
An Efficient Chromatin Immunoprecipitation Protocol for the Analysis of Histone Modification Distributions in the Brown Alga Ectocarpus

 ,
, 
Abstract
:1. Introduction
2. Experimental Design
2.1. Materials and Reagents
- Sporophytes and gametophytes of Ectocarpus male strain Ec32 (reference CCAP 1310/4 in the Culture Collection of Algae and Protozoa, Oban, UK)
- Sterile, filtered natural seawater
- Provasoli enrichment solution
- 140 mm Petri dishes (Corning, catalogue number: CLS430597)
- Parafilm (Sigma-Aldrich, Darmstadt, Germany, catalogue number P7793-1EA)
- Cell scraper (optional)
- Large-size cell strainer (for example, a 12.5 cm Finlandek permanent coffee filter) (optional, for tissue harvesting after culture)
- Dissection forceps
- 1 L Erlenmeyer flask
- 37% Formaldehyde
- PBS pH 7.5
- Sterile Miracloth (Millipore, Watford, UK, ref. 475855)
- 50 mL plastic Falcon tubes (Cellstar®, Bishop Stortford, UK, ref. 227 261)
- Refrigerated centrifuge for 50 mL plastic Falcon tubes (Eppendorf 5804R)
- Liquid nitrogen
- Pestle and mortar
- Pre-cooled spatula to transfer ground tissue from the mortar
- 7 mL Tenbroeck potter (Kontes, Mainz, Germany ref. 885000-0007)
- 15 mL plastic Falcon tubes (Cellstar®, Bishop Stortford, UK, ref. 188 271)
- 1.5 mL and 2 mL microtubes
- Refrigerated centrifuge for microtubes (Eppendorf 5427 R; maximum RCF 14,000× g)
- DAPI stock solution (2 mg/mL)
- Microscope slides
- Microscope coverslips (Knittel ref. MS0009)
- Covaris® microTUBEs (Covaris, Brighton, UK, AFA Fiber Pre-Slit Snap-Cap 6 mm × 16 mm, ref. 520045)
- SmartLadder and SmartLadder SF DNA size markers (Eurogentech, Seraing, Belgium, MW-1700-02 and MW-1800-02)
- Normal rabbit IgG naïve antibody (CST 2729)
- ChIP-grade antibodies against histone modifications of interest
- DynaBeads protein A (Invitrogen ref. 10002D)
- DynaBeads protein G (Invitrogen ref. 10004D)
- 1.5 mL and 2 mL Eppendorf® DNA LoBind microtubes (Cat. N° 022431021 and 022431048)
- Magnetic microtube rack (Invitrogen, Carlsbad, CA, USA, ref. 12321D)
- Glycine
- 5 M NaCl stock solution
- 0.5 M EDTA pH 8 stock solution
- 1 M Tris-HCl pH 6.5 stock solution
- Proteinase K 20 mg/mL (Invitrogen, Carlsbad, CA, USA, ref. AM2546)
- RNAse A, DNase, and protease-free 10 mg/mL (Thermo Fisher Scientific ref. EN0531)
- Pre-mixed phenol:chloroform:isoamyl alcohol (25:24:1)
- 100% Ethanol
- 70% (v/v) Ethanol
- 3 M Sodium acetate pH 5.2
- Glycogen (Thermo Fisher Scientific, Loughborough, UK, ref. R0561)
- MilliQ water
- High Sensitivity DNA (Agilent, Santa Clara, CA, USA, ref. 5076-4626)
- Qubit™ 1X High Sensitivity dsDNA (Invitrogen™ ref. Q33230)
2.2. Equipment
- Thermostatically controlled, illuminated growth cabinet (POL-EKO-APARATURE, Wodzisław Śląski, Poland, model: KK 240 FIT P) or growth room
- Thermomixer (Eppendorf, Saint-Quentin-Fallavier, France)
- Fluorescence microscope (Olympus, Southend-on-Sea, UK, model: CKX41)
- Covaris® M220 Focused-ultrasonicator
- Agarose gel electrophoresis system
- Bioanalyzer 2100 (Agilent, Santa Clara, CA, USA)
- Qubit 4.0 (Life Technologies, Paisley, UK)
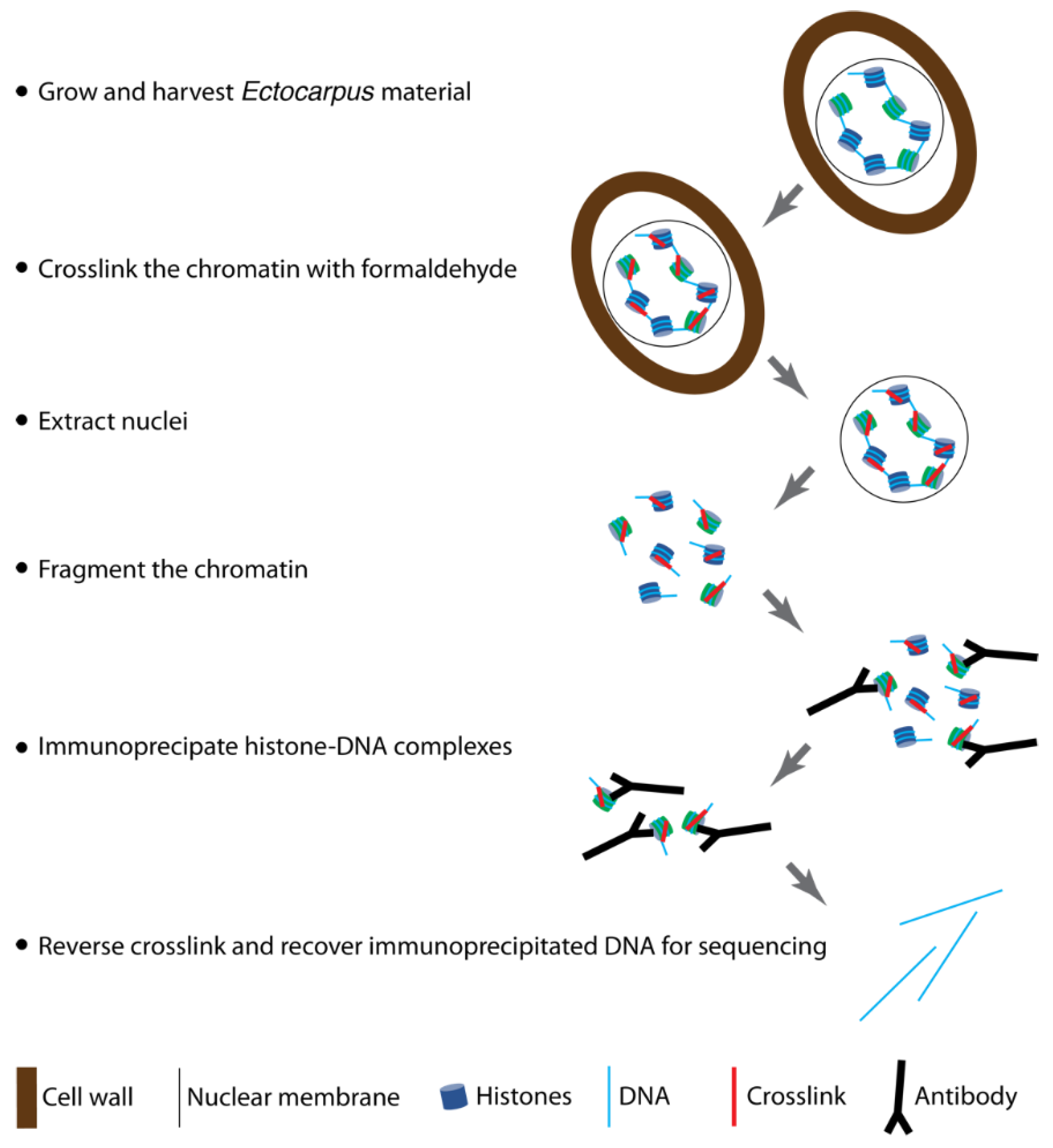
3. Procedure
3.1. Growth and Harvesting of Ectocarpus Cultures (Required Time: 2 Weeks)
- Inoculate 75 mL aliquots of Provasoli’s enriched natural seawater in transparent 140 mm diameter Petri dishes with the appropriate Ectocarpus strain, seal with Parafilm, and grow at 13 °C with a 12 h day/12 h night cycle and 20 µmol photons.m−2.s−1 irradiance [24]. Culture at least 1200 individual sporophytes or gametophytes at a density of six individuals per Petri dish.
- After culture, carefully transfer the algae with dissection forceps into a sterile Erlenmeyer flask containing 400 mL of sterile seawater. Two weeks of culture should produce approximately 4–5 g (FW) of tissue.
 CRITICAL STEP For some strains it may be necessary to grow more individuals per Petri dish and harvest at an earlier stage of development before cultures become fertile and produce the next generation of the life cycle. With some sporophyte strains it may be necessary to use a cell scraper to dislodge thalli from the bottom of the Petri dish. This material can be collected by filtering the seawater medium and then transferring the algal material with dissection forceps to the 400 mL of sterile seawater.
CRITICAL STEP For some strains it may be necessary to grow more individuals per Petri dish and harvest at an earlier stage of development before cultures become fertile and produce the next generation of the life cycle. With some sporophyte strains it may be necessary to use a cell scraper to dislodge thalli from the bottom of the Petri dish. This material can be collected by filtering the seawater medium and then transferring the algal material with dissection forceps to the 400 mL of sterile seawater.3.2. Crosslinking (Required Time: 2 h)
- 3.
- Crosslink the algal material by adding 400 mL of 2x crosslinking buffer to the 400 mL of algal material in seawater and incubating for exactly 5 min at room temperature under a chemical hood. Mix gently during the crosslinking.
 CRITICAL STEP Do not incubate for more than 5 min in the 2x crosslinking buffer as this could lead to excessive crosslinking, which may interfere with the chromatin extraction and immunoprecipitation steps. Note that formaldehyde is toxic if inhaled, ingested, or absorbed through skin.
CRITICAL STEP Do not incubate for more than 5 min in the 2x crosslinking buffer as this could lead to excessive crosslinking, which may interfere with the chromatin extraction and immunoprecipitation steps. Note that formaldehyde is toxic if inhaled, ingested, or absorbed through skin.- 4.
- Filter the tissue rapidly (15 s maximum) through a sterile piece of Miracloth to eliminate the formaldehyde. Transfer the tissue to a new 50 mL tube containing 50 mL of PBS quenching buffer for 5 min at room temperature. Mix gently during quenching.
- 5.
- Centrifuge at 3215× g 4 °C in an Eppendorf 5804R for 5 min for gametophytes or for 10 min for sporophytes.
- 6.
- Eliminate the buffer and resuspend in 50 mL of 1x phosphate-buffered saline (PBS) to wash the tissue. Centrifuge at 3215× g 4 °C in an Eppendorf 5804R for 5 min for gametophytes, or for 10 min for sporophytes.
- 7.
- Remove as much of the supernatant as possible by pipetting. You can invert the tube onto a piece of Miracloth placed on absorbent paper. Wrap approximately 1 g batches of tissue in aluminium foil, note the mass of tissue in each batch as this will be used to calculate the volume of nuclei isolation buffer to be added in the following steps. Freeze the crosslinked tissue rapidly in liquid nitrogen.
 PAUSE STEP The crosslinked tissue can be stored at −80 °C at this stage, but it should be used within the next few days if possible, and should not be stored for more than 1 month.
PAUSE STEP The crosslinked tissue can be stored at −80 °C at this stage, but it should be used within the next few days if possible, and should not be stored for more than 1 month. CRITICAL STEP The filamentous structure of the material may be partly dissociated after fixation, making it difficult to recover the pellet. If this is the case, scoop the pellet out of the Falcon tube with a spatula.
CRITICAL STEP The filamentous structure of the material may be partly dissociated after fixation, making it difficult to recover the pellet. If this is the case, scoop the pellet out of the Falcon tube with a spatula.3.3. Isolation of Semipure Nuclei (Required Time: 2 h)
- 8.
- Grind about 1 g of crosslinked tissue to an ultra-fine powder under liquid nitrogen using a pre-chilled mortar and pestle. Ensure that samples do not thaw during grinding.
 CRITICAL STEP It is important that the tissue is very thoroughly ground at this stage. To avoid cross-contamination use different mortars and pestles for different experimental conditions.
CRITICAL STEP It is important that the tissue is very thoroughly ground at this stage. To avoid cross-contamination use different mortars and pestles for different experimental conditions.- 9.
- Transfer the powder to a 15 mL tube containing pre-chilled nuclei isolation buffer with Triton X-100, β-mercaptoethanol, and protease inhibitor cocktail (approximatively 5 mL of buffer for 1 g of tissue). Resuspend well by pipetting up and down.
- 10.
- Transfer the extract into a 7 mL Tenbroeck potter. Grind 10 times slowly on ice, making hemicircular movements of the potter in the tube when you insert and remove it. To avoid cross-contamination, do not use the same Tenbroeck potter for the different experimental samples.
 CRITICAL STEP To effectively break open the Ectocarpus cells, the pestle of the Tenbroeck potter should fit tightly into the cylinder, and it should be quite difficult to move the pestle down the cylinder while making the hemicircular movements. When testing a Tenbroeck potter that has not been used previously, it is important to verify cell lysis using DAPI staining (see below). Grinding is less efficient if too much tissue is extracted; do not exceed 1.5 g.
CRITICAL STEP To effectively break open the Ectocarpus cells, the pestle of the Tenbroeck potter should fit tightly into the cylinder, and it should be quite difficult to move the pestle down the cylinder while making the hemicircular movements. When testing a Tenbroeck potter that has not been used previously, it is important to verify cell lysis using DAPI staining (see below). Grinding is less efficient if too much tissue is extracted; do not exceed 1.5 g.- 11.
- Incubate on ice for 20 min. Resuspend every 5 min.
- 12.
- Pre-wet two layers of Miracloth by pipetting 500 µL of nuclei isolation buffer with Triton X-100, β-mercaptoethanol, and protease inhibitor cocktail, and then filter the extract through the Miracloth into a 15 mL conical tube on ice. The two layers of Miracloth should be rotated at 90° to each other. Squeeze the Miracloth well to remove all liquid. This step is necessary to remove large debris.
- 13.
- Aliquot the filtered extract into several 2 mL microtubes and centrifuge for 20 min at 3000× g and 4 °C.
- 14.
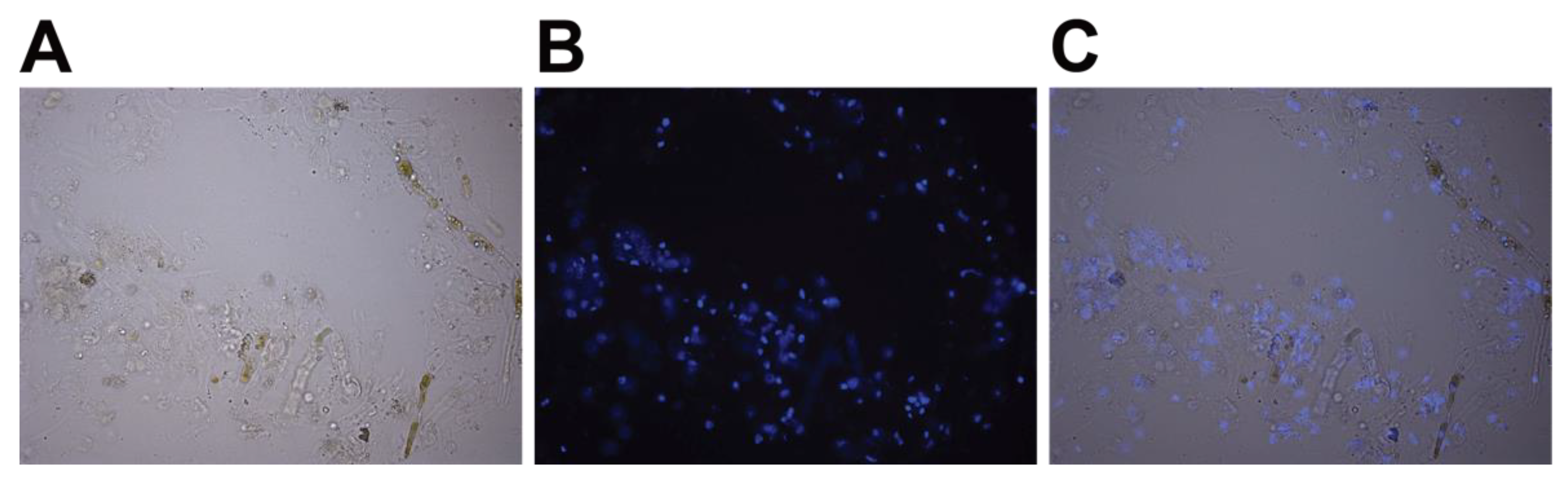
- Remove the supernatant and gently resuspend and combine the pellets from the same sample in 1 mL of nuclei isolation buffer with Triton X-100, β-mercaptoethanol, and protease inhibitor cocktail. Centrifuge the extract for 20 min at 3000× g and 4 °C. Repeat the wash with 1 mL of nuclei isolation buffer with Triton X-100, β-mercaptoethanol, and protease inhibitor cocktail, then centrifuge the extract for 20 min at 3000× g and 4 °C. The pellet should be a pale yellow or whitish colour at this stage, and the supernatant pale brown (Figure 2).
- 15.
- Remove the supernatant and gently resuspend the pellets in 1 mL of nuclei isolation buffer with β-mercaptoethanol and protease inhibitor cocktail but without Triton X-100.
- 16.
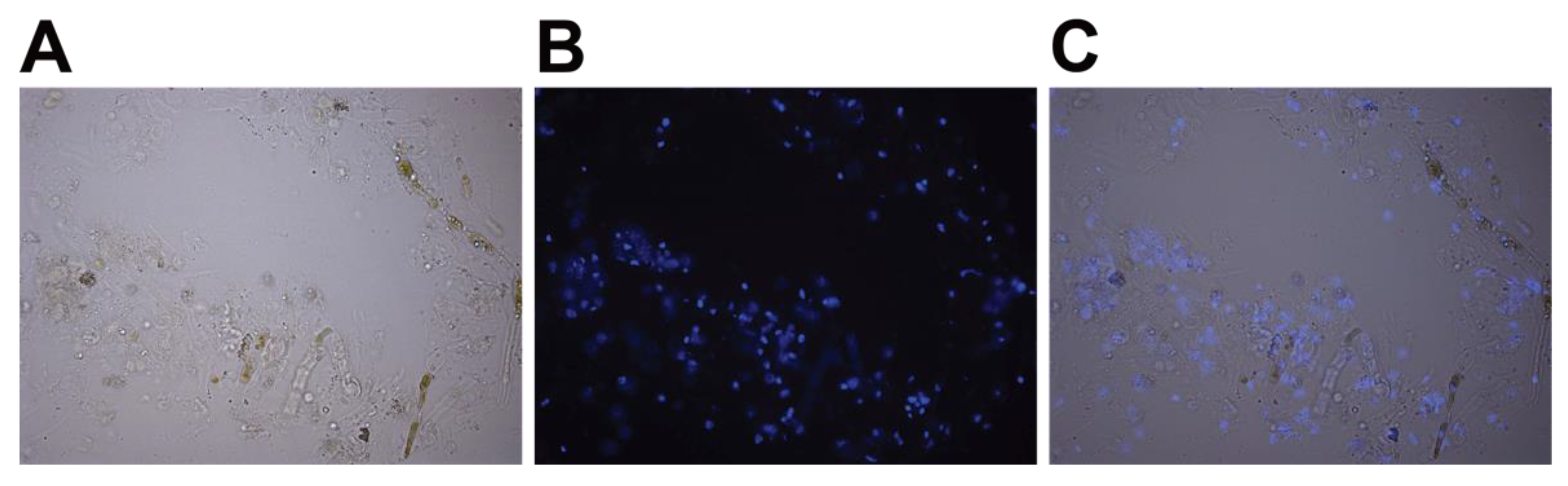
- OPTIONAL STEP Verify the release of the nuclei using microscopy. Dilute 1 µL of stock solution of 2 mg/mL DAPI in water (stored at −20 °C) in 100 µL of nuclei isolation buffer without Triton X-100, and then add 2 µL of this 10x preparation to 20 µL of extract. Incubate at room temperature for 10 min, then place between a slide and coverslip and visualize under a fluorescence microscope (Figure 3).
 CRITICAL STEP DAPI is toxic and a mutagen. Wear gloves while working with DAPI.
CRITICAL STEP DAPI is toxic and a mutagen. Wear gloves while working with DAPI.3.4. Lysis of Nuclei and Sonication (Required Time: 3 h)
- 17.
- Transfer to a new 2 mL microtube and centrifuge for 20 min at 3000× g and 4 °C. Prepare the nuclei lysis buffer and ChIP dilution buffer at this stage and cool to 4 °C.
- 18.
- Remove the supernatant and resuspend the pellet in 200 µL to 1 mL (depending on the quantity of starting tissue) of cold nuclei lysis buffer. For example, use 1 mL of nuclei lysis buffer for 1 g of tissue.
- 19.
- Keep a 5 µL aliquot of the chromatin extract to run on an agarose gel (see below) in order to compare with the chromatin extract after sonication.
 PAUSE STEP If necessary, the chromatin extract can be frozen at −80 °C overnight at this stage, but it is better to continue directly to the sonication step if possible.
PAUSE STEP If necessary, the chromatin extract can be frozen at −80 °C overnight at this stage, but it is better to continue directly to the sonication step if possible.- 20.
- Pipette 130 µL aliquots of the chromatin extract into new, clean Covaris® microTUBEs (AFA Fiber Pre-Slit Snap-Cap 6 mm × 16 mm). It is important to add exactly 130 µL to each microTUBE and to avoid foaming to ensure complete fragmentation.
- 21.
- Sonicate the chromatin extract in a Covaris® M220 Focused-ultrasonicator™ using the following parameters: duty 25%, peak power 75, cycles/burst 200, time duration 900 s, set point temperature 6 °C (range between 4 °C and 7 °C). Note that sonication could alternatively be carried out with another sonicator, such as the Diagenode Bioruptor Pico, with minor optimization.
- 22.
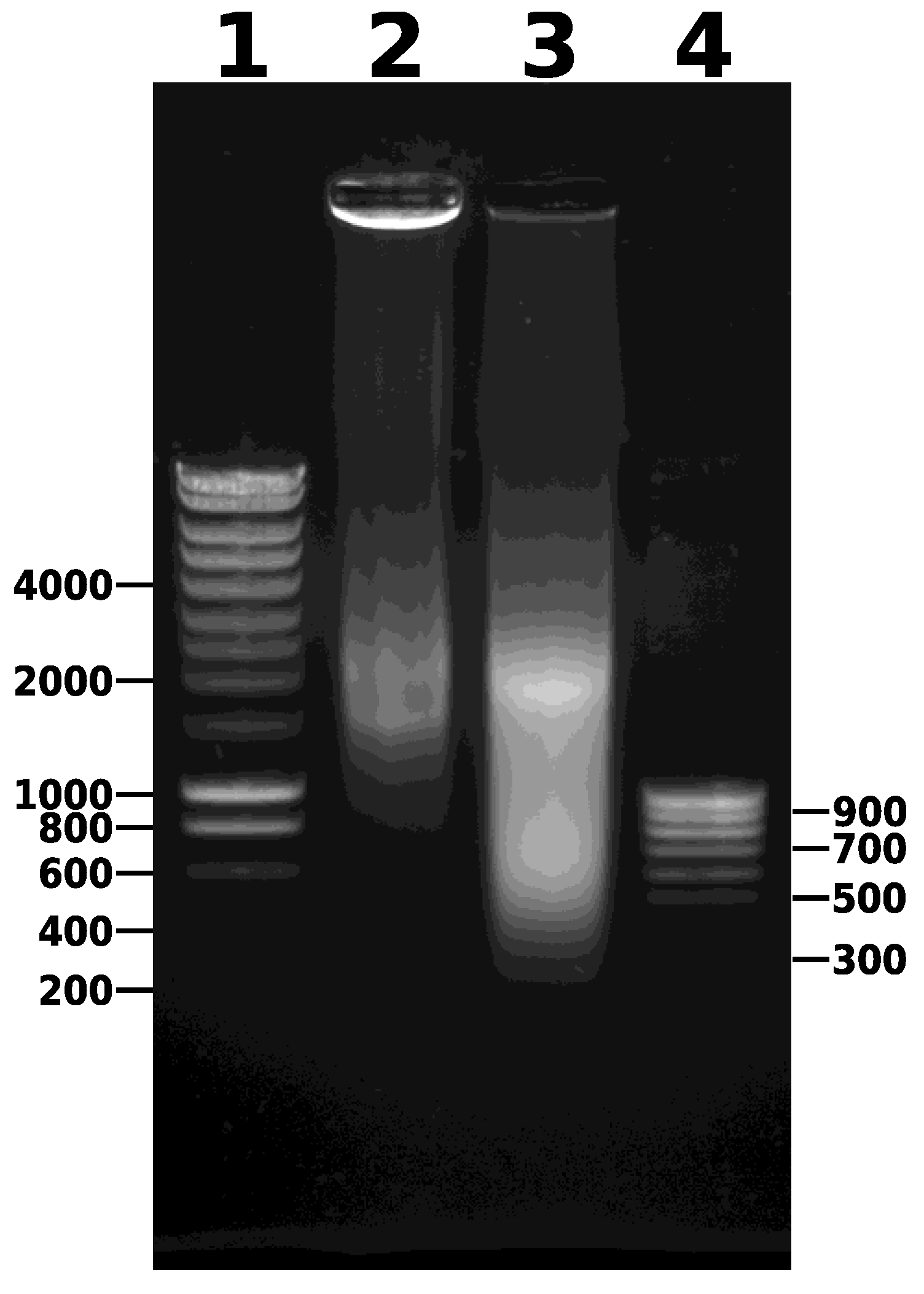
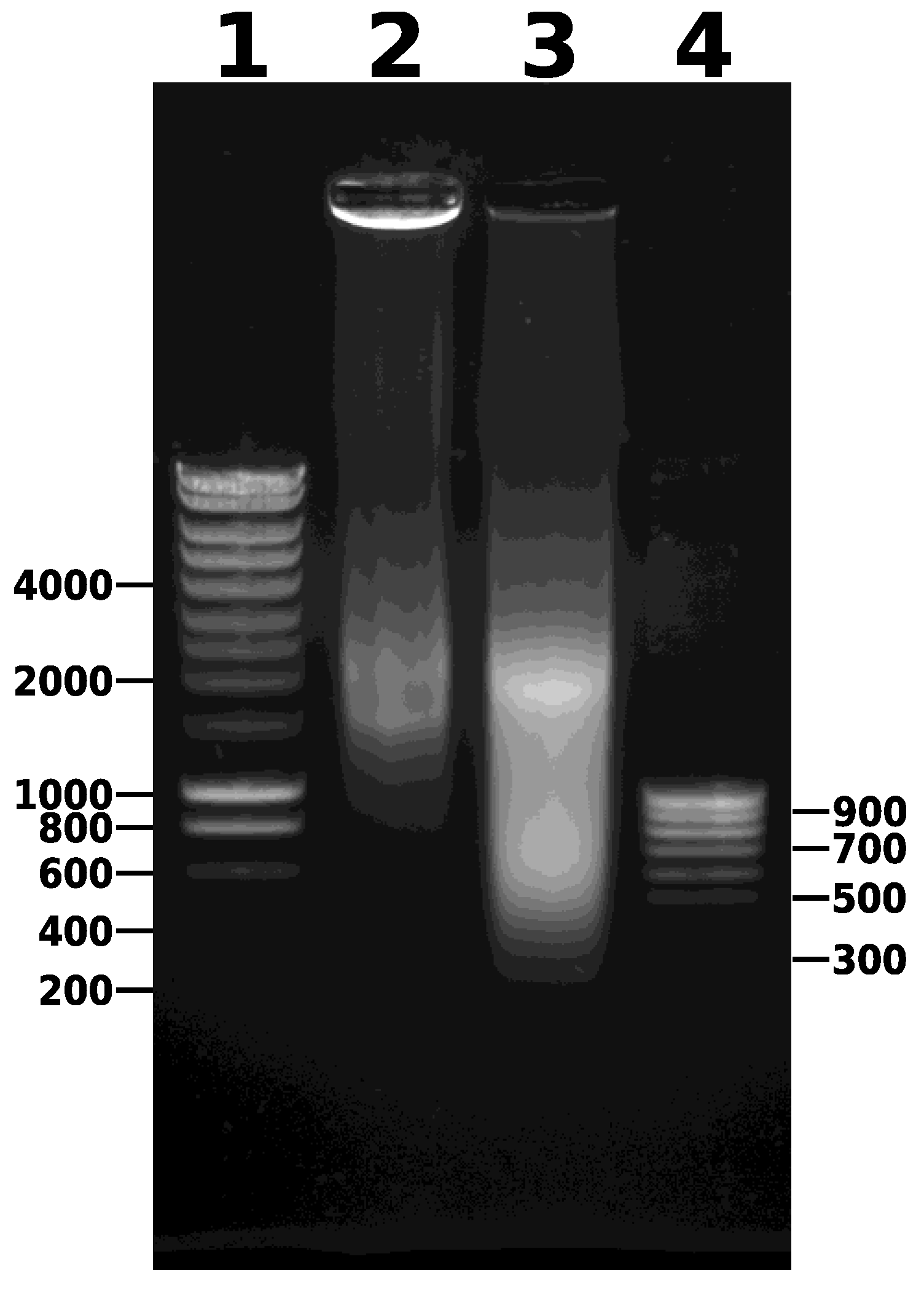
- Combine the contents of the microTUBEs after sonication and centrifuge for 5 min at 14,000× g and 4 °C to pellet the debris (Figure 2). Transfer the supernatant to a new 1.5 mL microtube. Keep 5 µL of this sample to run on a 0.8% agarose gel (along with the unsonicated sample taken earlier) to visualize the DNA fragments after sonication. After gel electrophoresis, a smear of sonicated chromatin should be observed between 100 bp and 1000 bp (Figure 4). Note that the samples are crosslinked chromatin at this stage (i.e., not naked DNA), so precise estimation of fragment size is not possible; however, gel electrophoresis provides an approximate estimation of the efficiency of fragmentation.
 CRITICAL STEP Note that the centrifugation step is essential to remove impurities that would otherwise interfere with and contaminate the immunoprecipitation step.
CRITICAL STEP Note that the centrifugation step is essential to remove impurities that would otherwise interfere with and contaminate the immunoprecipitation step.- 23.
- Measure the volume of supernatant and dilute tenfold with ChIP dilution buffer. For each sample, keep 50 μL of diluted, sonicated chromatin as an input control. The chromatin extract can be used directly to set up immunoprecipitations, or stored at –20 °C or −80 °C. Note that ChIP dilution buffer is also required to prepare the DynaBeads (Section 3.5). Store an aliquot of ChIP dilution buffer at 4 °C overnight for this.
 CRITICAL STEP Note that it is important to dilute from 1% to 0.1% SDS by adding ChIP dilution buffer because a high concentration of SDS can interfere with epitope–antibody interactions.
CRITICAL STEP Note that it is important to dilute from 1% to 0.1% SDS by adding ChIP dilution buffer because a high concentration of SDS can interfere with epitope–antibody interactions.3.5. Immunoprecipitation (Required Time: 24 h)
- 24.
- Prepare 500 μL aliquots of diluted chromatin (equivalent to 100 mg of starting tissue) in 1.5 mL Eppendorf DNA LoBind® microtubes. Add the recommended volume of your antibody. The volume is 5 µL for most antibodies, but optimal volumes may vary and should be determined empirically. As a negative control, add a naïve antibody, such as normal rabbit IgG CST N° 2729. As a mock control, carry out an immunoprecipitation without any antibody. Co-incubate the sonicated chromatin and antibody overnight at 4 °C on a rotating wheel (10 rpm).
- 25.
- The next day, prepare low-salt wash buffer, high-salt wash buffer, LiCl wash buffer, TE buffer, and elution buffer. Place the first four buffers on ice to cool and pre-heat the elution buffer to 65 °C.
- 26.
- Prior to use, resuspend DynaBeads protein A and DynaBeads protein G by vortexing. Pre-equilibrate the DynaBeads by pipetting 50 µL of each type of bead into separate, new microtubes, place in the magnetic rack, discard the supernatant and add 50 µL of ChIP dilution buffer, mix well by pipetting up and down. Repeat this wash twice and resuspend each lot of beads in 50 µL of ChIP dilution buffer.
- 27.
- Mix DynaBeads protein A and DynaBeads protein G in an equal volume (ratio 1:1). There should be 100 µL of beads for each immunoprecipitation.
- 28.
- Add 100 µL of pre-equilibrated beads to each 1.5 mL Eppendorf® DNA LoBind microtube containing chromatin extract plus antibody (i.e., to all the tubes from step 24). Mix for 2 h at 4 °C with gentle rotation (on a rotating wheel at 10 rpm).
- 29.
- Separate the supernatant and DynaBeads in a magnetic rack.
- 30.
- Carry out all washing steps in a cold room, if possible, otherwise keep the samples on ice. Collect immune complexes by placing the tubes in the magnetic rack for 2 min. Remove the supernatant.
- 31.
- Wash the DynaBeads–IgG–antigen–chromatin complexes twice with each of the following four buffers: low-salt wash buffer, high-salt wash buffer, LiCl wash buffer, and TE Buffer. For each buffer, carry out a rapid first wash, replacing the tube immediately in the magnetic rack, and then a 5 min second wash with the beads in suspension. Use 1 mL of each buffer and wash at 4 °C using only brief, gentle hand inversions to resuspend the beads. To recover the beads, place the tubes in the magnetic rack for 2 min. During this time, invert the rack once to recover any beads in the tube cover. After the 2 min, carefully remove the supernatant and add the next buffer.
- 32.
- Elute the immune complexes by adding 250 µL of elution buffer (made fresh at step 25 and pre-warmed to 65 °C) to the washed beads. Vortex and incubate in a Eppendorf Thermomixer at 65 °C for 20 min at 1000 rpm. Place the tubes in the magnetic rack and carefully transfer the supernatant fraction (eluate) to another Eppendorf® DNA LoBind tube. Repeat the elution with an additional 250 µL of elution buffer, and combine the two eluates. At the same time, add 450 µL of elution buffer to the 50 µL input control (positive control).
3.6. Reverse Crosslinking and RNA/Protein Digestion (Required Time: 18 h)
- 33.
- Add 20 µL 5 M NaCl to each tube and incubate at 65 °C for at least 12 h, or overnight, to reverse the crosslinking.
- 34.
- Add 10 µL of 0.5 M EDTA (pH 8), 20 µL of 1 M Tris-HCl (pH 6.5), 1 µL of 20 mg.mL−1 proteinase K, and 1 µL of 10 mg.mL−1 RNAse A to the eluate and incubate for 1.5 h at 45 °C.
3.7. DNA Extraction and Precipitation (Required Time: 2 h)
- 35.
- Add an equal volume (550 µL) of phenol:chloroform:isoamyl alcohol (25:24:1) and vortex briefly.
- 36.
- Centrifuge the samples in a microcentrifuge at 13,800× g for 15 min at 4 °C. Transfer the supernatant to a new 2 mL Eppendorf® DNA LoBind microtube.
- 37.
- To each tube, add 1.25 mL of 100% ethanol, 50 µL of 3 M sodium acetate (pH 5.2), and 4 µL glycogen (20 mg.mL−1). Incubate for 1 h, or overnight, at −80 °C to precipitate the DNA.
- 38.
- Centrifuge each sample at 13,800× g for 15 min at 4 °C.
- 39.
- Discard the supernatant, wash the pellet with 500 µL of 70% (v/v) ethanol, centrifuge again at 13,800× g for 10 min at 4 °C.
- 40.
- Discard the supernatant and dry the pellet briefly at room temperature.
 CRITICAL STEP Take care not to over-dry the glycogen-containing pellets as they may become insoluble.
CRITICAL STEP Take care not to over-dry the glycogen-containing pellets as they may become insoluble.- 41.
- Resuspend the pellets in 30 µL of DEPC water, store the DNA at −80 °C, and use within 3 months. Traces of phenol may be detected at this stage, but they do not interfere with library construction.
3.8. DNA Analysis (Required Time: 2 h)
- 42.
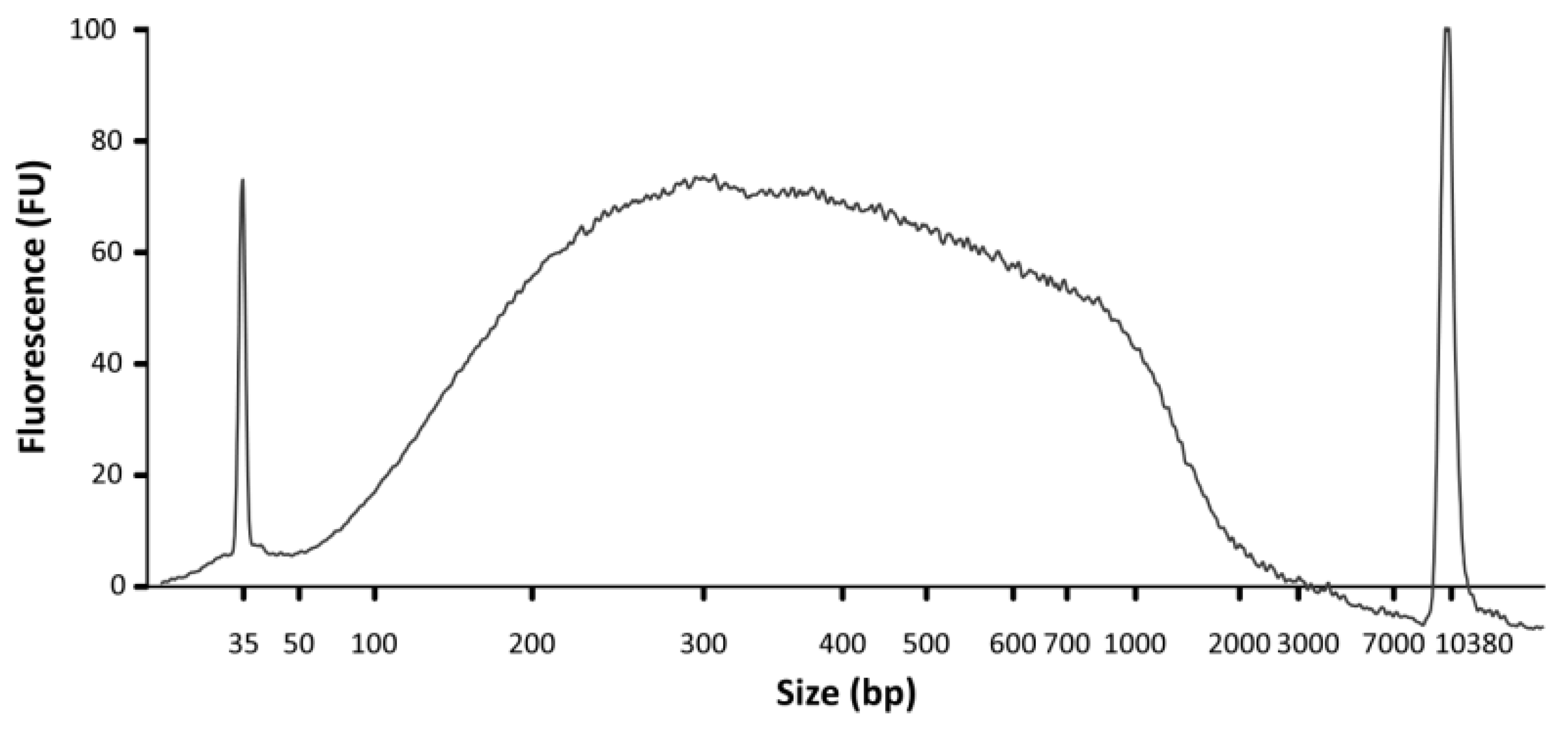
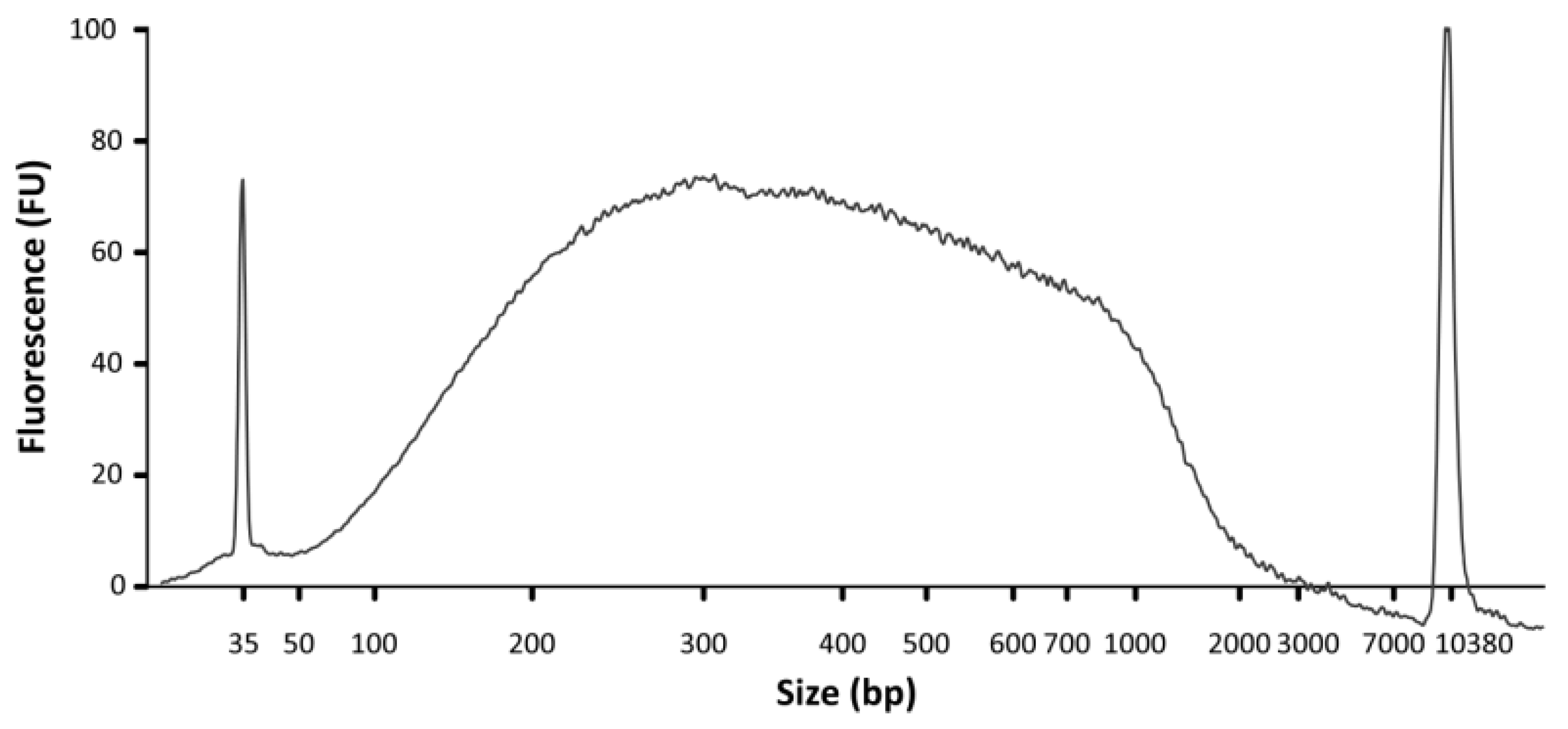
- DNA concentration and size range can be analyzed using a Bioanalyzer and a High Sensitivity DNA Chip (Agilent; Figure 5). In addition, DNA concentration should be assessed using a Qubit fluorometer and a dsDNA HS Assay kit.
4. Expected Results
5. Reagent Setup
Solutions
- 2x Crosslinking buffer (2% formaldehyde in natural seawater);
- PBS quenching buffer (1x PBS:2M glycine in a 4:1 ratio, the final concentration of glycine is 400 mM, PBS is at pH 7.5);
- Protease inhibitor cocktail (one Roche EDTA-free cOmplete ULTRA tablet dissolved in 1 mL MilliQ water—can be prepared in advance and frozen at −20 °C);
- Nuclei isolation buffer (125 mM sorbitol, 20 mM potassium citrate, 30 mM MgCl2, 5 mM EDTA, 55 mM HEPES pH 7.5; just before use, add 0.1% Triton X-100, 5 mM β-mercaptoethanol, and 2% protease inhibitor cocktail—note that nuclei isolation buffer can be stored for several months at 4 °C);
- Nuclei isolation buffer without Triton X-100 (125 mM sorbitol, 20 mM potassium citrate, 30 mM MgCl2, 5 mM EDTA, 55 mM HEPES pH 7.5; just before use, add 5 mM β-mercaptoethanol and 2% protease inhibitor cocktail);
- Nuclei lysis buffer (10 mM EDTA, 1% SDS, 50 mM Tris-HCl pH 8, 10% protease inhibitor cocktail—can be prepared in advance and kept at 4 °C);
- ChIP dilution buffer (1% Triton X-100, 1.2 mM EDTA, 167 mM NaCl, 16.7 mM Tris-HCl pH 8.0—note that ChIP dilution buffer can be stored for several months at 4 °C);
- Low-salt wash buffer (150 mM NaCl, 0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl pH 8.0);
- High-salt wash buffer (500 mM NaCl, 0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl pH 8.0);
- LiCl wash buffer (0.25 M LiCl, 1% NP40, 1% sodium deoxycholate, 1 mM EDTA, 10 mM Tris-HCl pH 8.0);
- TE buffer (1 mM EDTA, 10 mM Tris-HCl pH 8.0);
- Elution buffer (1% SDS, 0.1 M NaHCO3).
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klinger, T. The Role of Seaweeds in the Modern Ocean. Perspect. Phycol. 2015, 2, 31–39. [Google Scholar] [CrossRef]
- Duarte, C.M.; Wu, J.; Xiao, X.; Bruhn, A.; Krause-Jensen, D. Can Seaweed Farming Play a Role in Climate Change Mitigation and Adaptation? Front. Mar. Sci. 2017, 4, 100. [Google Scholar] [CrossRef]
- Cai, J.; Lovatelli, A.; Aguilar-Manjarrez, J.; Cornish, L.; Dabbadie, L.; Desrochers, A.; Diffey, S.; Garrido Gamarro, E.; Geehan, J.; Hurtado, A.; et al. Seaweeds and Microalgae: An Overview for Unlocking Their Potential in Global Aquaculture Development. FAO Fish. Aquac. Circ. 2021, 1229. [Google Scholar] [CrossRef]
- Coelho, S.; Cock, J. Brown Algal Model Organisms. Ann. Rev. Genet. 2020, 54, 71–92. [Google Scholar] [CrossRef]
- Cock, J.M.; Collén, J. Independent Emergence of Complex Multicellularity in the Brown and Red Algae. In Evolutionary Transitions to Multicellular Life; Ruiz-Trillo, I., Nedelcu, A.M., Eds.; Advances in Marine Genomics; Springer: Berlin/Heidelberg, Germany, 2015; pp. 335–361. [Google Scholar]
- Peters, A.F.; Marie, D.; Scornet, D.; Kloareg, B.; Cock, J.M. Proposal of Ectocarpus siliculosus (Ectocarpales, Phaeophyceae) as a Model Organism for Brown Algal Genetics and Genomics. J. Phycol. 2004, 40, 1079–1088. [Google Scholar] [CrossRef]
- Cock, J.M.; Sterck, L.; Rouzé, P.; Scornet, D.; Allen, A.E.; Amoutzias, G.; Anthouard, V.; Artiguenave, F.; Aury, J.; Badger, J.; et al. The Ectocarpus Genome and the Independent Evolution of Multicellularity in Brown Algae. Nature 2010, 465, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Cormier, A.; Avia, K.; Sterck, L.; Derrien, T.; Wucher, V.; Andres, G.; Monsoor, M.; Godfroy, O.; Lipinska, A.; Perrineau, M.-M.; et al. Re-Annotation, Improved Large-Scale Assembly and Establishment of a Catalogue of Noncoding Loci for the Genome of the Model Brown Alga Ectocarpus. New Phytol. 2017, 214, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Macaisne, N.; Liu, F.; Scornet, D.; Peters, A.F.; Lipinska, A.; Perrineau, M.-M.; Henry, A.; Strittmatter, M.; Coelho, S.M.; Cock, J.M. The Ectocarpus IMMEDIATE UPRIGHT Gene Encodes a Member of a Novel Family of Cysteine-Rich Proteins with an Unusual Distribution across the Eukaryotes. Development 2017, 144, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badis, Y.; Scornet, D.; Harada, M.; Caillard, C.; Godfroy, O.; Raphalen, M.; Gachon, C.M.M.; Coelho, S.M.; Motomura, T.; Nagasato, C.; et al. Targeted CRISPR-Cas9-Based Gene Knockouts in the Model Brown Alga Ectocarpus. New Phytol. 2021, 231, 2077–2091. [Google Scholar] [CrossRef] [PubMed]
- Arun, A.; Coelho, S.M.; Peters, A.F.; Bourdareau, S.; Pérès, L.; Scornet, D.; Strittmatter, M.; Lipinska, A.P.; Yao, H.; Godfroy, O.; et al. Convergent Recruitment of TALE Homeodomain Life Cycle Regulators to Direct Sporophyte Development in Land Plants and Brown Algae. eLife 2019, 8, e43101. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.F.; Scornet, D.; Ratin, M.; Charrier, B.; Monnier, A.; Merrien, Y.; Corre, E.; Coelho, S.M.; Cock, J.M. Life-Cycle-Generation-Specific Developmental Processes Are Modified in the immediate upright Mutant of the Brown Alga Ectocarpus siliculosus. Development 2008, 135, 1503–1512. [Google Scholar] [CrossRef] [Green Version]
- Godfroy, O.; Uji, T.; Nagasato, C.; Lipinska, A.P.; Scornet, D.; Peters, A.F.; Avia, K.; Colin, S.; Mignerot, L.; Motomura, T.; et al. DISTAG/TBCCd1 Is Required for Basal Cell Fate Determination in Ectocarpus. Plant Cell 2017, 29, 3102–3122. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Cock, J.M.; Pessia, E.; Luthringer, R.; Cormier, A.; Robuchon, M.; Sterck, L.; Peters, A.F.; Dittami, S.M.; Corre, E.; et al. A Haploid System of Sex Determination in the Brown Alga Ectocarpus sp. Curr. Biol. 2014, 24, 1945–1957. [Google Scholar] [CrossRef] [Green Version]
- Dittami, S.; Scornet, D.; Petit, J.; Ségurens, B.; Da Silva, C.; Corre, E.; Dondrup, M.; Glatting, K.; König, R.; Sterck, L.; et al. Global Expression Analysis of the Brown Alga Ectocarpus siliculosus (Phaeophyceae) Reveals Large-Scale Reprogramming of the Transcriptome in Response to Abiotic Stress. Genome Biol. 2009, 10, R66. [Google Scholar] [CrossRef] [Green Version]
- Lipinska, A.; Cormier, A.; Luthringer, R.; Peters, A.F.; Corre, E.; Gachon, C.M.M.; Cock, J.M.; Coelho, S.M. Sexual Dimorphism and the Evolution of Sex-Biased Gene Expression in the Brown Alga Ectocarpus. Mol. Biol. Evol. 2015, 32, 1581–1597. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.; Daujat, S.; Schneider, R. Lateral Thinking: How Histone Modifications Regulate Gene Expression. Trends Genet. 2016, 32, 42–56. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Han, W.; Teng, L.; Jiang, P.; Zhang, X.; Xu, D.; Li, C.; Pellegrini, M.; Wu, C.; Wang, Y.; et al. Single-Base Methylome Profiling of the Giant Kelp Saccharina japonica Reveals Significant Differences in DNA Methylation to Microalgae and Plants. New Phytol. 2020, 225, 234–249. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Tirichine, L.; Bowler, C. Protocol: Chromatin Immunoprecipitation (ChIP) Methodology to Investigate Histone Modifications in Two Model Diatom Species. Plant Methods 2012, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Coelho, M.A.; Shu, H.; Schotanus, K.; Thimmappa, B.C.; Yadav, V.; Chen, H.; Malc, E.P.; Wang, J.; Mieczkowski, P.A.; et al. Long Transposon-Rich Centromeres in an Oomycete Reveal Divergence of Centromere Features in Stramenopila-Alveolata-Rhizaria Lineages. PLoS Genet. 2020, 16, e1008646. [Google Scholar] [CrossRef]
- Bourdareau, S.; Tirichine, L.; Lombard, B.; Loew, D.; Scornet, D.; Wu, Y.; Coelho, S.M.; Cock, J.M. Histone Modifications during the Life Cycle of the Brown Alga Ectocarpus. Genome Biol. 2021, 22, 12. [Google Scholar] [CrossRef]
- Gueno, J.; Cossard, G.; Bourdareau, S.; Godfroy, O.; Lipinska, A.; Tirichine, L.; Cock, J.; Coelho, S. Chromatin Landscape Associated with Sexual Differentiation in a UV Sex Determination System. Nucl. Acids Res. 2022, in press. [Google Scholar] [CrossRef]
- Bourdareau, S. Genetic and Epigenetic Control of Life Cycle Transitions in the Brown Alga Ectocarpus sp. Ph.D. Thesis, Sorbonne Université, Paris, France, 2018. [Google Scholar]
- Coelho, S.M.; Scornet, D.; Rousvoal, S.; Peters, N.T.; Dartevelle, L.; Peters, A.F.; Cock, J.M. How to Cultivate Ectocarpus. Cold Spring Harb. Protoc. 2012, 2012, 258–261. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histone PTM or Control Sample | Antibody | DNA Yield/Fixed Material (ng/mg) |
|---|---|---|
| Input | n/a | 10–25 |
| Normal Rabbit IgG | (polyclonal) CST (ref:#2729) | 0.02–0.05 |
| Histone H3 | (D2B12) CST (ref:#4620) | 2–5 |
| H3K4me3 | (C42D8) CST (ref:#9751) | 0.1–0.3 |
| H3K9ac | (C5B11) CST (ref:#9649) | 0.2–0.8 |
| H4K20me3 | (D84D2) CST (ref:#5737) | 0.08–0.20 |
| H3K27ac | (polyclonal) Millipore (ref:07-360) | 0.05–0.30 |
| H3K36me3 | (polyclonal) Abcam (ref:ab9050) | 0.03–0.20 |
| H3K79me2 | (D15E8) CST (ref:#5427) | 0.20–0.40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourdareau, S.; Godfroy, O.; Gueno, J.; Scornet, D.; Coelho, S.M.; Tirichine, L.; Cock, J.M. An Efficient Chromatin Immunoprecipitation Protocol for the Analysis of Histone Modification Distributions in the Brown Alga Ectocarpus. Methods Protoc. 2022, 5, 36. https://doi.org/10.3390/mps5030036
Bourdareau S, Godfroy O, Gueno J, Scornet D, Coelho SM, Tirichine L, Cock JM. An Efficient Chromatin Immunoprecipitation Protocol for the Analysis of Histone Modification Distributions in the Brown Alga Ectocarpus. Methods and Protocols. 2022; 5(3):36. https://doi.org/10.3390/mps5030036
Chicago/Turabian StyleBourdareau, Simon, Olivier Godfroy, Josselin Gueno, Delphine Scornet, Susana M. Coelho, Leila Tirichine, and J. Mark Cock. 2022. "An Efficient Chromatin Immunoprecipitation Protocol for the Analysis of Histone Modification Distributions in the Brown Alga Ectocarpus" Methods and Protocols 5, no. 3: 36. https://doi.org/10.3390/mps5030036
APA StyleBourdareau, S., Godfroy, O., Gueno, J., Scornet, D., Coelho, S. M., Tirichine, L., & Cock, J. M. (2022). An Efficient Chromatin Immunoprecipitation Protocol for the Analysis of Histone Modification Distributions in the Brown Alga Ectocarpus. Methods and Protocols, 5(3), 36. https://doi.org/10.3390/mps5030036