarcoding Is More Sensitive than Backpack Electrofishing for Conducting Fish Surveys in Freshwater Streams. Mol. Ecol. 2021, 30, 3111–3126. [Google Scholar] [CrossRef]
- Di Muri, C.; Handley, L.L.; Bean, C.W.; Li, J.; Peirson, G.; Sellers, G.S.; Walsh, K.; Watson, H.V.; Winfield, I.J.; Hänfling, B. Read Counts from Environmental DNA (eDNA) Metabarcoding Reflect Fish Abundance and Biomass in Drained Ponds. Metabar. Metagenom. 2020, 4, e56959. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Ordination Methods, Diversity Analysis and Other Functions for Community and Vegetation Ecologists; R Package Version 2(0); Comprehensive R Archive Network; 2015; Volume 2. [Google Scholar]
- Berger, C.S.; Hernandez, C.; Laporte, M.; Côté, G.; Paradis, Y.; Kameni, T.; Kameni, T.D.W.; Normandeau, E.; Bernatchez, L. Fine-Scale Environmental Heterogeneity Shapes Fluvial Fish Communities as Revealed by eDNA Metabarcoding. Environ. DNA 2020, 2, 647–666. [Google Scholar] [CrossRef]
- Laporte, M.; Reny-Nolin, E.; Chouinard, V.; Hernandez, C.; Normandeau, E.; Bougas, B.; Côté, C.; Behmel, S.; Bernatchez, L. Proper Environmental DNA Metabarcoding Data Transformation Reveals Temporal Stability of Fish Communities in a Dendritic River System. Environ. DNA 2021, 3, 1007–1022. [Google Scholar] [CrossRef]
- Habera, J.W.; Kulp, M.A.; Moore, S.E.; Henry, T.B. Three-Pass Depletion Sampling Accuracy of Two Electric Fields for Estimating Trout Abundance in a Low-Conductivity Stream with Limited Habitat Complexity. N. Am. J. Fish. Manag. 2010, 30, 757–766. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Karr, J.R. Applying an Index of Biotic Integrity Based on Stream-Fish Communities: Considerations in Sampling and Interpretation. N. Am. J. Fish. Manag. 1986, 6, 418–429. [Google Scholar] [CrossRef]
- Karr, J.R.; Fausch, K.D.; Angermeier, P.L.; Yant, P.R.; Schlosser, I.J. Assessing Biological Integrity in Running Waters: A Method and Its Rationale. Ill. Nat. Hist. Surv. Spec. Publ. 1986, 5, 1–28. [Google Scholar]
- Nielsen, K.M.; Johnsen, P.J.; Bensasson, D.; Daffonchio, D. Release and Persistence of Extracellular DNA in the Environment. Environ. Biosaf. Res. 2007, 6, 37–53. [Google Scholar] [CrossRef]
- DeFlaun, M.; Paul, J.; Jeffrey, W. Distribution and Molecular Weight of Dissolved DNA in Subtropical Estuarine and Oceanic Environments. Mar. Ecol. Prog. Ser. 1987, 38, 65–73. [Google Scholar] [CrossRef]
- Saito, T.; Doi, H. A Model and Simulation of the Influence of Temperature and Amplicon Length on Environmental DNA Degradation Rates: A Meta-Analysis Approach. Front. Ecol. Evol. 2021, 9, 623831. [Google Scholar] [CrossRef]
- Barnes, M.A.; Turner, C.R. The Ecology of Environmental DNA and Implications for Conservation Genetics. Conserv. Genet. 2016, 17, 1–17. [Google Scholar] [CrossRef]
- Yamamoto, S.; Jo, T.; Murakami, H.; Minamoto, T.; Masuda, R. Effect of Water Temperature and Fish Biomass on Environmental DNA Shedding, Degradation, and Size Distribution. Ecol. Evol. 2019, 9, 1135–1146. [Google Scholar] [CrossRef]
- Pietramellara, G.; Ascher, J.; Borgogni, F.; Ceccherini, M.T.; Guerri, G.; Nannipieri, P. Extracellular DNA in Soil and Sediment: Fate and Ecological Relevance. Biol. Fertil. Soils 2009, 45, 219–235. [Google Scholar] [CrossRef]
- Zhu, B. Degradation of Plasmid and Plant DNA in Water Microcosms Monitored by Natural Transformation and Real-Time Polymerase Chain Reaction (PCR). Water Res. 2006, 40, 3231–3238. [Google Scholar] [CrossRef]
- Woodruff, S.P.; Johnson, T.R.; Waits, L.P. Evaluating the Interaction of Faecal Pellet Deposition Rates and DNA Degradation Rates to Optimize Sampling Design for DNA-Based Mark-Recapture Analysis of Sonoran Pronghorn. Mol. Ecol. Resour. 2015, 15, 843–854. [Google Scholar] [CrossRef]
- Bylemans, J.; Furlan, E.M.; Gleeson, D.M.; Hardy, C.M.; Duncan, R.P. Does Size Matter? An Experimental Evaluation of the Relative Abundance and Decay Rates of Aquatic Environmental DNA. Environ. Sci. Technol. 2018, 52, 6408–6416. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Yao, M. A Comprehensive and Comparative Evaluation of Primers for Metabarcoding eDNA from Fish. Methods Ecol. Evol. 2020, 11, 1609–1625. [Google Scholar] [CrossRef]
- Wei, N.; Nakajima, F.; Tobino, T. Effects of Treated Sample Weight and DNA Marker Length on Sediment eDNA Based Detection of a Benthic Invertebrate. Ecol. Indic. 2018, 93, 267–273. [Google Scholar] [CrossRef]
- Silsbee, D.G.; Larson, G.L. Water Quality of Streams in the Great Smoky Mountains National Park. Hydrobiologia 1982, 89, 97–115. [Google Scholar] [CrossRef]
- Bessey, C.; Jarman, S.N.; Berry, O.; Olsen, Y.S.; Bunce, M.; Simpson, T.; Power, M.; McLaughlin, J.; Edgar, G.J.; Keesing, J. Maximizing Fish Detection with eDNA Metabarcoding. Environ. DNA 2020, 2, 493–504. [Google Scholar] [CrossRef]
- Macher, T.H.; Schütz, R.; Arle, J.; Beermann, A.J.; Koschorreck, J.; Leese, F. Beyond Fish Edna Metabarcoding: Field Replicates Disproportionately Improve the Detection of Stream Associated Vertebrate Species. Metabar. Metagenom. 2021, 5, e66557. [Google Scholar] [CrossRef]
- Singer, G.A.C.; Fahner, N.A.; Barnes, J.G.; McCarthy, A.; Hajibabaei, M. Comprehensive Biodiversity Analysis via Ultra-Deep Patterned Flow Cell Technology: A Case Study of eDNA Metabarcoding Seawater. Sci. Rep. 2019, 9, 5991. [Google Scholar] [CrossRef]
- Brammell, B.F.; Strasko, E.K.; Brewer, S.A.; Piche, R.R.; Sams, C.M.; Mott, C.L.; Stull, M.A. Detecting Fossorial Salamanders Using eDNA: Development and Validation of Quantitative and End-Point PCR Assays for the Detection of Five Species of Ambystoma. Conserv. Genet. Resour. 2023, 15, 187–198. [Google Scholar] [CrossRef]
- Fujii, K.; Doi, H.; Matsuoka, S.; Nagano, M.; Sato, H.; Yamanaka, H. Environmental DNA Metabarcoding for Fish Community Analysis in Backwater Lakes: A Comparison of Capture Methods. PLoS ONE 2019, 14, e0210357. [Google Scholar] [CrossRef]
- Kumar, K.R.; Cowley, M.J.; Davis, R.L. Next-Generation Sequencing and Emerging Technologies. Semin. Thromb. Hemost. 2024, 50, 1026–1038. [Google Scholar] [CrossRef]
- Hänfling, B.; Handley, L.L.; Read, D.S.; Hahn, C.; Li, J.; Nichols, P.; Blackman, R.C.; Oliver, A.; Winfield, I.J. Environmental DNA Metabarcoding of Lake Fish Communities Reflects Long-Term Data from Established Survey Methods. Mol. Ecol. 2016, 25, 3101–3119. [Google Scholar] [CrossRef]
- Blabolil, P.; Harper, L.R.; Říčanová, Š.; Sellers, G.; Di Muri, C.; Jůza, T.; Vašek, M.; Sajdlová, Z.; Rychtecký, P.; Znachor, P.; et al. Environmental DNA Metabarcoding Uncovers Environmental Correlates of Fish Communities in Spatially Heterogeneous Freshwater Habitats. Ecol. Indic. 2021, 126, 107698. [Google Scholar] [CrossRef]
- Shu, L.; Chen, S.; Li, P.; Peng, Z. Environmental DNA Metabarcoding Reflects Fish DNA Dynamics in Lentic Ecosystems: A Case Study of Freshwater Ponds. Fishes 2022, 7, 257. [Google Scholar] [CrossRef]
- Deng, S.; Lin, B.; Luo, Y.; Dang, X.; Ma, C.; Zhou, Y.; Zhang, X.; Zhang, Y.; Xu, N.; Jiang, S.; et al. Assessment of Fish Species Biodiversity in the Yong River Basin Based on Environmental DNA Metabarcoding. Diversity 2025, 17, 35. [Google Scholar] [CrossRef]
- Flörl, L.; Cabrera, P.M.; Moccia, M.D.; Plüss, S.; Bokulich, N.A. HighALPS: Ultra-High-Throughput Marker-Gene Amplicon Library Preparation and Sequencing on the Illumina NextSeq and NovaSeq Platforms. Department of Health Sciences and Technology, Laboratory of Food Systems Biotechnology, Institute of Food, Nutrition, and Health, ETH Zurich, Switzerland. bioRxiv, 2024; Preprint uploaded to bioRxiv. [Google Scholar] [CrossRef]
- Meng, J.; Xu, F.; Yang, H.; Li, X.; Zhao, P. Exploring Microbiome and Plankton Responses and Interactions in the Mangrove Ecosystem through eDNA and Network Analysis. Sci. Total Environ. 2024, 930, 172581. [Google Scholar] [CrossRef]
- Fukaya, K.; Hasebe, Y. Occumb: An R Package for Site Occupancy Modeling of eDNA Metabarcoding Data. National Institute for Environmental Studies, Tsukuba, Ibaraki 305-8506, Japan. bioRxiv, 2025; Preprint uploaded to bioRxiv. [Google Scholar] [CrossRef]
- Deiner, K.; Fronhofer, E.A.; Mächler, E.; Walser, J.C.; Altermatt, F. Environmental DNA Reveals That Rivers Are Conveyer Belts of Biodiversity Information. Nat. Commun. 2016, 7, 12544. [Google Scholar] [CrossRef]
- Laporte, M.; Berger, C.S.; García-Machado, E.; Côté, G.; Morissette, O.; Bernatchez, L. Cage Transplant Experiment Shows Weak Transport Effect on Relative Abundance of Fish Community Composition as Revealed by eDNA Metabarcoding. Ecol. Indic. 2022, 137, 108785. [Google Scholar] [CrossRef]
- Fonseca, V.G.; Nichols, B.; Lallias, D.; Quince, C.; Carvalho, G.R.; Power, D.M.; Creer, S. Sample Richness and Genetic Diversity as Drivers of Chimera Formation in NSSU Metagenetic Analyses. Nucleic Acids Res. 2012, 40, e66. [Google Scholar] [CrossRef]
- Elbrecht, V.; Leese, F. Can DNA-Based Ecosystem Assessments Quantify Species Abundance? Testing Primer Bias and Biomass-Sequence Relationships with an Innovative Metabarcoding Protocol. PLoS ONE 2015, 10, e0130324. [Google Scholar] [CrossRef]
- Fonseca, V.G. Pitfalls in Relative Abundance Estimation Using Edna Metabarcoding. Mol. Ecol. Resour. 2018, 18, 923–926. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Kielgast, J.; Iversen, L.L.; Møller, P.R.; Rasmussen, M.; Willerslev, E. Detection of a Diverse Marine Fish Fauna Using Environmental DNA from Seawater Samples. PLoS ONE 2012, 7, e41732. [Google Scholar] [CrossRef]
- Sard, N.M.; Herbst, S.J.; Nathan, L.; Uhrig, G.; Kanefsky, J.; Robinson, J.D.; Scribner, K.T. Comparison of Fish Detections, Community Diversity, and Relative Abundance Using Environmental DNA Metabarcoding and Traditional Gears. Environ. DNA 2019, 1, 368–384. [Google Scholar] [CrossRef]
- Lawson Handley, L.; Read, D.S.; Winfield, I.J.; Kimbell, H.; Johnson, H.; Li, J.; Hahn, C.; Blackman, R.; Wilcox, R.; Donnelly, R.; et al. Temporal and Spatial Variation in Distribution of Fish Environmental DNA in England’s Largest Lake. Environ. DNA 2019, 1, 26–39. [Google Scholar] [CrossRef]
- Littlefair, J.E.; Hayhurst, L.D.; Yates, M.C.; Rennie, M.D.; Cristescu, M.E. A Strong Relationship between Environmental DNA Metabarcoding and Rank-Based Abundance of Fish. People and Nature Lab, Department of Genetics, Evolution, and Environment, University College London, United Kingdom. bioRxiv, 2025; Preprint uploaded to bioRxiv. [Google Scholar] [CrossRef]
- Nakagawa, H.; Fukushima, K.; Sakai, M.; Wu, L.; Minamoto, T. Relationships between the eDNA Concentration Obtained from Metabarcoding and Stream Fish Abundance Estimated by the Removal Method under Field Conditions. Environ. DNA 2022, 4, 1369–1380. [Google Scholar] [CrossRef]
- Lamb, P.D.; Hunter, E.; Pinnegar, J.K.; Creer, S.; Davies, R.G.; Taylor, M.I. How Quantitative Is Metabarcoding: A Meta-Analytical Approach. Mol. Ecol. 2019, 28, 420–430. [Google Scholar] [CrossRef]
- Lennon, R.E.; Parker, P.S. The Reclamation of Indian and Abrams Creeks, Great Smoky Mountains National Park; US Department of Interior, Fish and Wildlife Service: Washington, DC, USA, 1959. [Google Scholar]
- Tracy, B.H.; Rohde, F.C.; Hogue, G.M. An Annotated Atlas of the Freshwater Fishes of North Carolina. Southeast. Fishes Counc. Proc. 2020, 1, 1. [Google Scholar]
- Menhinick, E.F. The Freshwater Fishes of North Carolina; North Carolina Wildlife Resources Commission: Raleigh, NC, USA, 1991. [Google Scholar]
- Ciccotto, P. Observations of Co-Nest Building and Spawning in Native and Non-Native Chub Species (Nocomis spp. in the Swannanoa River, North Carolina. Southeast. Fish. Counc. Proc. 2024, 1, 5. [Google Scholar]
- Nico, L.; Fuller, P. Nocomis leptocephalus (Girard, 1856): U.S. Geological Survey, Nonindigenous Aquatic Species Database, Gainesville, FL. Available online: https://nas.er.usgs.gov/queries/factsheet.aspx?SpeciesID=575 (accessed on 2 January 2025).
- Lachner, E.A.; Jenkins, R.E. Systematics, Distribution, and Evolution of the Chub Genus Nocomis girard (Pisces, Cyprinidae) of Eastern United States, with Descriptions of New Species. Smithson. Contrib. Zool. 1971, 91, 1–27. [Google Scholar] [CrossRef]
- Etchison, L.; Owensby, D.; Rondel, C. Bringing Back the Natives: Reintroduction of Three Sucker Species in the Upper French Broad River. In Proceedings of the 34th Annual Meeting of the North Carolina Chapter of the American Fisheries Society, Durham, NC, USA, 21–23 February 2023. [Google Scholar]
- Bookwalter, J.; Niyas, A.M.M.; Caballero-López, B.; Villari, C.; Claramunt-López, B. Fecal Matters: Implementing Classical Coleoptera Species Lists with Metabarcoding Data from Passerine Bird Feces. J. Insect Conserv. 2023, 27, 557–569. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drainage | Watershed | # Samples |
|---|---|---|
| Abrams Creek | Abrams Creek | 5 |
| Big Creek | Big Creek | 1 |
| Calderwood Reservoir | Lake Cheoah-Little Tennessee River | 1 |
| Cataloochee River | Cataloochee Creek | 1 |
| Cataloochee River | Palmer Creek | 1 |
| Chilhowee Reservoir | Chilhowee Lake-Little Tennessee River | 1 |
| Cosby Creek | Cosby Creek | 1 |
| Deep Creek | Deep Creek | 1 |
| Dudley Creek | Dudley Creek | 1 |
| Eagle Creek | Eagle Creek | 1 |
| Fontana Reservoir | Fontana Lake-Tuckasegee River | 3 |
| Fontana Reservoir | Forney Creek | 1 |
| Fontana Reservoir | Hazel Creek | 1 |
| Fontana Reservoir | Lower Fontana Lake-Little Tennessee River | 3 |
| Fontana Reservoir | Noland Creek | 1 |
| Little Pigeon | Upper Little Pigeon River | 1 |
| Little Pigeon | Upper West Prong Little Pigeon River | 5 |
| Little Pigeon River | Middle Prong Little Pigeon River | 1 |
| Little River | Goshen Prong-Little River | 5 |
| Little River | West Prong Little River | 4 |
| Oconaluftee River | Lower Oconaluftee River | 1 |
| Oconaluftee River | Upper Oconaluftee River | 2 |
| Pigeon River | Cold Springs Creek-Pigeon River | 1 |
| Pigeon River | Cripple Creek-Pigeon River | 1 |
| Raven Fork | Lower Raven Fork | 1 |
| Roaring Fork | Roaring Fork | 1 |
| West Prong Little Pigeon | Le Conte Creek | 1 |
| West Prong Little Pigeon | Straight Fork-Raven Fork | 3 |
| Stream Order | Number Sampled |
|---|---|
| 6th | 1 |
| 5th | 9 |
| 4th | 14 |
| 3rd | 9 |
| 2nd | 9 |
| 1st | 8 |
| I.D. Level | Family | Genus/Species | Batra % | MiFish % |
|---|---|---|---|---|
| Species level identifications | Catostomidae | Catostomus commersonii | 0.34 | 0.40 |
| Hypentelium nigricans | 1.0 | 1.6 | ||
| Ictiobus bubalus | 0.008 | A | ||
| Moxostoma duquesnei | 0.04 | 0.34 | ||
| Moxostoma erythrurum | 0.003 | A | ||
| Centrarchidae | Ambloplites rupestris | 0.19 | 0.16 | |
| Lepomis cyanellus | 0.14 | 0.33 | ||
| Lepomis macrochirus | 0.35 | 0.38 | ||
| Micropterus dolomieu | 0.53 | 0.33 | ||
| Micropterus salmoides | A | 0.0006 | ||
| Clupeidae | Alosa chrysochloris | 0.005 | 0.0005 | |
| Cottidae | Cottus bairdii | 0.74 | 6.1 | |
| Cottus carolinae | 0.002 | 4.6 | ||
| Cyprinidae | Campostoma anomalum | 5.97 | 6.1 | |
| Clinostomus funduloides | A | 0.96 | ||
| Cyprinella galactura | 0.17 | 0.07 | ||
| Hybopsis amplops | 0.81 | 0.01 | ||
| Luxilus chrysocephalus | ∗ | A | ||
| Nocomis anomalum | 1.1 | 3.9 | ||
| Nocomis leptocephalus | 0.92 | A | ||
| Notropis leuciodus | 0.80 | 0.52 | ||
| Notropis rubellus | 0.67 | A | ||
| Notropis telescopus | ∗ | A | ||
| Pimephales notatus | 0.004 | A | ||
| Rhinichthys atratulus | 21.0 | 17.3 | ||
| Rhinichthys cataractae | 18.0 | 21.9 | ||
| Semotilus atromaculatus | 1.0 | 2.5 | ||
| Fundulidae | Fundulus catenatus | 0.012 | A | |
| Ictaluridae | Noturus baileyi | 0.013 | A | |
| Noturus flavipinnis | 0.005 | A | ||
| Percidae | Etheostoma blennioides | 0.37 | 0.40 | |
| Etheostoma flabellare | 0.23 | A | ||
| Etheostoma zonale | 0.001 | A | ||
| Etheostoma rufilineatum | A | 1.6 | ||
| Percina aurantiaca | 0.062 | 0.05 | ||
| Percina caprodes | A | 0.07 | ||
| Percina evides | 0.011 | 0.02 | ||
| Salmonidae | Oncorhynchus mykiss | 35.6 | 18.6 | |
| Salmo trutta | 2.1 | 1.4 | ||
| Salvelinus fontinalis | 7.6 | 10.3 | ||
| Genus level identifications | Catostomidae | Moxostoma | ∗ | |
| Centrarchidae | Micropterus | ∗ | ||
| Cottidae | Cottus | 0.003 |
| % Total Read Count | Stream Order | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | 5th | |||||
| Catostomidae | Catostomus commersonii | 0.36% | Low | ||||||
| Hypentelium nigricans | 1.25% | ||||||||
| Ictiobus bubalus | 0.01% | ||||||||
| Moxostoma duquesnei | 0.16% | ||||||||
| Moxostoma erythrurum | 0.002% | ||||||||
| Centrarchidae | Ambloplites rupestris | 0.18% | |||||||
| Lepomis cyanellus | 0.21% | ||||||||
| Lepomis macrochirus | 0.36% | ||||||||
| Micropterus dolomieu | 0.45% | High | |||||||
| Micropterus salmoides | 0.0002% | ||||||||
| Clupeidae | Alosa chrysochloris | 0.003% | |||||||
| Cottidae | Cottus bairdii | 2.82% | |||||||
| Cottus carolinae | 1.80% | ||||||||
| Cyprinidae | Campostoma anomalum | 6.02% | |||||||
| Clinostomus funduloides | 0.37% | ||||||||
| Cyprinella galactura | 0.13% | ||||||||
| Hybopsis amplops | 0.50% | ||||||||
| Luxilus chrysocephalus | 0.00% | ||||||||
| Nocomis leptocephalus | 0.57% | ||||||||
| Nocomis micropogon | 2.22% | ||||||||
| Notropis leuciodus | 0.69% | ||||||||
| Notropis rubellus | 0.41% | ||||||||
| Notropis telescopus | 0.0003% | ||||||||
| Pimephales notatus | 0.002% | ||||||||
| Rhinichthys atratulus | 19.60% | ||||||||
| Rhinichthys cataractae | 19.56% | ||||||||
| Semotilus atromaculatus | 1.59% | ||||||||
| Fundulidae | Fundulus catenatus | 0.01% | |||||||
| Ictaluridae | Noturus baileyi | 0.01% | |||||||
| Noturus flavipinnis | 0.003% | ||||||||
| Percidae | Etheostoma blennioides | 0.38% | |||||||
| Etheostoma rufilineatum | 0.61% | ||||||||
| Etheostoma flabellare | 0.14% | ||||||||
| Etheostoma zonale | 0.0004% | ||||||||
| Percina aurantiaca | 0.06% | ||||||||
| Percina caprodes | 0.03% | ||||||||
| Percina evides | 0.02% | ||||||||
| Salmonidae | Oncorhynchus mykiss | 29.02% | |||||||
| Salmo trutta | 1.83% | ||||||||
| Salvelinus fontinalis | 8.64% | ||||||||
| Stream Order | Elevation (m) | # Species | Shannon |
|---|---|---|---|
| 1st | 1448 | 3 | 1.172 |
| 1st | 1282 | 6 | 1.505 |
| 1st | 775 | 7 | 0.220 |
| 1st | 500 | 10 | 1.804 |
| 1st | 1462 | 4 | 0.015 |
| 1st | 530 | 4 | 1.078 |
| 1st | 527 | 5 | 1.618 |
| 1st | 526 | 3 | 1.032 |
| 2nd | 745 | 6 | 0.038 |
| 2nd | 338 | 10 | 1.346 |
| 2nd | 474 | 17 | 1.706 |
| 2nd | 1380 | 10 | 0.296 |
| 2nd | 593 | 4 | 0.839 |
| 2nd | 759 | 4 | 1.323 |
| 2nd | 1058 | 9 | 0.027 |
| 2nd | 1063 | 6 | 0.641 |
| 2nd | 499 | 2 | 0.833 |
| 3rd | 462 | 12 | 0.504 |
| 3rd | 430 | 10 | 1.251 |
| 3rd | 471 | 12 | 1.110 |
| 3rd | 526 | 6 | 1.047 |
| 3rd | 527 | 14 | 1.192 |
| 3rd | 534 | 8 | 0.981 |
| 3rd | 625 | 7 | 0.271 |
| 3rd | 1165 | 3 | 0.667 |
| 3rd | 668 | 7 | 0.810 |
| 4th | 521 | 13 | 1.449 |
| 4th | 488 | 7 | 0.697 |
| 4th | 442 | 16 | 1.863 |
| 4th | 506 | 9 | 1.702 |
| 4th | 407 | 10 | 1.201 |
| 4th | 857 | 9 | 0.867 |
| 4th | 832 | 10 | 1.285 |
| 4th | 691 | 4 | 0.522 |
| 4th | 514 | 9 | 1.351 |
| 4th | 515 | 10 | 0.628 |
| 4th | 801 | 10 | 1.284 |
| 4th | 523 | 9 | 1.141 |
| 4th | 564 | 9 | 1.216 |
| 4th | 555 | 13 | 1.829 |
| 5th | 410 | 19 | 2.387 |
| 5th | 564 | 17 | 2.022 |
| 5th | 417 | 19 | 1.867 |
| 5th | 338 | 26 | 2.463 |
| 5th | 577 | 9 | 1.389 |
| 5th | 616 | 14 | 1.795 |
| 5th | 751 | 10 | 1.417 |
| 5th | 529 | 15 | 1.872 |
| 5th | 683 | 10 | 1.515 |
| 6th | 336 | 24 | 2.639 |
| Site | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site Stream Order | 1st | 1st | 2nd | 2nd | 3rd | 3rd | 3rd | 3rd | ||||||||
| MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | |
| Campostoma anomalum | X | X | X | X | X | |||||||||||
| Catostomus commersonii | X | |||||||||||||||
| Cottus bairdii | X | |||||||||||||||
| Cottus carolinae | X | |||||||||||||||
| Etheostoma rufilineatum | X | X | X | |||||||||||||
| Hypentelium nigricans | X | |||||||||||||||
| Lepomis macrochirus | X | X | ||||||||||||||
| Micropterus dolomieu | X | |||||||||||||||
| Nocomis micropogon | X | |||||||||||||||
| Oncorhynchus mykiss | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||
| Rhinichthys atratulus | X | X | X | X | X | X | X | X | ||||||||
| Rhinichthys cataractae | X | X | X | X | X | X | X | X | X | X | ||||||
| Salmo trutta | X | X | X | |||||||||||||
| Salvelinus fontinalis | X | X | X | X | X | X | X | X | X | X | X | |||||
| Semotilus atromaculatus | X | X | X | X | X | X | ||||||||||
| % EF det. by eDNA | 100 | 100 | 100 | 100 | 100 | 100 | 67 | 67 | ||||||||
| Species det. only by eDNA | 5 | 3 | 4 | 0 | 6 | 1 | 3 | 5 | ||||||||
| Years of EF data | 1 | 33 | 17 | 18 | 1 | 25 | 6 | 2 | ||||||||
| Site Stream Order | 4th | 4th | 4th | 4th | 4th | 4th | 4th | 4th | 5th | 5th | 5th | 5th | 5th | 5th | 5th | 5th | 6th | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | |
| A. rupestris | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||
| C. anomalum | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||
| C. commersonii | X | X | X | X | X | X | X | X | X | X | X | |||||||||||||||||||||||
| C. funduloides | X | X | X | X | X | X | X | |||||||||||||||||||||||||||
| C. bairdii | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||||||||||||
| C. carolinae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||||||||||||||
| C. galactura | X | X | X | X | X | X | ||||||||||||||||||||||||||||
| E. monachus | X | |||||||||||||||||||||||||||||||||
| E. blennioides | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||
| E. chlorobranchium | X | X | X | X | X | X | ||||||||||||||||||||||||||||
| E. flabellare | X | X | X | X | X | X | X | X | X | |||||||||||||||||||||||||
| E. gutselli | X | |||||||||||||||||||||||||||||||||
| E. rufilineatum | X | X | X | X | X | X | X | |||||||||||||||||||||||||||
| E. simoterum | X | X | X | X | ||||||||||||||||||||||||||||||
| E. sitikuense | X | X | ||||||||||||||||||||||||||||||||
| E. swannanoa | X | |||||||||||||||||||||||||||||||||
| E. vulneratum | X | X | ||||||||||||||||||||||||||||||||
| E. zonale | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||
| F. catenatus | X | X | X | |||||||||||||||||||||||||||||||
| H. amblops | X | X | X | X | X | X | X | |||||||||||||||||||||||||||
| H. nigricans | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||||||||
| I. greeleyi | X | X | ||||||||||||||||||||||||||||||||
| I. bubalus | X | |||||||||||||||||||||||||||||||||
| L. cyanellus | X | X | X | X | X | |||||||||||||||||||||||||||||
| L. macrochirus | X | X | X | X | X | |||||||||||||||||||||||||||||
| L. appendix | X | |||||||||||||||||||||||||||||||||
| L. coccogenis | X | X | X | X | X | X | X | X | X | |||||||||||||||||||||||||
| M. dolomieu | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||
| M. punctulatus | X | |||||||||||||||||||||||||||||||||
| M. carinatum | X | |||||||||||||||||||||||||||||||||
| M. duquesnei | X | X | X | X | X | X | ||||||||||||||||||||||||||||
| M. erythrurum | X | |||||||||||||||||||||||||||||||||
| N. leptocephalus | X | X | X | X | X | X | X | |||||||||||||||||||||||||||
| N. micropogon | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||||||
| N. leuciodus | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||||||||||||||
| N. photogenis | X | X | ||||||||||||||||||||||||||||||||
| N. rubellus | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||
| N. rubricroceus | X | X | X | X | X | X | ||||||||||||||||||||||||||||
| N. spectrunculus | X | |||||||||||||||||||||||||||||||||
| N. telescopus | X | X | X | X | X | |||||||||||||||||||||||||||||
| N. baileyi | X | X | ||||||||||||||||||||||||||||||||
| N. flavipinnis | X | |||||||||||||||||||||||||||||||||
| O. mykiss | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |
| P. aurantiaca | X | X | ||||||||||||||||||||||||||||||||
| P. caprodes | X | |||||||||||||||||||||||||||||||||
| P. evides | X | X | X | X | X | |||||||||||||||||||||||||||||
| P. notatus | X | |||||||||||||||||||||||||||||||||
| R. atratulus | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||
| R. cataractae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||
| S. trutta | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||||||||
| S. fontinalis | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||||||||||||
| S. atromaculatus | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||||||||||||||
| % EF det. by eDNA | 78 | 100 | 71 | 100 | 100 | 100 | 67 | 44 | 73 | 86 | 88 | 100 | 77 | 100 | 88 | 75 | 90 | |||||||||||||||||
| Species det. only by eDNA | 6 | 7 | 4 | 0 | 5 | 7 | 3 | 3 | 4 | 10 | 3 | 6 | 9 | 10 | 3 | 6 | 6 | |||||||||||||||||
| Years of EF data | 22 | 4 | 1 | 15 | 5 | 1 | 11 | 2 | 2 | 10 | 18 | 18 | 1 | 2 | 1 | 1 | 5 | |||||||||||||||||
| Site Stream Order | 3rd | 3rd | 3rd | 4th | 4th | 5th | 5th | 5th | 5th | 5th | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | MB | EF | |
| Etheostoma zonale | X | X | ||||||||||||||||||
| Cottus carolinae | X | X | X | X | X | X | X | X | X | X | ||||||||||
| Hybopsis amblops | X | X | X | |||||||||||||||||
| Moxostoma duquesnei | X | X | ||||||||||||||||||
| Rhinichthys atratulus | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||
| Lepomis macrochirus | X | X | X | X | ||||||||||||||||
| Nocomis leptocephalus | X | |||||||||||||||||||
| Salvelinus fontinalis | X | X | X | X | X | X | X | X | ||||||||||||
| Salmo trutta | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||
| Campostoma anomalum | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||
| Alosa chrysochloris | X | |||||||||||||||||||
| Etheostoma sitikuense | X | |||||||||||||||||||
| Semotilus atromaculatus | X | X | X | X | X | X | X | X | ||||||||||||
| Etheostoma flabellare | X | X | X | |||||||||||||||||
| Percina evides | X | X | X | |||||||||||||||||
| Moxostoma erythrurum | X | X | ||||||||||||||||||
| Lepomis cyanellus | X | X | ||||||||||||||||||
| Etheostoma chlorobranchium | X | X | X | X | X | X | X | |||||||||||||
| Etheostoma blennioides | X | X | X | X | X | X | X | X | X | X | ||||||||||
| Micropterus salmoides | X | |||||||||||||||||||
| Percina caprodes | X | |||||||||||||||||||
| Rhinichthys cataractae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||
| Notropis spectrunculus | X | X | ||||||||||||||||||
| Cottus bairdii | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||
| Ichthyomyzon greeleyi | X | X | ||||||||||||||||||
| Hypentelium nigricans | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||
| Percina squamata | X | X | X | |||||||||||||||||
| Oncorhynchus mykiss | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |
| Etheostoma rufilineatum | X | X | X | X | X | X | ||||||||||||||
| Nocomis micropogon | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||
| Moxostoma carinatum | X | |||||||||||||||||||
| Ambloplites rupestris | X | X | X | X | X | X | ||||||||||||||
| Notropis rubellus | X | X | X | X | X | X | ||||||||||||||
| Clinostomus funduloides | X | X | X | X | X | |||||||||||||||
| Notropis rubricroceus | X | X | ||||||||||||||||||
| Micropterus dolomieu | X | X | X | X | ||||||||||||||||
| Noturus baileyi | X | X | ||||||||||||||||||
| Etheostoma simoterum | X | |||||||||||||||||||
| Etheostoma swannanoa | X | |||||||||||||||||||
| Percina aurantiaca | X | X | ||||||||||||||||||
| Notropis telescopus | X | X | X | X | X | |||||||||||||||
| Etheostoma tennesseense | X | X | X | |||||||||||||||||
| Notropis leuciodus | X | X | X | X | X | X | X | X | ||||||||||||
| Etheostoma gutselli | X | X | X | X | ||||||||||||||||
| Luxilus coccogenis | X | X | X | X | ||||||||||||||||
| Catostomus commersonii | X | X | X | |||||||||||||||||
| Cyprinella galactura | X | X | ||||||||||||||||||
| Perca flavescens | X | X | X | |||||||||||||||||
| Noturus flavipinnis | X | X | ||||||||||||||||||
| % EF det. by eDNA | 100 | 57 | 100 | 57 | 53 | 92 | 94 | 100 | 88 | 63 | ||||||||||
| Species det. only by eDNA | 8 | 2 | 3 | 5 | 3 | 7 | 11 | 4 | 5 | 4 | ||||||||||
| Years of EF data | 1 | 7 | 3 | 3 | 4 | 4 | 4 | 1 | 3 | 5 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brammell, B.F.; Brewer, S.A.; Fetter, K.S.; Slone, L.E.; Kulp, M.A.; McLaughlin, B.R.S. Ichthyofaunal Metabarcoding in the Southern Appalachians: Use of eDNA Metabarcoding in Fish Surveys in Lotic Systems of the Great Smoky Mountains National Park with Comparisons to Historic Electrofishing Data. Fishes 2025, 10, 145. https://doi.org/10.3390/fishes10040145
Brammell BF, Brewer SA, Fetter KS, Slone LE, Kulp MA, McLaughlin BRS. Ichthyofaunal Metabarcoding in the Southern Appalachians: Use of eDNA Metabarcoding in Fish Surveys in Lotic Systems of the Great Smoky Mountains National Park with Comparisons to Historic Electrofishing Data. Fishes. 2025; 10(4):145. https://doi.org/10.3390/fishes10040145
Chicago/Turabian StyleBrammell, Ben F., Sara A. Brewer, Karsner S. Fetter, Lauren E. Slone, Matt A. Kulp, and Ben R. S. McLaughlin. 2025. "Ichthyofaunal Metabarcoding in the Southern Appalachians: Use of eDNA Metabarcoding in Fish Surveys in Lotic Systems of the Great Smoky Mountains National Park with Comparisons to Historic Electrofishing Data" Fishes 10, no. 4: 145. https://doi.org/10.3390/fishes10040145
APA StyleBrammell, B. F., Brewer, S. A., Fetter, K. S., Slone, L. E., Kulp, M. A., & McLaughlin, B. R. S. (2025). Ichthyofaunal Metabarcoding in the Southern Appalachians: Use of eDNA Metabarcoding in Fish Surveys in Lotic Systems of the Great Smoky Mountains National Park with Comparisons to Historic Electrofishing Data. Fishes, 10(4), 145. https://doi.org/10.3390/fishes10040145