
Retinoblastoma Cell Growth In Vitro and Tumor Formation In Ovo—Influence of Different Culture Conditions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
- (1)
- RB medium composed of Dulbecco’s modified Eagle’s medium (DMEM; PAN-Biotech; Aidenbach; Germany; Cat. No.: P04-04510) with 15% fetal bovine serum (FBS; PAN-Biotech; Aidenbach; Germany; P30-3702), 100 U penicillin/ml and 100 µg streptomycin/ml (Gibco; Grand Island, NEB, USA; Cat. No.: 15140), 4 mM L-glutamine (Gibco; Grand Island; USA; Cat. No.: 25030-024), 50 µM ß-mercaptoethanol (Roth; Germany; Karlsruhe; Cat. No.: M6250) and 10 µg insulin/ml (PAN-Biotech; Aidenbach; Germany; Cat. No.: P07-04300). Cultivation at 10% CO2.
- (2)
- DMEM medium composed of Dulbecco’s modified Eagle’s medium (DMEM; PAN-Biotech; Aidenbach; Germany; P04-04510) with 15% fetal bovine serum (FBS; PAN-Biotech; Aidenbach, Germany; P30-3702), 100 U penicillin/ml and 100 µg streptomycin/ml (Gibco; Grand Island; USA; Cat. No.: 15140) and 4 mM L-glutamine (Gibco; Grand Island; USA; Cat. No.: 25030-024). Cultivation at 5% CO2.
- (3)
- RPMI medium containing biotin, vitamin B12 and PABA (Gibco; Grand Island; USA; Cat. No.: 11875-093) with 15% fetal bovine serum (FBS; PAN-Biotech; Aidenbach, Germany; P30-3702), 100 U penicillin/ml and 100 µg streptomycin/ml (Gibco; Grand Island; USA; Cat. No.: 15140) and 4 mM L-glutamine (Gibco; Grand Island; USA; Cat. No.: 25030-024). Cultivation at 5% CO2.
2.2. Cell Proliferation and Apoptosis Detection
2.3. Growth Curves
2.4. Gene Expression Analysis
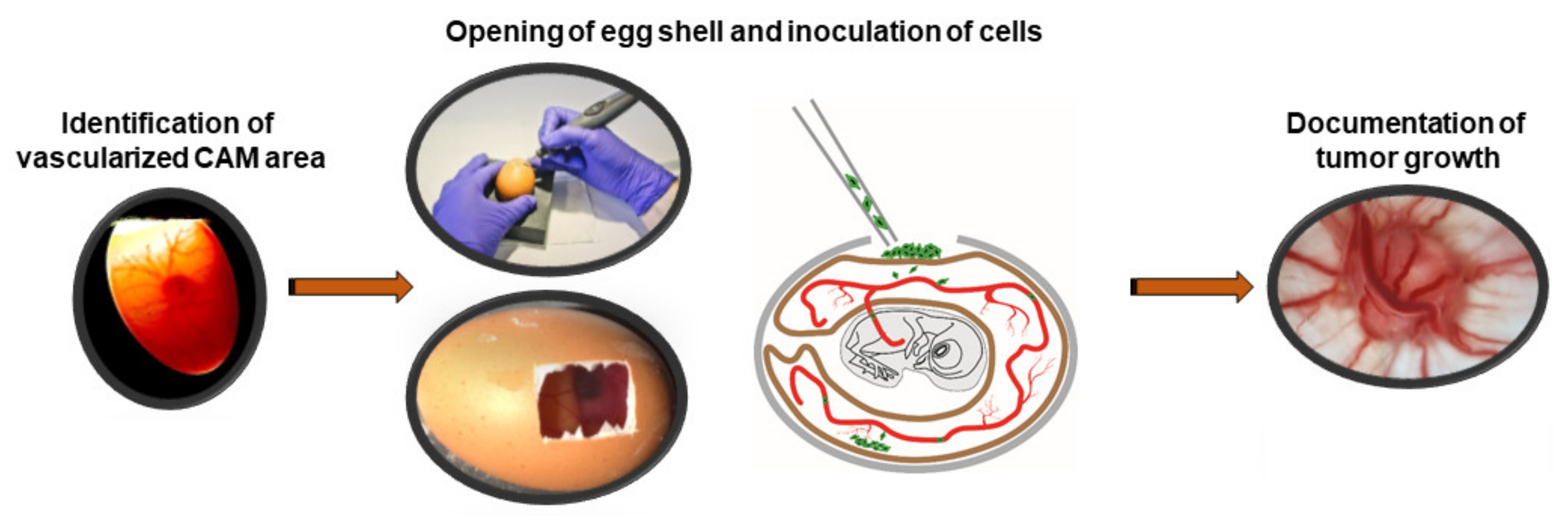
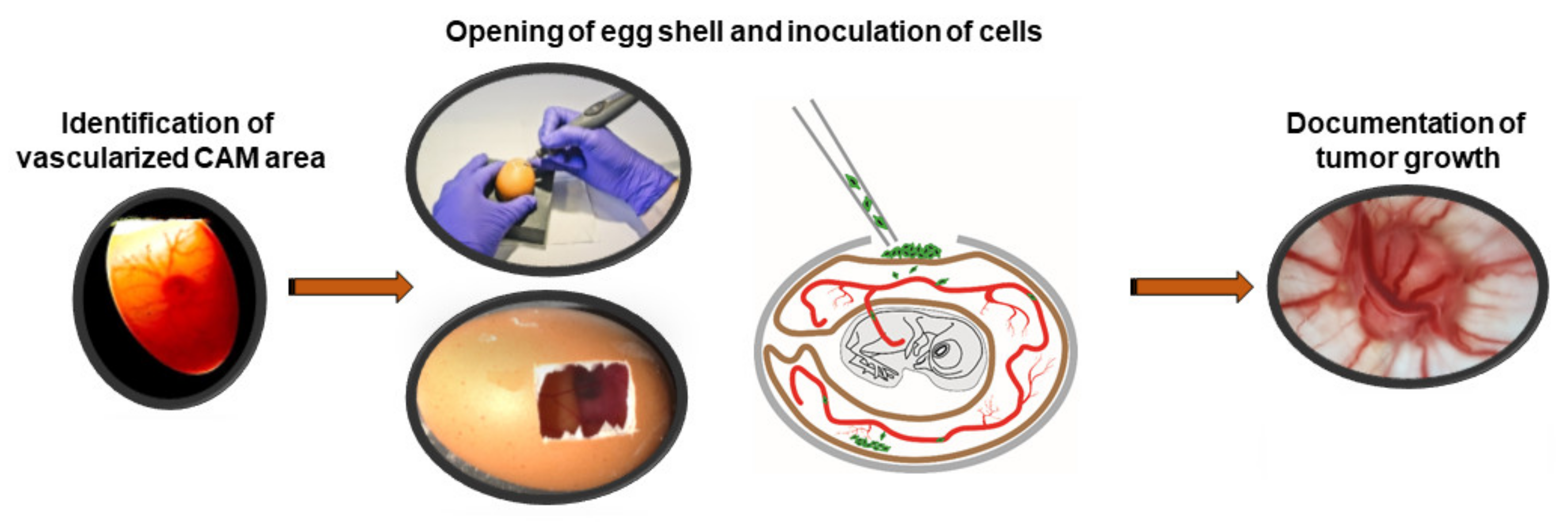
2.5. CAM Assays
2.6. Grafting of RB Cells
2.7. Harvesting of Tissue
2.8. Statistical Analysis
3. Results
3.1. RB Cell Line Morphology under Different Cell Culture Conditions
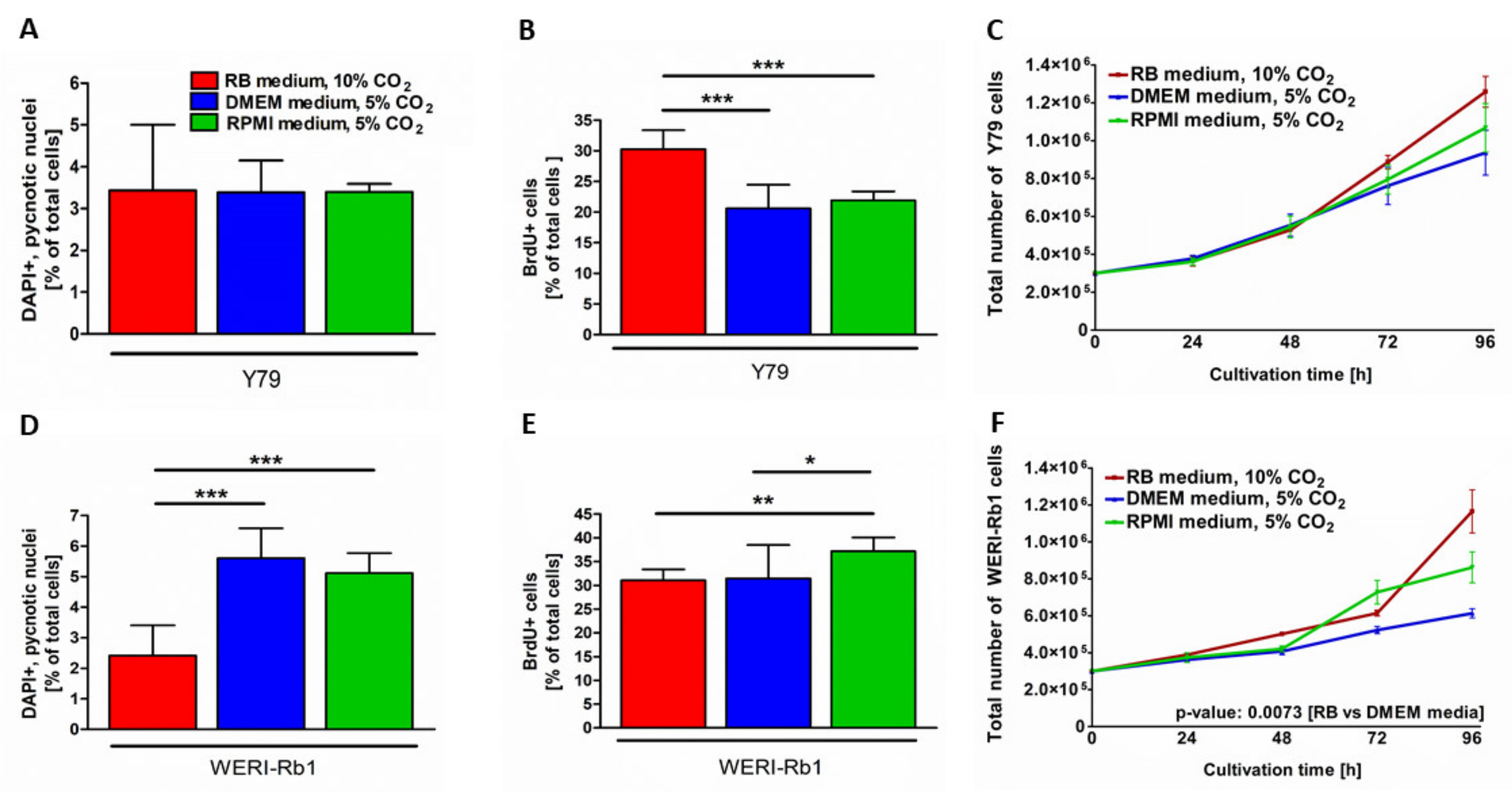
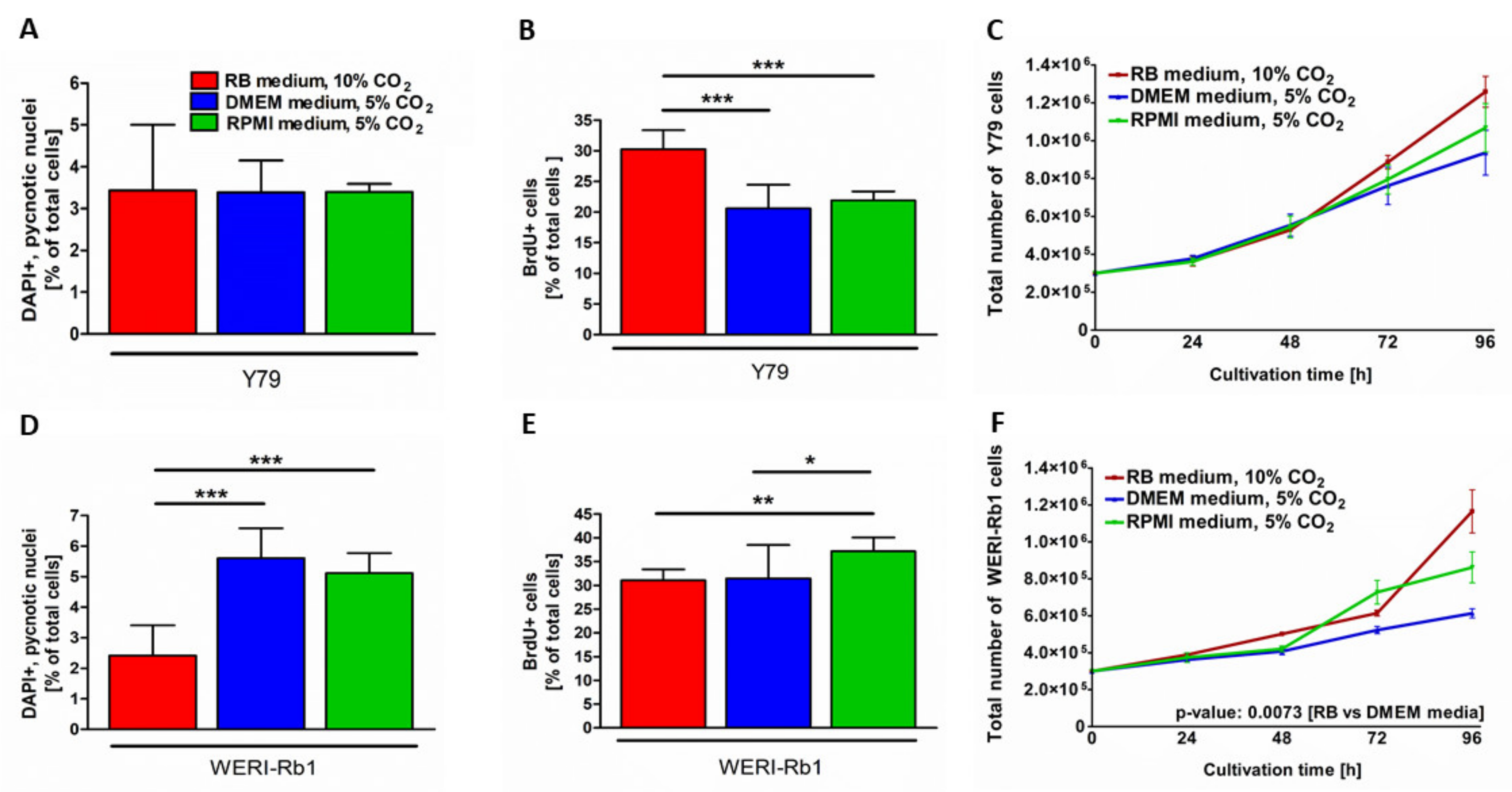
3.2. Different Culture Conditions Changed Apoptosis, Proliferation and Growth Kinetics of RB Cell Lines
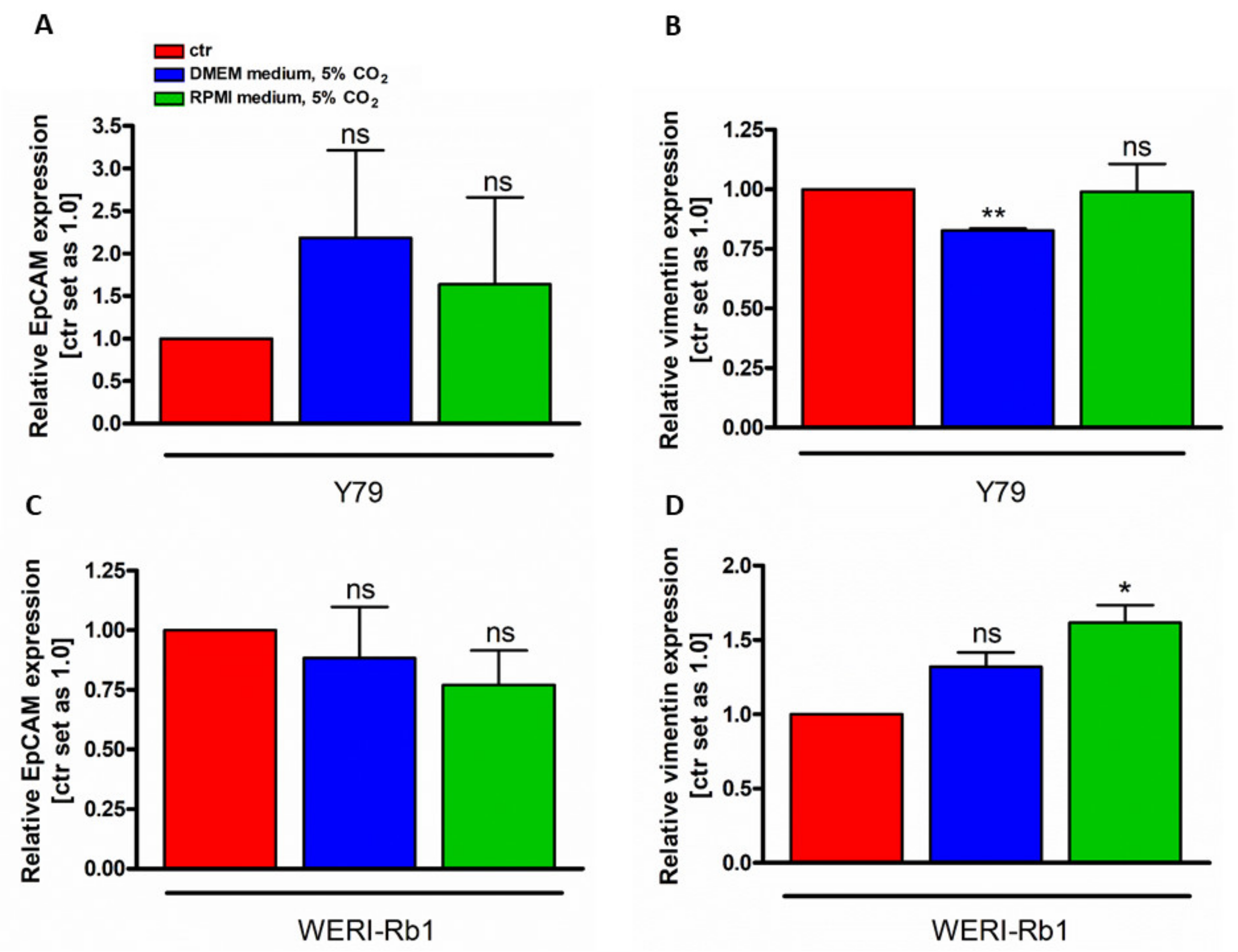
3.3. Vimentin and EpCAM Gene Expression Analysis under Different Cell Culture Conditions
3.4. Systematic Analysis of In Ovo RB Tumor Growth in the CAM Model
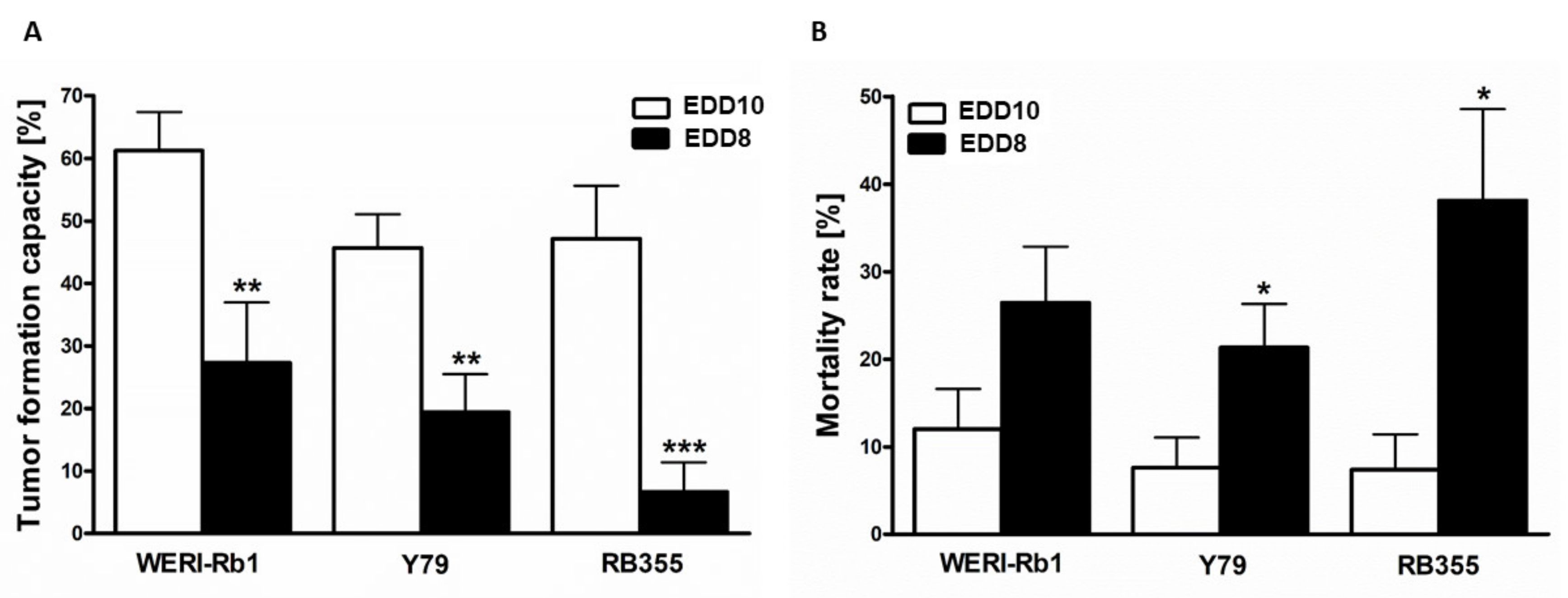
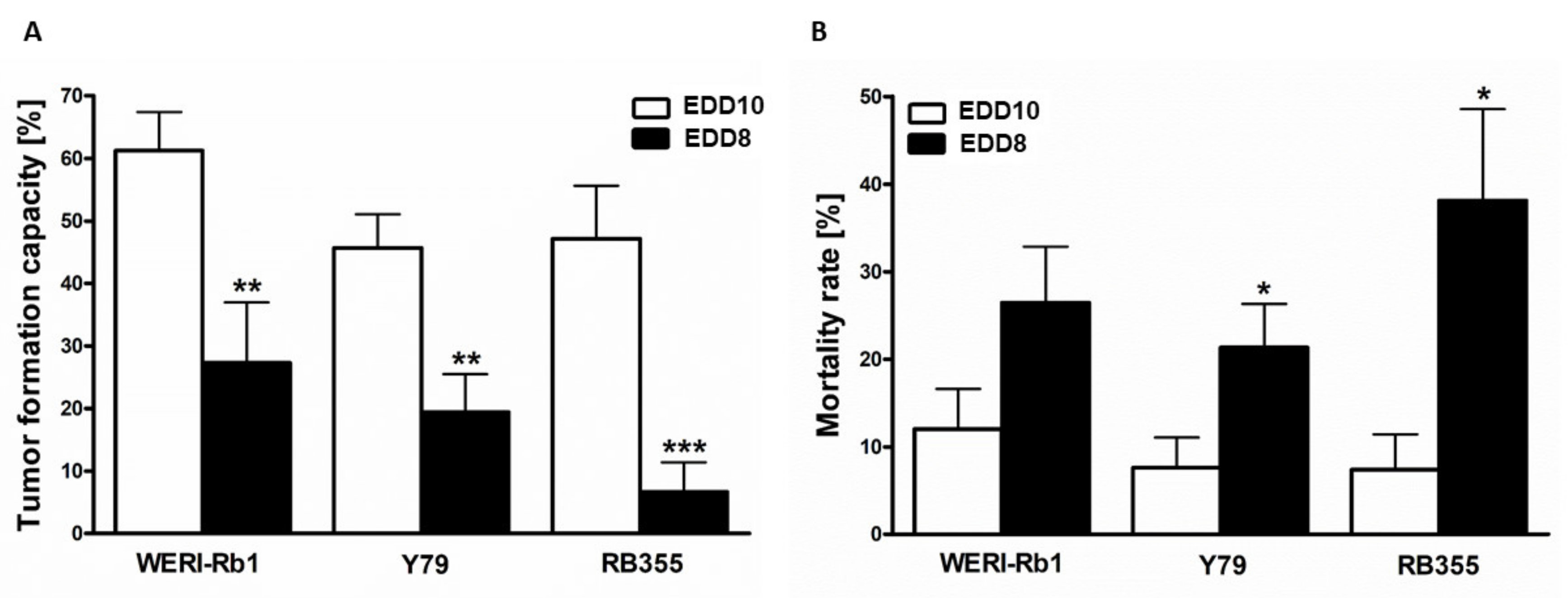
3.5. In Ovo RB Cell Tumor Development and Mortality Rate of Chick Embryos Depend on the Time Point of CAM Experiments
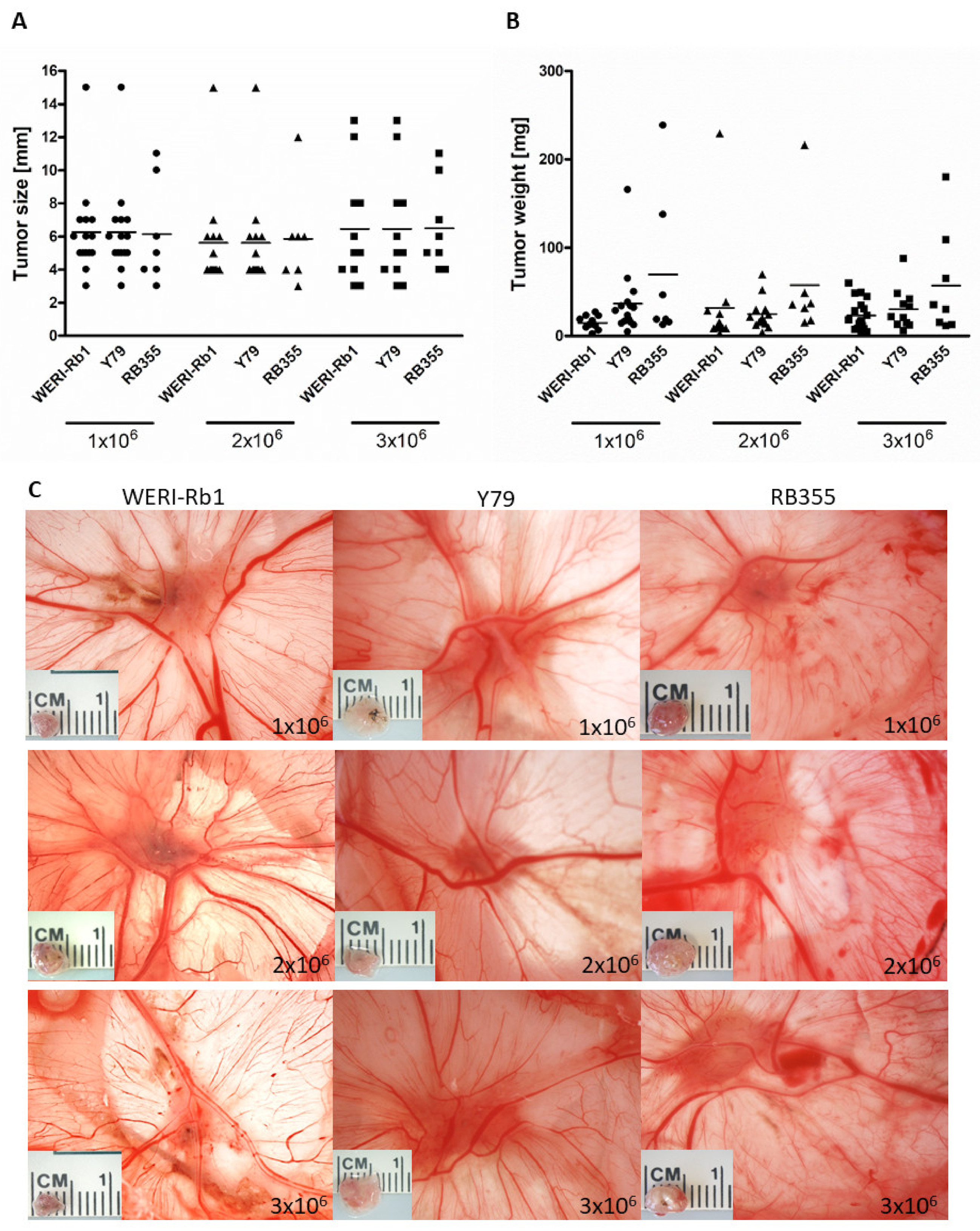
3.6. Inoculated RB Tumor Cell Concentration Did Not Influence Tumor Size and Weight In Ovo
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimaras, H.; Kimani, K.; Dimba, E.A.O.; Gronsdahl, P.; White, A.; Chan, H.S.L.; Gallie, B.L. Retinoblastoma. Lancet 2012, 379, 1436–1446. [Google Scholar] [CrossRef] [Green Version]
- Dimaras, H.; Corson, T.W. Retinoblastoma, the visible CNS tumor: A review. J. Neurosci. Res. 2018, 97, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Griegel, S.; Hong, C.; Frötschl, R.; Hülser, D.F.; Greger, V.; Horsthemke, B.; Rajewsky, M.F. Newly established human retinoblastoma cell lines exhibit an “immortalized” but not an invasive phenotype in vitro. Int. J. Cancer 1990, 46, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Temming, P.; Arendt, M.; Viehmann, A.; Eisele, L.; Le Guin, C.H.D.; Schündeln, M.M.; Biewald, E.; Astrahantseff, K.; Wieland, R.; Bornfeld, N.; et al. Incidence of second cancers after radiotherapy and systemic chemotherapy in heritable retinoblastoma survivors: A report from the German reference center. Pediatr. Blood Cancer 2017, 64, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Reid, T.W.; Albert, D.M.; Rabson, A.S.; Russell, P.; Craft, J.; Chu, E.W.; Tralka, T.S.; Wilcox, J.L. Characteristics of an established cell line of retinoblastoma. J. Nat. Cancer Inst. 1974, 53, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.; Philippeit, C.; Weise, A.; Dünker, N. Re-characterization of established human retinoblastoma cell lines. Histochem. Cell Biol. 2015, 143, 325–338. [Google Scholar] [CrossRef]
- Jayabal, P.; Ma, X.; Shiio, Y. EZH2 suppresses endogenous retroviruses and an interferon response in cancers. Genes Cancer 2021, 12, 96–105. [Google Scholar] [CrossRef]
- Gratias, S.; Schüler, A.; Hitpass, L.K.; Stephan, H.; Rieder, H.; Schneider, S.; Horsthemke, B.; Lohmann, D.R. Genomic gains on chromosome 1q in retinoblastoma: Consequences on gene expression and association with clinical manifestation. Int. J. Cancer 2005, 116, 555–563. [Google Scholar] [CrossRef]
- Deryugina, E.I.; Quigley, J.P. Chick embryo chorioallantoic membrane model systems to study and visualize human tumor cell metastasis. Histochem. Cell Biol. 2008, 130, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D. The CAM assay in the study of the metastatic process. Exp. Cell Res. 2021, 400, 112510. [Google Scholar] [CrossRef]
- Pawlikowska, P.; Tayoun, T.; Oulhen, M.; Faugeroux, V.; Rouffiac, V.; Aberlenc, A.; Pommier, A.L.; Honore, A.; Marty, V.; Bawa, O.; et al. Exploitation of the chick embryo chorioallantoic membrane (CAM) as a platform for anti-metastatic drug testing. Sci. Rep. 2020, 10, 16876. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Stock, R.; Ribatti, D. The CAM Assay as an Alternative In Vivo Model for Drug Testing. Handb. Exp. Pharmacol. 2021, 265, 303–323. [Google Scholar] [CrossRef] [PubMed]
- Eckrich, J.; Kugler, P.; Buhr, C.R.; Ernst, B.P.; Mendler, S.; Baumgart, J.; Brieger, J.; Wiesmann, N. Monitoring of tumor growth and vascularization with repetitive ultrasonography in the chicken chorioallantoic-membrane-assay. Sci. Rep. 2020, 10, 18585. [Google Scholar] [CrossRef] [PubMed]
- Kain, K.H.; Miller, J.W.I.; Jones-Paris, C.R.; Thomason, R.T.; Lewis, J.D.; Bader, D.M.; Barnett, J.V.; Zijlstra, A. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. 2014, 243, 216–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohle, D.S.; Pasa, S.D.; Gustmann, S.; Laub, M.; Wissler, J.H.; Jennissen, H.P.; Dünker, N. Chick ex ovo culture and ex ovo CAM assay: How it really works. J. Vis. Exp. 2009, 30, e1620. [Google Scholar] [CrossRef] [Green Version]
- Busch, M.; Grosse-Kreul, J.; Wirtz, J.J.; Beier, M.; Stephan, H.; Royer-Pokora, B.; Metz, K.; Dunker, N. Reduction of the tumorigenic potential of human retinoblastoma cell lines by TFF1 overexpression involves p53/caspase signaling and miR-18a regulation. Int. J. Cancer 2017, 141, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Dünker, N.; Jendrossek, V. Implementation of the Chick Chorioallantoic Membrane (CAM) Model in Radiation Biology and Experimental Radiation Oncology Research. Cancers 2019, 11, 1499. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Tamma, R.; Annese, T. Chorioallantoic membrane vascularization. A meta-analysis. Exp. Cell Res. 2021, 405, 112716. [Google Scholar] [CrossRef]
- Liu, M.; Scanlon, C.S.; Banerjee, R.; Russo, N.; Inglehart, R.C.; Willis, A.L.; Weiss, S.J.; D’Silva, N.J. The Histone Methyltransferase EZH2 Mediates Tumor Progression on the Chick Chorioallantoic Membrane Assay, a Novel Model of Head and Neck Squamous Cell Carcinoma. Transl. Oncol. 2013, 6, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D. The chick embryo chorioallantoic membrane as a model for tumor biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R. The chick embryo chorioallantoic membrane as an in vivo experimental model to study human neuroblastoma. J. Cell. Physiol. 2018, 234, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFall, R.C.; Sery, T.W.; Makadon, M. Characterization of a new continuous cell line derived from a human retinoblastoma. Cancer Res. 1977, 37, 1003–1010. [Google Scholar] [PubMed]
- Haubold, M.; Weise, A.; Stephan, H.; Dünker, N. Bone morphogenetic protein 4 (BMP4) signaling in retinoblastoma cells. Int. J. Biol. Sci. 2010, 6, 700–715. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, A.; Mellor, R.; Panzarella, G.; Aimes, R.T.; Hooper, J.D.; Marchenko, N.D.; Quigley, J.P. A quantitative analysis of rate-limiting steps in the metastatic cascade using human-specific real-time polymerase chain reaction. Cancer Res. 2002, 62, 7083–7092. [Google Scholar]
- Palmer, T.D.; Lewis, J.; Zijlstra, A. Quantitative analysis of cancer metastasis using an avian embryo model. J. Vis. Exp. 2011, 51, e2815. [Google Scholar] [CrossRef] [Green Version]
- Elso, C.M.; Roberts, L.J.; Smyth, G.K.; Thomson, R.J.; Baldwin, T.M.; Foote, S.J.; Handman, E. Leishmaniasis host response loci (lmr1-3) modify disease severity through a Th1/Th2-independent pathway. Genes Immun. 2004, 5, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-J.; Lei, K.-F.; Han, F. Tumor microenvironment: Recent advances in various cancer treatments. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3855–3864. [Google Scholar] [CrossRef]
- Kreft, M.E.; Tratnjek, L.; Lasič, E.; Hevir, N.; Rižner, T.L.; Kristan, K. Different Culture Conditions Affect Drug Transporter Gene Expression, Ultrastructure, and Permeability of Primary Human Nasal Epithelial Cells. Pharm. Res. 2020, 37, 170. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doege, A.; Steens, R.; Dünker, N.; Busch, M.A. Retinoblastoma Cell Growth In Vitro and Tumor Formation In Ovo—Influence of Different Culture Conditions. Methods Protoc. 2022, 5, 21. https://doi.org/10.3390/mps5020021
Doege A, Steens R, Dünker N, Busch MA. Retinoblastoma Cell Growth In Vitro and Tumor Formation In Ovo—Influence of Different Culture Conditions. Methods and Protocols. 2022; 5(2):21. https://doi.org/10.3390/mps5020021
Chicago/Turabian StyleDoege, Annika, Rebecca Steens, Nicole Dünker, and Maike Anna Busch. 2022. "Retinoblastoma Cell Growth In Vitro and Tumor Formation In Ovo—Influence of Different Culture Conditions" Methods and Protocols 5, no. 2: 21. https://doi.org/10.3390/mps5020021
APA StyleDoege, A., Steens, R., Dünker, N., & Busch, M. A. (2022). Retinoblastoma Cell Growth In Vitro and Tumor Formation In Ovo—Influence of Different Culture Conditions. Methods and Protocols, 5(2), 21. https://doi.org/10.3390/mps5020021