EvoDevo: An Ongoing Revolution?


{kind=link}
Abstract
1. Introduction: The House That Charles Built
2. One, No One, and Hundred Thousand EvoDevo
2.1. One EvoDevo
2.2. No One EvoDevo
2.3. One Hundred Thousand EvoDevo
2.4. The Search for Unity
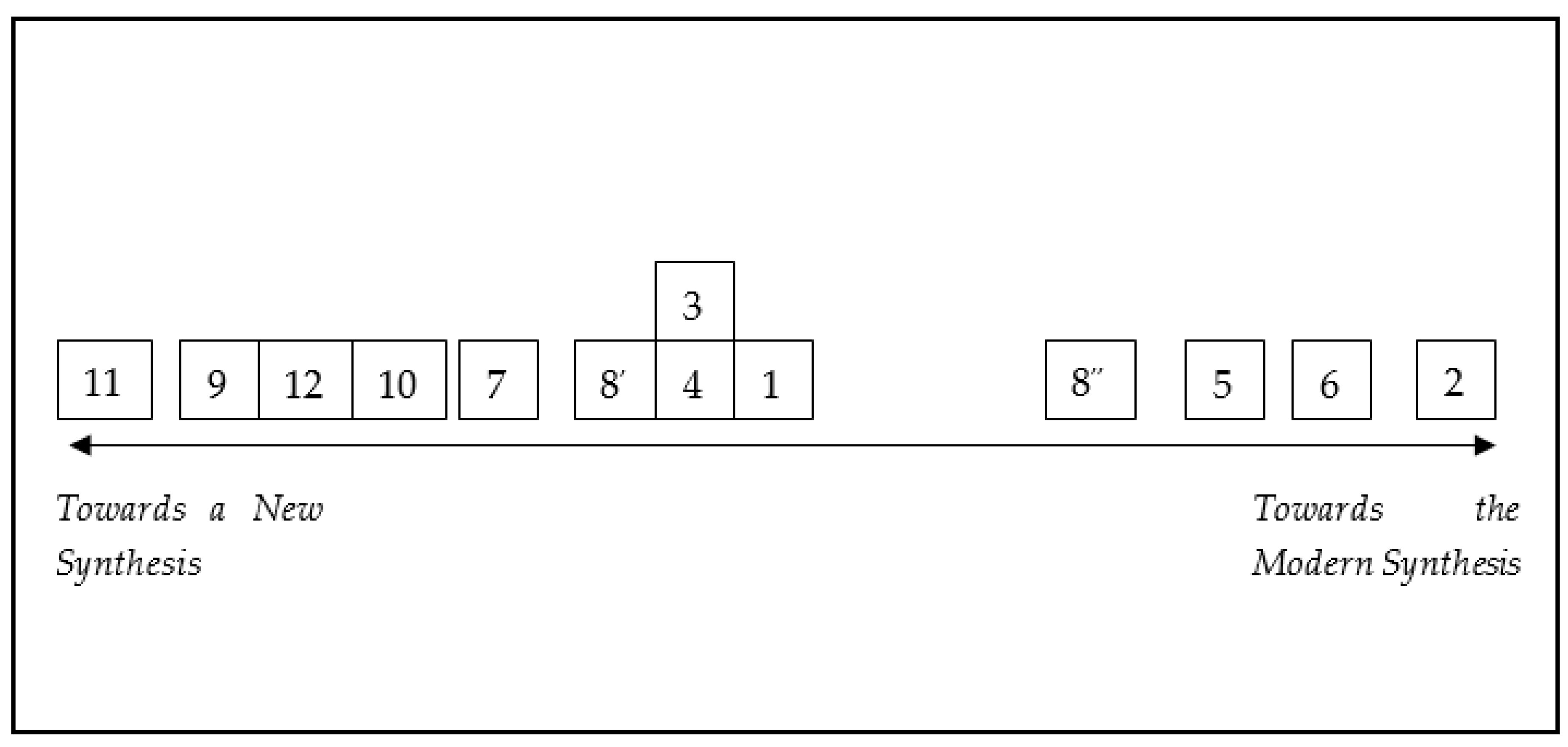
3. A Synthesis within a Synthesis
On the Synthesis and Expansion of Theories
4. Conclusions: Towards a Theory of What?
Funding
Acknowledgments
Conflicts of Interest
References
- Toulmin, S. Human Understanding, Vol. I, General Introduction and Part I; Clarendon Press: Oxford, UK, 1972; ISBN 9780691071855. [Google Scholar]
- Darwin, C. The Origin of Species by Means of Natural Selection, or the Preservation of Favored Race in the Struggle of Life; Cambridge University Press: New York, NY, USA, 2009; ISBN 9781108005487. [Google Scholar]
- Robert, S.J. Embryology, Epigenesis, and Evolution: Taking Development Seriously; Cambridge University Press: Cambridge, UK, 2004; ISBN 9780521030861. [Google Scholar]
- Kirschner, M.W.; Gerhart, J.C. Facilitated Variation. In Evolution the Extended Synthesis; Müller, G.B., Pigliucci, M., Eds.; The MIT Press: Cambridge, MA, USA, 2010; pp. 253–280. ISBN 9780262315142. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1951; ISBN 1120819306. [Google Scholar]
- Dobzhansky, T.; Ayala, J.F.; Stebbins, G.L.; Valentine, J.W. Evoluciòn; Edicion Omega: Barcelona, Spain, 1983. [Google Scholar]
- Withfield, J. Postmodern Evolution? Nature 2008, 455, 281–284. [Google Scholar] [CrossRef][Green Version]
- Hall, B.K. Unlocking the Black Box between Genotype and Phenotype: Cell condensation as Morphogenetic (modular) Units. Biol. Philos. 2003, 18, 219–247. [Google Scholar] [CrossRef]
- Waddington, C.H. The Evolution of an Evolutionist; Edinburgh University Press: Edinburgh, UK, 1975. [Google Scholar]
- Hall, B.K. Evo-devo or devo-evo—Does it matter? Evol. Dev. 2000, 2, 177–178. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F. Evo-Devo, Devo-Evo, and Devgen-Popgen. Biol. Philos. 2003, 18, 347–352. [Google Scholar] [CrossRef]
- Müller, G.B. Evo-Devo: Extending the evolutionary synthesis. Nat. Rev. Genet. 2007, 8, 943–949. [Google Scholar] [CrossRef]
- Müller, G.B.; Wagner, G.P. Novelty in Evolution: Restructuring the concept. Annu. Rev. Ecol. Syst. 1991, 22, 229–256. [Google Scholar] [CrossRef]
- Müller, G.B.; Newman, S.A. The innovation triad: An EvoDevo agenda. J. Exp. Zool. 2005, 304, 487–503. [Google Scholar] [CrossRef]
- Müller, G.B. Homology: The evolution of morphological organization. In Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology; Müller, G.B., Newman, S.A., Eds.; MIT Press: Cambridge, MA, USA, 2003; pp. 51–69. [Google Scholar]
- Minelli, A.; Fusco, G. Homology. In The Philosophy of Biology: A Companion for Educators; Kampourakis, K., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 289–322. ISBN 9789400765375. [Google Scholar]
- Carroll, S.B. Endless Forms Most Beautiful: The New Science of EvoDEvo and the Making of the Animal Kingdom; Norton: New York, NY, USA, 2005; ISBN 9780393327793. [Google Scholar]
- Wagner, G. Homology, Genes, and Evolutionary Innovation; Princeton University Press: Princeton, NJ, USA, 2014; ISBN 9780691180670. [Google Scholar]
- Kirschner, M.; Gerhart, J. Evolvability. Proc. Natl. Acad. Sci. USA 1998, 95, 8420–8427. [Google Scholar] [CrossRef]
- Minelli, A. Evolvability and Its Evolvability. In Challenging the Modern Synthesis: Adapatation, Development, and Inheritance; Huneman, P., Walsh, D., Eds.; Oxford University Press: Oxford, UK, 2017; pp. 211–238. ISBN 978-0199377176. [Google Scholar]
- Arthur, W. The interaction between developmental bias and natural selection: From centipede segments to a general hypothesis. Heredity 2002, 89, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, S. Modularity, comparative embryology and evo-devo: Developmental dissection of evolving body plans. Dev. Biol. 2009, 332, 61–69. [Google Scholar] [CrossRef]
- Robert, J.S. Evo-Devo. In The Oxford Handbook of Philosophy of Biology; Ruse, M., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 291–309. ISBN 9780199737260. [Google Scholar]
- Laubichler, M.D. Evolutionary Developmental Biology. In The Cambridge Companion to the Philosophy of Biology; Hull, D., Ruse, M., Eds.; Cambridge University Press: Cambirdge, UK, 2008; pp. 342–360. ISBN 9781139001588. [Google Scholar]
- Love, A.C. Evolutionary Developmental Biology: Philosophical Issues. In Handbook of Evolutionary Thinking in the Sciences; Heams, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 265–283. [Google Scholar]
- Müller, G.B. Evolutionary Developmental Biology. In Handbook of Evolution, Vol. 2: The Evolution of Living Systems (Including Hominids); Wuketis, F.M., Ayala, F.J., Eds.; Wiley-VCH: Hoboken, NJ, USA, 2005; pp. 87–115. ISBN 9783527308385. [Google Scholar]
- Minelli, A. Evolutionary Developmental Biology does not Offer a Significant Challenge to the Neo-Darwinian Paradigm. In Contemporary Debates in Philosophy of Biology; Ayala, F.J., Arp, R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 213–226. ISBN 9781405159982. [Google Scholar]
- Laubichler, M.D. Evolutionary Developmental Biology Offers a Significant Challenge to the Neo-Darwinian Paradigm. In Contemporary Debates in Philosophy of Biology; Ayala, F.J., Arp, R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 198–212. ISBN 9781405159982. [Google Scholar]
- Carroll, R.L. Towards a new evolutionary synthesis. Tree 2000, 15, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Raff, R.A. Evo-Devo: The evolution of a new discipline. Nat. Rev. Gen. 2000, 1, 74–79. [Google Scholar] [CrossRef]
- Minelli, A. Grand challenges in evolutionary developmental biology. Front. Ecol. Evol. 2015, 2, 1–11. [Google Scholar] [CrossRef]
- Müller, G.B. Evo-devo as a discipline. In Evolving Pathways: Key themes in Evolutionary Developmental Biology; Minelli, A., Fusco, G., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 5–30. ISBN 9781849722698. [Google Scholar]
- Hall, B.K.; Olson, W.M. Keywords and Concepts in Evolutionary Developmental Biology; Harvard University Press: Cambridge, MA, USA, 2003; ISBN 9781849722698. [Google Scholar]
- Müller, G.B. Six Memos for Evo-Devo. In From Embryology to EvoDevo: A History of Developmental Evolution; Laubichler, M.D., Maienschein, J., Eds.; The MIT Press: Cambridge, UK, 2007; pp. 499–524. [Google Scholar]
- Minelli, A. Fusco, Evolving Pathways: Key themes in Evolutionary Developmental Biology; Cambridge University Press: Cambridge, UK, 2008; ISBN 9781107405455. [Google Scholar]
- Burian, R.M. On conflicts between genetic and developmental viewpoints—And their attempted resolution in molecular biology. In Structures and Norms in Science; Dalla Chiara, M.L., Doets, K., Mundici, D., van Benthem, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 243–264. ISBN 978-0-7923-4384-4. [Google Scholar]
- Laubichler, M.D.; Wagner, G.P. 2004—Introduction to the papers of the 2001 Kowalevsky Medal Winner Symposium. J. Exp. Zool. Mol. Dev. Evol. 2004, 302, 1–4. [Google Scholar] [CrossRef]
- McGinnis, W.; Levine, M.S.; Hafen, E.; Kuroiwa, A.; Gehring, W.J. A Conserved DNA sequence in homeotic genes of the Drosophila Antennapedia and bithorax complexes. Nature 1984, 308, 428–433. [Google Scholar] [CrossRef]
- Wakimoto, B.T.; Kaufman, T.C. Analysis of Larval Segmentation in Lethal Genotypes Associated with the Antennapedia Gene Complex in Drosophila Melanogaster. Dev. Biol. 1981, 81, 51–64. [Google Scholar] [CrossRef]
- Carroll, S.B. Endless Forms: The Evolution of Gene Regulation and Morphological Diversity. Cell 2000, 101, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Slack, J. A Rosetta Stone for pattern formation in animals? Nature 1984, 310, 364–365. [Google Scholar] [CrossRef]
- Gould, S.J. The Structure of Evolutionary Theory; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 2002; ISBN 9780674006133. [Google Scholar]
- Davidson, E.H. The Regulatory Genome: Gene Regulatory Networks in Development and Evolution; Academic Press: New York, NY, USA, 2006; ISBN 9781280641329. [Google Scholar]
- Carroll, S.B. Evo-Devo and an Expanding Evolutionary Synthesis: A Genetic Theory of Morphological Evolution. Cell 2008, 134, 25–36. [Google Scholar] [CrossRef]
- Carroll, S.B. 2001 The Big Picture. Nature 2001, 409, 669. [Google Scholar] [CrossRef] [PubMed]
- Sterelny, K. Development, Evolution, and Adaptation. Philos. Sci. 2000, 67, 369–387. [Google Scholar] [CrossRef]
- Futuyma, D.J. Evolutionary biology today and the call for an extended synthesis. Interface Focus 2017, 7, 20160145. [Google Scholar] [CrossRef]
- Hoekstra, H.E.; Coyne, J.A. The Locus of Evolution: EvoDevo and the genetics of adaptations. Evolution 2007, 61, 995–1016. [Google Scholar] [CrossRef] [PubMed]
- Nikla, K.J. Thinking outside the Hox. Biol. Theory 2006, 1, 128–129. [Google Scholar] [CrossRef]
- Medina, M.L. Two “EvoDevos”. Biol. Theory 2010, 5, 7–11. [Google Scholar] [CrossRef]
- Amundson, R. The Changing Role of the Embryo in Evolutionary Thought; Cambridge University Press: Cambridge, UK, 2005; ISBN 9780521703970. [Google Scholar]
- Gehring, W.J.; Hiromi, Y. Homeotic Genes and the Homeobox. Annu. Rev. Genet. 1986, 20, 147–173. [Google Scholar] [CrossRef]
- Gehring, W.J. Master Control Genes in Development and Evolution: The Homeobox Story; Yale University Press: New Haven, CT, USA, 1998; ISBN 9780300074093. [Google Scholar]
- Forgacs, G.; Newman, S.A. Biological Physics of the Developing Embryo; Cambridge University Press: Cambridge, UK, 2005; ISBN 9780521783378. [Google Scholar]
- Kauffman, S. The Origins of Order: Self-Organizating and Selection in Evolution; Oxford University Press: Oxford, UK, 1993; ISBN 9780195058116. [Google Scholar]
- Newman, S.A.; Muller, G.B. Epigenetic mechanisms of character origination. J. Exp. Zool. B Mol. Dev. Evol. 2000, 288, 304–317. [Google Scholar] [CrossRef]
- Callebaut, W.; Muller, G.B.; Newman, S.A. The Organismic Systems Approach: Evo-Devo and the Streamlining of the Naturalistic Agenda. In Integrating Evolution and Development: From Theory to Practice; Samson, R., Brandon, R., Eds.; The MIT Press: Cambridge, MA, USA, 2007; ISBN 9780262195607. [Google Scholar]
- Müller, G.B.; Newman, S. Origination of Organismal Form: The Forgotten Cause in Evolutionary Theory. In Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology; Müller, G.B., Newman, S., Eds.; MIT Press: Cambridge, MA, USA, 2003; pp. 3–10. ISBN 9780262280327. [Google Scholar]
- Müller, G.B.; Newman, S. Generation, Integration, Autonomy: Three Steps in the Evolution of Homology. Homology (Novartis Foundation Symposium 222); Wiley: Chichester, UK, 1999; pp. 65–79. ISBN 9780470515662. [Google Scholar]
- Goodwin, B. Evolution and the generative order. In Theoretical Biology: Epigenetic and Evolutionary Order from Complex Systems; Goodwin, B., Saunders, P.T., Eds.; Edinburgh University Press: Edinburgh, UK, 1989; pp. 89–100. ISBN 9780801845192. [Google Scholar]
- Webster, G.; Goodwin, B. Form and Transformation: Generative and Relational Principles in Biology; Cambridge University Press: Cambridge, UK, 1996; ISBN 9780521207430. [Google Scholar]
- Ho, M.W.; Saunders, P.T. Beyond Neo-darwinism: And epigenetic approach to evolution. J. Theor. Biol. 1979, 78, 573–591. [Google Scholar] [CrossRef]
- Callebaut, W. Self-organization and optimization: Conflicting or complementary approaches? In Evolutionary Systems; Van de Vijver, G., Salthe, S.N., Delpos, M., Eds.; Kluwer: Dordrecht, The Netherlands, 1998; pp. 79–100. [Google Scholar]
- Weber, B.H.; Depew, P.J. Natural selection and self-organization. Biol. Philos. 1996, 11, 33–65. [Google Scholar] [CrossRef]
- Fontana, W. Modelling EvoDevo with RNA. BioEssays 2002, 24, 1164–1177. [Google Scholar] [CrossRef]
- Fontana, W. The topology of the possible. In Understanding Change: Models, Methodologies and Metaphors; Wimmer, A., Kössler, R., Eds.; Palgrave Macmillan: London, UK, 2006; pp. 67–84. ISBN 9780230524644. [Google Scholar]
- Lewontin, R.C. The Docrtine of DNA: Biology as Ideology; Penguin: New York, NY, USA, 1993; ISBN 9780140232196. [Google Scholar]
- Rose, S.; Lewontin, R.C.; Konin, L. Not in Our Genes: Biology, Ideology and Human Nature; Penguin Books: Harmondsworth, UK, 1990; ISBN 9780394508177. [Google Scholar]
- Lewontin, R.C. The Triple Helix: Gene, Organism, and Environment; Harvard University Press: Cambridge, MA, USA, 2000; ISBN 9780674006775. [Google Scholar]
- Levins, R.; Lewontin, R.C. A program for biology. Biol. Theory 2006, 1, 333–335. [Google Scholar] [CrossRef]
- Kohl, P.; Crampin, E.J.; Quinn, T.A.; Noble, D. Systems Biology: An Approach. Clin. Pharmacol. Ther. 2010, 88, 25–33. [Google Scholar] [CrossRef]
- Kitano, H. Systems Biology: Toward System-Level Understanding of Biological Systems. In Foundations of Systems Biology; Kitano, H., Ed.; The MIT Press: Cambridge, MA, USA, 2001; pp. 1–36. ISBN 9780262277204. [Google Scholar]
- Kirschner, M.W. The Meaning of Systems Biology. Cell 2005, 121, 503–504. [Google Scholar] [CrossRef]
- Krohs, U.; Callebaut, W. Data without models merging with models without data. In Systems Biology: Philosophical Foundations; Boogerd, F.C., Bruggeman, F.J., Hofmeyer, S.J.-H., Westerhoff, H.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 181–213. ISBN 9780444520852. [Google Scholar]
- Noble, D. The Aims of Systems Biology: Between Molecules and Organisms. Pharmacopsychiatry 2011, 44 (Suppl. 1), 9–14. [Google Scholar] [CrossRef]
- Kitano, H. Systems Biology: A Brief Overview. Science 2002, 295. [Google Scholar] [CrossRef]
- Newman, S.A. The Fall and Rise of Systems Biology: Recovering from a Halfcentury Gene Hing; GeneWatch: London, UK, 2003; Volume 16, pp. 8–12. [Google Scholar]
- Noble, D. A theory of biological relativity: No privileged level of causation. Interface Focus 2012, 2, 55–64. [Google Scholar] [CrossRef]
- Noble, D. The Music of Life: Biology beyond the Genome; Oxford University Press: Oxford, UK, 2006; ISBN 9780199295739. [Google Scholar]
- Piaget, J. Behavior and Evolution; Pantheon Books: New York, NY, USA, 1978; ISBN 9780394735887. [Google Scholar]
- Weiss, P. Cellular Dynamics. Rev. Mod. Phys. 1959, 31, 11–20. [Google Scholar] [CrossRef]
- Griesemer, J.R. Reproduction and the reduction of genetics. In The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives; Beurton, P.J., Falk, R., Rheinberger, H.J., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 240–285. ISBN 9780521060240. [Google Scholar]
- Griesemer, J. The units of evolutionary transitions. Selection 2000, 1, 67–80. [Google Scholar] [CrossRef]
- Gottlieb, G. Probabilistic Epigenesis. Dev. Sci. 2007, 10, 1–11. [Google Scholar] [CrossRef]
- Griesemer, J.R. Reproduction and scaffolded developmental processes. In Towards a Theory of Development; Minelli, A., Pradeu, T., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 182–202. ISBN 9780199671434. [Google Scholar]
- Wimsatt, W.C.; Griesemer, J.R. Reproducing entrenchments to scaffold culture. In Integrating Evolution and Development: From Theory to Practice; Sanson, R., Brandon, R.N., Eds.; The MIT Press: Cambridge, MA, USA, 2007; pp. 227–323. ISBN 9780262195607. [Google Scholar]
- Oyama, S.; Griffiths, P.E.; Gray, R.D. Introduction: What is developmental Systems Theory? In Cycles of Contingency: Developmental Systems and Evolution; Oyama, S., Griffiths, P.E., Gray, R.D., Eds.; The MIT Press: Cambridge, MA, USA, 2001; ISBN 97802626506321-12. [Google Scholar]
- Oyama, S. The Ontogeny of Information: Developmental Systems and Evolution; Duke University Press: Durham, NC, USA, 2000; ISBN 9780822324317. [Google Scholar]
- Robert, J.S.; Hall, B.K.; Olson, W.M. Bridging the gap between developmental systems theory and evolutionary developmental biology. BioEssays 2001, 23, 954–962. [Google Scholar] [CrossRef]
- Caporael, L.R. Evolution, Groups, and Scaffolded Mings. In Developing Scaffolds in Evolution, Culture, and Cognition; Caporael, L.R., Griesemer, J.R., Wimsatt, W.C., Eds.; The MIT Press: Cambridge, MA, USA, 2014; pp. 57–76. ISBN 9781461952367. [Google Scholar]
- Caporael, L.R. The Evolution of Truly Social Cognition: The Core Configurations Model. Pers. Soc. Psychol. Rev. 1997, 1, 276–298. [Google Scholar] [CrossRef]
- West-Eberhard, M.-J. Developmental Plasticity and Evolution; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- West-Eberhard, M.-J. Developmental Plasticity and the Origin of Species Differences. Proc. Natl. Acad. Sci. USA 2005, 102 (Suppl. 1), 6543–6549. [Google Scholar] [CrossRef]
- Kupiec, K.J. The Origin of Individuals; Worlds Scientific Publishing: Singapore, 2009; ISBN 9789812704993. [Google Scholar]
- Kupiec, K.J. L’Ontophylogenese: Evolution des Especes et Developpement de L’individu; Editions Quæ: Versailles, France, 2012; ISBN 9782759217861. [Google Scholar]
- Gottlieb, G. Individual Development and Evolution: The Genesis of Novel Behavior; Oxford University Press: New York, NY, USA, 1992; ISBN 9780415648486. [Google Scholar]
- Abouheif, E.; Favé, M.-J.; Ibarraran-Viniegra, A.S.; Lesoway, M.P.; Rafiqi, A.M.; Rajakumar, R. Eco-Evo-Devo: The times has come. In Ecological Genomics: Ecology and the Evolution of Genes and Genomes; Laundry, C.R., Aubin-North, N., Eds.; Advances in Experimental Medicine and Biology 781; Springer: Dordrecht, The Netherlands, 2014; pp. 107–125. [Google Scholar]
- Gilbert, S.F.; Epel, D. Ecological Developmental Biology: The Environmental Regulation of Development, Health, and Evolution; Sinauer Associates: Sunderland, MA, USA, 2015; ISBN 9781605353449. [Google Scholar]
- Bateson, P.P.G. Behaviour, Development and Evolution; Cambridge University Press: Cambridge, UK, 2017; ISBN 9781783742486. [Google Scholar]
- Stotz, K. Extended evolutionary psychology: The importance of transgenerational developmental plasticity. Front. Psychol. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Wilkins, A.S. The Evolution of Developmental Pathways; Sinauer Associates: Sunderland, MA, USA, 2002; ISBN 9780878939169. [Google Scholar]
- Griesemer, J. Development, Culture, and the Units of Inheritance. Philos. Sci. 2000, 348–368. [Google Scholar] [CrossRef]
- Giere, R.N. Scientific Perspectivism; The University of Chicago Press: Chicago, IL, USA, 2006; ISBN 9780226292137. [Google Scholar]
- Robert, J.S. How developmental is evolutionary developmental biology? Biol. Philos. 2002, 17, 591–611. [Google Scholar] [CrossRef]
- Maynard-Smith, J. Shaping Life: Genes, Embryos and Evolution; Yale University Press: New Haven, CT, USA, 1998; ISBN 9780300080223. [Google Scholar]
- Nuno de la Rosa, L. Computing the Extended Synthesis: Mapping the Dynamics and Conceptual Structure of the Evolvability Research Front. J. Exp. Zool. Mol. Dev. Evol. 2017, 328, 395–396. [Google Scholar] [CrossRef]
- Winther, R.G. Evo-devo as a Trading zone. In Conceptual Change in Biology, Boston Studies in the Philosophy and History of Science; Love, A.C., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 459–482. ISBN 9789401794121. [Google Scholar]
- Pigliucci, M. Do we need an extended evolutionary synthesis? Evolution 2007, 61, 2743–2749. [Google Scholar] [CrossRef]
- Pigliucci, M.; Muller, G.B. Evolution: The Extended Synthesis; The MIT Press: Cambridge, MA, USA, 2010; ISBN 9780262315142. [Google Scholar]
- Hall, B.K. Evolutionary Developmental Biology (Evo-Devo): Past, Present, and Future. Evol. Educ. Outreach 2012, 5, 184–193. [Google Scholar] [CrossRef]
- Laland, K.N.; Uller, T.; Feldman, M.W.; Sterelny, K.; Müller, G.B.; Moczek, A.P.; Jablonka, E.; Odling-Smee, J.; Wray, G.A.; Hoekstra, H.E.; et al. Does evolutionary theory need a rethink? Nat. Cell Biol. 2014, 514, 161–164. [Google Scholar] [CrossRef]
- Wray, G.A.; Hoekstra, H.E.; Futuyma, D.J.; Lensky, R.E.; Mackay, T.F.C.; Schluter, D.; Strassmann, J.E. Does evolutionary theory need a rethink? No, all is well. Nature 2014, 514, 161–164. [Google Scholar] [CrossRef]
- Laland, K.N.; Uller, T.; Feldman, M.W.; Sterelny, K.; Müller, G.B.; Moczek, A.P.; Jablonka, E.; Odling-Smee, F.J. The extended evolutionary synthesis: Its structure, assumptions and predictions. Proc. R. Soc. B Boil. Sci. 2015, 282, 20151019. [Google Scholar] [CrossRef]
- Müller, G.B. EvoDevo Shapes the Extended Synthesis. Biol. Theory 2014, 9, 119–121. [Google Scholar] [CrossRef][Green Version]
- Futuyma, D.J. Can Modern Evolutionary Theory Explain Macroevolution? In Macroevolution; Serrelli, E., Gontier, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 29–85. ISBN 978-3-319-15044-4. [Google Scholar]
- Fàbregas-Tejeda, A.; Vergara-Silva, F. The emerging structure of the Extended Evolutionary Synthesis: Where does Evo-Devo fit in? Theory Biosci. 2018, 137, 169–184. [Google Scholar] [CrossRef]
- Sarkar, S. Evolutionary Theory in the 1920s: The Nature of the “Synthesis”. Philos. Sci. 2004, 71, 1215–1226. [Google Scholar] [CrossRef][Green Version]
- Love, A.C. Evolutionary Morphology, Innovation, and the Synthesis of Evolutionary and Developmental Biology. Biol. Philos. 2003, 18, 309–345. [Google Scholar] [CrossRef]
- Griesemer, J. Towards a theory of Extended Developments. In Perspective on Evolutionary and Developmental Biology; Fusco, G., Ed.; Padova University Press: Padova, Italy, 2019; pp. 319–334. ISBN 978-88-6938-140-9. [Google Scholar]
- Craig, L.R. The So-Called Extended Synthesis and Population Genetics. Biol. Theory 2010, 5, 117–123. [Google Scholar] [CrossRef]
- Minelli, A.; Pradeu, T. (Eds.) Towards a Theory of Development; Oxford University Press: Oxford, UK, 2014; ISBN 9780199671434. [Google Scholar]
- Laubichler, M.D.; Prohaska, S.J.; Stadler, P.F. Toward a mechanistic explanation of phenotypic evolution: The need for a theory of theory integration. J. Exp. Zool. Part B Mol. Dev. Evol. 2018, 330, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Jenner, R.A. EvoDevo’s identity: From model organisms to developmental types. In Evolving Pathways: Key Themes in Evolutionary Developmental Biology; Minelli, A., Fusco, G., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 100–119. [Google Scholar]
- Minelli, A. Understanding Development; Cambridge University Press: Cambridge, UK, forthcoming.
- Pradeu, T.; Laplane, L.; Prévot, K.; Hoquet, T.; Reynaud, V.; Fusco, G.; Minelli, A.; Orgogozo, V.; Vervoort, M. Defining “Development”. Curr. Top. Dev. Biol. 2016, 117, 171–183. [Google Scholar] [CrossRef]
- Minelli, A. The Development of Animal Form; Cambridge University Press: Cambridge, UK, 2003; ISBN 9780521808514. [Google Scholar]
- Minelli, A. Forms of Becoming: The Evolutionary Biology of Development; Princeton University Press: Princeton, NJ, USA, 2009; ISBN 9780691135687. [Google Scholar]
- Minelli, A. Animal Development, an Open-Ended Segment of Life. Biol. Theory 2011, 6, 4–15. [Google Scholar] [CrossRef]
- Badyaev, A.V.; Uller, T. Parental effects in ecology and evolution: Mechanisms, processes and implications. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1169–1177. [Google Scholar] [CrossRef]
- Chiu, L.; Gilbert, S.F. The Birth of the Holobiont: Multi-species Birthing Through Mutual Scaffolding and Niche Construction. Biosemiotics 2015, 8, 191–210. [Google Scholar] [CrossRef]
- Caporael, L.R.; Griesemer, J.R.; Wimsatt, W.C. Developing Scaffold in Evolution, Culture and Cognition; The MIT Press: Cambridge, MA, USA, 2014; ISBN 9781461952367. [Google Scholar]
- Griffiths, P. Stotz, Genetics and Philosophy: An Introduction; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Griffiths, P.E.; Stotz, K. Genes in the Postgenomic Era. Theor. Med. Bioeth. 2006, 27, 499–521. [Google Scholar] [CrossRef]
- Gerstein, M.B.; Bruce, C.; Rozowsky, J.S.; Zheng, D.; Du, J.; Korbel, J.O.; Emanuelsson, O.; Zhang, Z.D.; Weissman, S.; Snyder, M. What is a gene, post-ENCODE? History and updated definition. Genome Res. 2007, 17, 669–681. [Google Scholar] [CrossRef]
- Shapiro, J.A. How life changes itself: The Read–Write (RW) genome. Phys. Life Rev. 2013, 10, 287–323. [Google Scholar] [CrossRef]
- Gilbert, S.F. The Reactive Genome. In Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology; Muller, G.B., Newman, S.A., Eds.; The MIT Press: Cambridge, MA, USA, 2003; pp. 87–101. ISBN 9780262280327. [Google Scholar]
- Lamm, E. The genome as a developmental organ. J. Physiol. 2014, 592, 2283–2293. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.B.; Pigliucci, M. Extended Synthesis: Theory Expansion or Alternative? Biol. Theory 2010, 5, 275–276. [Google Scholar] [CrossRef]
- Jablonka, E.; Lamb, M.J. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life; The MIT Press: Cambridge, MA, USA, 2005; ISBN 9780262525848 0262525844. [Google Scholar]
- Bonduriansky, R.; Day, T. Extended Heredity: A New Understanding of Inheritance and Evolution; Princeton University Press: Princeton, NJ, USA, 2018; ISBN 9781400890156. [Google Scholar]
- Gilbert, S.F. Genes Classical and Genes Developmental: The different use genes in Evolutionary Synthesis. In The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives; Falk, R., Rheinberger, H.-J., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 178–192. ISBN 0521060249 9780521060240. [Google Scholar]
- Moss, L. What Genes Can’t Do; The MIT Press: Cambridge, MA, USA, 2003; ISBN 9780262632973. [Google Scholar]
- Falk, R. The Gene: A concept in tension. In The Concept of the Gene in Development and Evolution: Historical and Epistemological Perspectives; Falk, R., Rheinberger, H.-J., Eds.; Cambridge University Press: New York, NY, USA, 2000; pp. 178–192. ISBN 0521060249 9780521060240. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amato, S.I. EvoDevo: An Ongoing Revolution? Philosophies 2020, 5, 35. https://doi.org/10.3390/philosophies5040035
Amato SI. EvoDevo: An Ongoing Revolution? Philosophies. 2020; 5(4):35. https://doi.org/10.3390/philosophies5040035
Chicago/Turabian StyleAmato, Salvatore Ivan. 2020. "EvoDevo: An Ongoing Revolution?" Philosophies 5, no. 4: 35. https://doi.org/10.3390/philosophies5040035
APA StyleAmato, S. I. (2020). EvoDevo: An Ongoing Revolution? Philosophies, 5(4), 35. https://doi.org/10.3390/philosophies5040035