
In Situ Study on the Influence of a Dark Environment on the Upstream Behaviors of Plateau Fishes in Fishways: A Pilot Study

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
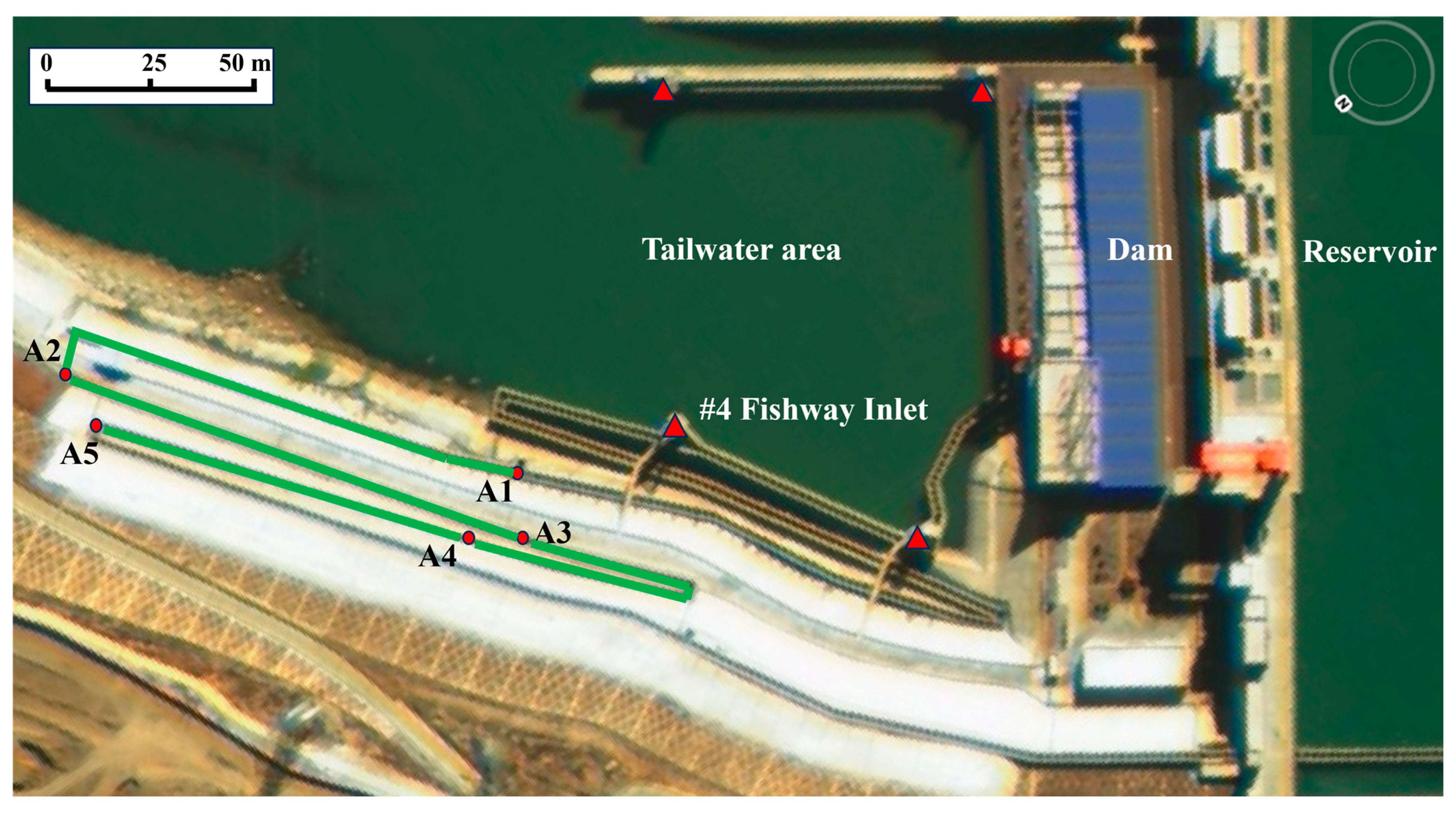
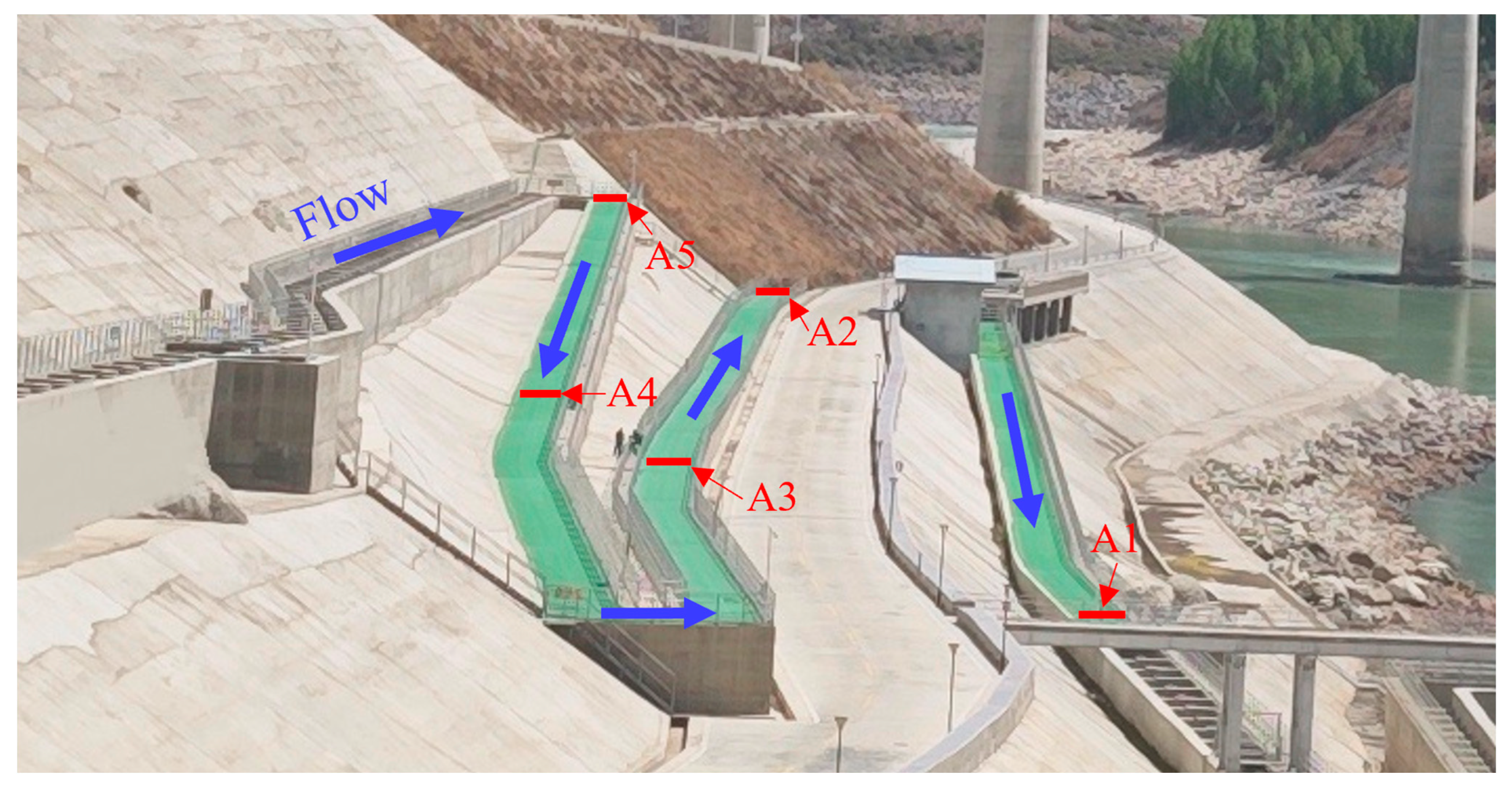
2.1. Study Area
2.2. Experimental Design
2.3. Data Analysis
3. Results
3.1. Entry Efficiency at the Experimental Segment and Passage Efficiency at Each Antenna
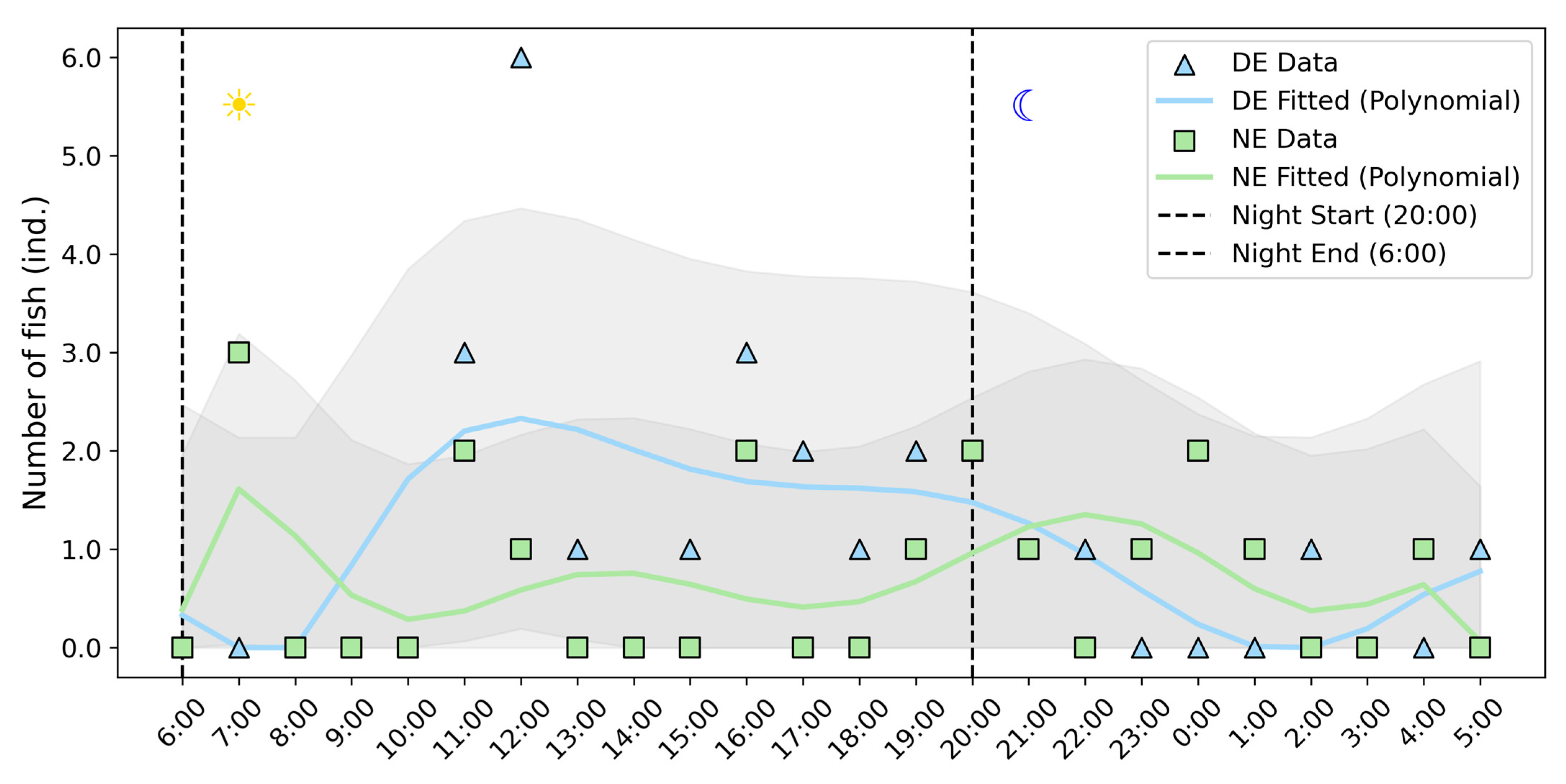
3.2. Upstream Rhythms of Fish in Different Environments
3.3. Fish Transit Speed and Influencing Factors
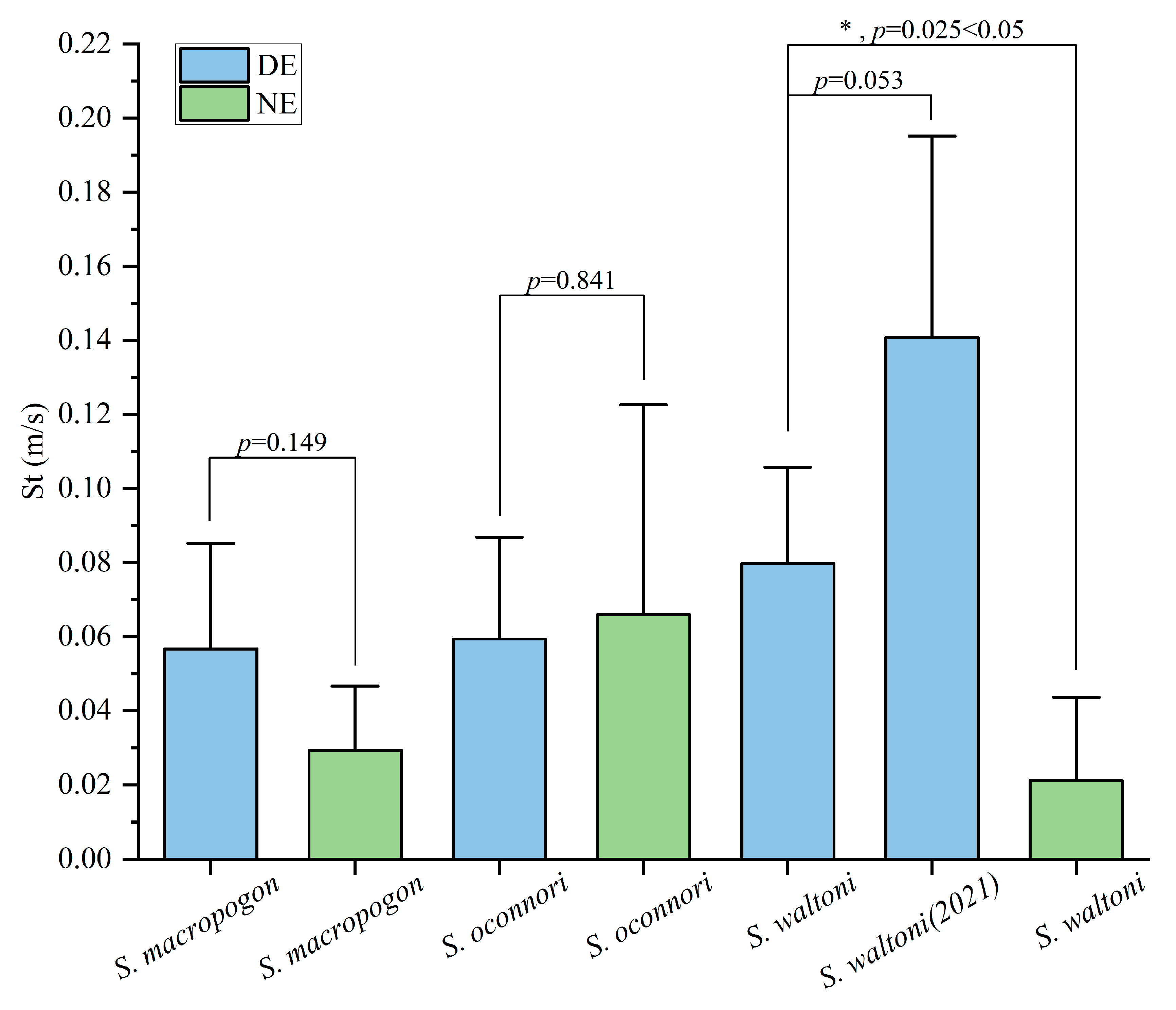
3.3.1. Analysis of Environmental Differences in Transit Speed
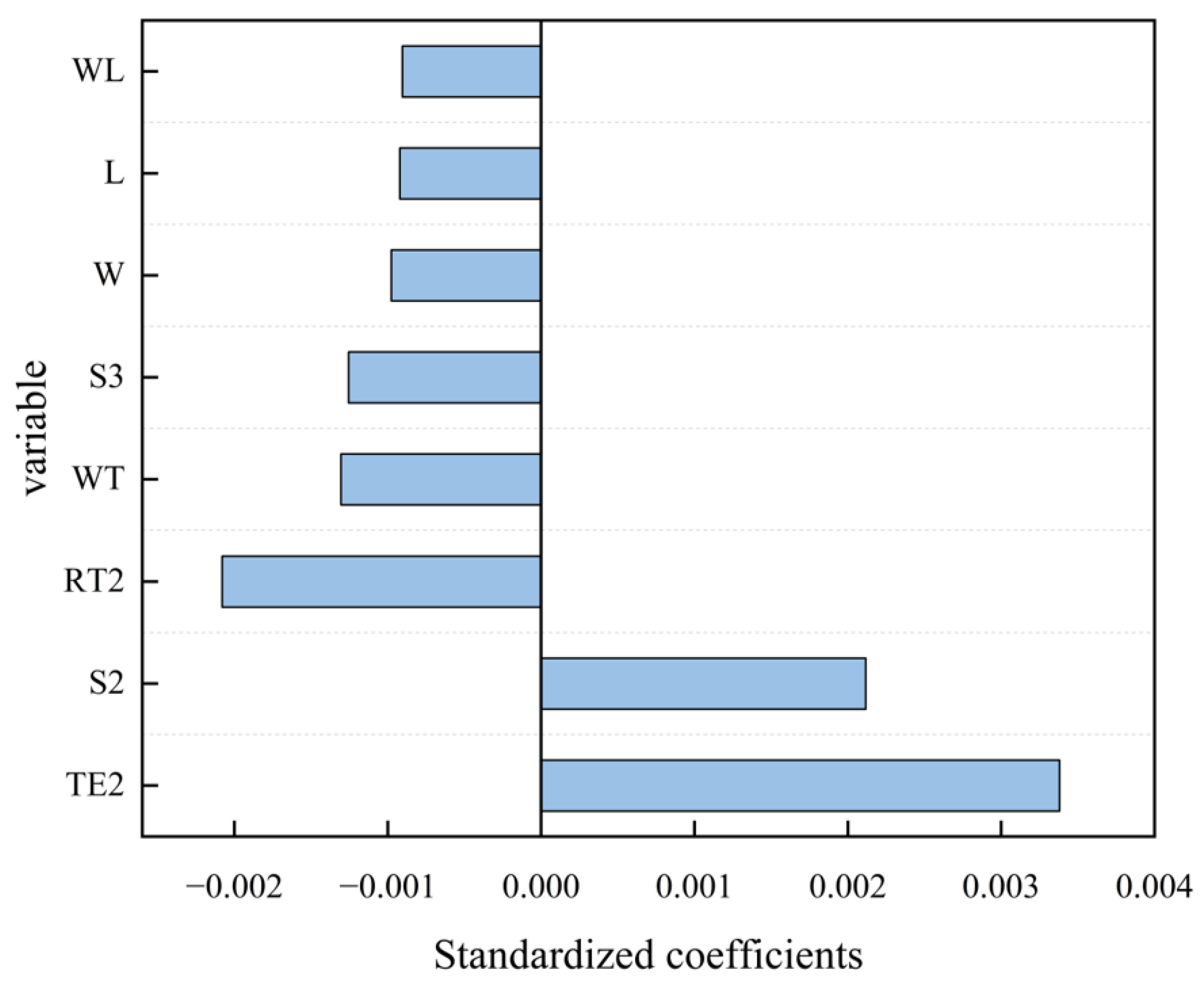
3.3.2. Analysis of the Key Factors Influencing Transit Speeds
4. Discussion
4.1. Analysis of Fish Upstream Behaviors in Different Environments
4.2. Identification of the Key Factors Influencing Transit Speeds
4.3. Establishment of a Basin-Wide PIT Database
5. Conclusions
- There were no significant differences in the entry efficiencies at the experimental segment for the three species in the dark and natural light environments. However, when facing the dark environment, the three fishes had entry efficiencies at the experimental segment of 66.7%, 88.9%, and 66.7%, respectively, all higher than their performances in the natural light environment;
- The passage efficiencies of S. macropogon and S. waltoni showed no significant differences between the dark and natural light environments, with the passage efficiencies at the A5 antenna ranging from a minimum of 16.7% (S. waltoni in the dark environment) to a maximum of 40.0% (S. macropogon in the natural light environment). In contrast, S. oconnori exhibited a significant difference in the passage efficiencies between the two environments, with a passage efficiency at the A5 antenna of 0 in the natural light environment, which increased significantly to 75.0% in the dark environment;
- The dark environment disrupted the crepuscular migration rhythms of the three plateau schizothoracids. In the natural light environment, these fishes exhibit a distinct diel rhythm, with pronounced migration peaks during the morning (07:00–07:59) and evening (19:00–21:59) twilight periods. However, in the dark environment, the majority (76%) of the individuals migrate upstream during the daytime (06:00–19:59);
- The average transit speeds of S. macropogon and S. waltoni in the dark environment were 2 times and 3.8 times higher, respectively, than those in the natural light environment, while the transit speeds of S. oconnori remained relatively stable across both environments. The dark environment had a dominant and positive effect on the transit speeds of all three species. The differences in the upstream behaviors among the species under varying light conditions highlighted species-specific adaptive responses. S. oconnori outperformed S. macropogon and S. waltoni in upstream migration, while S. waltoni exhibited greater adaptability to the dark environment.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
- The following abbreviations are used in this manuscript:
| DE | dark environment |
| NE | natural light environment |
| Ee | entry efficiency at the experimental segment |
| Ep | passage efficiency |
| St | transit speed |
References
- Freeman, M.C.; Pringle, C.M.; Jackson, C.R. Hydrologic connectivity and the contribution of stream headwaters to ecological integrity at regional scales. J. Am. Water Resour. Assoc. 2007, 43, 5–14. [Google Scholar] [CrossRef]
- Amoros, C.; Bornette, G. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshw. Biol. 2002, 47, 761–776. [Google Scholar] [CrossRef]
- Blanchet, S.; Rey, O.; Etienne, R.; Lek, S.; Loot, G. Species-specific responses to landscape fragmentation: Implications for management strategies. Evol. Appl. 2010, 3, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Gouskov, A.; Reyes, M.; Wirthner-Bitterlin, L.; Vorburger, C. Fish population genetic structure shaped by hydroelectric power plants in the upper Rhine catchment. Evol. Appl. 2016, 9, 394–408. [Google Scholar] [CrossRef]
- Hall, C.J.; Jordaan, A.; Frisk, M.G. The historic influence of dams on diadromous fish habitat with a focus on river herring and hydrologic longitudinal connectivity. Landsc. Ecol. 2011, 26, 95–107. [Google Scholar] [CrossRef]
- Rosenberg, D.M.; McCully, P.; Pringle, C.M. Global-scale environmental effects of hydrological alterations: Introduction. Bioscience 2000, 50, 746–751. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef]
- Pan, D.; Lichuan, K.; Ying, C.; Peng, D.; Xin, G.; Hao, X. Research on the Structure and Application of canal-tunnel combined vertical slot fishway. Sichuan Water Resour. 2024, 45, 60–63. [Google Scholar]
- Xiaoli, W. CHN ENERGY started construction of the longest ecological fishway project in Asia. Constr. Enterp. Manag. 2023, 4, 120. [Google Scholar]
- Xianren, Z.; Chao, Y. Research on the design and application of fishway engineering of the BaLin Dam. Jilin Water Resour. 2024, 6, 69–73. [Google Scholar] [CrossRef]
- Qingyun, T.; Jie, T. The Application of Numerical Simulation in the Smooth Design of the Fishway of the Balin Dam. Sci. Technol. Innov. 2024, 5–8. [Google Scholar]
- Jones, M.J.; Hale, R. Using knowledge of behaviour and optic physiology to improve fish passage through culverts. Fish Fish. 2020, 21, 557–569. [Google Scholar] [CrossRef]
- Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. Patterns of avoidance behaviours in the light/dark preference test in young juvenile zebrafish: A pharmacological study. Behav. Brain Res. 2011, 222, 15–25. [Google Scholar] [CrossRef]
- Thorpe, J.E.; Morgan, R.I.G.; Pretswell, D.; Higgins, P.J. Movement Rhythms in Juvenile Atlantic Salmon, Salmo-Salar L. J. Fish Biol. 1988, 33, 931–940. [Google Scholar] [CrossRef]
- Huang, J.; Lin, C.; Shi, X.; Zhang, N.; Shi, S.; Zhang, B.; Mo, W. Light environment optimization of culvert fishway based on the phototaxis of Schizothorax prenanti. Chin. J. Ecol. 2021, 40, 2155–2163. [Google Scholar] [CrossRef]
- Jones, M.J.; Baumgartner, L.J.; Zampatti, B.P.; Beyer, K. Low light inhibits native fish movement through a vertical-slot fishway: Implications for engineering design. Fish. Manag. Ecol. 2017, 24, 177–185. [Google Scholar] [CrossRef]
- Hadderingh, R.H.; Van Aerssen, G.; De Beijer, R.; Van der Velde, G. Reaction of silver eels to artificial light sources and water currents: An experimental deflection study. Regul. Rivers-Res. Manag. 1999, 15, 365–371. [Google Scholar] [CrossRef]
- Tarena, F.; Comoglio, C.; Candiotto, A.; Nyqvist, D. Artificial light at night affects fish passage rates in two small-sized Cypriniformes fish. Ecol. Freshw. Fish 2024, 33, 13. [Google Scholar] [CrossRef]
- Lin, C.Y.; Deng, Z.D.; Shi, X.T.; Dai, H.C.; Wang, J.X.; Huang, W.Q.; Luo, J.; Huang, J. Mutually promoting or constraining? Disentangling the superimposed effect of velocity and illuminance on fish motion in low-velocity flows with a novel metric. Freshw. Biol. 2022, 67, 1468–1480. [Google Scholar] [CrossRef]
- Xu, J.; Sang, W.; Dai, H.; Lin, C.; Ke, S.; Mao, J.; Wang, G.; Shi, X. A Detailed Analysis of the Effect of Different Environmental Factors on Fish Phototactic Behavior: Directional Fish Guiding and Expelling Technique. Animals 2022, 12, 240. [Google Scholar] [CrossRef]
- Wang, H.T.; Jiang, X.T.; Liu, K.J.; Pu, X.C.; Wang, Y.M. Swimming ability of Schizothoracinae fishes in Yarlung Zangbo River of China. J. Fish Biol. 2024, 105, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Franklin, A.E.; Haro, A.; Castro-Santos, T.; Noreika, J. Evaluation of Nature-Like and Technical Fishways for the Passage of Alewives at Two Coastal Streams in New England. Trans. Am. Fish. Soc. 2012, 141, 624–637. [Google Scholar] [CrossRef]
- Robinson, W.; Baumgartner, L.J.; Homsombath, K.; Ning, N.; Phommachanh, K.; Phommavong, T.; Poomchaivej, T.; Pomorin, K.; Simmanivong, D.; Singhanouvong, D.; et al. PIT tagging systems are suitable for assessing cumulative impacts of Mekong River hydropower plants on (upstream) fish migrations in Lao PDR. Fish Res. 2024, 274, 5. [Google Scholar] [CrossRef]
- Xu, J.W.; Li, D.Q.; Hu, X.Z.; Jiao, Y.L.; Wang, J.P.; Wu, Y.J.; Lin, C.Y.; Ke, S.F.; Bai, T.X.; Wang, N.N.; et al. Quantitative Assessment and Regulation of Passage and Entrance Attraction Efficiency of Vertical-Slot Fishway on Heishuihe River in Southwest China. Animals 2024, 14, 2365. [Google Scholar] [CrossRef]
- Bao, J.H.; Li, W.W.; Zhang, C.S.; Mi, X.Y.; Li, H.T.; Zhao, X.J.; Cao, N.; Twardek, W.M.; Cooke, S.J.; Duan, M. Quantitative assessment of fish passage efficiency at a vertical-slot fishway on the Daduhe River in Southwest China. Ecol. Eng. 2019, 141, 12. [Google Scholar] [CrossRef]
- Keep, J.K.; Watson, J.R.; Cramp, R.L.; Jones, M.J.; Gordos, M.A.; Ward, P.J.; Franklin, C.E. Low light intensities increase avoidance behaviour of diurnal fish species: Implications for use of road culverts by fish. J. Fish Biol. 2021, 98, 634–642. [Google Scholar] [CrossRef]
- Liang, Y.; Hou, Y.Q.; Hu, W.B.; Johnson, D.; Wang, J.X. Flow velocity preference of Schizothorax oconnori Lloyd swimming upstream. Glob. Ecol. Conserv. 2021, 32, 10. [Google Scholar] [CrossRef]
- Hou, Y.Q.; Yang, Z.; An, R.D.; Cai, L.; Chen, X.J.; Zhao, X.J.; Zou, X. Water flow and substrate preferences of Schizothorax wangchiachii (Fang, 1936). Ecol. Eng. 2019, 138, 1–7. [Google Scholar] [CrossRef]
- Keefer, M.L.; Caudill, C.C.; Peery, C.A.; Moser, M.L. Context-dependent diel behavior of upstream-migrating anadromous fishes. Environ. Biol. Fishes 2013, 96, 691–700. [Google Scholar] [CrossRef]
- Ono, K.; SimenstadSchool, C.A. Reducing the effect of overwater structures on migrating juvenile salmon: An experiment with light. Ecol. Eng. 2014, 71, 180–189. [Google Scholar] [CrossRef]
- Pulgar, J.; Manríquez, P.H.; Widdicombe, S.; Garcia-Huidobro, R.; Quijón, P.A.; Carter, M.; Aldana, M.; Quintanilla-Ahumada, D.; Duarte, C. Artificial Light at Night (ALAN) causes size-dependent effects on intertidal fish decision-making. Mar. Pollut. Bull. 2023, 193, 6. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Xie, D.; Li, B.; Liu, Y.; Hu, S.; Liu, H.; Shi, Y.; Zhu, S. Influence of water temperature, habitat complexity and light on the predatory performance of the dark sleeper Odontobutis potamophila (Gunther, 1861). J. Freshw. Ecol. 2020, 35, 367–378. [Google Scholar] [CrossRef]
- Koeda, K.; Touma, H.; Tachihara, K. Nighttime migrations and behavioral patterns of Pempheris schwenkii. PeerJ 2021, 9, 15. [Google Scholar] [CrossRef]
- Peterson, A.N.; Soto, A.P.; McHenry, M.J. Pursuit and Evasion Strategies in the Predator–Prey Interactions of Fishes. Integr. Comp. Biol. 2021, 61, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Thurow, R.F.; Peterson, J.T.; Chandler, G.L.; Moffitt, C.M.; Bjornn, T.C. Concealment of juvenile bull trout in response to temperature, light, and substrate: Implications for detection. PLoS ONE 2020, 15, 17. [Google Scholar] [CrossRef]
- Maximino, C.; de Brito, T.M.; Dias, C.; Gouveia, A.; Morato, S. Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 2010, 5, 209–216. [Google Scholar] [CrossRef]
- Ali, M.A.; Sakurai, S.; Collin, S.P. Adaptative Radiation of the Retina in Galaxiidae (Salmoniformes). Aust. J. Zool. 1990, 38, 173–186. [Google Scholar] [CrossRef]
- Amtstaetter, F.; O’Connor, J.; Borg, D.; Stuart, I.; Moloney, P. Remediation of upstream passage for migrating Galaxias (Family: Galaxiidae) through a pipe culvert. Fisheries Manag. Ecol. 2017, 24, 186–192. [Google Scholar] [CrossRef]
- Bogaard, M.; Gido, K.; McKinstry, M.; Pennock, C. Water temperature predicts razorback sucker Xyrauchen texanus spawning migrations. Environ. Biol. Fishes 2023, 106, 1503–1517. [Google Scholar] [CrossRef]
- Katopodis, C.; Kells, J.; Acharya, M. Nature-Like and Conventional Fishways: Alternative Concepts? Can. Water Resour. J. 2001, 26, 211–232. [Google Scholar] [CrossRef]
- Hockley, F.; Wilson, C.; Brew, A.; Cable, J. Fish responses to flow velocity and turbulence in relation to size, sex and parasite load. J. R. Soc. Interface 2013, 11, 20130814. [Google Scholar] [CrossRef] [PubMed]
- Matley, J.K.; Klinard, N.V.; Jaine, F.R.A.; Lennox, R.J.; Koopman, N.; Reubens, J.T.; Harcourt, R.G.; Cooke, S.J.; Huveneers, C. Long-term effects of tagging fishes with electronic tracking devices. Fish. Fish. 2024, 25, 1009–1025. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | River | Dam Height/Water Level (m) | Total Length (m) | Tunnel Length (m) | Tunnel Proportion (%) | Status | Type |
|---|---|---|---|---|---|---|---|
| Gongzui Hydropower Station Fishway | Dadu River | 85.6 | 1565 | 674 | 43.1 | Completed | Vertical-Slot Fishway |
| Batang Hydropower Station Fishway | Jinsha River | 69 | 2176 | 570 | 26.2 | Under Construction | Vertical-Slot Fishway |
| Jinchuan Hydropower Station Fishway | Dadu River | 111.5 | 5020 | 962 | 19.2 | Under Construction | Natural-like Fishway |
| Zhala Hydropower Station Fishway | Yuqiao River | 67 | 2970 | 821 | 27.6 | Under Construction | Vertical-Slot Fishway |
| Xiangqi Hydropower Station Fishway | Xiangjiang River | 75 | 556 | 475 | 85.4 | Completed | Vertical-Slot Fishway |
| Balin Dam Fishway | Mengjiang Bawang River | 46 | 1830 | 265 | 14.5 | Planned | Natural-like Fishway |
| The Fishway Used in This Experiment | Yarlung Zangbo River | 42 | 2597 | 655 | 25.2 | - | Vertical-Slot Fishway |
| Test Environment | NE | DE | NE | DE | NE | DE | DE | |
|---|---|---|---|---|---|---|---|---|
| Species | S. macropogon | S. oconnori | S. waltoni | S. waltoni (2021) | ||||
| N0 | 10 | 9 | 12 | 18 | 18 | 9 | - | |
| Fish Length (cm) | Mean ± SD (range) | 42.5 ± 4.13 (35.6–49.8) | 45.7 ± 3.01 (40.5–48.6) | 45.6 ± 9.02 (32.2–61.5) | 44.4 ± 3.51 (37.6–50.6) | 43.3 ± 5.96 (32.1–51.2) | 42.7 ± 10.96 (25.1–54.4) | - |
| Fish weight (g) | Mean ± SD (range) | 796.9 ± 290.96 (389.3–1335.3) | 881.6 ± 135.31 (653.0–1088.2) | 1101.5 ± 756.38 (357.6–2733.0) | 770.9 ± 147.63 (450.8–975.7) | 886.6 ± 323.37 (404.3–1432.5) | 748.5 ± 492.71 (95.2–1376.3) | - |
| N1 | 5 | 6 | 9 | 16 | 8 | 6 | 8 | |
| N2 | 5 | 4 | 8 | 15 | 6 | 3 | 8 | |
| N3 | 3 | 4 | 5 | 15 | 5 | 3 | 7 | |
| N4 | 2 | 3 | 2 | 12 | 3 | 1 | 3 | |
| N5 | 2 | 2 | 0 | 12 | 2 | 1 | 3 | |
| Ee | 50.0% | 66.7% | 75.0% | 88.9% | 44.4% | 66.7% | - | |
| Ep2 | 100.0% | 66.7% | 88.9% | 93.8% | 75.0% | 50.0% | 100.0% | |
| Ep3 | 60.0% | 66.7% | 55.6% | 93.8% | 62.5% | 50.0% | 87.5% | |
| Ep4 | 40.0% | 50.0% | 22.2% | 75.0% | 37.5% | 16.7% | 37.5% | |
| Ep5 | 40.0% | 33.3% | 0% | 75.0% | 25.0% | 16.7% | 37.5% | |
| St (m/s) | Mean ± SD (range) | 0.029 ± 0.017 (0.012–0.050) | 0.057 ± 0.027 (0.026–0.091) | 0.066 ± 0.057 (0.004–0.122) | 0.059 ± 0.027 (0.014–0.118) | 0.021 ± 0.022 (0.004–0.061) | 0.080 ± 0.026 (0.063–0.110) | 0.141 ± 0.054 (0.068–0.216) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Yao, F.; Lv, J.; Li, H.; Wang, Z.; Huang, Y.; Chen, K.; He, W.; Wang, X.; Li, J. In Situ Study on the Influence of a Dark Environment on the Upstream Behaviors of Plateau Fishes in Fishways: A Pilot Study. Fishes 2025, 10, 136. https://doi.org/10.3390/fishes10030136
Wang B, Yao F, Lv J, Li H, Wang Z, Huang Y, Chen K, He W, Wang X, Li J. In Situ Study on the Influence of a Dark Environment on the Upstream Behaviors of Plateau Fishes in Fishways: A Pilot Study. Fishes. 2025; 10(3):136. https://doi.org/10.3390/fishes10030136
Chicago/Turabian StyleWang, Biao, Fei Yao, Jianzhang Lv, Hongze Li, Zhe Wang, Yongzeng Huang, Kaixiao Chen, Wei He, Xiaogang Wang, and Jingjuan Li. 2025. "In Situ Study on the Influence of a Dark Environment on the Upstream Behaviors of Plateau Fishes in Fishways: A Pilot Study" Fishes 10, no. 3: 136. https://doi.org/10.3390/fishes10030136
APA StyleWang, B., Yao, F., Lv, J., Li, H., Wang, Z., Huang, Y., Chen, K., He, W., Wang, X., & Li, J. (2025). In Situ Study on the Influence of a Dark Environment on the Upstream Behaviors of Plateau Fishes in Fishways: A Pilot Study. Fishes, 10(3), 136. https://doi.org/10.3390/fishes10030136