
Update on Chitin and Chitosan from Insects: Sources, Production, Characterization, and Biomedical Applications



Abstract

1. Introduction
2. Sources of Chitin and Chitosan from Insects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Species | Demineralization | Deproteinization | Decoloration | Chitin Yield (%) | Ref. |
|---|---|---|---|---|---|
| Lepidoptera | |||||
| Silkworm | 1 M HCl in 30 °C for 2 h | 1 M NaOH in 90 °C for 2 h | 2% KMnO4 for 2 h, 2% H2C2O4 for 2 h | NA | [21] |
| Flour moth, Ephestia kuehniella | 1 M HCl at 100 °C for 20 min | 1 M NaOH at 85 °C for 60 min | 1% KMnO4 for 60 min | 9.5–10.5 | [22] |
| Butterfly, Argynnis pandora | 2 M HCl at 50 °C for 24 h | 2 M NaOH solution at 50 °C for 24 h | Distilled water, methanol, and chloroform (4:2:1) for 10 min | Wings—22 Body without wings—8 | [23] |
| Clanis bilineata | 7% (v/v) HCl at 25 °C for 24 h | 10% (w/v) NaOH at 60 °C for 24 h | NA | NA | [24] |
| Clanis bilineata | 7% (v/v) HCl at 25 °C for 24 h | 10% (w/v) NaOH at 60 °C for 24 h | NA | NA | [25] |
| Clanis bilineata larvae | 7% (v/v) HCl at 25 °C for 24 h | 10% (w/v) NaOH at 60 °C for 24 h | NA | NA | [26] |
| Coleoptera | |||||
| Mealworm, Tenebrio molitor | 1 M HCl in 30 °C for 2 h | 1 M NaOH in 90 °C for 2 h | 2% KMnO4 for 2 h, 2% H2C2O4 for 2 h | NA | [21] |
| Comb-clawed beetles, Omophlus sp. | 2 M HCl for 4 h at 50 °C | 2 M NaOH for 20 h at 100 °C | Methanol, chloroform, and water (2:1:4) | NA | [27] |
| Cockchafer, Melolontha melolontha | 50 mL of 4 M HCl at 75 °C for 2 h | 4 M NaOH at 150 °C for 18 h | Water, alcohol, and chloroform (4:2:1) for 20 min | 13–14 | [28] |
| Cockchafer, Melolontha sp. | 2 M HCl at 60 °C for 20 h | 1 M of NaOH for 20 h at 100 °C | Distilled water, methanol, and chloroform (4:2:1) for 30 min | Male—16.60 Female—15.66 | [29] |
| Colorado potato beetle, Leptinotarsa decemlineata | 100 mL of 2 M HCl at 65–75 °C for 2 h | 50 mL of 2 M NaOH at 80–90 °C for 16 h | Chloroform, methanol, and water (in a ratio of 1:2:4) for 1 h | Adults—20 Larvae—7 | [30] |
| Catharsius molossus L. | 1.30 M HCl at 80 °C for 30 min | 4.0 M NaOH at 90 °C for 6 h | 2% oxalic acid at 70 °C for 30 min | 24% | [31] |
| Calosoma rugosa | 1 M HCl | 1.0 M NaOH at 100 °C for 8 h | NA | 5.0 | [32] |
| Calosoma rugosa | * 36.5% HCl | * 1.0 M NaOH | NA | NA | [33] |
| Mealworm, Tenebrio molitor | 3 h in 2 M HCl at 20 °C | 500 mL 5% NaOH at 95 °C for 3 h | NA | 18.01 | [34] |
| Mealworm, Tenebrio molitor | 2 M HCl at 50 °C for 24 h | 2 M NaOH solution at 50 °C for 24 h | NA | NA | [35] |
| Mealworm, Tenebrio molitor | * 1.5 M HCl at 20 °C, 120 rpm for 6 h | * 1.25 M NaOH at 80 °C for 24 h | NA | 4.72% | [36] |
| Tenebrio Molitor | 2 M HCl at 65–75 °C for 2 h | 2 M NaOH at 80 to 90 °C | Chloroform, methanol, and water (1:2:4) for 1 h | 17.7% | [37] |
| Mealworm beetle, Tenebrio molitor, Zophobas morio | * 7% (v/v) HCl at 25 °C for 24 h | * NaOH at 80 °C for 24 h | NA | Larvae—4.60 Adult—8.40 Superworm—3.90 | [38] |
| Rhinoceros beetle, Allomyrina dichotoma | Larvae—10.53 Pupa—12.70 Adult—14.20 | ||||
| Mealworm, Zophobas morio | 1.0 M HCl at 35 °C | 0.5 M, 1.0 M and 2.0 M NaOH at 80 °C for 20 h | Glacial acetone for 30 min | 0.5 M-5.43 1.0 M-5.22 2.0 M-4.77 | [39] |
| Mealworm, Zophobas morio | 20% HCI for 10 min | 1 M NaOH 1:10 (g/mL) at 80 °C for 3 h | NA | NA | [40] |
| European stag beetle, Lucanus cervus | 1 M HCl at 90 °C for 1 h | 1 M NaOH in 90 °C for 14 h | Chloroform, methanol, and water (1:2:4, v/v) | 10.9 | [41] |
| Pine chafer, Polyphylla fullo | 11.3 | ||||
| Pentodon algerinum | 5% acetic acid at 55 °C for 2 h | 10% KOH at 40 °C for 48 h | NA | NA | [42] |
| Wheat weevil, Sitophilus granarius | 1 M HCl for 0.5 h | 1 M NaOH at 100 °C, 8 h | Ethanol and acetone | NA | [43] |
| Dor beetle, Anoplotrupes stercorosus | 2 M of HCl at 100 °C for 2 h | 2 M NaOH at 140 °C for 20 h | Chloroform, methanol, and water (1:2:4, v/v) for 2 h at room temperature | 20.1 | [44] |
| Blaps tibialis | 25.2 | ||||
| Rose chafer, Cetonia aurata | 18.2 | ||||
| Dor beetle, Geotrupes stercorarius | 20.4 | ||||
| Blaps lethifera | 1 M HCl for 1 h at 40 °C | 1 M NaOH at 80 °C for 2 h | 10 v/v % H2O2 for 30 min at 50 °C | NA | [45] |
| Pimelia fernandezlopezi | |||||
| Banana weevil, Cosmopolites sordidus | 1.0 M HCl at 50 °C for 24 h | 1.0 M NaOH 80 °C for 8 h | NA | 11.8 | [46] |
| Orthoptera | |||||
| Grasshopper | 1 M HCl in 30 °C for 2 h | 1 M NaOH in 90 °C for 2 h | 2% KMnO4 for 2 h, 2% H2C2O4 for 2 h | NA | [21] |
| Shistocerca gregarea Forsskal | 5% acetic acid at 55 °C for 2 h | 10% KOH at 40 °C for 48 h | NA | NA | [42] |
| Mexican katydid, Pterophylla beltrani | NA | NA | NA | 11.8 | [47] |
| Moroccan locust, Dociostaurus maroccanus | 2 M HCl in 55 °C for 1 h | 2 M NaOH in 50 °C for 18 h | Methanol, chloroform, and distilled water (2:1:4) | Nymphs—12 Adults—14 | [48] |
| House cricket, Brachytrupes | * Oxalic acid for 3 h at room temperature | * 1 M NaOH at 95 °C for 6 h | 1% sodium hypochlorite for 3 h | 4.3–7.1 | [49] |
| Celes variabilis, | 4 M HCl at 75 °C for 2 h | 4 M NaOH for 20 h at 150 °C | NA | 4.71–11.84 | [50] |
| Wart-biter, Decticus verrucivorus, | |||||
| Desert cricket, Melanogryllus desertus, | |||||
| Paracyptera labiata | |||||
| Calliptamus barbarus Oedaleus decorus | 1 M HCl at 100 °C for 30 min | 1 M NaOH at 80–90 °C for 21 h | Chloroform, methanol, and distilled water solution (1:2:4) for 1 h | 20.5 16.5 | [51] |
| Ailopus simulatrix | 4 M HCl at 75 °C for 1 h | 2 M NaOH at 175 °C for 18 h | Chloroform, methanol, and distilled water (1:2:4) | 5.3 | [52] |
| Ailopus strepens | 7.4 | ||||
| Duroniella fracta | 5.7 | ||||
| Duroniella laticornis | 6.5 | ||||
| Red-winged grasshopper, Oedipoda miniata | 8.1 | ||||
| Blue-winged grasshopper, Oedipoda caerulescens | 8.9 | ||||
| Pyrgomorpha cognata | 6.6 | ||||
| Two-spotted cricket, Gryllus bimaculatus | 2 M HCl | 1.25 M NaOH | NA | 20.9–23.3 | [53] |
| Calosoma rugosa | 1 M HCl | 1.0 M NaOH at 100 °C for 8 h | NA | 12.2 | [32] |
| Bradyporus sureyai | 1 M HCl in 90 °C for 1 h | 1 M NaOH in 90 °C for 14 h | Chloroform, methanol, and water (1:2:4, v/v) | 9.8 | [41] |
| European mole cricket, Gryllotalpa gryllotalpa | 10.1 | ||||
| Two-spotted cricket, Gryllus bimaculatus | * Oxalic acid for 3 h at room temperature | * 1 M NaOH at 95 °C and 130 rpm for 6 h | APS solution (50% (w/v)) at 50 °C for 30 min | 5.1 | [54] |
| Two-spotted cricket, Gryllus bimaculatus | 2 M HCl at 21 °C for 3 h | 1.25 M NaOH at 95 °C for 3 h | 50% NaOH (w/w) at 95 °C and 105 °C for 3 h | 79.03–91.14 | [55] |
| Brachystola magna | 2 M HCl at 50 °C for 24 h | 2 M NaOH solution at 50 °C for 24 h | NA | NA | [35] |
| House cricket, Acheta domesticus | * 1 M HCl for 2 h at 98 °C | * 1 M NaOH at 80 °C for 24 h | NA | NA | [56] |
| House cricket, Acheta domesticus Gryllodes sigillatus | 0.25 M HCl at 85 °C for 15 min | 1 L NaOH at 70 °C for 22 h | NA | NA | [57] |
| House cricket, Acheta domesticus | * HCl, 1 M for 2 h at 98 °C | * NaOH, 1 M at 80 °C for 22 h | NA | 7.34 | [58] |
| House cricket, Acheta domesticus | 1 M NaOH (1:2 w/v) at 70 °C for 22 h | 0.25 M HCl (1:2 w/v) at 85 °C for 15 min | NA | 5.7 ± 0.10 | [59] |
| Gryllodes sigillatus | 3.4 ± 0.10 | ||||
| Hymenoptera | |||||
| Western honey bee, Apsis mellifera | 2 M HCl at 80 °C for 6 h | 2 M of NaOH and refluxed for 20 h at 100 °C | Distilled water (40 mL), methanol (20 mL), and chloroform (20 mL) | Head—8.9 Thorax—6.79 Abdomen—8.61 Legs—13.25 Wings—7.64 | [60] |
| Western honey bee, Apsis mellifera | 1 N HCl | 1 M NaOH for 12 h at ambient temperature (20 °C) | NA | 8.8 | [61] |
| Western honey bee, Apsis mellifera | 1 M HCl | 1.0 M NaOH at 100 °C for 8 h | NA | 2.5 | [32] |
| Western honey bee, Apsis mellifera | 36.5% HCl | 1.0 M NaOH | NA | NA | [33] |
| Western honey bee, Apis mellifera | * 6.7% HCl at 25 °C for 3 h | * 8% NaOH at 90 °C for 1 h | 33% H2O2 | 23 | [62] |
| European hornet, Vespa crabro Oriental hornet, Vespa orientalis German wasp, Vespula germanica | 2 M HCl at 75 °C for 2 h | 4 M NaOH at 150 °C for 18 h | Distilled water, methanol, and chloroform (4:2:1) for 2 h | 8.3 6.4 11.9 | [63] |
| Asian hornet, Vespa velutina | 100 mL of 1 M HCl at 50 °C for 3 h | 1 M NaOH (100 mL) at 60 °C for 8 h | 100 mL 1% sodium hypochlorite | 11.7 | [64] |
| Oriental hornet, Vespa orientalis | 1.0 M HCl to a solid ratio of 15 mL/g at 100 °C for 20 min | 1.0 M sodium hydroxide at 85 °C | H2O2/33% HCl 9:1, v/v | NA | [65] |
| European hornet, Vespa crabro | 1 M HCl at 50 °C for 6 h | 60 °C in 1 M NaOH solution for 16 h | Distilled water, methanol, and chloroform (4:2:1) solution at room temperature for 40 min at 250 rpm | Larvae—2.2 Pupa—6.2 Adult—10.3 | [66] |
| Oriental hornet, Vespa orientalis | 5% acetic acid at 55 °C for 2 h | 10% KOH at 40 °C for 48 h | NA | NA | [42] |
| Red-tailed bumblebee, Bombus lapidaries Formica clara | 2 M of HCl at 100 °C for 2 h | 2 M NaOH at 140 °C for 20 h | Chloroform, methanol, and water (1:2:4, v/v) for 2 h at room temperature | 9.3 7.8 | [44] |
| Diptera | |||||
| Black soldier fly, Hermetia illucens | 1:10 (m/v) with HCl 1 M at room temperature for 1 h | 1 M NaOH treatment (solid: liquid ratio of 1:25 (m/v), 1 h at 80 °C | NA | Larvae—96.3 ± 3.7 Prepupae—94.5 ± 1.5 Pupae—93.9 ± 2.0 Shedding—75.7 ± 4.0 Cocoons—96.8 ± 1.8 Flies—95.7 | [67] |
| Black soldier fly, Hermetia illucens | 1 M HCl for 1 h | 1 M NaOH at 80 °C for 24 h | 1% KMnO4 | NA | [68] |
| Black soldier fly, Hermetia illucens | 1 M HCl at 100 °C for 30 min | 1 M NaOH at 80 °C for 24 h | NA | Pupae exuviae—9 Imago—23 | [69] |
| Black soldier fly, Hermetia illucens | NA | 1 M NaOH 1 h at 80 °C | NA | 8.5 ± 0.1 | [70] |
| Black soldier fly, Hermetia illucens | 2 M HCl at 55 °C for 1 h | 2 M NaOH at 50 °C for 18 h | NaClO at 80 °C for 4 h | Larvae—3.6 Prepupa—3.1 Puparium—14.1 Adults—2.9 | [71] |
| Black soldier fly, Hermetia illucens | HCl at 2 h | NaOH at 90 °C for 3 h | NA | 21.3 | [72] |
| Black soldier fly, Hermetia illucens | 2% HCl for 2 h at 20 °C | NaOH 50 °C for 2 h | NA | 7 | [73] |
| Black soldier fly, Hermetia illucens | * 2 N HCl for 24 h at 15 min | * 40 mL of 2 N HCl for 24 h at room temperature | NA | 9 | [74] |
| Black soldier fly, Hermetia illucens | 1 M HCl, 1:10 (w/v) for 2 h | 1 M NaOH, 1 g/10 mL, at 80 °C for 6 h | 1% KMnO4 in a 1:30 w/v ratio at room temperature for 4 h | Late larvae—3.025 Prepupae—5.371 Pupal exuviae—18.800 Imagoes—11.846 | [75] |
| Black soldier fly, Hermetia illucens | * 1 M HCl for 2 h | * 1 M NaOH 4 h | NA | 10.18 ± 0.42 | [76] |
| Black soldier fly, Hermetia illucens | 0.5 M CH2O2 for 1h at room temperature | 2 M NaOH for 2 h at 80 °C | 5% H2O2 for 1 h at 90 °C | NA | [77] |
| Black soldier fly, Hermetia illucens | 1 M HCl for 2 h | 1 M NaOH 4 h | NA | NA | [78] |
| Black soldier fly, Hermetia illucens | 1 M HCl for 2 h at 100 °C | 1 M NaOH for 4 h at 100 °C | NA | 10.18 | [79] |
| Black soldier fly, Hermetia illucens | 0.5 M formic acid for 1 h at room temperature | 2 M NaOH, 2 h at 80 °C | 5% (v/v) H2O2 for 30–60 min at 90 °C | Larvae—10 ± 0.7 Pupal exuviae—23 ± 1.9 Dead adults—6 ± 0.1 | [80] |
| Black soldier fly, Hermetia illucens | 0.5 M HCl at room temperature for 2 h | 1.9 M NaOH for 2 h at 50 °C | 5% H2O2 | NA | [81] |
| Black soldier fly, Hermetia illucens | 7% HCl for 2 h at room temperature | 10% NaOH at 80 °C for 24 h | NA | NA | [82] |
| Black soldier fly, Hermetia illucens | 1% HCl at 20 °C for 2 h | 30% (w/w) NaOH at room temperature for 30 min, and then at 100 °C for 2 h | NA | NA | [83] |
| Black soldier fly, Hermetia illucens | 1 M HCl at 22 °C for 1 h | 1 M NaOH at 80 °C for 24 h | 1—Without decoloration 2—Water at 100 °C for 24 h 3—9% H2O2 at 80 °C for 2.5 h 4—9% H2O2 at 80 °C for 5 h 5—1% KMnO4 at 80 °C for 20 min | 1—7.95 ± 0.20 2—7.97 ± 0.10 3—7.01 ± 0.12 4—5.98 ± 0.08 5—5.69 ± 0.28 | [84] |
| Common fruit fly Drosophila melanogaster | 2 M HCl solution for 3 h at 4 °C | NaOH (8% w/w) solution for 20 h at 70 °C | Methanol, chloroform, and distilled water (in a ratio of 2:1:4) for 30 min | 7.85 | [85] |
| Calliphora vicina | 2 M of HCl at 100 °C for 2 h | 2 M NaOH at 140 °C for 20 h | Chloroform, methanol, and water (1:2:4, v/v) for 2 h at room temperature | 8.1 | [44] |
| Housefly, Musca domestica | 1 M HCl for 1 h at 40 °C | 2 h of 1 M NaOH at 80 °C | 10 v/v % H2O2 for 30 at 50 °C | NA | [45] |
| Housefly, Musca domestica | 3 h in 500 mL of 2 N HCl solution at room temperature | 500 mL of 1.25 N NaOH at 95 °C for 3 h | NA | 8.02 | [86] |
| Tabanus bovinus | 1 M HCl for 12 h at room temperature | 1 M NaOH for 18 h at 70 °C | Water, methanol, and chloroform (1:2:4) | NA | [87] |
| Hemiptera | |||||
| Green bug, Nezara viridula | 5% acetic acid at 55 °C for 2 h | 10% KOH at 40 °C for 48 h | NA | NA | [42] |
| Dock bug, Coreus marginatus | 2 M of HCl at 100 °C for 2 h | 2 M NaOH at 140 °C for 20 h | Chloroform, methanol, and water (1:2:4, v/v) for 2 h at room temperature | 14.5 | [44] |
| Black-and-red bug, Lygaeus equestris | 11.1 | ||||
| Pyrrhocoris apterus | 10.6 | ||||
| Cicada slough | 1 M HCl in 30 °C for 2 h | 1 M NaOH in 90 °C for 2 h | 2% KMnO4 for 2 h, 2% H2C2O4 for 2 h | NA | [21] |
| Aquatic bug, Ranatra linearis | 100 mL of 1 M HCl at 90 °C for 1 h | 1 M NaOH at 110 °C for 18 h | Chloroform, methanol, and water (1:2:4) | 15–16 | [88] |
| Cicada lodosi | 2 M HCl for 2 h at 100 °C | 2 M NaOH at 100 °C for 20 h | Water, methanol, and chloroform mixed at a ratio of 4:2:1 | 4.97 | [89] |
| Cicada mordoganensis | 6.49 | ||||
| Cicadatra platyptera | 8.84 | ||||
| Cicadatra atra | 6.70 | ||||
| Cicadatra hyaline | 5.51 | ||||
| Cicadivetta tibialis | 5.88 | ||||
| Cicada Cryptotympana atrata | 1000 mL of 7% (w/w) HCl at room temperature for 24 h | 1000 mL of 10% (w/w) NaOH at 60 °C for 24 h | NA | 62.42 | [24] |
| Coridius nepalensis | 1 M HCl for 1 h | 1 M NaOH at 80 °C for 24 h | 1% sodium hypochlorite for 1 h | 43.97 | [90] |
| Dictyoptera | |||||
| Eupolyphaga sinensis | 1.3 M HCl at 80 °C for 1 h, soaked at room temperature for 24 h | 4 M NaOH at 90 °C for 6 h | 10% H2O2 at 80 °C for 30 min | 11.63 ± 0.80 | [91] |
| Brazilian cockroach, Blaberus giganteus | NA | 2 M NaOH at 90 °C for 9 h | Chloroform, methanol, and water (1:2:2) at room temperature for 1.5 h | Wings—26.9 Dorsal pronotum—21.2 | [92] |
| German cockroach, Blattela germanic | 5% acetic acid at 55 °C for 2 h | 10% KOH at 40 °C for 48 h | NA | NA | [42] |
| German cockroach, Blattella germanica | 2 M of HCl at 100 °C for 2 h | 2 M NaOH at 140 °C for 20 h | Chloroform, methanol, and water (1:2:4, v/v) for 2 h at room temperature | 4.7 | [44] |
| American cockroach, Periplaneta americana | 1% sodium hypochlorite solution (1%, w/v) | 1 M NaOH at 100 °C for 24 h | NA | Nymph—8.4 Adult—15 | [93] |
| German cockroach, Blattella germanica | Nymph—5.4 Adult—6.2 | ||||
| American cockroach, Periplaneta americana | 2 N HCl at room temperature for 3 h | 1.25 N NaOH at 95 °C for 3 h | NA | 3.36 | [53] |
| American cockroach, Periplaneta americana | * 20 mL of 1% HCl for 24 h | * 4% of NaOH for 1 h | 50 mL of 2% NaOH for 1 h | NA | [94] |
| American cockroach, Periplaneta americana | 4 M HCl for 2 h at 75 °C | 4 M NaOH for 20 h at 150 °C | Water, methanol, and chloroform (ratio of 4:2:1) for 4 h at 30 °C | Wings—18 Without wings—13 | [95] |
| American cockroach, Periplaneta americana | * 0.5 M HCl at 60 °C and 500 rpm for 1 h | * 0.5 M NaOH at 95 °C and 500 rpm for 20 min, then 4 M NaOH for 160 min | 10% H2O2 at 80 °C and 500 rpm for 3 h | NA | [96] |
| American cockroach, Pariplaneta americana linnaeus | 6.7% HCl at 25 °C for 3 h | 8% NaOH at 90 °C for 1 h | 33% H2O2 | 42 | [62] |
| American cockroach, Periplaneta americana | * 1 M HCl for 2 h at 75 °C | * 2.5% (w/v) NaOH for 6 h at 100 °C | Acetone at 50 °C for 2 h | NA | [97] |
| American cockroach, Periplaneta americana | 5% acetic acid at 55 °C for 2 h | 10% KOH at 40 °C for 48 h | NA | NA | [42] |
| American cockroach, Periplaneta americana | NA | 4% NaOH for 48 h at 90 °C | NA | NA | [98] |
| Odonata | |||||
| Dragonfly, Sympetrum fonscolombii | 1 M HCl at room temperature for 1 h | 1 M NaOH solution at 50 °C for 15 h | Chloroform, methanol, and distilled water (1:2:4, v/v) | 20.3 ± 0.85 | [99] |
| Downy emerald, Cordulia aenea | 2 M of HCl at 100 °C for 2 h | 2 M NaOH at 140 °C for 20 h | Chloroform, methanol, and water (1:2:4, v/v) for 2 h at room temperature | 9.5 | [44] |
| Four-spotted chaser, Libellula quadrimaculata | 10.1 | ||||
| Ephemeroptera | |||||
| Mayfly | 2 M HCl at 50 °C | 2 M NaOH at 100 °C | Methanol and chloroform (1:1) | 10.21 | [100] |
| Insect Species | Deacetylation Conditions | Chitosan Yield (%) | DDA (%) | Molecular Weight (Da) | Moisture Content (%) | Ash Content (%) | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|
| Chitin | Chitosan | Chitin | Chitosan | ||||||
| Lepidoptera | |||||||||
| Silkworm | 60% NaOH in 100 °C for 8 h | 3.1 | 85.5 | (4.090 ± 0.059) × 104 | NA | 0.07 ± 0.008 | NA | 0.05 ± 0.003 | [20] |
| Mediterranean flour moth, Ephestia kuehniella | NA | NA | NA | NA | 9.1 ± 0.4 | NA | 0.14 ± 0.08 | NA | [21] |
| Clanis bilineata | 55% NaOH (w/w), 120 °C for 4 h | 95.9 | NA | NA | NA | 3.8 | NA | 0.3 | [24] |
| Coleoptera | |||||||||
| Mealworm, Tenebrio molitor | 60% NaOH in 100 °C for 8 h | 2.5 | 85.9 | (3.975 ± 0.072) × 104 | NA | 0.19 ± 0.012 | NA | 0.50 ± 0.016 | [20] |
| Mealworm, Tenebrio molitor | 500 mL of NaOH at 95 or 105 °C for 3 h or 5 h | 9.2 | 95.5 | NA | [33] | ||||
| Mealworm, Tenebrio molitor | 50% NaOH at 100 °C for 3h | 78.26 | 75.84 | NA | [36] | ||||
| Mealworm, Tenebrio molitor | 60% NaOH contained NaBH4 (0.004 g), 120 °C for 2 h | NA | 88.55 | 8.123×105 | NA | [33] | |||
| Mealworm, Tenebrio molitor | NaOH 40% (w/v) at 90 °C, 500 rpm for 8 h | 31.9 | 53.9 | NA | 6.2 ± 0.5 | 4.2 ± 0.1 | 3.6 ± 0.2 | 3.7 ± 0.1 | [101] |
| Mealworm, Tenebrio molitor | 50% NaOH at 80 °C for 4 h | NA | 89.4 | NA | [35] | ||||
| Mealworm beetle, Tenebrio molitor, Zophobas morio | 55% (w/v) NaOH at 90 °C for 9 h | Larvae—80.00 Adult—78.33 Superworm—83.33 | Larvae—75.59 Adult—75.63 Superworm—75.67 | NA | [37] | ||||
| Rhinoceros beetle, Allomyrina dichotoma | Larvae—83.37 Pupa—83.37 Adult—75.00 | Larvae—75.66 Pupa—75.67 Adult—74.66 | |||||||
| Mealworm beetle, Zophobas morio | 50 wt % NaOH in 90 °C for 30 h | 65.84, 70.88, 75.52 | 81.06, 64.82, 74.14 | NA | [39] | ||||
| Mealworm beetle, Zophobas morio | NaOH 100 g/mL at 80 °C for 16 h | 15–18% | NA | NA | [40] | ||||
| Catharsius molossus | 8 M NaOH at room temperature for 24 h | 17% | 94.9 ± 0.85 | 4.5 ± 0.07 ×105 | NA | 6.55 ± 0.05 | NA | 0.34 ± 0.04 | [31] |
| Calosoma rugosa | 50% NaOH (15 mL/g) at 100 °C for 8 h | NA | 95 | NA | NA | 8.8 | NA | 2.0 | [32] |
| Calosoma rugosa | 50% NaOH at 100 °C for 8 h | NA | 95 | NA | [33] | ||||
| Colorado potato beetle, Leptinotarsa decemlineata | 50% NaOH (w/v, 1:20) at 100 °C for 3 h | Adult—72 Larvae—67 | Adult—82 Larvae—76 | Adults—2.722 × 103 Larvae—2.676 × 103 | NA | [102] | |||
| European stag beetle, Lucanus cervus, | NA | NA | NA | NA | 6.6 | NA | 0.6 | NA | [41] |
| Pine chafer, Polyphylla fullo | 5.9 | 1.7 | |||||||
| Blaps lethifera, | 50 w/v% NaOH | 50.0 ± 0.3 | 87.1 ± 0.2 | NA | NA | 14.3 ± 0.3 | NA | 1.5 ± 0.1 | [45] |
| Pimelia fernandezlopezi | 41.7 ± 0.5 | 88.2 ± 0.1 | 17.2 ± 0.2 | 2.0 ± 0.1 | |||||
| Banana weevil, Cosmopolites sordidus | 50% NaOH at 90 °C for 10 h | 70.2 | 77.8 ± 0.39 | (343 ± 37.3) × 103 | NA | 2.4 | 6.4 | 2.2 | [46] |
| Orthoptera | |||||||||
| Grasshopper | 60% NaOH in 100 °C for 8 h | 5.7 | 89.7 | (3.989 ± 0.021) × 104 | NA | 1.8 ± 0.213 | NA | 0.89 ± 0.025 | [21] |
| Mexican katydid, Pterophylla beltrani | 70% NaOH in 1:3 ratio for 1.5 h at 120 °C | 58.8 | NA | NA | NA | NA | NA | NA | [46] |
| Moroccan locust, Dociostaurus maroccanus | 60% NaOH at 150 °C for 4 h | Nymphs—77.38 Adults—81.69 | NA | Adults—7.2 × 103 Nymphs—5.6 × 103 | NA | NA | NA | NA | [48] |
| Brachytrupes | 50% (w/v) NaOH at 121 °C for 5 h | 2.4–5.8 | NA | NA | 4.00 | 3.33 | 1.00 | 1.00 | [49] |
| Calliptamus barbarus, Oedaleus decorus | 50% NaOH (w/v 1:15) at 130 °C for 2 h | 70–75 74–76 | 70–75 | NA | NA | NA | NA | NA | [51] |
| Desert locust, Schistocerca gregaria | 50% NaOH (15 mL/g) at 100 °C for 8 h | NA | 98 | NA | NA | NA | 14.1 | 1.6 | [32] |
| Bradyporus sureyai, | NA | NA | NA | NA | 5.2 | NA | 3.8 | NA | [41] |
| European mole cricket, Gryllotalpa gryllotalpa | 6.0 | 2.1 | |||||||
| Two-spotted cricket, Gryllus bimaculatus | (50–67% NaOH) at 95 °C at 130 rpm | 41.75 | 56.47–84.98 | NA | NA | NA | NA | NA | [54] |
| Brachystola magna | 60% NaOH contained NaBH4 (0.004 g), 120 °C for 2 h | NA | 89.89 ± 1.34 | 696.95 × 103 | NA | NA | NA | NA | [35] |
| House crickets, Acheta domesticus | 50% NaOH for 3 h at 130 °C | 88.0 | 62.9 | 86.8 × 103 | NA | NA | NA | NA | [56] |
| House crickets, Acheta domesticus | 40% NaOH at 120 °C for 2 h | 90.6 | 88.5 | NA | NA | NA | NA | NA | [103] |
| House crickets, Acheta domesticus | 67% w/v NaOH for 2, 4 6, 10 h | 2 h-76.0 ± 6.7 4 h-77.3 ± 1.9 6 h-80.5 ± 2.1 10 h-69.0 ± 2.2 | 2 h-72.5 ± 1.0 4 h-76.3 ± 1.3 6 h-79.1 ± 1.9 10 h-79.4 ± 1.3 | 344 × 103 | NA | NA | NA | NA | [59] |
| Gryllodes sigillatus | 2 h-65.0 ± 1.6 4 h-63.7 ± 1.2 6 h-60.3 ± 3.3 10 h-62.3 ± 0.9 | 2 h-73.5 ± 1.4 4 h-74.9 ± 1.3 6 h-77.2 ± 1.8 10 h-81.3 ± 1.1 | 524 × 103 | ||||||
| Hymenoptera | |||||||||
| Western honey bee, Apsis mellifera | NA | NA | NA | NA | 7.7 ± 0.09 | NA | 2.4 ± 0.03 | NA | [61] |
| Western honey bee, Apsis mellifera | 50% NaOH at 100 °C for 8 h | NA | 96 | NA | NA | 17.6 | NA | 9.2 | [32] |
| Oriental hornet, Vespa orientalis | 50% NaOH at 100 °C for 2 h | NA | 96 | NA | NA | NA | NA | NA | [65] |
| Diptera | |||||||||
| Black soldier fly, Hermetia illucens | NaOH at 100 °C for 2 h | 32 | 90 | NA | NA | NA | NA | NA | [73] |
| Black soldier fly, Hermetia illucens | 50% (w/v) NaOH (1: 50) for 4 h at 95 °C | Late larvae—81.034 Prepupae—73.656 Pupal exuviae—79.701 Imagoes—63.158 | NA | NA | NA | NA | NA | NA | [75] |
| Black soldier fly, Hermetia illucens | 40% NaOH for 8 h | 6.58 | NA | NA | NA | NA | NA | NA | [76] |
| Black soldier fly, Hermetia illucens | 12 M NaOH for 4 h at 100 °C | Larvae—3 Pupal exuviae—10 Dead adults—3 | Larvae—92 Pupal exuviae—90 Dead adults—93 | Larvae—21 × 103 Pupal exuviae—35 × 103 Dead adults—36 × 103 | NA | NA | NA | NA | [80] |
| Black soldier fly, Hermetia illucens | 100 °C for 2 h | 81 | 66 | 505 × 103 | NA | NA | NA | NA | [83] |
| Common fruit fly, Drosophila melanogaster | 10 mL of NaOH solution (60%, w/w) for 48 h at 150 °C | 70.91 | NA | NA | NA | NA | NA | NA | [85] |
| Oriental blue fly, Chrysomya megacephala | 100 mL NaOH (1 mol/L) at 95 °C for 6 h | 26.2 | 89.6 | 501 × 103 | NA | NA | NA | NA | [104] |
| Housefly, Musca domestica | 50% NaOH | 57.9 ± 0.2 | 84.1 ± 0.3 | NA | NA | 7.8 ± 0.1 | NA | 8.2 ± 0.2 | [45] |
| Hemiptera | |||||||||
| Cicada slough | 60% NaOH at 100 °C for 8 h | 28.2 | 84.1 | (3.779 ± 0.068) × 104 | NA | 0.18 ± 0.016 | NA | 0.03 ± 0.004 | [21] |
| Aquatic bug, Ranatra linearis | NA | 70% | NA | NA | NA | NA | NA | NA | [88] |
| Dictyoptera | |||||||||
| American cockroach, Periplaneta americana | 50% NaOH in 100 °C for 4 h | 5.80 | 36.8 | NA | NA | NA | NA | NA | [105] |
| German cockroach, Blattella germanica | 2.95 | 31.5 | |||||||
| American cockroach, Periplaneta americana | 40% NaOH in 120 °C for 2 h | 99.7 | 90.3 | NA | NA | NA | NA | NA | [103] |
| American cockroach, Periplaneta americana | 10% NaOH at 80 °C for 3 h | NA | 90.85 ± 3.37 | (16 ± 0.746) × 103 | NA | NA | NA | NA | [96] |
| American cockroach, Periplaneta americana | 50% (w/v) NaOH for 8 h at 120 °C | 74.51 | NA | NA | NA | NA | NA | NA | [97] |
| Eupolyphaga sinensis | 50% NaOH at 90 °C for 24 h | 5.48 ± 0.32 | 96.57 ± 0.48 | (127.79 ± 1.35) × 103 | NA | 5.19 ± 0.11 | NA | 0.55 ± 0.05 | [91] |
| Ephemeroptera | |||||||||
| Mayfly | 60% NaOH at 150 °C for 6 h | 78.43 | 84.3 | 3.69 × 103 | NA | NA | NA | NA | [100] |
| Insect Species | Chitin (%) | Chitosan (%) | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Carbon (C) | Hydrogen (H) | Nitrogen (N) | Carbon/Nitrogen | Carbon (C) | Hydrogen (H) | Nitrogen (N) | Carbon/Nitrogen | ||
| Flour moth, Ephestia kuehniella | 43.12 | 6.11 | 5.89 | 7.057 | NA | NA | NA | NA | [22] |
| Butterfly, wings, Argynnis pandora | 44.89 | 6.53 | 6.62 | 6.781 | NA | NA | NA | NA | [23] |
| Butterfly, OBP, Argynnis pandora | 44.91 | 6.45 | 6.48 | 6.931 | NA | NA | NA | NA | |
| Common cockchafer, Melolontha melolontha | 45.09 | 6.29 | 6.72 | 6.71 | NA | NA | NA | NA | [28] |
| Superworm, Zophobas morio | 0.5 M-43.27 1.0 M-43.07 2.0 M-43.32 | 0.5 M-6.77 1.0 M-6.73 2.0 M-6.77 | 0.5 M-6.60 1.0 M-6.38 2.0 M-6.29 | 0.5 M-6.56 1.0 M-6.75 2.0 M-6.89 | 42.76 42.27 42.08 | 7.47 7.09 7.40 | 6.55 6.76 6.56 | 6.53 6.25 6.41 | [39] |
| European stag beetle, Lucanus cervus | 45.9 | 7.6 | 5.3 | 8.5 | NA | NA | NA | NA | [41] |
| Pine chafer, Polyphylla fullo | 45.4 | 7.5 | 5.1 | 8.9 | |||||
| Moroccan locust, Dociostaurus maroccanus | Adult—42.35 Nymph—47.32 | Adult—5.64 Nymph—6.64 | Adult—4.63 Nymph—5.66 | Adult—9.15 Nymph—8.36 | Adult—41.63 Nymph—42.01 | Adult—6.38 Nymph—6.28 | Adult—7.20 Nymph—6.45 | Adult—5.78 Nymph—6.51 | [48] |
| House cricket, Brachytrupes | 41.30 | NA | 6.022 | 6.858 | 38.98 | NA | 5.932 | 6.571 | [49] |
| Celes variabilis | 45.05–49.0 | 6.31–6.92 | 5.68–6.43 | 7.01–8.24 | NA | NA | NA | NA | [50] |
| Wart-biter, Decticus verrucivorus | |||||||||
| Melanogryllus desertus | |||||||||
| Paracyptera labiata | |||||||||
| Bradyporus sureyai European mole cricket, Gryllotalpa gryllotalpa | 46.6 ± 0.1 44.2 ± 0.1 | 7.7 ± 0.1 7.6 ± 0.1 | 5.3 ± 0.1 5.0 ± 0.1 | 8.8 8.8 | NA | NA | NA | NA | [41] |
| European hornet, Vespa crabro | 46.62 | 6.42 | 6.85 | 6.81 | NA | NA | NA | NA | [63] |
| Oriental hornet, Vespa orientalis | 46.01 | 6.34 | 6.71 | 6.86 | |||||
| German wasp, Vespula germanica | 44.94 | 5.95 | 6.90 | 6.51 | |||||
| Asian hornet, Vespa velutina | 43.47 | 6.94 | 6.85 | 6.35 | NA | NA | NA | NA | [64] |
| European hornet, Vespa crabro | Larval—45.6 Pupa—46.2 Adult—45.5 | Larval—6.5 Pupa—6.4 Adult—6.3 | Larval—6.5 Pupa—6.3 Adult—6.49 | Larval—7.02 Pupa—7.33 Adult—7.01 | NA | NA | NA | NA | [66] |
| Black soldier fly, Hermetia illucens | Pupal exuviae—35.23 Imago—32.09 | Pupal exuviae—5.11 Imago—4.80 | Pupal exuviae—3.73 Imago—3.9 | Pupal exuviae—9.45 Imago—8.23 | NA | NA | NA | NA | [68] |
| Black soldier fly, Hermetia illucens | Pupae exuviae—43.74 Imago—39.74 | Pupae exuviae—5.82 Imago—5.46 | Pupae exuviae—6.14 Imago—6.00 | Pupae exuviae—6.62 Imago—7.12 | NA | NA | NA | NA | [69] |
| Black soldier fly, Hermetia illucens | 41.84 | 6.74 | 5.96 | 7.02 | NA | NA | NA | NA | [73] |
| Oriental blue fly, Chrysomya megacephala | NA | NA | NA | NA | 39.06 ± 0.20 | 7.16 ± 0.08 | 7.30 ± 0.08 | 5.35 | [104] |
| American cockroach, Periplaneta americana | 45.74 | 6.59 | 6.69 | 6.84 | NA | NA | NA | NA | [95] |
| Dragonfly, Sympetrum fonscolombii | 47.09 | 6.65 | 6.83 | 6.89 | NA | NA | NA | NA | [99] |
| American cockroach, Pariplaneta Americana linnaeus | 47.23 | 7.32 | 7.20 | 6.56 | NA | NA | NA | NA | [62] |
| Western honey bee, Apis mellifera linneaus | 52.65 | 8.42 | 5.55 | 9.49 | NA | NA | NA | NA | [62] |
| Coridius nepalensis | 42.175 | 6.551 | 6.878 | 6.13 | NA | NA | NA | NA | [90] |
| American cockroach, Periplaneta americana | 43.84 | 6.93 | 6.33 | 6.92 | NA | NA | NA | NA | [98] |
| German cockroach, Blattella germanica | 46.28 | 6.84 | 6.84 | 6.77 | NA | NA | NA | NA | [44] |
| Dor beetle, Anoplotrupes stercorosus | 40.6 | 7.66 | 6.35 | 6.39 | |||||
| Blaps tibialis | 45.17 | 6.85 | 6.43 | 7.02 | |||||
| Rose chafer, Cetonia aurata | 43.6 | 7.13 | 6.87 | 6.35 | |||||
| Geotrupes stercorariu | 43.1 | 6.49 | 6.77 | 6.37 | |||||
| Calliphora vicina | 48.9 | 6.54 | 6.79 | 7.2 | |||||
| Dock bug, Coreus marginatus | 39.2 | 6.95 | 6.03 | 6.5 | |||||
| Black-and-red bug, Lygaeus equestris | 46.59 | 6.34 | 6.74 | 6.91 | |||||
| Pyrrhocoris apterus | 46.38 | 6.02 | 6.77 | 6.85 | |||||
| Red-tailed bumblebee, Bombus lapidaries | 40.1 | 7.48 | 6.11 | 6.56 | |||||
| Formica clara | 46.48 | 6.45 | 6.59 | 7.05 | |||||
| Downy emerald, Cordulia aenea | 44.6 | 6.86 | 6.66 | 6.70 | |||||
| Four-spotted chaser, Libellula quadrimaculata | 43.0 | 6.95 | 6.42 | 6.70 | |||||
| Pale giant horse-fly, Tabanus bovinus | 47.60 | 6.55 | 6.57 | 7.24 | 41.99 | 6.42 | 7.18 | 5.85 | [87] |
| Insect Species | Chitin (%) | Chitosan (%) | Ref. | ||
|---|---|---|---|---|---|
| XRD Peaks at 2θ | CrI (%) | XRD Peaks at 2θ | CrI (%) | ||
| Lepidoptera | |||||
| Silkworm | 9.6, 19.7, 12.7, 23.2, 26.3, 39 | 59.21 | 10, 20 | 32.9 | [21] |
| Butterfly, Argynnis pandora | Wings—9.3, 9.3, 12.84, 21.04, 22.9, 26.36 OBP—8.5, 19.3, 12.84, 21.14, 23.06, 6.66 | Wings—64 OBP—66 | NA NA | NA NA | [23] |
| Coleoptera | |||||
| Mealworm | 9.6, 19.7, 12.7, 23.2, 26.3, 28.1, 39 | 81.11 | 10, 20 | 51.9 | [21] |
| Omophlus sp. | 9.42, 12.72, 19.34, 20.84, 23.32, 26.44 | 82.9 | NA | NA | [27] |
| Cockchafer, Melolontha melolontha | NA | 75.2 | NA | NA | [28] |
| Cockchafer, Melolontha sp. | 9.32–9.70, 12.12–13.22, 12.78–13.22, 19.18–19.76, 20.06–21.48, 23.06–23.78, 26.02–26.80 | 74.1–88.9 | NA | NA | [29] |
| Colorado potato beetle, Leptinotarsa decemlineata | Larvae—9.6,13.22, 19.68, 21.42, 23.26, 26.7 Adults—9.66, 13.18, 19.48, 21.06, 23.16, 26.76 | Larvae—72 Adults—76 | NA | NA | [30] |
| Calosoma rugosa | NA | NA | 9.7, 20.3, 22.6 | 49 | [32] |
| Calosoma rugosa | NA | NA | 9.7, 20.3 | 49 | [33] |
| Mealworm beetle, Tenebrio molitor | 9.6, 19.6, 21.1, 23.7, 36 | 57.85 | NA | NA | [36] |
| Mealworm beetle, Tenebrio molitor | 9.011, 9.034, 19.119, 19.167, 21.38, 21.44, 22.68, 22.74 | NA | NA | NA | [106] |
| Mealworm beetle, (Tenebrio molitor, Zophobas morio) | NA | NA | 10.62, 20.02 | 58.11 | [38] |
| Rhinoceros beetle, Allomyrina dichotoma | NA | NA | 10.74, 19.92 | 62.77 | |
| European stag beetle, Lucanus cervus | 9.67, 12.40, 19.60, 23.41, 26.26, 39.1 | 85.2 | NA | NA | [41] |
| Pine chafer, Polyphylla fullo | 9.2, 12.40, 19.46, 23.50, 26.21, 28.1, 39.5 | 86.1 | |||
| Wheat weevil, Sitophilus granarius | 8.9, 9.2, 18.7, 25.6, 12.3, 22.8 | 78.77 | NA | NA | [43] |
| Dor beetle, Anoplotrupes stercorosus | 9.58, 13.36, 19.66, 21.14, 23.18, 26.52 | 83.5 | NA | NA | [44] |
| Blaps tibialis | 9.48, 12.76, 19.38, 21.08, 23.04, 26.64 | 80.1 | |||
| Rose chafer, Cetonia aurata | 9.44, 13.04, 19.52, 21.28, 23.46, 26 | 86.3 | |||
| Geotrupes stercorarius | 9.64, 13.14, 19.56, 21.38, 23.22, 26.76 | 80.1 | |||
| Blaps lethifera | NA | NA | 10.7, 19.9 | 84 | [45] |
| Pimelia fernandezlopezi | 11.5, 20.4 | 73 | |||
| Banana weevil, Cosmopolites sordidus | NA | NA | 19.5 | 41 | [46] |
| Orthoptera | |||||
| Grasshopper | 9.6, 19.7, 12.7, 23.2, 26.3, 28.1, 39.0 | 83.4 | 10, 20 | 50.1 | [21] |
| Moroccan locust, Dociostaurus maroccanus | Adult—9.56, 12.76, 19.72, 21.12, 23.96, 26.64 Nymph—9.42, 12.86, 19.72, 21.56, 23.38, 26.66 | Adult—71 Nymph—74 | Adult—10.96, 20.14 Nymph—10.76, 20.3 | NA | [48] |
| House cricket, Brachytrupes | 9.4, 12.8, 17.1, 19.4, 21.1, 23.2, 26.3, 28.5, 35.0, 39.0 | 88.02 | 9.6, 19.6, 21.2 | 86.64 | [49] |
| Celes variabilis | 9, 19, 12, 21, 23, 26 | 75–80 | NA | NA | [50] |
| Wart-biter, Decticus verrucivorus | |||||
| Melanogryllus desertus | |||||
| Paracyptera labiata | |||||
| Calliptamus barbarus | 9.26, 19.28, 21.24, 23.28, 26.36, 31.78 | 70.9 | 10.92, 20.08 | NA | [51] |
| Oedaleus decorus | 9.6, 19.6, 21.1,23.7, 26.64 | 76.8 | 10.08, 20.14 | ||
| Ailopus simulatrix | 9.3, 12.7, 19.6, 21.1, 23.8, 26.6 | 76 | NA | NA | [52] |
| Ailopus strepens | 9.5, 12.8, 19.6, 20.8, 23.8, 26.4 | 75 | |||
| Duroniella fracta | 9.5, 12.6, 19.4, 20.9, 23.5, 26.8 | 72 | |||
| Duroniella laticornis | 9.5, 12.8, 19.3, 20.7, 23.2, 26.5 | 71 | |||
| Red-winged grasshopper, Oedipoda miniata | 9.7, 12.9, 19.6, 21, 23.7, 26,8 | 74 | |||
| Oedipoda caerulescens | 9.3, 12.7, 19.3, 20.7, 23.1, 26.9 | 74 | |||
| Pyrgomorpha cognata | 9.4, 13.3, 19.6, 20.9, 23.4, 26,9 | 63 | |||
| Desert locust, Schistocerca gregaria | NA | NA | 9.3, 20.2, 24.4 | 69 | [32] |
| Bradyporus sureyai | 9.62, 12.5, 19.72, 23.74, 26.22, 27.8, 39.2 | 83.1 | NA | NA | [41] |
| European mole cricket, Gryllotalpa gryllotalpa | 9.44, 12.3, 19.41, 23.31, 26.2, 27.9, 39.0 | 80.6 | |||
| Two-spotted cricket, Gryllus bimaculatus | NA | NA | 10.50, 20.07 | 57.52 | [54] |
| Shistocerca gregarea Forsskal | 9.2, 19.1,12.6, 22.9, 26.2 | 71.4 | NA | NA | [42] |
| Hymenoptera | |||||
| Western honey bee, Apsis mellifera | NA | NA | 9.7, 20.3 | 59 | [32] |
| Western honey bee, Apsis mellifera | NA | NA | 9.7, 20.3 | 59 | [33] |
| European hornet, Vespa crabro | 9.64, 12.74, 19.38, 20.94, 23.92, 26.88 | 69.88 | NA | NA | [63] |
| Oriental hornet, Vespa orientalis | 9.68, 12.72, 19.32, 21.6, 23.74, 26.8 | 53.92 | |||
| European wasp, Vespula germanica | 9.32, 12.92, 20.l04, 21.24, 23.16, 25.9 | 50 | |||
| Oriental hornet, Vespa orientalis | 9.2, 19.1,12.6, 22.9, 26.2 | 39.4 | NA | NA | [42] |
| Red-tailed bumblebee, Bombus lapidarius | 9.64, 13.02, 19.58, 21.22, 23.44, 26.78 | 75.5 | NA | NA | [44] |
| Formica clara | 9.5, 13.38, 19.78, 20.84, 23.1, 26.76 | 69.8 | |||
| Diptera | |||||
| Musca domestica | NA | NA | 10.5, 20.2 | 81 | [45] |
| Black soldier fly, Hermetia illucens | 9.4, 13.0, 19.3, 20.8, 23.2, 29.5 | Prepupae—94 Cocoons—94 Sheddings—89 Larvae—89 | NA | NA | [67] |
| Black soldier fly, Hermetia illucens | 9, 19, 22, 24, 30 | Larval—35 Imago—24.9 | NA | NA | [68] |
| Black soldier fly, Hermetia illucens | 9.3, 19.8, 23, 26.0 | Pupae exuvia—25.2 Imago—49.4 | NA | NA | [69] |
| Black soldier fly, Hermetia illucens | Larvae—9.30, 12.78, 19.26, 21.82, 23.31, 26.41 Prepupa—9.38, 12.93, 19.33, 21.19, 23.42, 26.37 Puparium—9.30, 12.67, 19.29, 20.77, 23.38, 26.45 Adult—9.50, 12.82, 19.33, 20.81, 23.31, 26.34 | Larvae—33.09 Prepupa—35.14 Puparium—68.44 Adult—87.92 | NA | NA | [71] |
| Black soldier fly, Hermetia illucens | 9, 19, 13, 21, 23, 26 | Larvae—84 Pupal exuviae—62 Dead adults—93 | 10, 20 | Larvae—77 Pupal exuviae—80 Dead adults—86 | [80] |
| Calliphora vicina | 9.38, 12.88, 19.3, 20.8, 22.84, 26.8 | 67.1 | NA | NA | [44] |
| Hemiptera | |||||
| Cicada slough | 9.6, 19.7, 12.7, 23.2, 26.3, 39 | 85.21 | 10, 20, 23.2 | 49.1 | [21] |
| Aquatic bug, Ranatra linearis | 9.34, 12.38, 19.66, 20.88, 23.22, 26.56, 38.96 | 84.8 | NA | NA | [88] |
| Coridius nepalensis | 9, 20, 20.5, 22.8, 26.4 | 86.33 | NA | NA | [90] |
| Dock bug, Coreus marginatus | 9.7, 13.2, 19.86, 21.24, 23.42, 26.54 | 76.9 | NA | NA | [44] |
| Black-and-red bug, Lygaeus equestris | 9.64, 13, 19.76, 21.16, 22.8, 26.7 | 59.7 | |||
| Pyrrhocoris apterus | 9.44, 12.52, 19.14, 20.84, 22.66, 26.7 | 62.1 | |||
| Dictyoptera | |||||
| American cockroach, Periplaneta americana | 9.14, 19.58, 12.88, 20.98, 23.12, 26.8 | 86.7 | NA | NA | [95] |
| American cockroach, Periplaneta americana | 12, 19, 20.5, 21.5, 23, 26 | 83.72 | NA | NA | [98] |
| German cockroach, Blattela germanica | 9.2, 19.1, 12.6, 22.9, 26.2 | 44.2 | NA | NA | [42] |
| German cockroach, Blattella germanica | 9.4, 12.7, 19.5, 20.68, 23.33, 26.66 | 70.1 | NA | NA | [44] |
| Odonata | |||||
| Dragonfly, Sympetrum, fonscolombii | 9, 13, 19, 21, 26 | 96.4 | NA | NA | [99] |
| Downy emerald, Cordulia aenea | 9.54, 13.18, 19.62, 21.4, 23.76, 26.92 | 73.2 | NA | NA | [44] |
| Four-spotted chaser, Libellula quadrimaculata | 9.54, 13.24, 19.68, 21.06, 23.1, 26.88 | 63.9 | |||
| Insect Species | Chitin (%) | Chitosan (%) | Ref. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| First | Second | DTGmax (°C) | First Peak | Second Peak | DTGmax (°C) | ||||||
| Mass Loss (%) | T (°C) | Mass Loss (%) | T (°C) | Mass Loss (%) | T (°C) | Mass Loss (%) | T (°C) | ||||
| Lepidoptera | |||||||||||
| Silkworm, Bombyx mori | NA | NA | NA | NA | NA | NA | 80–90 | NA | 290–300 | NA | [21] |
| Butterfly, Argynnis pandora | Wings—4.85 OBP—4.82 | 30–100 | Wings—82.23 OBP—80 | 100–650 | Wings—386.9 OBP—399.6 | NA | NA | NA | NA | NA | [23] |
| Coleoptera | |||||||||||
| Mealworm, Tenebrio molitor | NA | NA | 80–90 | NA | 290–300 | NA | [21] | ||||
| Omophlus sp. | 3.6% (α) 7.7% (β) | NA | 78.8% (α) 63.5% (β) | NA | 385.3 (α) 334.2 (β) | NA | NA | NA | NA | NA | [27] |
| Cockchafer, Melolontha melolontha | 4 | 0–150 | 78 | 150–600 | 380 | NA | NA | NA | NA | NA | [28] |
| Cockchafer, Melolontha sp. | 1.1–5.4 | NA | 51.2–94.5 | NA | Female—392.2 Male—378.3 | NA | NA | NA | NA | NA | [29] |
| Colorado potato beetle, Leptinotarsa decemlineata | 3–5 | NA | Adult—74 Larval—48 | NA | Adult—379 Larval—307 | 3–5 | NA | 94–96 | NA | Adult—289 Larval—292 | [102] |
| Catharsius molossus | NA | 13.37 | 25–290 | 45.76 | 290–400 | 317.70 | [31] | ||||
| European stag beetle, Lucanus cervus | 6.6 | 100 | 70 | 300–420 | 379.9 | NA | NA | NA | NA | NA | [41] |
| Pine chafer, Polyphylla fullo | 5.9 | 100 | 73 | 374.7 | |||||||
| Dor beetle, Anoplotrupes stercorosus | 4 | NA | 74 | NA | 387 | NA | NA | NA | NA | NA | [44] |
| Blaps tibialis | 7 | 76 | 385 | ||||||||
| Rose chafer, Cetonia aurata | 7 | 82 | 361 | ||||||||
| Geotrupes stercorarius | 5 | 77 | 390 | ||||||||
| Orthoptera | |||||||||||
| Grasshopper | NA | NA | 80–90 | NA | 290–300 | NA | [21] | ||||
| Moroccan locust, Dociostaurus maroccanus | 4 | NA | Adult—77 Nymph—82 | NA | Adult—386 Nymph—383 | Adult—5 Nymph—7 | NA | Adult—62 Nymph—59 | NA | Adult—302 Nymph—308 | [48] |
| Celes variabilis | 3–6 | 0–150 | 73–96 | 150–400 | 350–387 | NA | NA | NA | NA | NA | [50] |
| Wart-biter, Decticus verrucivorus | |||||||||||
| Melanogryllus desertus | |||||||||||
| Paracyptera labiata | |||||||||||
| Calliptamus barbarus | 8 | 0–150 | 72 | 150–650 | 381 | 8 | 0–150 | 61 | 300 | 296 | [51] |
| Oedaleus decorus | 6 | 77 | 390 | 6 | 57 | 305 | |||||
| Ailopus simulatrix | NA | 0–150 | 82 | 150–600 | 383 | NA | NA | NA | NA | NA | [52] |
| Ailopus strepens | 78 | 382 | |||||||||
| Duroniella fracta | 74 | 381 | |||||||||
| Duroniella laticornis | 72 | 382 | |||||||||
| Red-winged grasshopper, Oedipoda miniata | 76 | 385 | |||||||||
| Oedipoda caerulescens | 77 | 384 | |||||||||
| Pyrgomorpha cognata | 74 | 384 | |||||||||
| Bradyporus sureyai | 5.2 | 0–100 | 72 | 300–420 | 382.4 | NA | NA | NA | NA | NA | [41] |
| European mole cricket, Gryllotalpa gryllotalpa | 6.0 | 70 | 300–412 | 374.6 | |||||||
| House crickets, Acheta domesticus | 7.4 | 0–150 | 53 | 200–400 | 359.2 | NA | NA | NA | NA | NA | [56] |
| Hymenoptera | |||||||||||
| Western honey bee, Apsis mellifera | Head—6 Thorax—4 Abdomen—3 Legs—5 Wings—3 | NA | Head—67 Thorax—56 Abdomen—68 Legs—68 Wings—60 | NA | Head—308 Thorax—360 Abdomen—367 Legs—359 Wings—359 | NA | NA | NA | NA | NA | [60] |
| European hornet, Vespa crabro | Larvae—3.51 Pupa—2.7 Adult—6.5 | 30–180 | Larvae—88.7 Pupa—69.9 Adult—78.3 | NA | Larvae—384.8 Pupa—381.7 Adult—382.4 | NA | NA | NA | NA | NA | [66] |
| Red-tailed bumblebee, Bombus lapidarius | 5 | NA | 72 | NA | 384 | NA | NA | NA | NA | NA | [44] |
| Formica clara | 4 | 78 | 374 | ||||||||
| Diptera | |||||||||||
| Black soldier fly, Hermetia illucens | 2–3 | 0–122 | 62–63 | 122–450 | Imago—387 Larval—389 | NA | NA | NA | NA | NA | [68] |
| Black soldier fly, Hermetia illucens | 5–6 | 74–110 | 70–80 | 250 | Imago—363 Pupae exuvia—371 | NA | NA | NA | NA | NA | [69] |
| Black soldier fly, Hermetia illucens | Larvae—4.42 Prepupa—6.74 Puparium—8.52 Adults—7.5 | 0–150 | Larvae—69.48 Prepupa—71.16 Puparium—71.25 Adults—3.31 | 150–400 | Larvae—372 Prepupa—373 Puparium—371 Adults—372 | NA | NA | NA | NA | NA | [71] |
| Common fruit fly, Drosophila melanogaster | 4.4 | NA | 75.6 | NA | 378.7 | 4.47 | NA | 61.12 | NA | 304.7 | [85] |
| Calliphora vicina | 4 | NA | 65 | NA | 379 | NA | NA | NA | NA | NA | [44] |
| Pale giant horse-fly, Tabanus bovinus | 4.6 | 30–150 | 80.2 | 200–650 | 370.50 | 7.3 | 30–150 | 59.3 | 200–650 | 295.4 | [87] |
| Hemiptera | |||||||||||
| Cicada slough | NA | NA | 80–90 | NA | 290–300 | NA | [21] | ||||
| Aquatic bug, Ranatra linearis | 6 | 0–150 | 78 | 150–650 | 393 | 9 | 0–150 | 50 | 150–650 | 289 | [88] |
| Cicada lodosi | 4.41 | 0–200 | 83.94 | 200–750 | 411.70 | NA | NA | NA | NA | NA | [89] |
| Cicada mordoganensis | 4.88 | 80.44 | 412.40 | ||||||||
| Cicadatra platyptera | 3.80 | 81.78 | 412.20 | ||||||||
| Cicadatra atra | 4.54 | 83.75 | 411.50 | ||||||||
| Cicadatra hyaline | 5.4 | 66.78 | 412.70 | ||||||||
| Cicadivetta tibialis | 4.04 | 73.49 | 339.90–402.30 | ||||||||
| Dock bug, Coreus marginatus Black-and-red bug, Lygaeus equestris Pyrrhocoris apterus | 9 3 5 | NA | 73 66 78 | NA | 389 375 387 | NA | NA | NA | NA | NA | [44] |
| Dictyoptera | |||||||||||
| American cockroach, Periplaneta americana | 5 | 100 | 76 | 350–390 | 389 | NA | NA | NA | NA | NA | [95] |
| German cockroach, Blattella germanica | 4 | NA | 77 | NA | 389 | NA | NA | NA | NA | NA | [44] |
| Odonata | |||||||||||
| Dragonfly, Sympetrum fonscolombii | 2.9 | 25–100 | 73.2 | 100–750 | 369.2 | NA | NA | NA | NA | NA | [99] |
| Downy emerald, Cordulia aenea | 4 | NA | 75 | NA | 378 | NA | NA | NA | NA | NA | [44] |
| Four-spotted chaser, Libellula quadrimaculata | 6 | 76 | 384 | ||||||||
| Insect Species | Chemical Shift (ppm) | Ref. | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | C=O | CH3 | ||
| Blowfly larvae, chitosan | 104.47 | 56.78 | 75.14 | 85.31 | 75.14 | 60.41 | NA | 22.64 | [59] |
| Black soldier fly, chitin | 104.6 | 55.7 | 74.2 | 84.0 | 76.4 | 61.5 | 173.9 | 23.4 | [69] |
| European rhinoceros beetle, Oryctes nasicornis, chitin film | 104 | 55 | 73 | 83 | 75 | 61 | NA | NA | [107] |
| Asian hornet, Vespa velutina, chitin | 104.12 | 55.94 | NA | 82.74 | 75.34 | 60.34 | 173.17 | 22.79 | [64] |
| Black soldier fly, Hermetia illucens Pupal, chitin | 104.3 | 55.2 | 73.5 | 83.2 | 75.8 | 60.9 | 173.1 | 22.9 | [108] |
| Mealworm, Tenebrio molitor Linnaeus 1758, chitin | 104.63 | 55.71 | 76.14 | 83.63 | 76.14 | 61.46 | 173 | 23 | [109] |
| Monarchs, Swallowtails, chitin | 104 | NA | 73 | 83 | 76 | NA | 173 | 23 | [110] |
3. Extraction of Chitin and Chitosan from Insects
3.1. Chemical Extraction
3.1.1. Delipidation
3.1.2. Demineralization
3.1.3. Deproteinization
3.1.4. Decolorization
3.1.5. Deacetylation
3.2. Biological Extraction
3.3. Other Extraction Methods
4. Characterization and Modification of Chitin and Chitosan from Insects
4.1. Physicochemical and Structural Characterization
4.1.1. Extraction Yield
4.1.2. Degree of Deacetylation (DDA)
4.1.3. Molecular Weight
4.1.4. Moisture Content and Ash Content
4.1.5. Elemental Analysis
4.1.6. Solubility
4.1.7. Fourier Transform Infrared Spectroscopy
4.1.8. Crystalline Properties
4.1.9. Thermogravimetric Analysis
4.1.10. Nuclear Magnetic Resonance Spectroscopy
4.1.11. Scanning Electron Microscopy
5. Biomedical Applications of Chitin and Chitosan from Insects
5.1. Antioxidant and Anti-Agng Activity
5.2. Antibacterial Activity
| Insect Order | Insect Species | Bacteria Strains | Ref. |
|---|---|---|---|
| Coleoptera | Cosmopolites sordidus | Escherichia coli, Klebsiella pneumoniae | [46] |
| Tenebrio molitor | Proteus mirabilis, Klebsiella pneumoniae, Enterococcus faecalis, Staphylococcus epidermidis | [105] | |
| Tenebrio Molitor | Bacillus cereus, Listeria monocytogenes, Escherischia coli, Staphylococcus aureus | [38] | |
| Hylobius abietis L. | Enterococcus faecalis, Enterobacter aerogenes, Bacillus subtilis, Streptococcus pneumonia, Vibrio parahoe molyticus, Acinetobacter baumannii, Bacillus megaterium, Bacillus cereus, Micrococcus luteus Klebsiella oxytoca, Pseudomonas aeruginosa, Shigella sonnei, Staphylococcus epidermitis wt, Staphylococcus aureus, Klebsiella pneumoniae subsp. Pneumoniae, Pseudomonas aeruginosa, Pseudomonas aeruginosa, Escherichia coli | [147] | |
| Tenebrio molitor | Pseudomonas aeruginosa | [37] | |
| Blaps lethifera, Pimelia fernandezlopezi | Listeria innocua, Bacillus subtiliis, Staphylococcus aureus, Salmonella typhimurium, Pseudomonas aeruginosa | [45] | |
| Tenebrio molitor | Proteus mirabilis, Klebsiella pneumoniae, Enterococcus faecalis, Staphylococcus epidermidis | [105] | |
| Dictyoptera | American Cockroach German Cockroach | Micrococcus luteus, Staphylococcus aureus, Pseudomonas aeruginosa, Escherichia coli | [93] |
| American cockroach, Periplaneta americana | Escherichia coli, Klebsiella pneumoniae, Staphylococcus aureus, Bacillus subtilis | [97] | |
| Periplaneta americana | Serratia marcescens, Escherichia coli | [96] | |
| Periplaneta americana Blattella germanica | Proteus mirabilis, Klebsiella pneumoniae, Enterococcus faecalis, Staphylococcus epidermidis | [105] | |
| Diptera | black soldier fly, Hermetia illucens | Pseudomonas aeruginosa, Serratia marcescens | [75] |
| Escherichia coli, Bacillus subtilis, Pseudomonas aeruginosa, Staphylococcus aureus, Candida albicans | [76] | ||
| Escherichia coli, Micrococcus favus | [77] | ||
| Escherichia coli, Staphylococcus epidermidis | [83] | ||
| Musca domestica | Listeria innocua, Bacillus subtiliis, Staphylococcus aureus, Salmonella typhimurium, Pseudomonas aeruginosa | [45] | |
| Orthoptera | Calliptamus barbarous, Oedaleus decorus | Listeria monocytogenes, Bacillus subtilis, Salmonella enteritidis, Yersinia enterocolitica, Candida albicans | [51] |
| Acheta domesticus, Gryllodes sigillatus | Escherichia coli, Listeria innocua | [59] | |
| Hymenoptera | Vespa orientalis | Klebsiella pneumoniae, Escherichia coli, Pseudomonas aeruginosa | [65] |
5.3. Anticancer Activity
5.4. Insect Chitosan-Based Biomimetic Materials for Wound Management
5.5. Insect Chitosan-Based Biomimetics Drug Delivery System
5.6. Other Biomedical Applications
6. Conclusion and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Khattak, S.; Wahid, F.; Liu, L.P.; Jia, S.R.; Chu, L.Q.; Xie, Y.-Y.; Li, Z.-X.; Zhong, C. Applications of Cellulose and Chitin/Chitosan Derivatives and Composites as Antibacterial Materials: Current State and Perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 1989–2006. [Google Scholar] [CrossRef] [PubMed]
- Hahn, T.; Tafi, E.; Paul, A.; Salvia, R.; Falabella, P.; Zibek, S. Current State of Chitin Purification and Chitosan Production from Insects. J. Chem. Technol. Biotechnol. 2020, 95, 2775–2795. [Google Scholar] [CrossRef]
- Marzieh, M.-N.; Zahra, F.; Tahereh, E.; Sara, K.-N. Comparison of the Physicochemical and Structural Characteristics of Enzymatic Produced Chitin and Commercial Chitin. Int. J. Biol. Macromol. 2019, 139, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Salavati, M. Mechanical Properties of α-Chitin and Chitosan Biocomposite: A Molecular Dynamic Study. J. Compos. Sci. 2023, 7, 464. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A. Chitin Nanostructures in Living Organisms. In Chitin; Gupta, N.S., Ed.; Topics in Geobiology; Springer: Dordrecht, The Netherlands, 2011; Volume 34, pp. 1–34. ISBN 978-90-481-9683-8. [Google Scholar]
- Feng, M.; Lu, X.; Hou, D.; Zhang, S. Solubility, Chain Characterization, and Derivatives of Chitin. In Handbook of Chitin and Chitosan; Elsevier: Amsterdam, The Netherlands, 2020; pp. 101–129. ISBN 978-0-12-817970-3. [Google Scholar]
- Kumari, S.; Kishor, R. Chitin and Chitosan: Origin, Properties, and Applications. In Handbook of Chitin and Chitosan; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–33. ISBN 978-0-12-817970-3. [Google Scholar]
- Yen, M.-T.; Mau, J.-L. Selected Physical Properties of Chitin Prepared from Shiitake Stipes. LWT-Food Sci. Technol. 2007, 40, 558–563. [Google Scholar] [CrossRef]
- Sagheer, F.A.A.; Al-Sughayer, M.A.; Muslim, S.; Elsabee, M.Z. Extraction and Characterization of Chitin and Chitosan from Marine Sources in Arabian Gulf. Carbohydr. Polym. 2009, 77, 410–419. [Google Scholar] [CrossRef]
- Kaya, M.; Mujtaba, M.; Ehrlich, H.; Salaberria, A.M.; Baran, T.; Amemiya, C.T.; Galli, R.; Akyuz, L.; Sargin, I.; Labidi, J. On Chemistry of γ-Chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Morin-Crini, N.; Lichtfouse, E.; Torri, G.; Crini, G. Fundamentals and Applications of Chitosan. In Sustainable Agriculture Reviews 35; Crini, G., Lichtfouse, E., Eds.; Sustainable Agriculture Reviews; Springer International Publishing: Cham, Switzerland, 2019; Volume 35, pp. 49–123. ISBN 978-3-030-16537-6. [Google Scholar]
- Pellis, A.; Guebitz, G.M.; Nyanhongo, G.S. Chitosan: Sources, Processing and Modification Techniques. Gels 2022, 8, 393. [Google Scholar] [CrossRef]
- Bumgardner, J.D.; Murali, V.P.; Su, H.; Jenkins, O.D.; Velasquez-Pulgarin, D.; Jennings, J.A.; Sivashanmugam, A.; Jayakumar, R. Characterization of Chitosan Matters. In Chitosan Based Biomaterials Volume 1; Elsevier: Amsterdam, The Netherlands, 2017; pp. 81–114. ISBN 978-0-08-100230-8. [Google Scholar]
- Leke-Aladekoba, A.A. Comparison of Extraction Methods and Characterisation of Chitin and Chitosan with Antimicrobial and Antioxidant Properties from Black Soldier Fly (Hermetia illucens) Meal; Dalhousie University: Halifax, NS, Canada, 2018. [Google Scholar]
- Jiménez-Gómez, C.P.; Cecilia, J.A. Chitosan: A Natural Biopolymer with a Wide and Varied Range of Applications. Molecules 2020, 25, 3981. [Google Scholar] [CrossRef]
- Zainol Abidin, N.A.; Kormin, F.; Zainol Abidin, N.A.; Mohamed Anuar, N.A.F.; Abu Bakar, M.F. The Potential of Insects as Alternative Sources of Chitin: An Overview on the Chemical Method of Extraction from Various Sources. Int. J. Mol. Sci. 2020, 21, 4978. [Google Scholar] [CrossRef]
- Rehman, K.U.; Hollah, C.; Wiesotzki, K.; Heinz, V.; Aganovic, K.; Rehman, R.U.; Petrusan, J.-I.; Zheng, L.; Zhang, J.; Sohail, S.; et al. Insect-Derived Chitin and Chitosan: A Still Unexploited Resource for the Edible Insect Sector. Sustainability 2023, 15, 4864. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, J.; Zhu, K.Y. Chito-Protein Matrices in Arthropod Exoskeletons and Peritrophic Matrices. In Extracellular Sugar-Based Biopolymers Matrices; Cohen, E., Merzendorfer, H., Eds.; Biologically-Inspired Systems; Springer International Publishing: Cham, Switzerland, 2019; Volume 12, pp. 3–56. ISBN 978-3-030-12918-7. [Google Scholar]
- Muthukrishnan, S.; Merzendorfer, H.; Arakane, Y.; Kramer, K.J. Chitin Metabolism in Insects. In Insect Molecular Biology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2012; pp. 193–235. ISBN 978-0-12-384747-8. [Google Scholar]
- Mishyna, M.; Glumac, M. So Different, yet so Alike Pancrustacea: Health Benefits of Insects and Shrimps. J. Funct. Foods 2021, 76, 104316. [Google Scholar] [CrossRef]
- Luo, Q.; Wang, Y.; Han, Q.; Ji, L.; Zhang, H.; Fei, Z.; Wang, Y. Comparison of the Physicochemical, Rheological, and Morphologic Properties of Chitosan from Four Insects. Carbohydr. Polym. 2019, 209, 266–275. [Google Scholar] [CrossRef]
- Mehranian, M.; Pourabad, R.F.; Bashir, N.S.; Taieban, S. Physicochemical Characterization of Chitin from the Mediterranean Flour Moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). J. Macromol. Sci. Part A 2017, 54, 720–726. [Google Scholar] [CrossRef]
- Kaya, M.; Bitim, B.; Mujtaba, M.; Koyuncu, T. Surface Morphology of Chitin Highly Related with the Isolated Body Part of Butterfly (Argynnis Pandora). Int. J. Biol. Macromol. 2015, 81, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-J.; Pan, S.-K.; Wang, H.-B.; Wu, J.-H. Preparation of Chitooligosaccharides from Cicada Slough and Their Antibacterial Activity. Int. J. Biol. Macromol. 2013, 62, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Chen, J.; Wu, S. Hypolipidemic Activity of the Chitooligosaccharides from Clanis Bilineata (Lepidoptera), an Edible Insect. Int. J. Biol. Macromol. 2013, 59, 96–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lu, M.; Wang, S. Antiageing Activities of Water-Soluble Chitosan from Clanis Bilineata Larvae. Int. J. Biol. Macromol. 2017, 102, 376–379. [Google Scholar] [CrossRef]
- Kaya, M.; Sargin, I.; Aylanc, V.; Tomruk, M.N.; Gevrek, S.; Karatoprak, I.; Colak, N.; Sak, Y.G.; Bulut, E. Comparison of Bovine Serum Albumin Adsorption Capacities of α-Chitin Isolated from an Insect and β-Chitin from Cuttlebone. J. Ind. Eng. Chem. 2016, 38, 146–156. [Google Scholar] [CrossRef]
- Kaya, M.; Baublys, V.; Can, E.; Šatkauskienė, I.; Bitim, B.; Tubelytė, V.; Baran, T. Comparison of Physicochemical Properties of Chitins Isolated from an Insect (Melolontha melolontha) and a Crustacean Species (Oniscus asellus). Zoomorphology 2014, 133, 285–293. [Google Scholar] [CrossRef]
- Kaya, M.; Bulut, E.; Mujtaba, M.; Sivickis, K.; Sargin, I.; Akyuz, B.; Erdogan, S. Gender Influences Differentiation of Chitin among Body Parts: Differentiation of Chitin Among Body Parts. Arch. Insect Biochem. Physiol. 2016, 93, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Saman, I.; Menteş, A.; Cakmak, Y.S.; Baran, T.; Kaya, M.; Asan Ozusaglam, M. Physicochemical Characterization of Chitin and Chitosan Obtained from Resting Eggs of Ceriodaphnia Quadrangula (Branchiopoda: Cladocera: Daphniidae). J. Crustac. Biol. 2014, 34, 283–288. [Google Scholar] [CrossRef]
- Ma, J.; Xin, C.; Tan, C. Preparation, Physicochemical and Pharmaceutical Characterization of Chitosan from Catharsius Molossus Residue. Int. J. Biol. Macromol. 2015, 80, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Marei, N.H.; El-Samie, E.A.; Salah, T.; Saad, G.R.; Elwahy, A.H.M. Isolation and Characterization of Chitosan from Different Local Insects in Egypt. Int. J. Biol. Macromol. 2016, 82, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Marei, N.; Elwahy, A.H.M.; Salah, T.A.; El Sherif, Y.; El-Samie, E.A. Enhanced Antibacterial Activity of Egyptian Local Insects’ Chitosan-Based Nanoparticles Loaded with Ciprofloxacin-HCl. Int. J. Biol. Macromol. 2019, 126, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kim, M.; Moon, C.; Seo, D.; Han, Y.S.; Jo, Y.H.; Noh, M.Y.; Park, Y.; Kim, S.; Kim, Y.W.; et al. Extraction of Chitin and Chitosan from Larval Exuvium and Whole Body of Edible Mealworm, Tenebrio molitor. Entomol. Res. 2018, 48, 227–233. [Google Scholar] [CrossRef]
- Saenz-Mendoza, A.I.; Zamudio-Flores, P.B.; García-Anaya, M.C.; Velasco, C.R.; Acosta-Muñiz, C.H.; De Jesús Ornelas-Paz, J.; Hernández-González, M.; Vargas-Torres, A.; Aguilar-González, M.Á.; Salgado-Delgado, R. Characterization of Insect Chitosan Films from Tenebrio molitor and Brachystola Magna and Its Comparison with Commercial Chitosan of Different Molecular Weights. Int. J. Biol. Macromol. 2020, 160, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.-J.; Hwang, I.-K.; Nho, C.W.; Kim, S.M.; Kim, S.H. Determination of Carbohydrate Composition in Mealworm (Tenebrio molitor L.) Larvae and Characterization of Mealworm Chitin and Chitosan. Foods 2021, 10, 640. [Google Scholar] [CrossRef]
- Nafary, A.; Mousavi Nezhad, S.; Jalili, S. Extraction and Characterization of Chitin and Chitosan from Tenebrio molitor Beetles and Investigation of Its Antibacterial Effect against Pseudomonas Aeruginosa. Adv. Biomed. Res. 2023, 12, 96. [Google Scholar] [CrossRef]
- Shin, C.-S.; Kim, D.-Y.; Shin, W.-S. Characterization of Chitosan Extracted from Mealworm Beetle (Tenebrio molitor, Zophobas morio) and Rhinoceros Beetle (Allomyrina dichotoma) and Their Antibacterial Activities. Int. J. Biol. Macromol. 2019, 125, 72–77. [Google Scholar] [CrossRef]
- Soon, C.Y.; Tee, Y.B.; Tan, C.H.; Rosnita, A.T.; Khalina, A. Extraction and Physicochemical Characterization of Chitin and Chitosan from Zophobas morio Larvae in Varying Sodium Hydroxide Concentration. Int. J. Biol. Macromol. 2018, 108, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Zhatkanbayev, Y.; Zhatkanbayeva, Z.; Iskakova, Z.; Kolpek, A.; Serikov, A.; Moldagulova, N.; Danlybayeva, G.; Sarsenova, A.; Anuarbekova, S. Application of Chitosan-Based Hydrogel Obtained from Insects in Pine Planting. Int. J. Biomater. 2023, 2023, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kabalak, M.; Aracagök, D.; Torun, M. Extraction, Characterization and Comparison of Chitins from Large Bodied Four Coleoptera and Orthoptera Species. Int. J. Biol. Macromol. 2020, 145, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Badawy, M.E.I.; Rabea, E.I. A Biopolymer Chitosan and Its Derivatives as Promising Antimicrobial Agents against Plant Pathogens and Their Applications in Crop Protection. Int. J. Carbohydr. Chem. 2011, 2011, 1–29. [Google Scholar] [CrossRef]
- Jagdale, P.; Mharsale, N.; Gotarne, R.; Magdum, S. Extraction and Characterization of Chitin from Granary Weevil, Sitophilus granaries L. (Coleoptera: Curculionidae). Arthropods 2022, 11, 176–185. [Google Scholar]
- Kaya, M.; Baublys, V.; Sargin, I.; Šatkauskienė, I.; Paulauskas, A.; Akyuz, B.; Bulut, E.; Tubelytė, V.; Baran, T.; Seyyar, O.; et al. How Taxonomic Relations Affect the Physicochemical Properties of Chitin. Food Biophys. 2016, 11, 10–19. [Google Scholar] [CrossRef]
- Amor, I.B.; Hemmami, H.; Laouini, S.E.; Abdelaziz, A.G.; Barhoum, A. Influence of Chitosan Source and Degree of Deacetylation on Antibacterial Activity and Adsorption of AZO Dye from Water. Biomass Convers. Biorefinery 2023. [Google Scholar] [CrossRef]
- Ssekatawa, K.; Byarugaba, D.K.; Wampande, E.M.; Moja, T.N.; Nxumalo, E.; Maaza, M.; Sackey, J.; Ejobi, F.; Kirabira, J.B. Isolation and Characterization of Chitosan from Ugandan Edible Mushrooms, Nile Perch Scales and Banana Weevils for Biomedical Applications. Sci. Rep. 2021, 11, 4116. [Google Scholar] [CrossRef]
- Torres-Castillo, J.A.; Sinagawa-García, S.R.; Lara-Villalón, M.; Martínez-Ávila, G.C.G.; Mora-Olivo, A.; Reyes-Soria, F.A. Evaluation of Biochemical Components from Pterophylla beltrani (Bolivar & Bolivar) (Orthoptera: Tettigoniidae): A Forest Pest from Northeastern Mexico. Southwest. Entomol. 2015, 40, 741–751. [Google Scholar] [CrossRef]
- Erdogan, S.; Kaya, M. High Similarity in Physicochemical Properties of Chitin and Chitosan from Nymphs and Adults of a Grasshopper. Int. J. Biol. Macromol. 2016, 89, 118–126. [Google Scholar] [CrossRef]
- Ibitoye, E.B.; Lokman, I.H.; Hezmee, M.N.M.; Goh, Y.M.; Zuki, A.B.Z.; Jimoh, A.A. Extraction and Physicochemical Characterization of Chitin and Chitosan Isolated from House Cricket. Biomed. Mater. 2018, 13, 025009. [Google Scholar] [CrossRef]
- Kaya, M.; Lelešius, E.; Nagrockaitė, R.; Sargin, I.; Arslan, G.; Mol, A.; Baran, T.; Can, E.; Bitim, B. Differentiations of Chitin Content and Surface Morphologies of Chitins Extracted from Male and Female Grasshopper Species. PLoS ONE 2015, 10, e0115531. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Baran, T.; Asan-Ozusaglam, M.; Cakmak, Y.S.; Tozak, K.O.; Mol, A.; Mentes, A.; Sezen, G. Extraction and Characterization of Chitin and Chitosan with Antimicrobial and Antioxidant Activities from Cosmopolitan Orthoptera Species (Insecta). Biotechnol. Bioprocess Eng. 2015, 20, 168–179. [Google Scholar] [CrossRef]
- Kaya, M.; Erdogan, S.; Mol, A.; Baran, T. Comparison of Chitin Structures Isolated from Seven Orthoptera Species. Int. J. Biol. Macromol. 2015, 72, 797–805. [Google Scholar] [CrossRef]
- Kim, M.-W.; Song, Y.-S.; Seo, D.-J.; Han, Y.S.; Jo, Y.H.; Noh, M.Y.; Yang, Y.C.; Park, Y.-K.; Kim, S.-A.; Choi, C.; et al. Extraction of Chitin and Chitosan from the Exoskeleton of the Cockroach (Periplaneta americana L.). J. Chitin Chitosan 2017, 22, 76–81. [Google Scholar] [CrossRef]
- Chae, K.-S.; Shin, C.-S.; Shin, W.-S. Characteristics of Cricket (Gryllus bimaculatus) Chitosan and Chitosan-Based Nanoparticles. Food Sci. Biotechnol. 2018, 27, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Song, Y.; Han, Y.S.; Jo, Y.H.; Choi, M.H.; Park, Y.; Kang, S.H.; Kim, S.; Choi, C.; Jung, W. Production of Chitin and Chitosan from the Exoskeleton of Adult Two-spotted Field Crickets (Gryllus bimaculatus). Entomol. Res. 2017, 47, 279–285. [Google Scholar] [CrossRef]
- Psarianos, M.; Ojha, S.; Schneider, R.; Schlüter, O.K. Chitin Isolation and Chitosan Production from House Crickets (Acheta domesticus) by Environmentally Friendly Methods. Molecules 2022, 27, 5005. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Liceaga, A.M.; San Martin-Gonzalez, F.; Jones, O.G.; Garcia-Bravo, J.M.; Kaplan, I. Development of Chitosan Films from Edible Crickets and Their Performance as a Bio-Based Food Packaging Material. Polysaccharides 2021, 2, 744–758. [Google Scholar] [CrossRef]
- Psarianos, M.; Dimopoulos, G.; Ojha, S.; Cavini, A.C.M.; Bußler, S.; Taoukis, P.; Schlüter, O.K. Effect of Pulsed Electric Fields on Cricket (Acheta Domesticus) Flour: Extraction Yield (Protein, Fat and Chitin) and Techno-Functional Properties. Innov. Food Sci. Emerg. Technol. 2022, 76, 102908. [Google Scholar] [CrossRef]
- Malm, M.; Liceaga, A.M. Physicochemical Properties of Chitosan from Two Commonly Reared Edible Cricket Species, and Its Application as a Hypolipidemic and Antimicrobial Agent. Polysaccharides 2021, 2, 339–353. [Google Scholar] [CrossRef]
- Kaya, M.; Mujtaba, M.; Bulut, E.; Akyuz, B.; Zelencova, L.; Sofi, K. Fluctuation in Physicochemical Properties of Chitins Extracted from Different Body Parts of Honeybee. Carbohydr. Polym. 2015, 132, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Tsaneva, D.; Petkova, Z.; Petkova, N.; Stoyanova, M.; Stoyanova, A.; Denev, P. Isolation and Characterization of Chitin and Biologically Active Substances from Honeybee (Apis mellifera). J. Pharm. Sci. 2018, 10, 884–888. [Google Scholar]
- Kovaleva, E.; Pestov, A.; Stepanova, D.; Molochnikov, L. Characterization of Chitin and Its Complexes Extracted from Natural Raw Sources. AIP Conf. Proc. 2016, 1772, 050007. [Google Scholar]
- Kaya, M.; Bağrıaçık, N.; Seyyar, O.; Baran, T. Comparison of Chitin Structures Derived from Three Common Wasp Species (Vespa Crabro LINNAEUS, 1758, Vespa Orientalis LINNAEUS, 1771 and Vespula Germanica (FABRICIUS, 1793)). Arch. Insect Biochem. Physiol. 2015, 89, 204–217. [Google Scholar] [CrossRef]
- Feás, X.; Vázquez-Tato, M.P.; Seijas, J.A.; Pratima, G.; Nikalje, A.; Fraga-López, F. Extraction and Physicochemical Characterization of Chitin Derived from the Asian Hornet, Vespa Velutina Lepeletier 1836 (Hym.: Vespidae). Molecules 2020, 25, 384. [Google Scholar] [CrossRef] [PubMed]
- Essa, E.E.; Hamza, D.; Khalil, M.M.H.; Zaher, H.; Salah, D.; Alnemari, A.M.; Rady, M.H.; Mo`men, S.A.A. The Antibacterial Activity of Egyptian Wasp Chitosan-Based Nanoparticles against Important Antibiotic-Resistant Pathogens. Molecules 2022, 27, 7189. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Sofi, K.; Sargin, I.; Mujtaba, M. Changes in Physicochemical Properties of Chitin at Developmental Stages (Larvae, Pupa and Adult) of Vespa Crabro (Wasp). Carbohydr. Polym. 2016, 145, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Soetemans, L.; Uyttebroek, M.; Bastiaens, L. Characteristics of Chitin Extracted from Black Soldier Fly in Different Life Stages. Int. J. Biol. Macromol. 2020, 165, 3206–3214. [Google Scholar] [CrossRef]
- Waśko, A.; Bulak, P.; Polak-Berecka, M.; Nowak, K.; Polakowski, C.; Bieganowski, A. The First Report of the Physicochemical Structure of Chitin Isolated from Hermetia illucens. Int. J. Biol. Macromol. 2016, 92, 316–320. [Google Scholar] [CrossRef]
- Purkayastha, D.; Sarkar, S. Physicochemical Structure Analysis of Chitin Extracted from Pupa Exuviae and Dead Imago of Wild Black Soldier Fly (Hermetia illucens). J. Polym. Environ. 2020, 28, 445–457. [Google Scholar] [CrossRef]
- D’Hondt, E.; Soetemans, L.; Bastiaens, L.; Maesen, M.; Jespers, V.; Van Den Bosch, B.; Voorspoels, S.; Elst, K. Simplified Determination of the Content and Average Degree of Acetylation of Chitin in Crude Black Soldier Fly Larvae Samples. Carbohydr. Res. 2020, 488, 107899. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Rehman, K.U.; Feng, W.; Yang, D.; Rehman, R.U.; Cai, M.; Zhang, J.; Yu, Z.; Zheng, L. Physicochemical Structure of Chitin in the Developing Stages of Black Soldier Fly. Int. J. Biol. Macromol. 2020, 149, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Antonov, A.; Ivanov, G.; Pastukhova, N.; Bovykina, G. Production of Chitin from Dead Hermetia illucens. IOP Conf. Ser. Earth Environ. Sci. 2019, 315, 042003. [Google Scholar] [CrossRef]
- Khayrova, A.; Lopatin, S.; Varlamov, V. Black Soldier Fly Hermetia illucens as a Novel Source of Chitin and Chitosan. Int. J. Sci. 2019, 8, 81–86. [Google Scholar] [CrossRef]
- Caligiani, A.; Marseglia, A.; Leni, G.; Baldassarre, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of Black Soldier Fly Prepupae and Systematic Approaches for Extraction and Fractionation of Proteins, Lipids and Chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Teo, H.P.; Law, K.W.; Eric Chan, W.C.; Michelle Soo, O.Y. Antibacterial Properties of Chitosan Isolated from the Black Soldier Fly, Hermetia illucens. Sains Malays. 2022, 51, 3923–3935. [Google Scholar] [CrossRef]
- Lagat, M.K.; Were, S.; Ndwigah, F.; Kemboi, V.J.; Kipkoech, C.; Tanga, C.M. Antimicrobial Activity of Chemically and Biologically Treated Chitosan Prepared from Black Soldier Fly (Hermetia illucens) Pupal Shell Waste. Microorganisms 2021, 9, 2417. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, A.; Triunfo, M.; Scieuzo, C.; Ianniciello, D.; Tafi, E.; Hahn, T.; Zibek, S.; Salvia, R.; De Bonis, A.; Falabella, P. Antimicrobial Properties of Chitosan from Different Developmental Stages of the Bioconverter Insect Hermetia illucens. Sci. Rep. 2022, 12, 8084. [Google Scholar] [CrossRef]
- Kemboi, V.; Kipkoech, C.; Njire, M.; Were, S.; Lagat, M.; Ndwiga, F.; Wesonga, J.; Tanga, C. Biocontrol Potential of Chitin and Chitosan Extracted from Black Soldier Fly Pupal Exuviae against Bacterial Wilt of Tomato. Microorganisms 2022, 10, 165. [Google Scholar] [CrossRef]
- Lagat, M.K. Biological and Chemical Extraction of Chitin and Chitosan from the Black Soldier Fly (Hermetia illucens) Exoskeleton and Antimicrobial Activity Against Selected Human Pathogenic Microbes. Master’s Thesis, Jomo Kenyatta University of Agriculture and Technology, Juja, Kenya, 2022. [Google Scholar]
- Triunfo, M.; Tafi, E.; Guarnieri, A.; Salvia, R.; Scieuzo, C.; Hahn, T.; Zibek, S.; Gagliardini, A.; Panariello, L.; Coltelli, M.B.; et al. Characterization of Chitin and Chitosan Derived from Hermetia illucens, a Further Step in a Circular Economy Process. Sci. Rep. 2022, 12, 6613. [Google Scholar] [CrossRef] [PubMed]
- Maria-Beatrice, C.; Luca, P.; Alessandro, V.; Vito, G.; Alessandro, G.; Pierfrancesco, M.; Patrizia, C.; Andrea, L.; Angela, D.B.; Patrizia, F. Chitin and Its Derivatives: Nanostructured Materials from Different Marine and Terrestrial Sources. Chem. Eng. Trans. 2022, 93, 295–300. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, S.C.; Nam, K.D.; Kim, T.H.; Jung, B.O.; Park, Y.-I.; Synytsya, A.; Park, J.K. Chitosan Isolated from Black Soldier Flies Hermetia illucens: Structure and Enzymatic Hydrolysis. Process Biochem. 2022, 118, 171–181. [Google Scholar] [CrossRef]
- Khayrova, A.; Lopatin, S.; Shagdarova, B.; Sinitsyna, O.; Sinitsyn, A.; Varlamov, V. Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan. Molecules 2022, 27, 577. [Google Scholar] [CrossRef] [PubMed]
- Złotko, K.; Waśko, A.; Kamiński, D.M.; Budziak-Wieczorek, I.; Bulak, P.; Bieganowski, A. Isolation of Chitin from Black Soldier Fly (Hermetia illucens) and Its Usage to Metal Sorption. Polymers 2021, 13, 818. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Akyuz, B.; Bulut, E.; Sargin, I.; Eroglu, F.; Tan, G. Chitosan Nanofiber Production from Drosophila by Electrospinning. Int. J. Biol. Macromol. 2016, 92, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Han, Y.S.; Jo, Y.H.; Choi, M.H.; Kang, S.H.; Kim, S.; Jung, W. Extraction of Chitin and Chitosan from Housefly, Musca Domestica, Pupa Shells. Entomol. Res. 2016, 46, 324–328. [Google Scholar] [CrossRef]
- İlk, S.; Ramanauskaitė, A.; Koç Bilican, B.; Mulerčikas, P.; Çam, D.; Onses, M.S.; Torun, I.; Kazlauskaitė, S.; Baublys, V.; Aydın, Ö.; et al. Usage of Natural Chitosan Membrane Obtained from Insect Corneal Lenses as a Drug Carrier and Its Potential for Point of Care Tests. Mater. Sci. Eng. C 2020, 112, 110897. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Mentes, A.; Asaroglu, M.; Sezen, G.; Tozak, K.O. Extraction and Characterization of α-Chitin and Chitosan from Six Different Aquatic Invertebrates. Food Biophys. 2014, 9, 145–157. [Google Scholar] [CrossRef]
- Mol, A.; Kaya, M.; Mujtaba, M.; Akyuz, B. Extraction of High Thermally Stable and Nanofibrous Chitin from Cicada (Cicadoidea). Entomol. Res. 2018, 48, 480–489. [Google Scholar] [CrossRef]
- Sharbidre, A.; Sargar, S.; Gogoi, H.; Patil, R. Characterization of Chitin Content Extracted from Edible Insect, Coridius Nepalensis (Westwood, 1837) (Hemiptera: Dinidoridae). Int. J. Trop. Insect Sci. 2021, 41, 1893–1900. [Google Scholar] [CrossRef]
- Jiang, F.; Li, X.; Duan, Y.; Li, Q.; Qu, Y.; Zhong, G.; Qiu, M.; Zhang, J.; Zhang, C.; Pan, X. Extraction and Characterization of Chitosan from Eupolyphaga Sinensis Walker and Its Application in the Preparation of Electrospinning Nanofiber Membranes. Colloids Surf. B Biointerfaces 2023, 222, 113030. [Google Scholar] [CrossRef]
- Kaya, M.; Sargin, I.; Sabeckis, I.; Noreikaite, D.; Erdonmez, D.; Salaberria, A.M.; Labidi, J.; Baublys, V.; Tubelytė, V. Biological, Mechanical, Optical and Physicochemical Properties of Natural Chitin Films Obtained from the Dorsal Pronotum and the Wing of Cockroach. Carbohydr. Polym. 2017, 163, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Basseri, H.; Bakhtiyari, R.; Hashemi, S.J.; Baniardelani, M.; Shahraki, H.; Hosainpour, L. Antibacterial/Antifungal Activity of Extracted Chitosan From American Cockroach (Dictyoptera: Blattidae) and German Cockroach (Blattodea: Blattellidae). J. Med. Entomol. 2019, 56, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Wanule, D.; Balkhande, J.V.; Ratnakar, P.U.; Kulkarni, A.N.; Bhowate, C.S. Extraction and FTIR Analysis of Chitosan from American Cockroach, Periplaneta americana. Int. J. Eng. Sci. Innov. Technol. 2014, 3, 299–304. [Google Scholar]
- Kaya, M.; Baran, T. Description of a New Surface Morphology for Chitin Extracted from Wings of Cockroach (Periplaneta americana). Int. J. Biol. Macromol. 2015, 75, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wei, X.; Sui, Z.; Guo, M.; Geng, J.; Xiao, J.; Huang, D. Preparation of Antioxidant and Antibacterial Chitosan Film from Periplaneta americana. Insects 2021, 12, 53. [Google Scholar] [CrossRef]
- Mahmoud, T.M.; Mostafa, I.H.; Ahmed, S.B.; Aly, F.M.; Mohammad, R.K.A.-S. Antibacterial and Antiviral Activities of Chitosan Nanoparticles from the American cockroach, Periplaneta americana. J. Appl. Pharm. Sci. 2022, 12, 202–208. [Google Scholar] [CrossRef]
- Kamal, M.; Adly, E.; Alharbi, S.A.; Khaled, A.S.; Rady, M.H.; Ibrahim, N.A. Exploring Simplified Methods for Insect Chitin Extraction and Application as a Potential Alternative Bioethanol Resource. Insects 2020, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Sargin, I.; Al-jaf, I.; Erdogan, S.; Arslan, G. Characteristics of Corneal Lens Chitin in Dragonfly Compound Eyes. Int. J. Biol. Macromol. 2016, 89, 54–61. [Google Scholar] [CrossRef]
- Tan, G.; Kaya, M.; Tevlek, A.; Sargin, I.; Baran, T. Antitumor Activity of Chitosan from Mayfly with Comparison to Commercially Available Low, Medium and High Molecular Weight Chitosans. Vitro Cell. Dev. Biol.-Anim. 2018, 54, 366–374. [Google Scholar] [CrossRef]
- Jantzen Da Silva Lucas, A.; Quadro Oreste, E.; Leão Gouveia Costa, H.; Martín López, H.; Dias Medeiros Saad, C.; Prentice, C. Extraction, Physicochemical Characterization, and Morphological Properties of Chitin and Chitosan from Cuticles of Edible Insects. Food Chem. 2021, 343, 128550. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Baran, T.; Erdoğan, S.; Menteş, A.; Aşan Özüsağlam, M.; Çakmak, Y.S. Physicochemical Comparison of Chitin and Chitosan Obtained from Larvae and Adult Colorado Potato Beetle (Leptinotarsa decemlineata). Mater. Sci. Eng. C 2014, 45, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Umar, Z.G.; Abalaka, M.E.; Daniyan, S.Y.; Babayi, H.; Adeniyi, K.A. Physicochemical and Bio-Metabolite Characterizations of Chitosan Isolated from American Cockroach (Periplaneta americana) and Cricket (Acheta domesticus). BIOMED Nat. Appl. Sci. 2022, 02, 25–36. [Google Scholar] [CrossRef]
- Song, C.; Yu, H.; Zhang, M.; Yang, Y.; Zhang, G. Physicochemical Properties and Antioxidant Activity of Chitosan from the Blowfly Chrysomya megacephala Larvae. Int. J. Biol. Macromol. 2013, 60, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Cheraghi, E.; Kababian, M.; Moradi-Asl, E.; Mousavi Bafrouyi, S.M.; Saghafipour, A. Structure and Antibacterial Activity of Chitosan from the American Cock-Roach, the German Cockroach and the Mealworm Beetle. J. Arthropod-Borne Dis. 2023, 16, 325–339. [Google Scholar] [CrossRef]
- Song, Y.; Jo, Y.H.; Han, Y.S.; Jung, W. Production of Chitin- and Chitosan-oligosaccharide Using the Edible Insect, Tenebrio molitor. Entomol. Res. 2022, 52, 207–213. [Google Scholar] [CrossRef]
- Kaya, M.; Salaberria, A.M.; Mujtaba, M.; Labidi, J.; Baran, T.; Mulercikas, P.; Duman, F. An Inclusive Physicochemical Comparison of Natural and Synthetic Chitin Films. Int. J. Biol. Macromol. 2018, 106, 1062–1070. [Google Scholar] [CrossRef]
- Hahn, T.; Tafi, E.; Von Seggern, N.; Falabella, P.; Salvia, R.; Thomä, J.; Febel, E.; Fijalkowska, M.; Schmitt, E.; Stegbauer, L.; et al. Purification of Chitin from Pupal Exuviae of the Black Soldier Fly. Waste Biomass Valorization 2022, 13, 1993–2008. [Google Scholar] [CrossRef]
- Chalghaf, M.; Charradi, K.; Ksouri, R.; Alsulami, Q.A.; Jaouani, A.; Keshk, S.M.A.S.; Hayouni, E.A. Physicochemical Characterization of Chitin Extracted by Different Treatment Sequences from an Edible Insect. Int. J. Biol. Macromol. 2023, 253, 127156. [Google Scholar] [CrossRef]
- Goularte, N.F.; Kallem, T.; Cegelski, L. Chemical and Molecular Composition of the Chrysalis Reveals Common Chitin-Rich Structural Framework for Monarchs and Swallowtails. J. Mol. Biol. 2022, 434, 167456. [Google Scholar] [CrossRef] [PubMed]
- Greven, H.; Kaya, M.; Junker, K.; Akyuz, L.; Amemiya, C.T. Characterization of Tongue Worm (Pentastomida) Chitin Supports α- Rather than β-Chitin. Zool. Anz. 2019, 279, 111–115. [Google Scholar] [CrossRef]
- Nafisah, A.; Nahrowi; Mutia, R.; Jayanegara, A. Chemical Composition, Chitin and Cell Wall Nitrogen Content of Black Soldier Fly (Hermetia illucens) Larvae after Physical and Biological Treatment. IOP Conf. Ser. Mater. Sci. Eng. 2019, 546, 042028. [Google Scholar] [CrossRef]
- Srinivasan, H.; Kanayairam, V.; Ravichandran, R. Chitin and Chitosan Preparation from Shrimp Shells Penaeus Monodon and Its Human Ovarian Cancer Cell Line, PA-1. Int. J. Biol. Macromol. 2018, 107, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Synowiecki, J.; Al-Khateeb, N.A. Production, Properties, and Some New Applications of Chitin and Its Derivatives. Crit. Rev. Food Sci. Nutr. 2003, 43, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.N.; Chin, Y.L.; Chen, W.N. Comparison of Sustainable Lipid and Protein Removal Methods for the Isolation of Insect Chitin from Black Soldier Fly Exoskeleton. ACS Food Sci. Technol. 2021, 1, 698–706. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Liang, S.-H.; Lai, W.-L.; Lee, J.-X.; Wang, Y.-P.; Liu, Y.-T.; Wang, S.-H.; Lee, M.-H. Sustainable Extraction of Chitin from Spent Pupal Shell of Black Soldier Fly. Processes 2021, 9, 976. [Google Scholar] [CrossRef]
- Egorova, K.S.; Gordeev, E.G.; Ananikov, V.P. Biological Activity of Ionic Liquids and Their Application in Pharmaceutics and Medicine. Chem. Rev. 2017, 117, 7132–7189. [Google Scholar] [CrossRef]
- Shamshina, J.L.; Berton, P.; Rogers, R.D. Advances in Functional Chitin Materials: A Review. ACS Sustain. Chem. Eng. 2019, 7, 6444–6457. [Google Scholar] [CrossRef]
- Morais, E.S.; Lopes, A.M.D.C.; Freire, M.G.; Freire, C.S.R.; Coutinho, J.A.P.; Silvestre, A.J.D. Use of Ionic Liquids and Deep Eutectic Solvents in Polysaccharides Dissolution and Extraction Processes towards Sustainable Biomass Valorization. Molecules 2020, 25, 3652. [Google Scholar] [CrossRef]
- Li, Z.; Liu, C.; Hong, S.; Lian, H.; Mei, C.; Lee, J.; Wu, Q.; Hubbe, M.A.; Li, M.-C. Recent Advances in Extraction and Processing of Chitin Using Deep Eutectic Solvents. Chem. Eng. J. 2022, 446, 136953. [Google Scholar] [CrossRef]
- Huet, G.; Hadad, C.; González-Domínguez, J.M.; Courty, M.; Jamali, A.; Cailleu, D.; Van Nhien, A.N. IL versus DES: Impact on Chitin Pretreatment to Afford High Quality and Highly Functionalizable Chitosan. Carbohydr. Polym. 2021, 269, 118332. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, M.-C.; Liu, C.; Liu, X.; Lu, Y.; Zhou, G.; Liu, C.; Mei, C. Microwave-Assisted Deep Eutectic Solvent Extraction of Chitin from Crayfish Shell Wastes for 3D Printable Inks. Ind. Crops Prod. 2023, 194, 116325. [Google Scholar] [CrossRef]
- Marmier, T.; Szczepanski, C.R.; Candet, C.; Zenerino, A.; Godeau, R.-P.; Godeau, G. Investigation on Mecynorhina Torquata Drury, 1782 (Coleoptera, Cetoniidae, Goliathini) Cuticle: Surface Properties, Chitin and Chitosan Extraction. Int. J. Biol. Macromol. 2020, 164, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Godeau, X.Y.; Andrianandrasana, F.J.; Volkova, O.; Szczepanski, C.R.; Zenerino, A.; Montreuil, O.; Godeau, R.-P.; Kuzhir, P.; Godeau, G. Investigation on Dung Beetle’s (Heliocopris Hope, 1838) Chitosan Valorisation for Hydrogel 3D Printing. Int. J. Biol. Macromol. 2022, 199, 172–180. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.M.; Franco, T.T.; Oliveira Junior, E.N.D. Physicochemical Characterization of Thermally Treated Chitosans and Chitosans Obtained by Alkaline Deacetylation. Int. J. Polym. Sci. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Gzyra-Jagieła, K.; Pęczek, B.; Wiśniewska-Wrona, M.; Gutowska, N. Physicochemical Properties of Chitosan and Its Degradation Products. In Chitin and Chitosan; Broek, L.A.M., Boeriu, C.G., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 61–80. ISBN 978-1-119-45043-6. [Google Scholar]
- Abdel-Rahman, R.M.; Hrdina, R.; Abdel-Mohsen, A.M.; Fouda, M.M.G.; Soliman, A.Y.; Mohamed, F.K.; Mohsin, K.; Pinto, T.D. Chitin and Chitosan from Brazilian Atlantic Coast: Isolation, Characterization and Antibacterial Activity. Int. J. Biol. Macromol. 2015, 80, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Sedaghat, F.; Yousefzadi, M.; Toiserkani, H.; Najafipour, S. Chitin from Penaeus Merguiensis via Microbial Fermentation Processing and Antioxidant Activity. Int. J. Biol. Macromol. 2016, 82, 279–283. [Google Scholar] [CrossRef]
- Qiao, C.; Ma, X.; Wang, X.; Liu, L. Structure and Properties of Chitosan Films: Effect of the Type of Solvent Acid. LWT 2021, 135, 109984. [Google Scholar] [CrossRef]
- Kapadnis, G.; Dey, A.; Dandekar, P.; Jain, R. Effect of Degree of Deacetylation on Solubility of Low-molecular-weight Chitosan Produced via Enzymatic Breakdown of Chitosan. Polym. Int. 2019, 68, 1054–1063. [Google Scholar] [CrossRef]
- Roy, J.C.; Salaün, F.; Giraud, S.; Ferri, A.; Chen, G.; Guan, J. Solubility of Chitin: Solvents, Solution Behaviors and Their Related Mechanisms. In Solubility of Polysaccharides; Xu, Z., Ed.; InTech: Houston, TX, USA, 2017; ISBN 978-953-51-3649-1. [Google Scholar]
- Wang, W.; Meng, Q.; Li, Q.; Liu, J.; Zhou, M.; Jin, Z.; Zhao, K. Chitosan Derivatives and Their Application in Biomedicine. Int. J. Mol. Sci. 2020, 21, 487. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Gulbake, A.; Shilpi, S.; Jain, A.; Hurkat, P.; Jain, S.K. A New Horizon in Modifications of Chitosan: Syntheses and Applications. Crit. Rev. Ther. Drug Carrier Syst. 2013, 30, 91. [Google Scholar] [CrossRef] [PubMed]
- Chisty, A.H.; Masud, R.A.; Hasan, M.M.; Khan, M.N.; Mallik, A.K.; Rahman, M.M. PEGylated Chitin and Chitosan Derivatives. In Handbook of Chitin and Chitosan; Elsevier: Amsterdam, The Netherlands, 2020; pp. 59–100. ISBN 978-0-12-817970-3. [Google Scholar]
- Liu, T.G.; Wang, Y.T.; Li, B.; Deng, H.B.; Huang, Z.L.; Qian, L.W.; Wang, X. Urea Free Synthesis of Chitin-Based Acrylate Superabsorbent Polymers under Homogeneous Conditions: Effects of the Degree of Deacetylation and the Molecular Weight. Carbohydr. Polym. 2017, 174, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, R.; Prabaharan, M.; Sudheesh Kumar, P.T.; Nair, S.V.; Tamura, H. Biomaterials Based on Chitin and Chitosan in Wound Dressing Applications. Biotechnol. Adv. 2011, 29, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Jafari, H.; Pirouzifard, M.; Khaledabad, M.A.; Almasi, H. Effect of Chitin Nanofiber on the Morphological and Physical Properties of Chitosan/Silver Nanoparticle Bionanocomposite Films. Int. J. Biol. Macromol. 2016, 92, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Furuike, T.; Komoto, D.; Hashimoto, H.; Tamura, H. Preparation of Chitosan Hydrogel and Its Solubility in Organic Acids. Int. J. Biol. Macromol. 2017, 104, 1620–1625. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.; Szczepanski, C.R.; Godeau, R.-P.; Godeau, G. Chitosan Extraction from Goliathus Orientalis Moser, 1909: Characterization and Comparison with Commercially Available Chitosan. Biomimetics 2020, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Kasaai, M. A Review of Several Reported Procedures to Determine the Degree of N-Acetylation for Chitin and Chitosan Using Infrared Spectroscopy. Carbohydr. Polym. 2008, 71, 497–508. [Google Scholar] [CrossRef]
- Engwa, G.A. Free Radicals and the Role of Plant Phytochemicals as Antioxidants Against Oxidative Stress-Related Diseases. In Phytochemicals-Source of Antioxidants and Role in Disease Prevention; Asao, T., Asaduzzaman, M., Eds.; InTech: Houston, TX, USA, 2018; ISBN 978-1-78984-377-4. [Google Scholar]
- Shetty, A.K.; Kodali, M.; Upadhya, R.; Madhu, L.N. Emerging Anti-Aging Strategies-Scientific Basis and Efficacy. Aging Dis. 2018, 9, 1165. [Google Scholar] [CrossRef]
- Cui, X.; Lin, Q.; Liang, Y. Plant-Derived Antioxidants Protect the Nervous System from Aging by Inhibiting Oxidative Stress. Front. Aging Neurosci. 2020, 12, 209. [Google Scholar] [CrossRef]
- Tafi, E.; Triunfo, M.; Guarnieri, A.; Ianniciello, D.; Salvia, R.; Scieuzo, C.; Ranieri, A.; Castagna, A.; Lepuri, S.; Hahn, T.; et al. Preliminary investigation on the effect of insect-based chitosan on preservation of coated fresh cherry tomatoes. Sci. Rep. 2023, 13, 7030. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Su, Y.; Chen, C.; Jia, G.; Wang, H.; Wu, J.C.G.; Lin, J. Relationship between Antibacterial Activity of Chitosan and Surface Characteristics of Cell Wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar] [PubMed]
- Bhavsar, P.S.; Dalla Fontana, G.; Zoccola, M. Sustainable Superheated Water Hydrolysis of Black Soldier Fly Exuviae for Chitin Extraction and Use of the Obtained Chitosan in the Textile Field. ACS Omega 2021, 6, 8884–8893. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Sargin, I.; Mulerčikas, P.; Labidi, J.M.; Salaberria, A.S.; Cakmak, Y.; Kazlauskaitė, S.; Erdonmez, D.; Baublys, V. Conversion of Waste Parasitic Insect (Hylobius abietis L.) into Antioxidative, Antimicrobial and Biodegradable Films. J. Renew. Mater. 2019, 7, 215–226. [Google Scholar] [CrossRef]
- Mahboub, M.; Hassan, M.; Bream, A.; Mohamed, A.; Abdel-Samad, M. Preparation, Characterization and Anticancer Activity of Chitosan Prepared from the American cockroach, Periplaneta americana. Egypt. Acad. J. Biol. Sci. Entomol. 2021, 14, 163–171. [Google Scholar] [CrossRef]
- Bano, I.; Arshad, M.; Yasin, T.; Ghauri, M.A.; Younus, M. Chitosan: A Potential Biopolymer for Wound Management. Int. J. Biol. Macromol. 2017, 102, 380–383. [Google Scholar] [CrossRef]
- Marei, N.H.; El-Mazny, W.; El-Shaer, A.; Zaki, K.D.; Hussein, Z.S.; Abd-El-Samie, E.M. Enhanced Wound Healing Activity of Desert Locust (Schistocerca Gregaria) vs. Shrimp (Penaeus Monodon) Chitosan Based Scaffolds. Int. J. Biol. Macromol. 2017, 97, 23–33. [Google Scholar] [CrossRef]
- Jana, S.; Jana, S. (Eds.) Functional Chitosan: Drug Delivery and Biomedical Applications; Springer: Singapore, 2019; ISBN 9789811502620. [Google Scholar]
- Ahsan, S.M.; Thomas, M.; Reddy, K.K.; Sooraparaju, S.G.; Asthana, A.; Bhatnagar, I. Chitosan as Biomaterial in Drug Delivery and Tissue Engineering. Int. J. Biol. Macromol. 2018, 110, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Denkbas, E.B.; Ottenbrite, R.M. Perspectives on: Chitosan Drug Delivery Systems Based on Their Geometries. J. Bioact. Compat. Polym. 2006, 21, 351–368. [Google Scholar] [CrossRef]
- Głąb, M.; Kudłacik-Kramarczyk, S.; Drabczyk, A.; Guigou, M.D.; Sobczak-Kupiec, A.; Mierzwiński, D.; Gajda, P.; Walter, J.; Tyliszczak, B. Multistep Chemical Processing of Crickets Leading to the Extraction of Chitosan Used for Synthesis of Polymer Drug Carriers. Materials 2021, 14, 5070. [Google Scholar] [CrossRef]
- Albanna, M.Z.; Bou-Akl, T.H.; Blowytsky, O.; Walters, H.L.; Matthew, H.W.T. Chitosan Fibers with Improved Biological and Mechanical Properties for Tissue Engineering Applications. J. Mech. Behav. Biomed. Mater. 2013, 20, 217–226. [Google Scholar] [CrossRef] [PubMed]
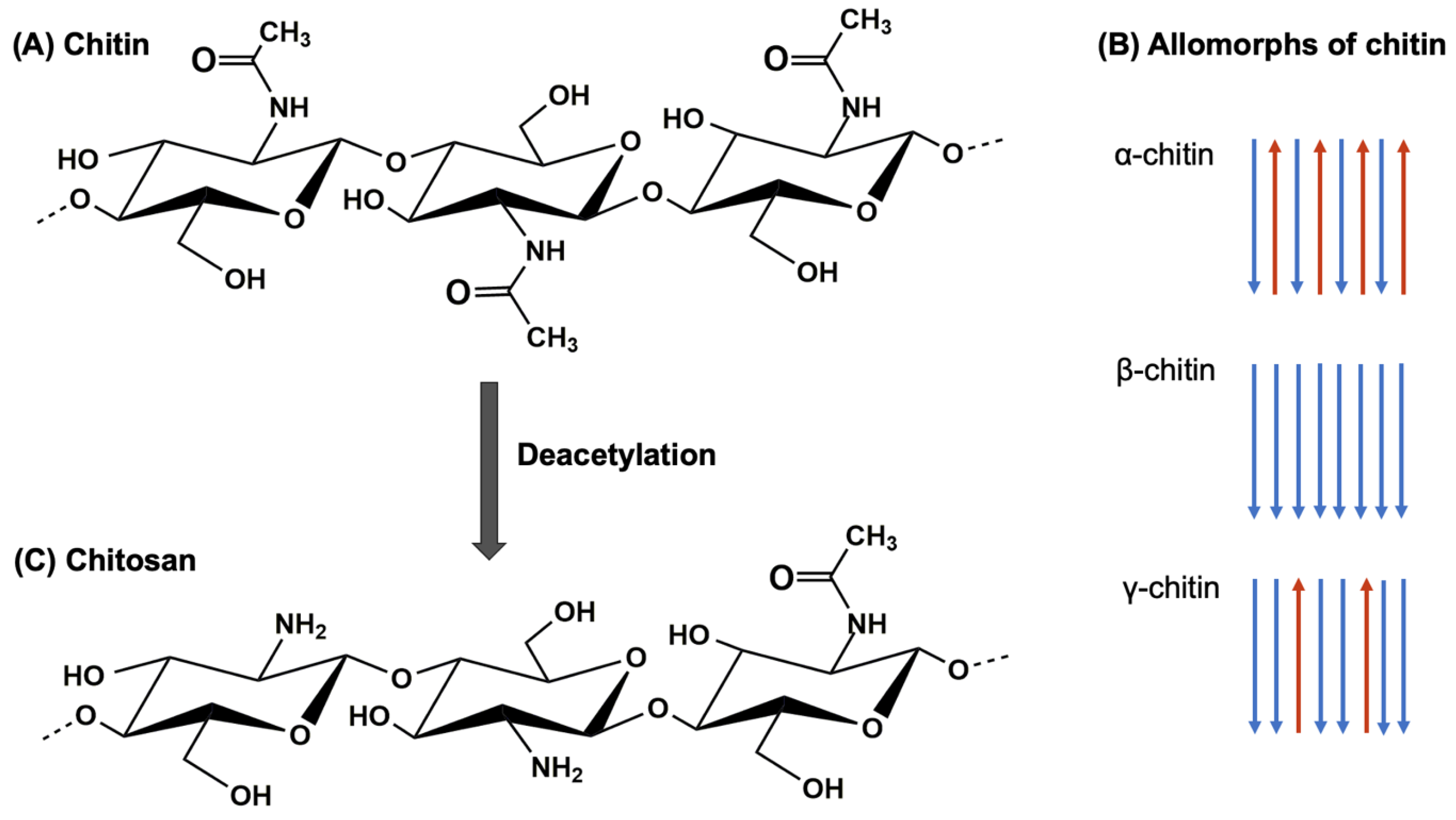
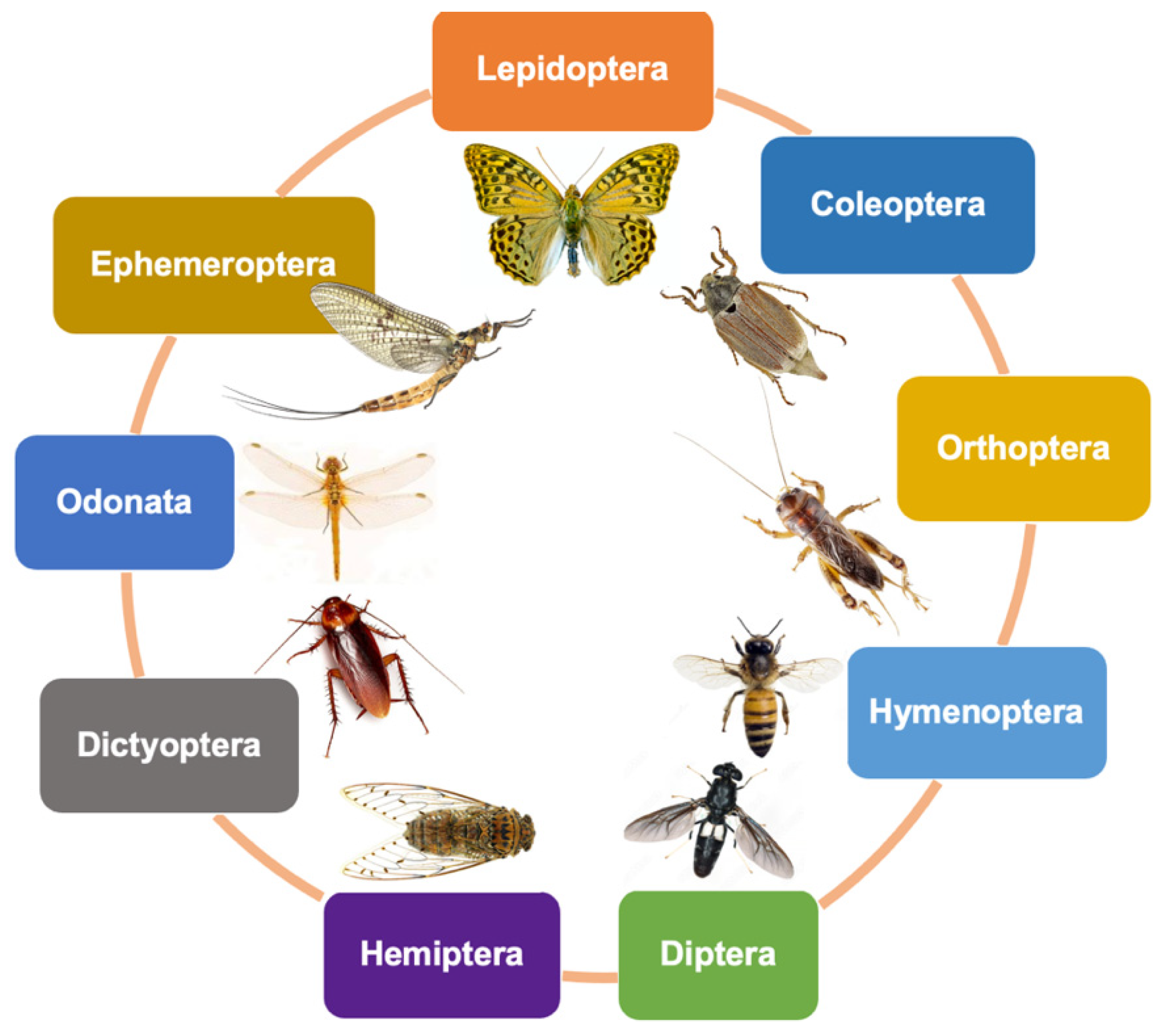
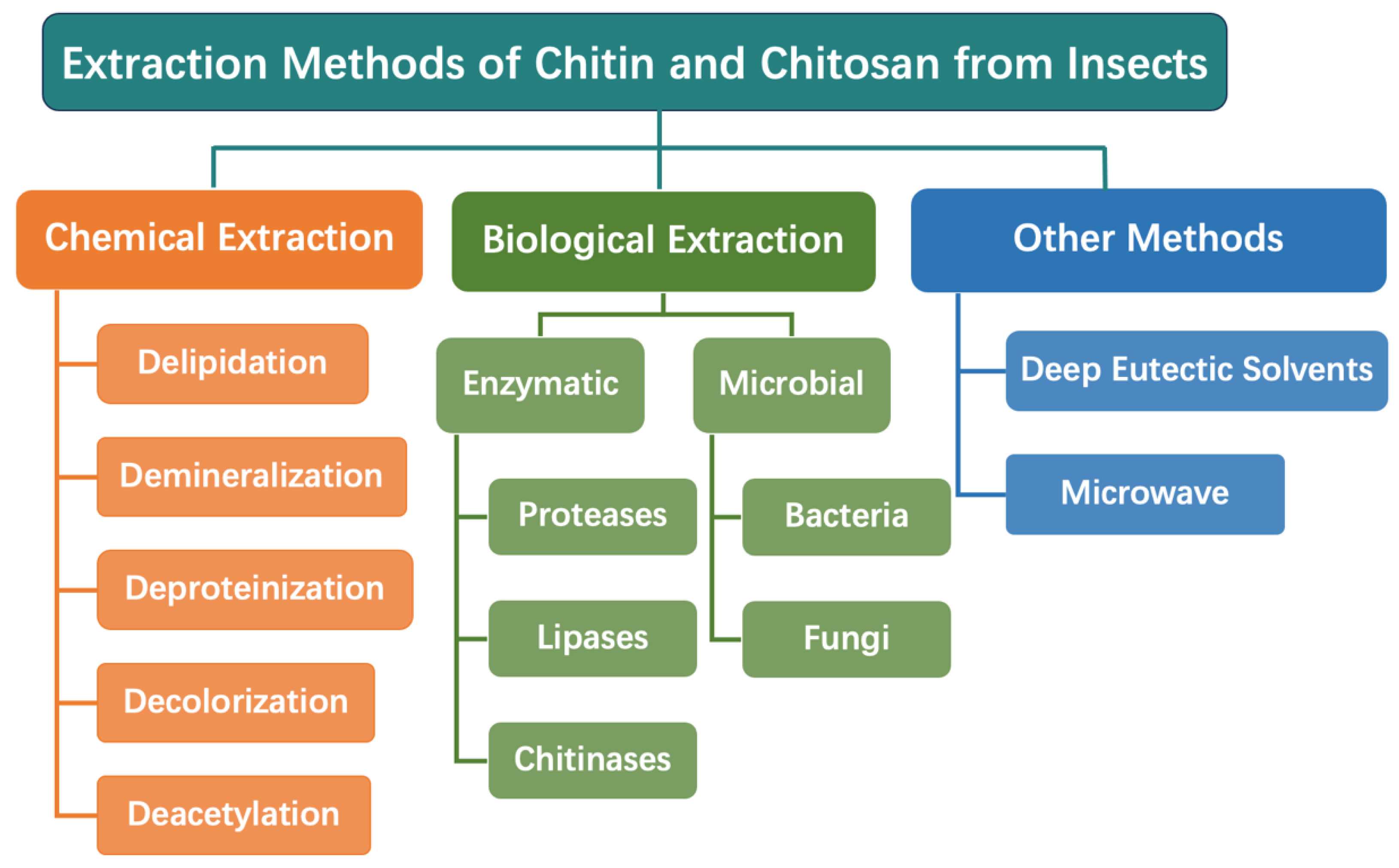
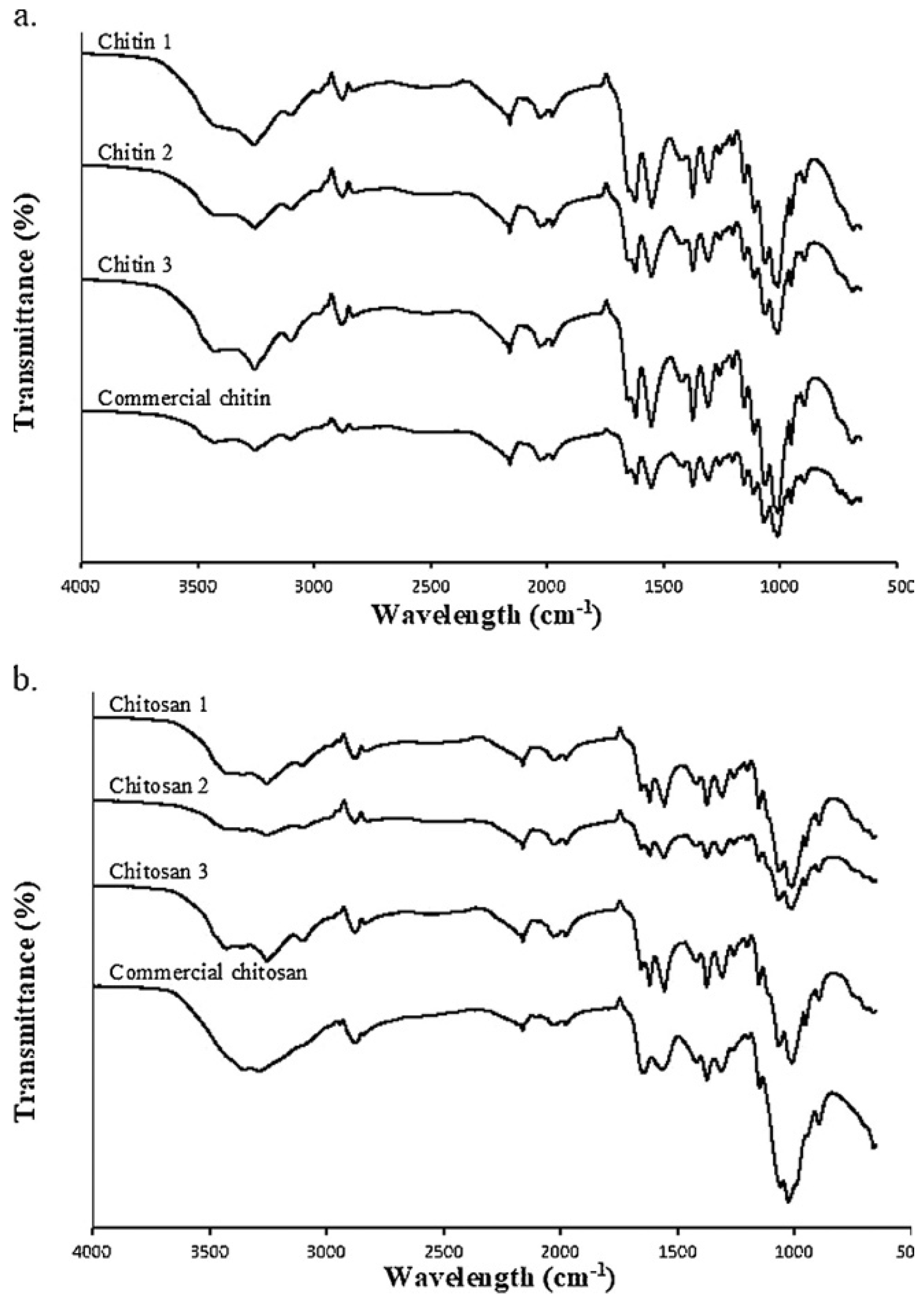
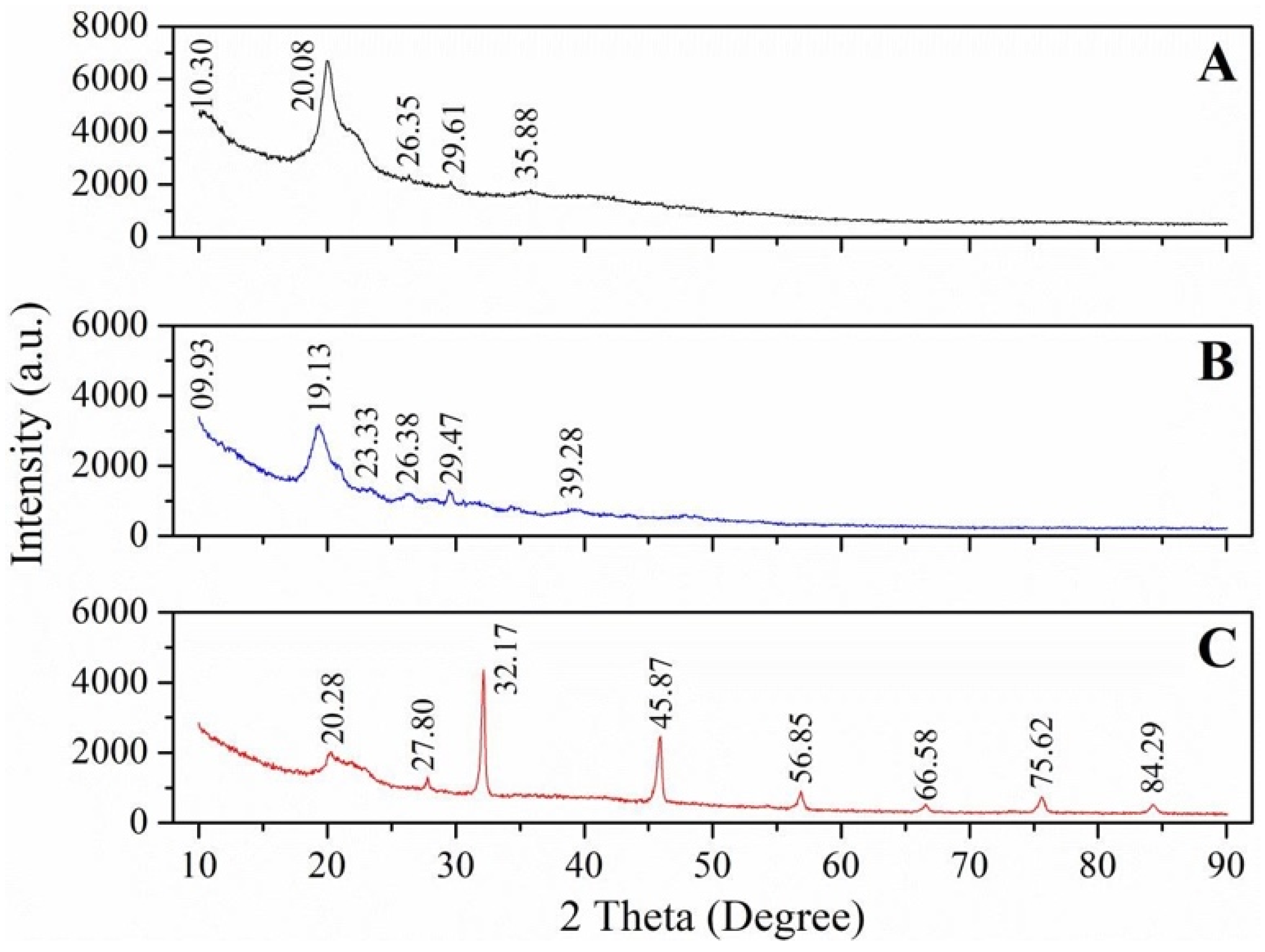
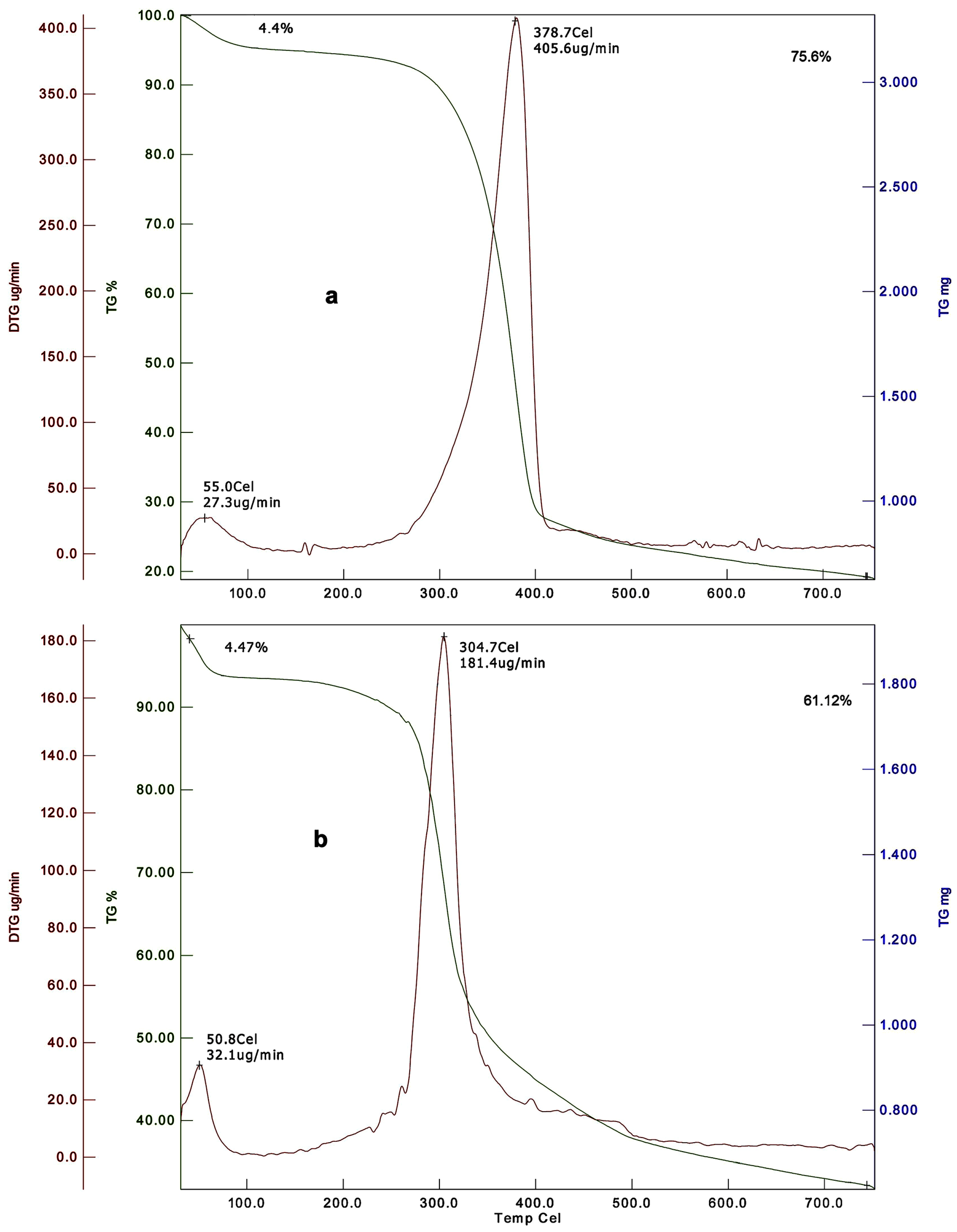
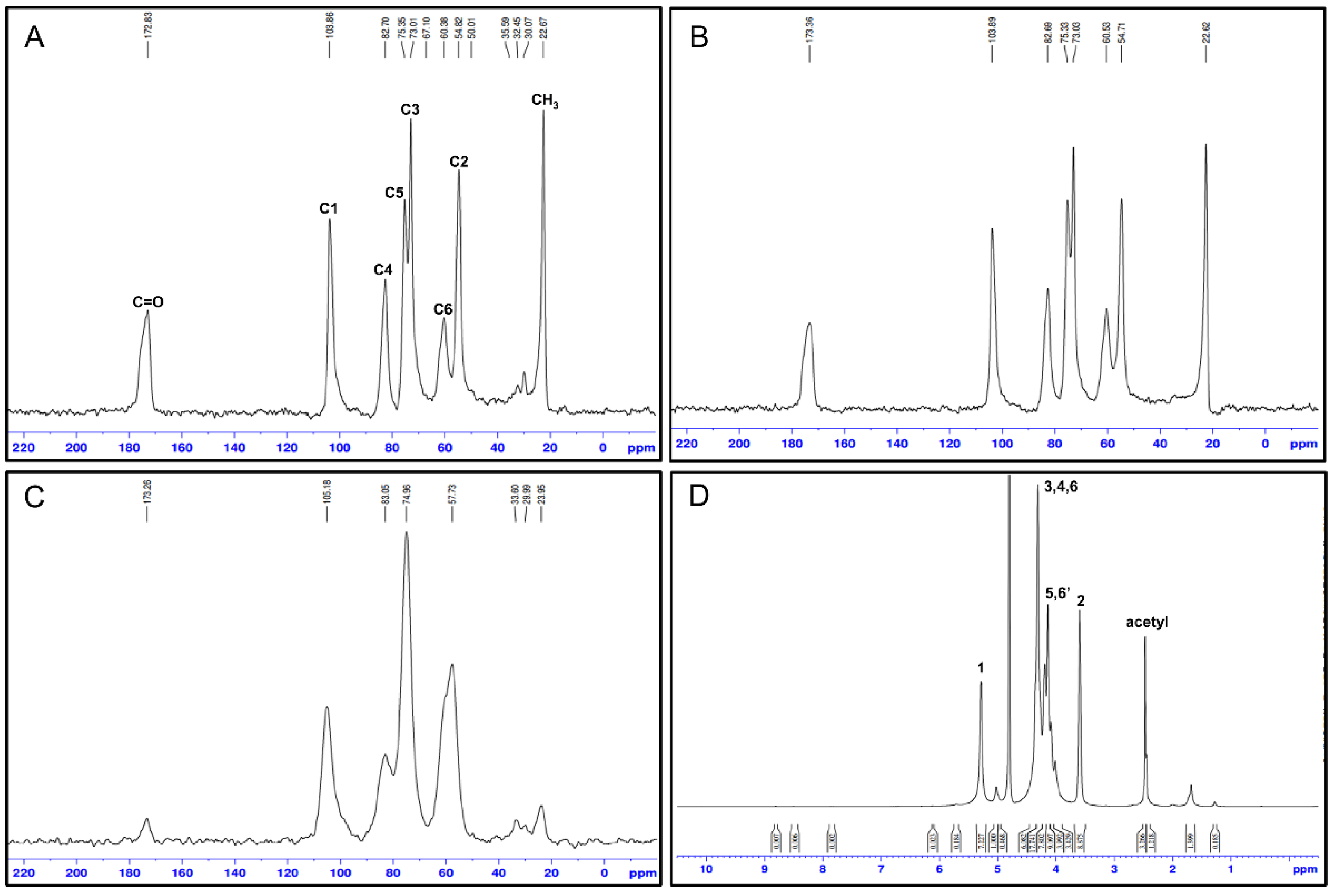

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, Z.; Kuzhir, P.; Godeau, G. Update on Chitin and Chitosan from Insects: Sources, Production, Characterization, and Biomedical Applications. Biomimetics 2024, 9, 297. https://doi.org/10.3390/biomimetics9050297
Mei Z, Kuzhir P, Godeau G. Update on Chitin and Chitosan from Insects: Sources, Production, Characterization, and Biomedical Applications. Biomimetics. 2024; 9(5):297. https://doi.org/10.3390/biomimetics9050297
Chicago/Turabian StyleMei, Zhenying, Pavel Kuzhir, and Guilhem Godeau. 2024. "Update on Chitin and Chitosan from Insects: Sources, Production, Characterization, and Biomedical Applications" Biomimetics 9, no. 5: 297. https://doi.org/10.3390/biomimetics9050297
APA StyleMei, Z., Kuzhir, P., & Godeau, G. (2024). Update on Chitin and Chitosan from Insects: Sources, Production, Characterization, and Biomedical Applications. Biomimetics, 9(5), 297. https://doi.org/10.3390/biomimetics9050297