
Biofeedback-Based Closed-Loop Phytoactuation in Vertical Farming and Controlled-Environment Agriculture


Abstract
1. Introduction
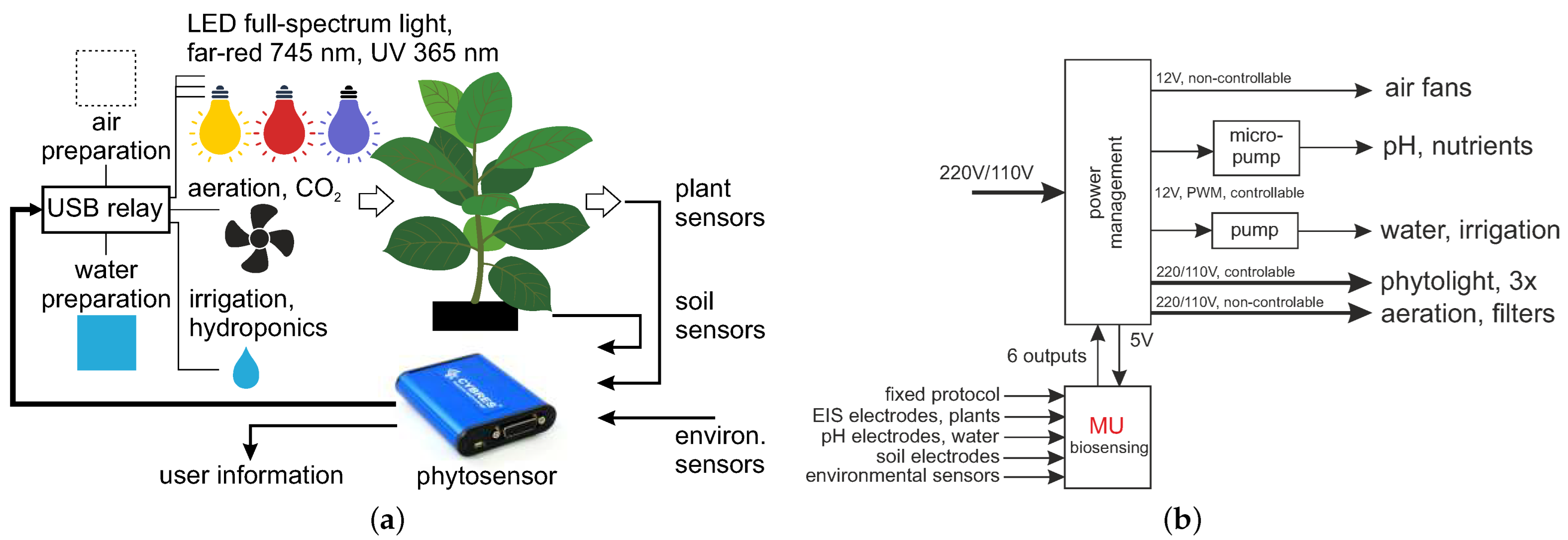
2. Methods and Setup
3. Measurements
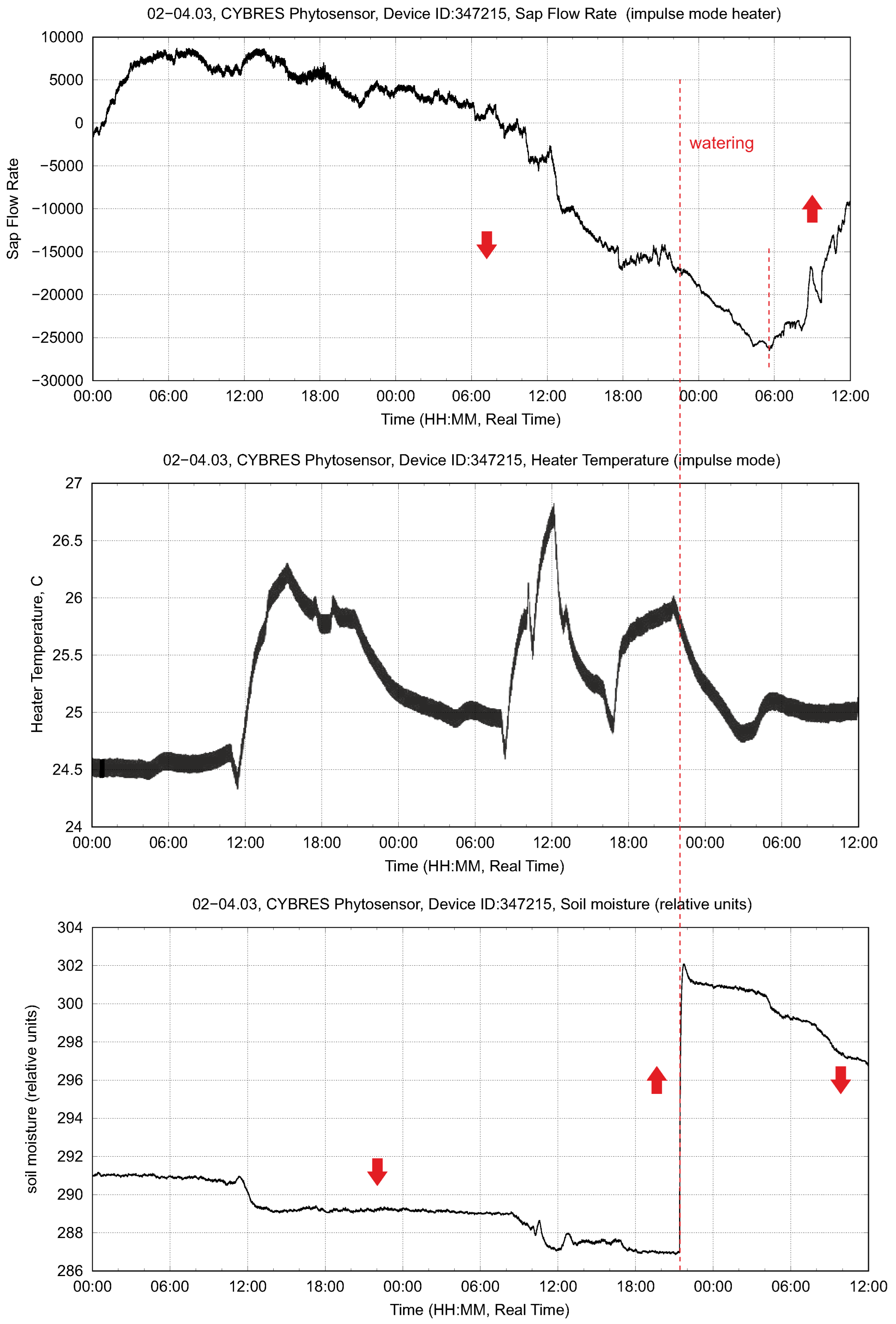
3.1. Heat-Based Sap Sensors for Long-Term Measurements
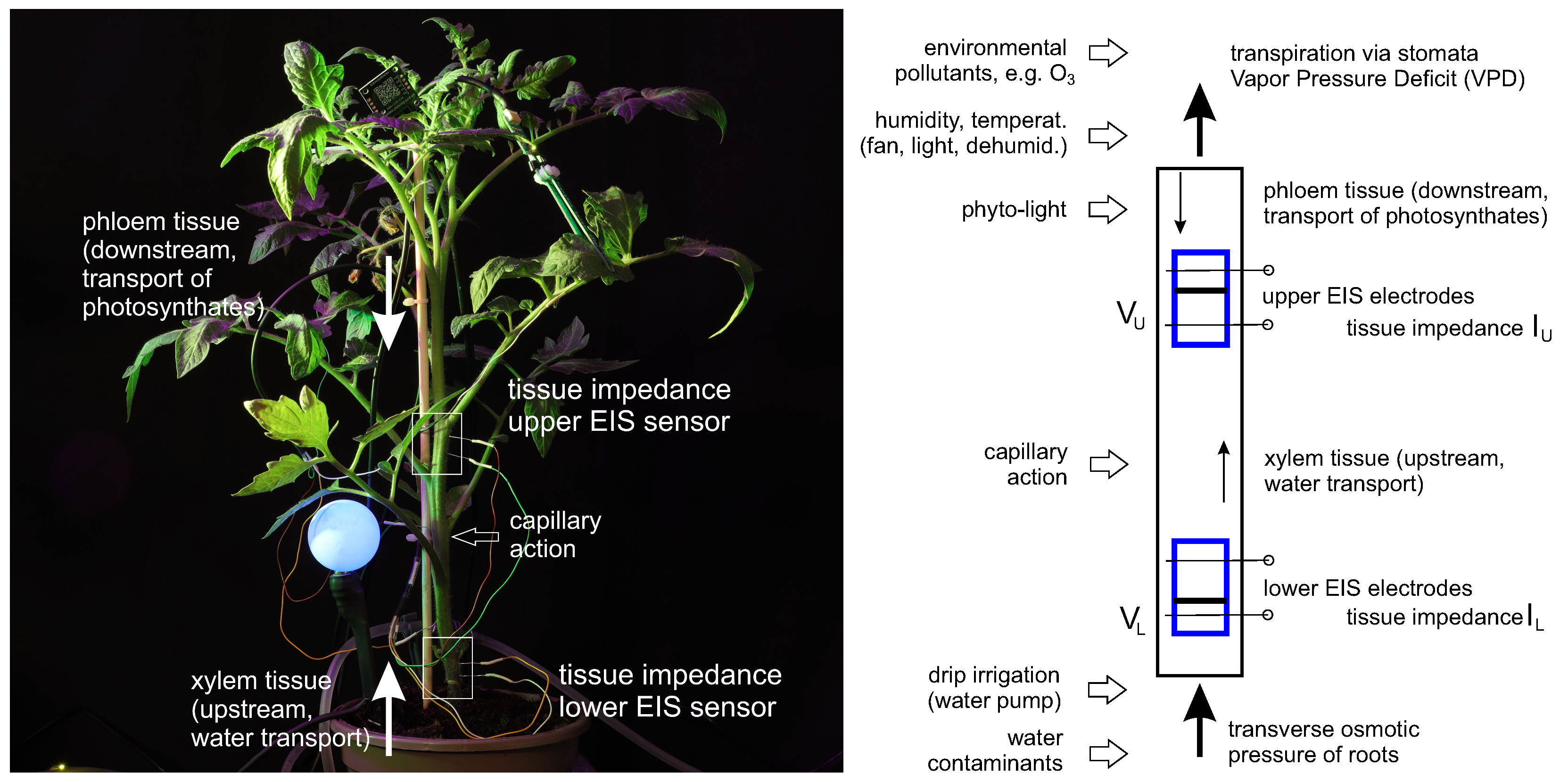
3.2. Fluid Dynamics in the Stem, Measured by the Electrochemical Method
3.3. Electrochemical Analysis of Upflow and Downflow Fluids
3.4. Hydrodynamic Model
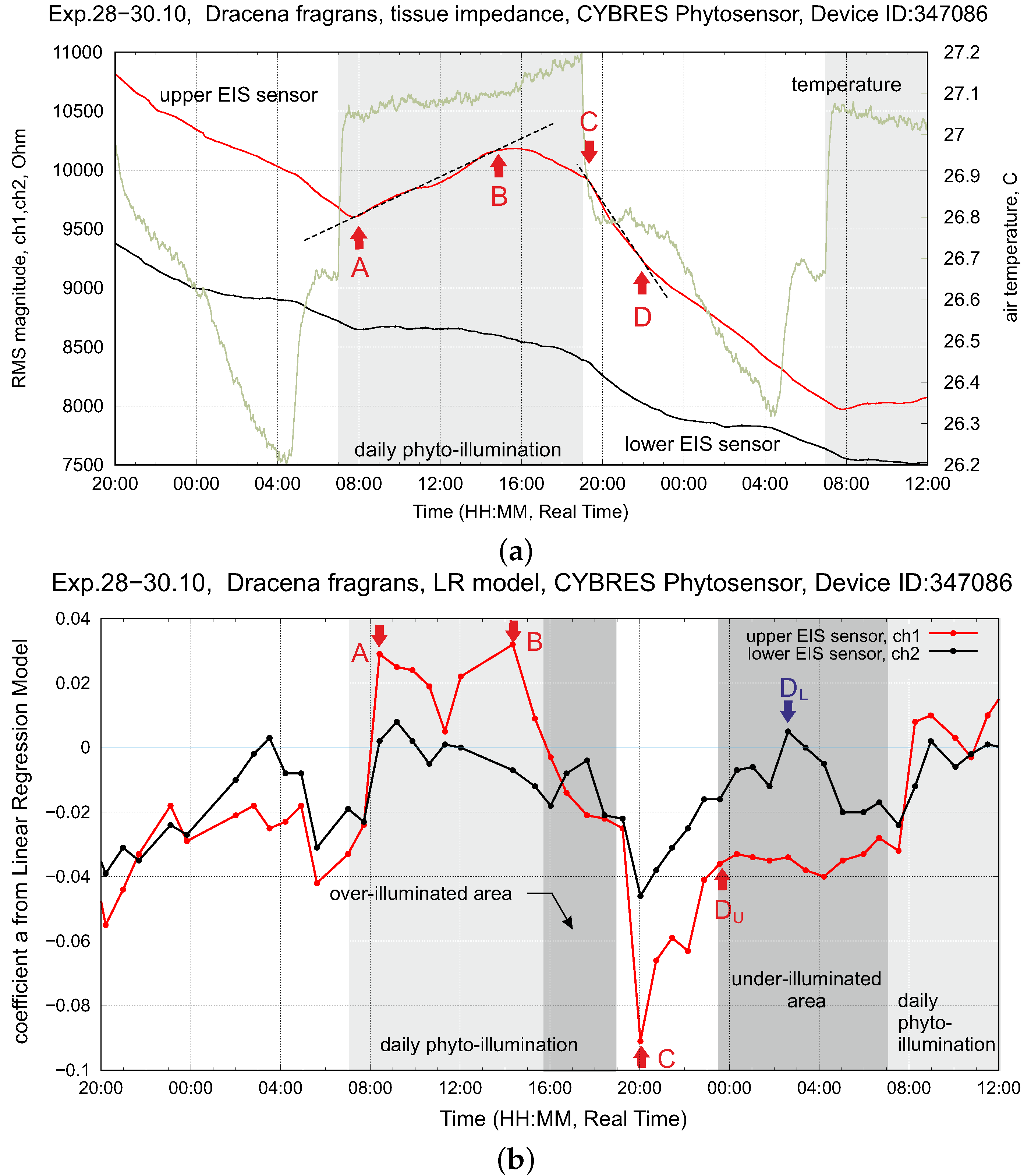
3.5. Characterization of EIS-Based Dynamics in the Hydrodynamic Model
4. Applications of Biofeedback-Based Control
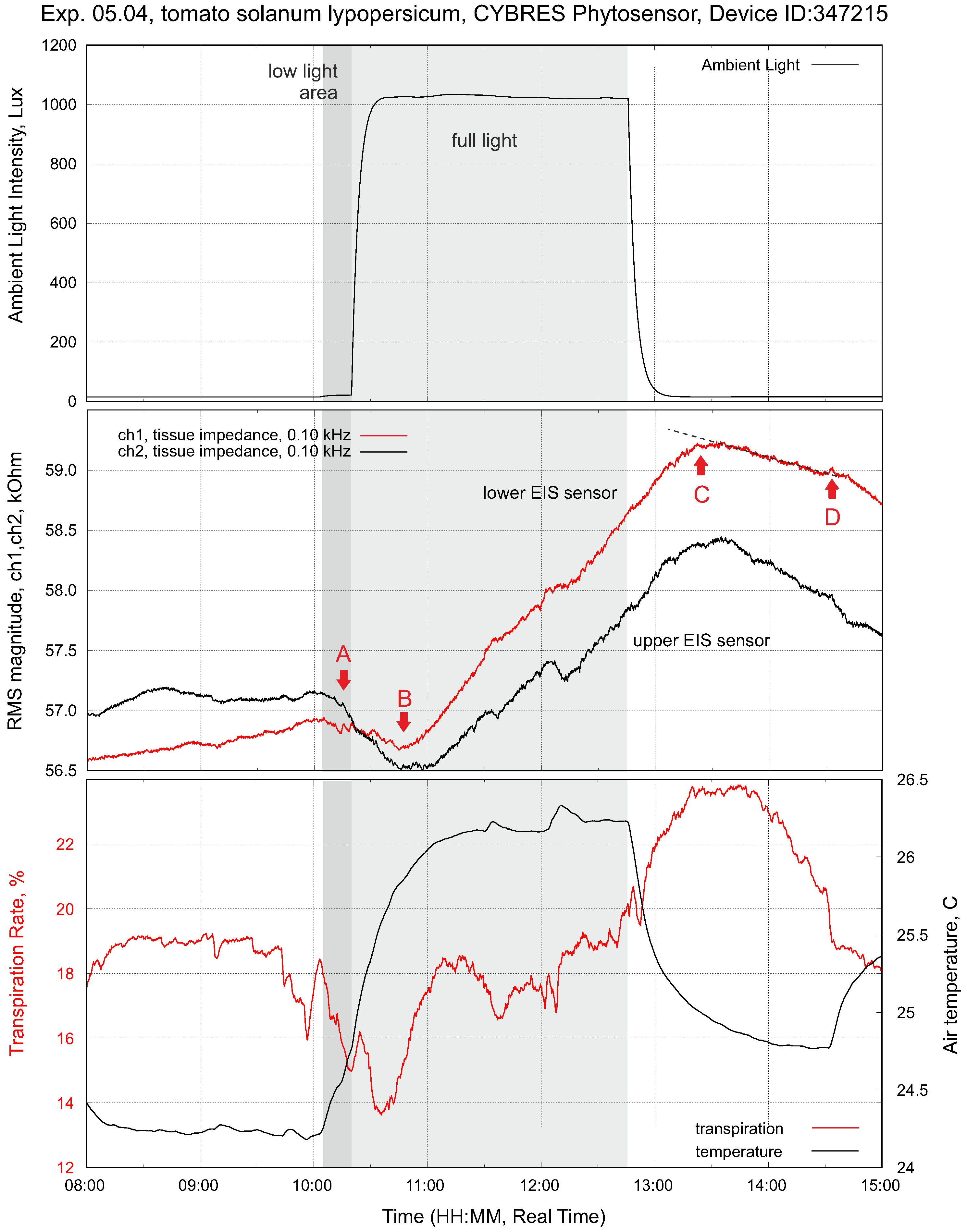
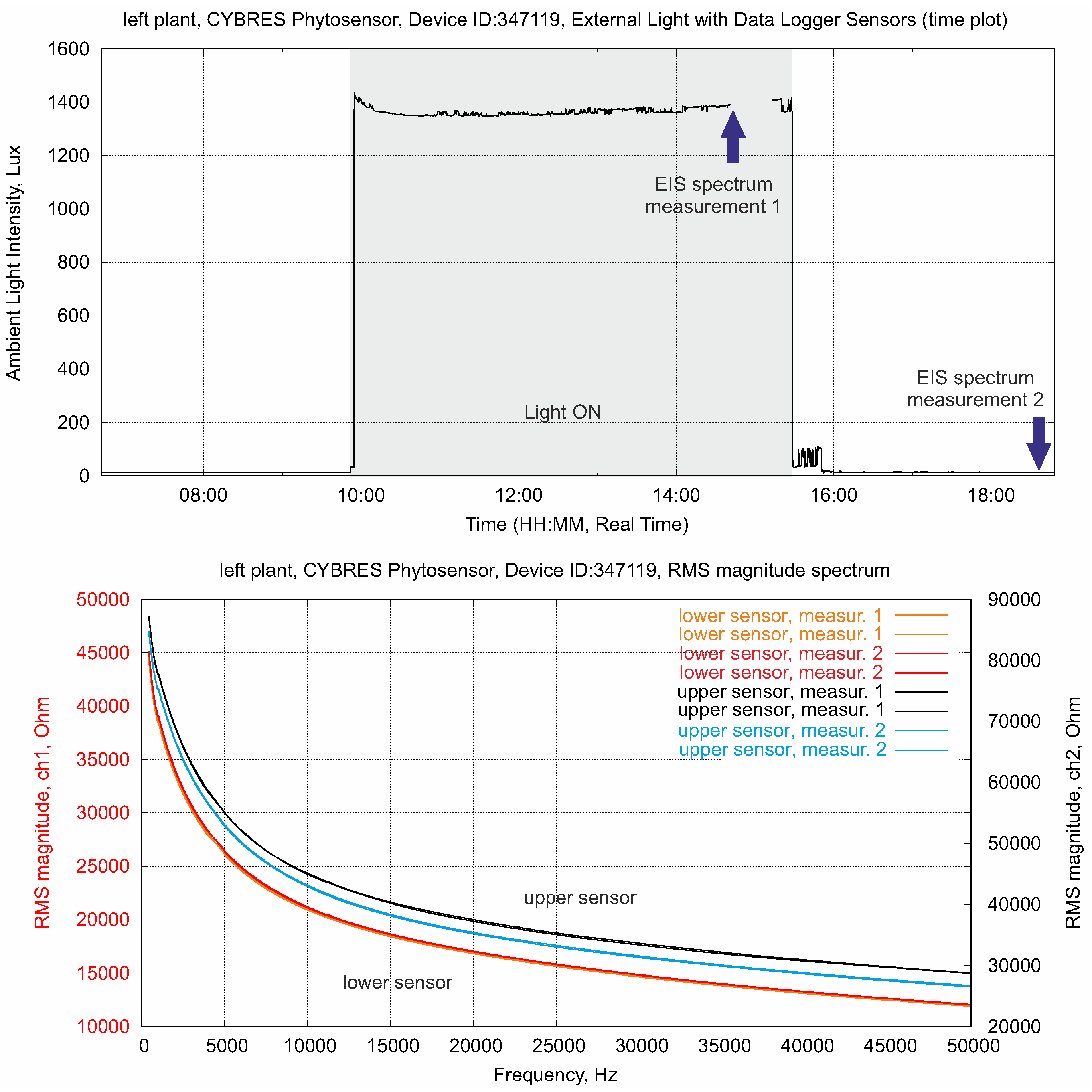
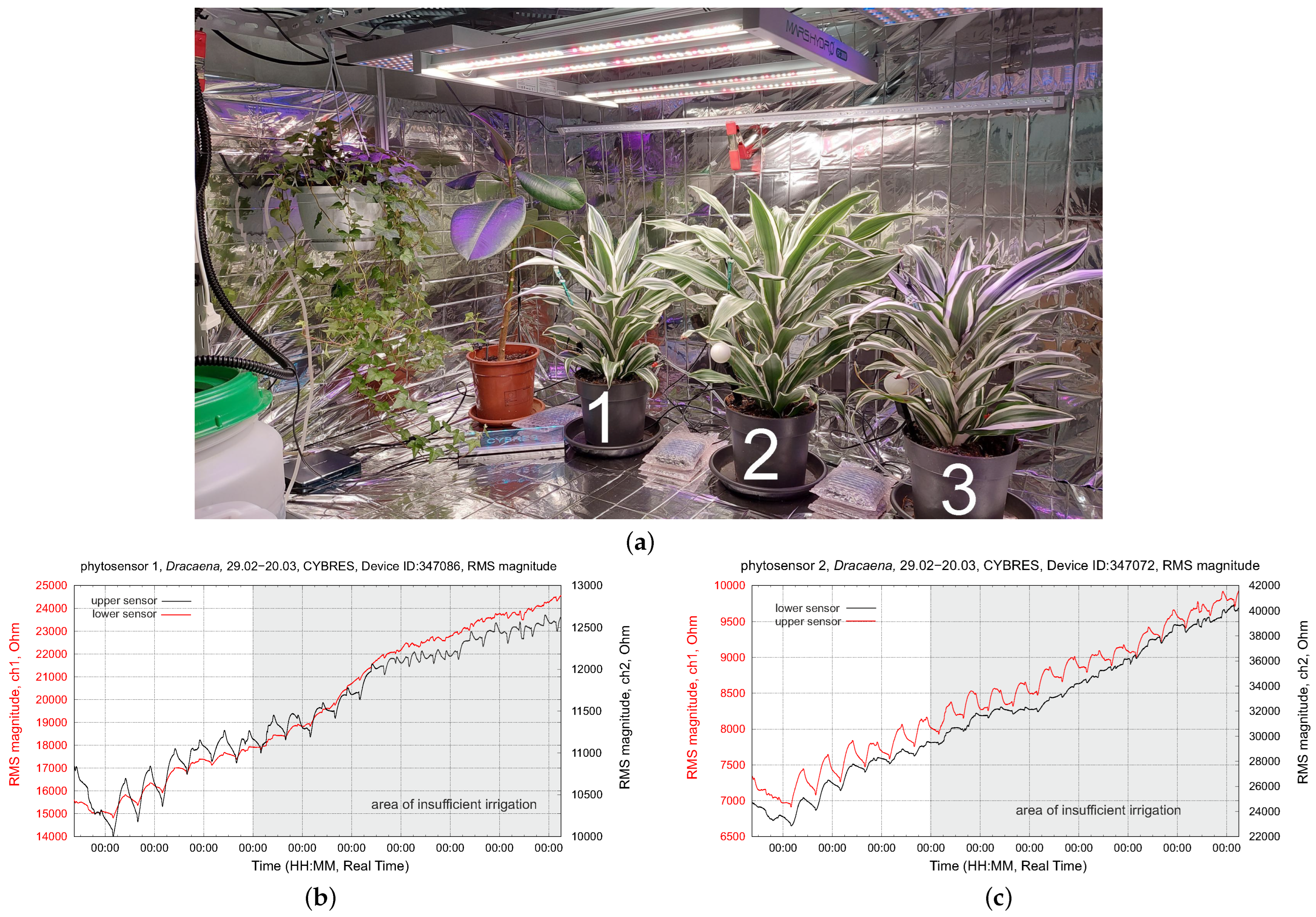
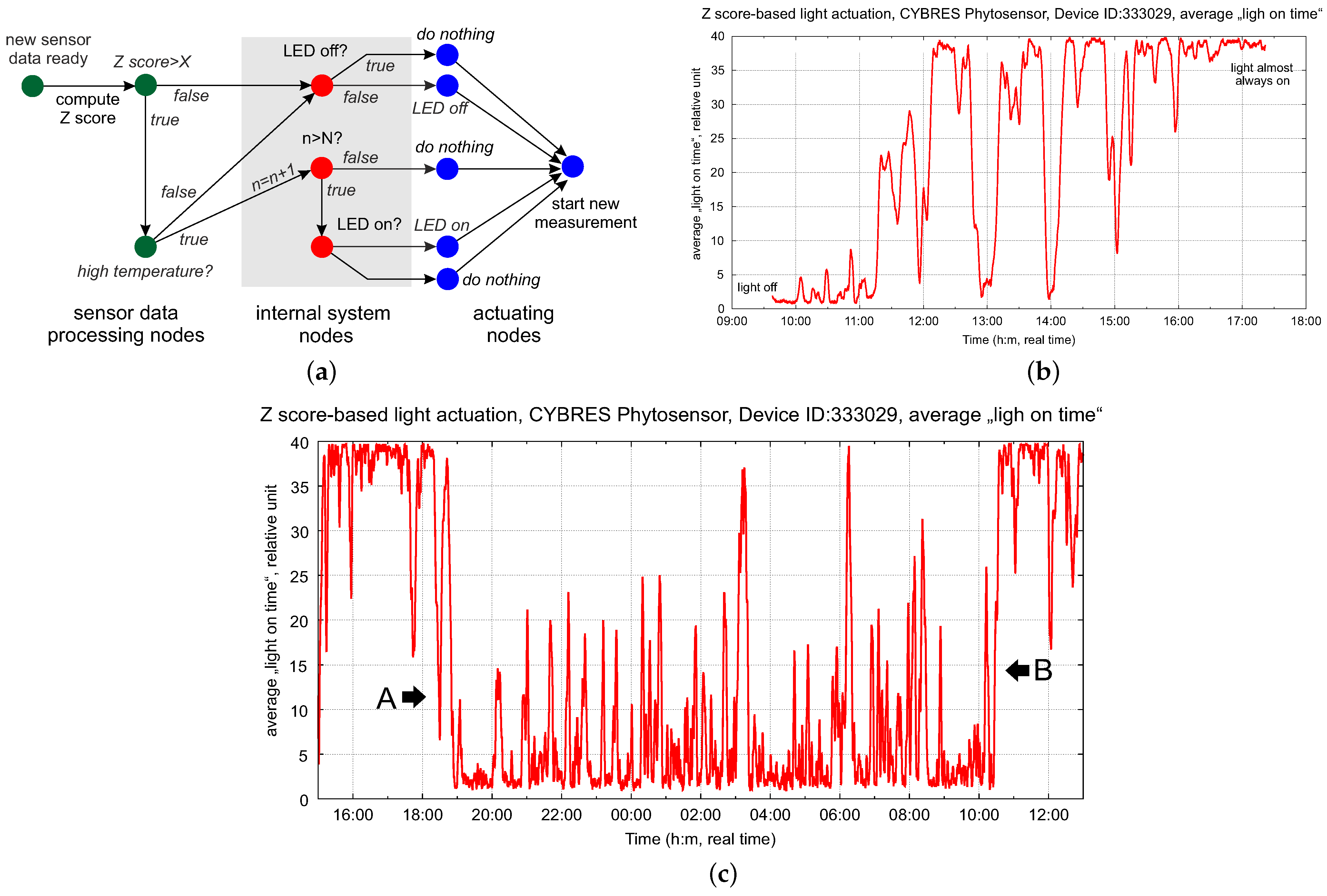
4.1. Biofeedback-Based Control of Phytolight
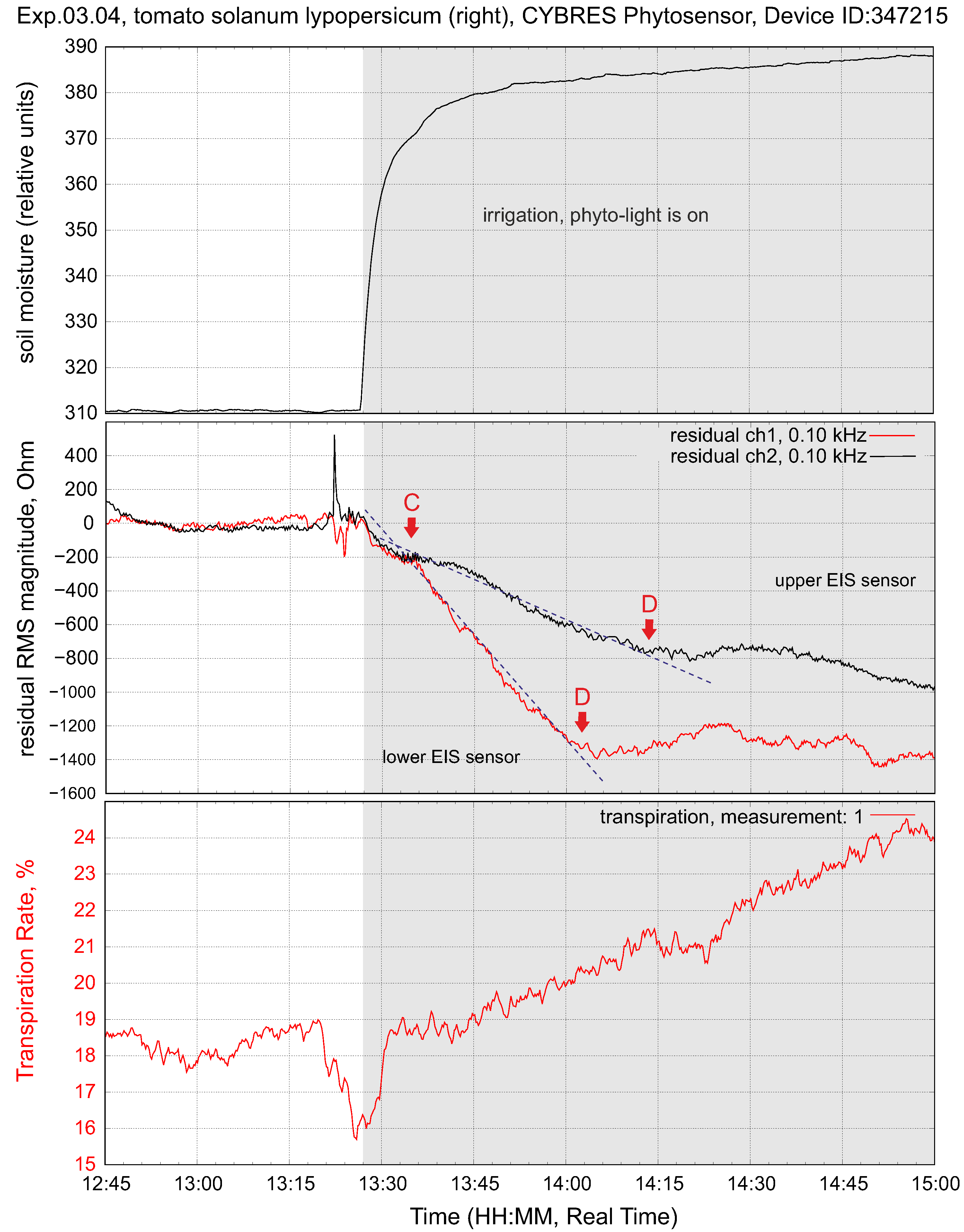
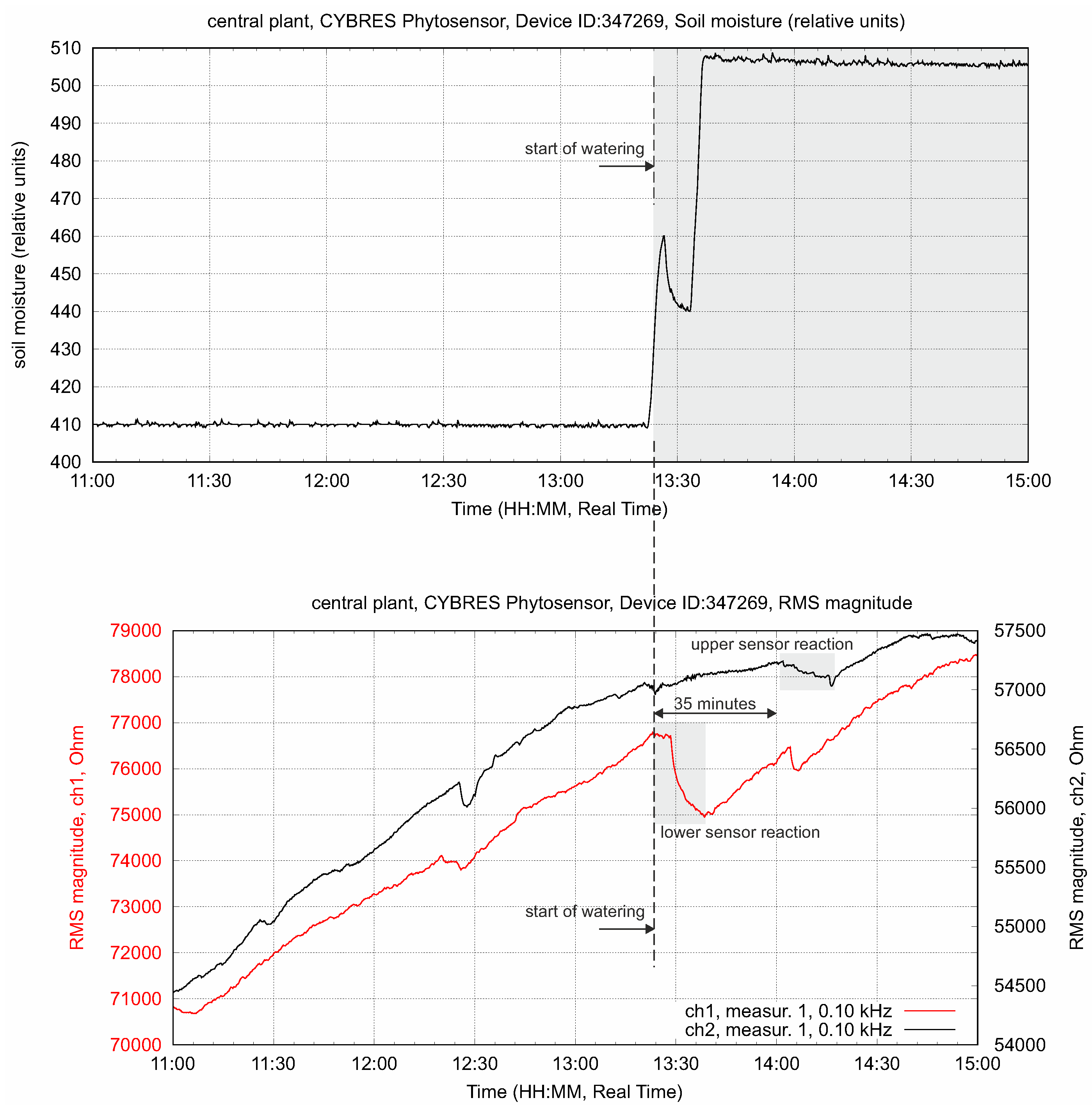
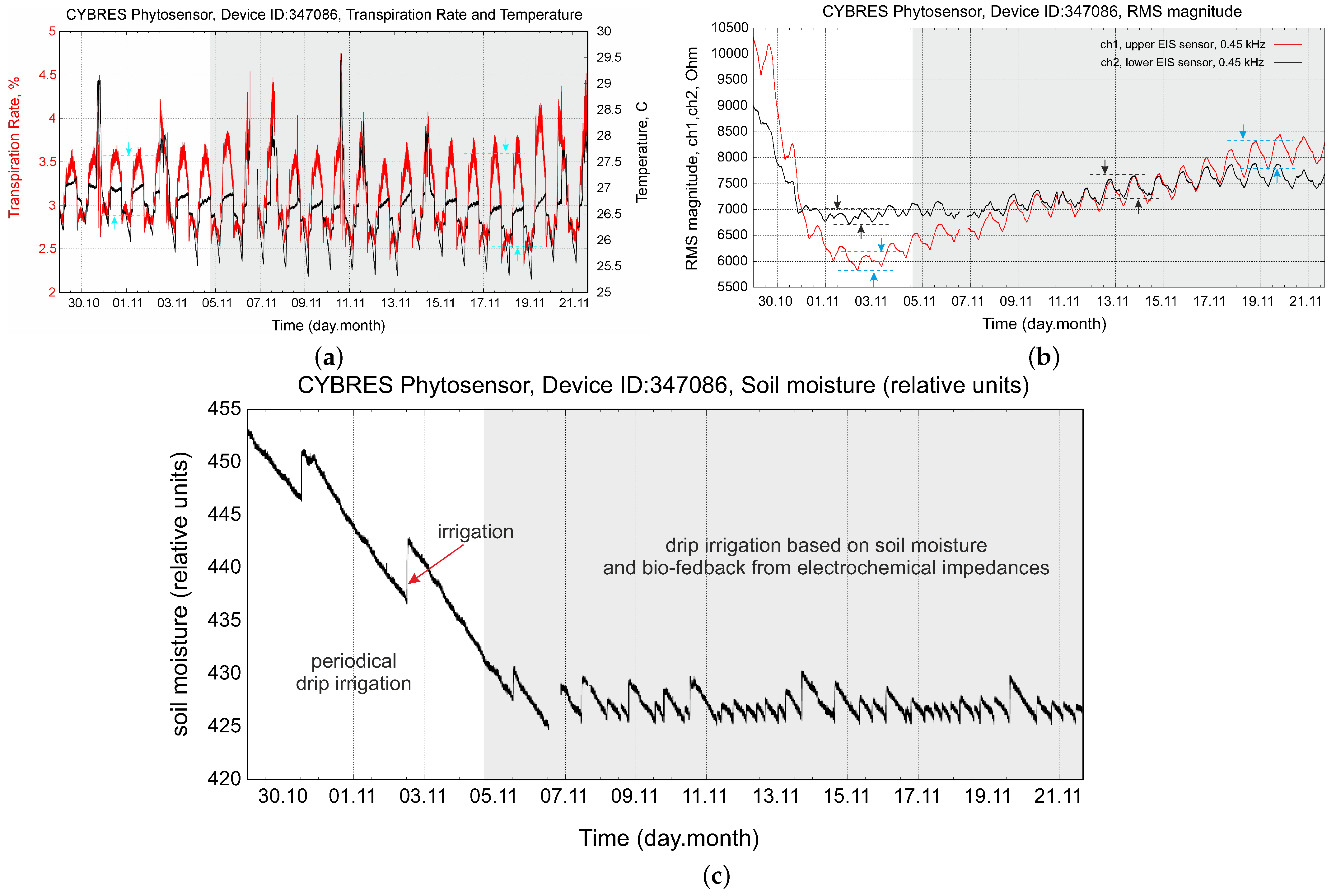
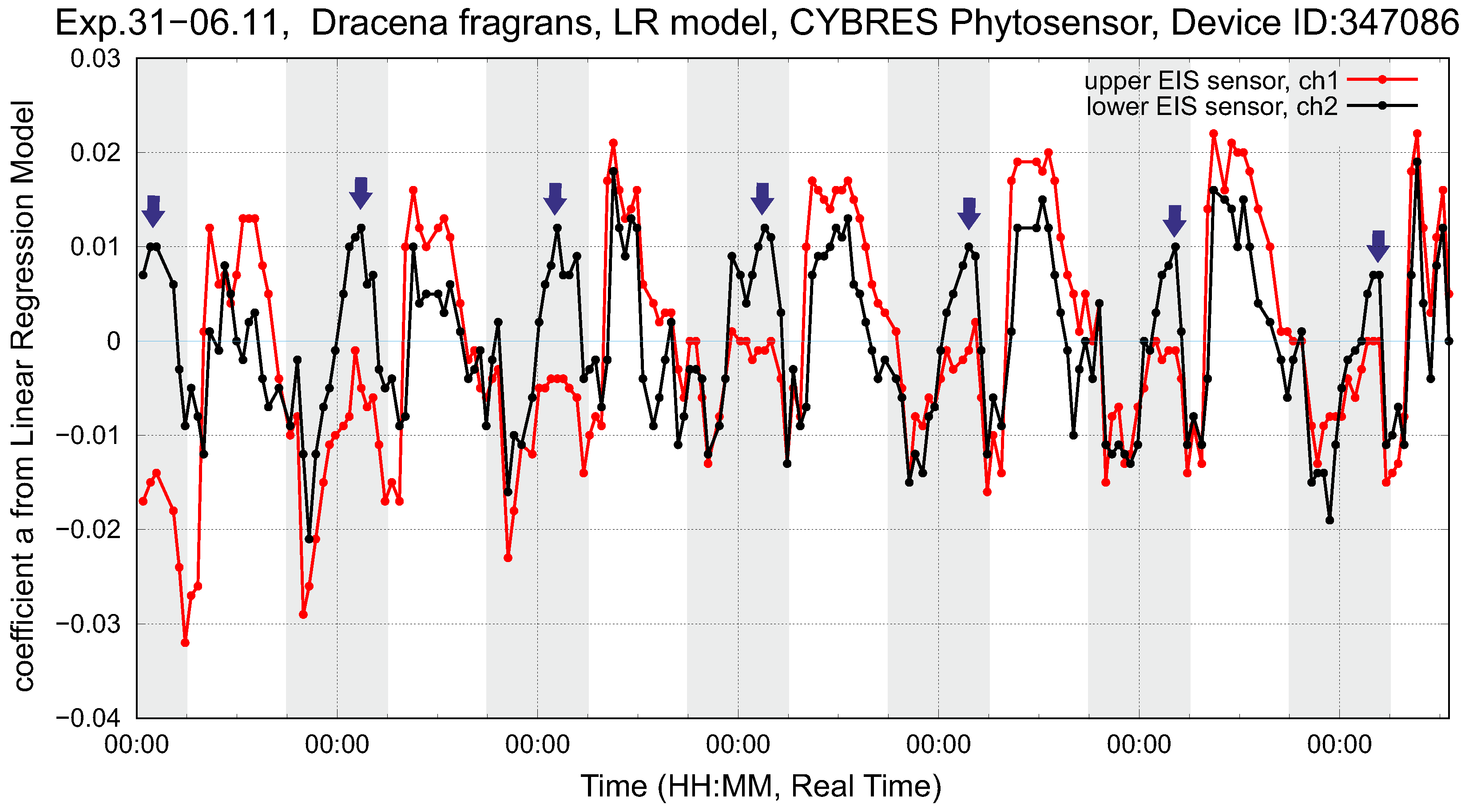
4.2. Biofeedback-Based Irrigation
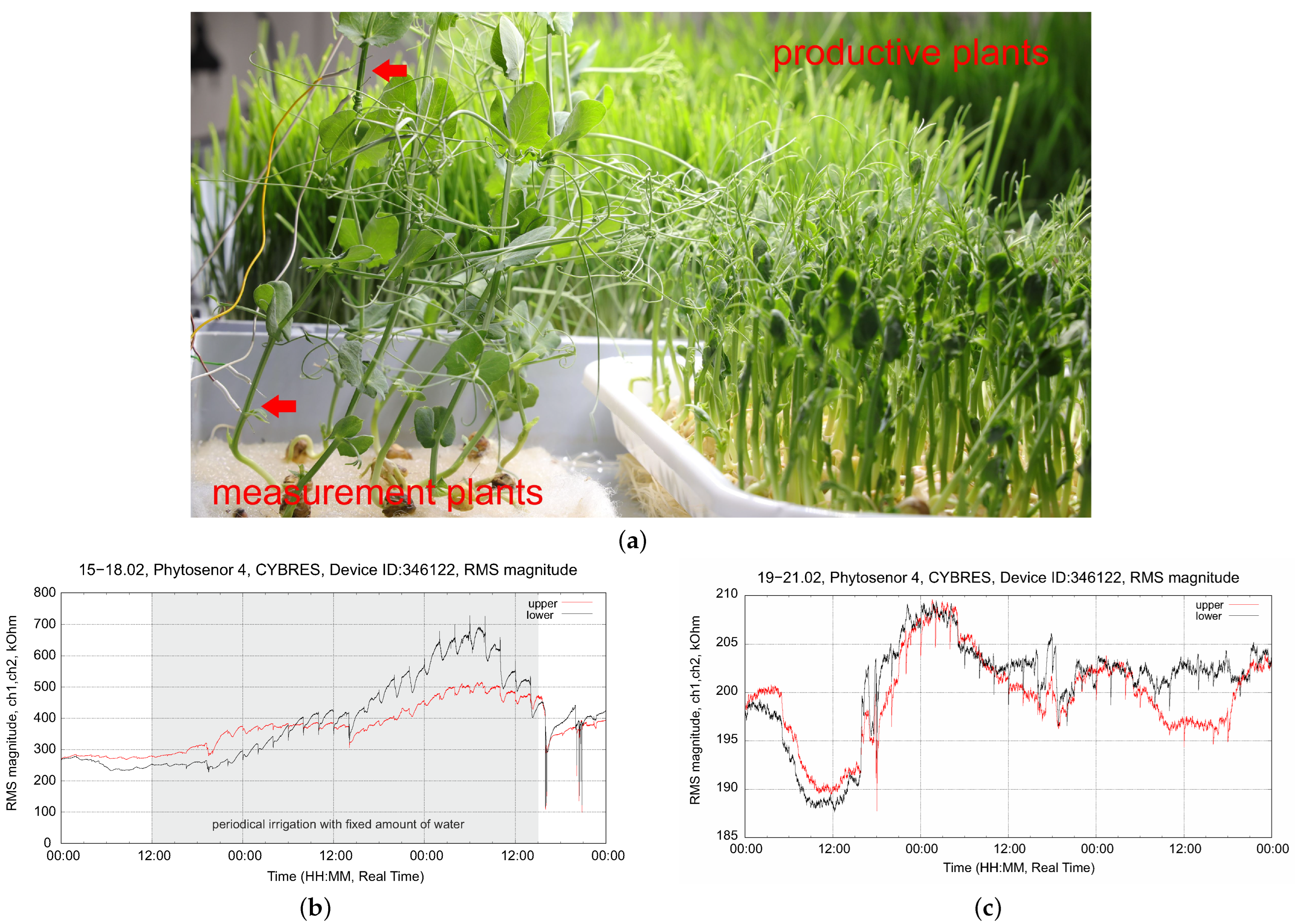
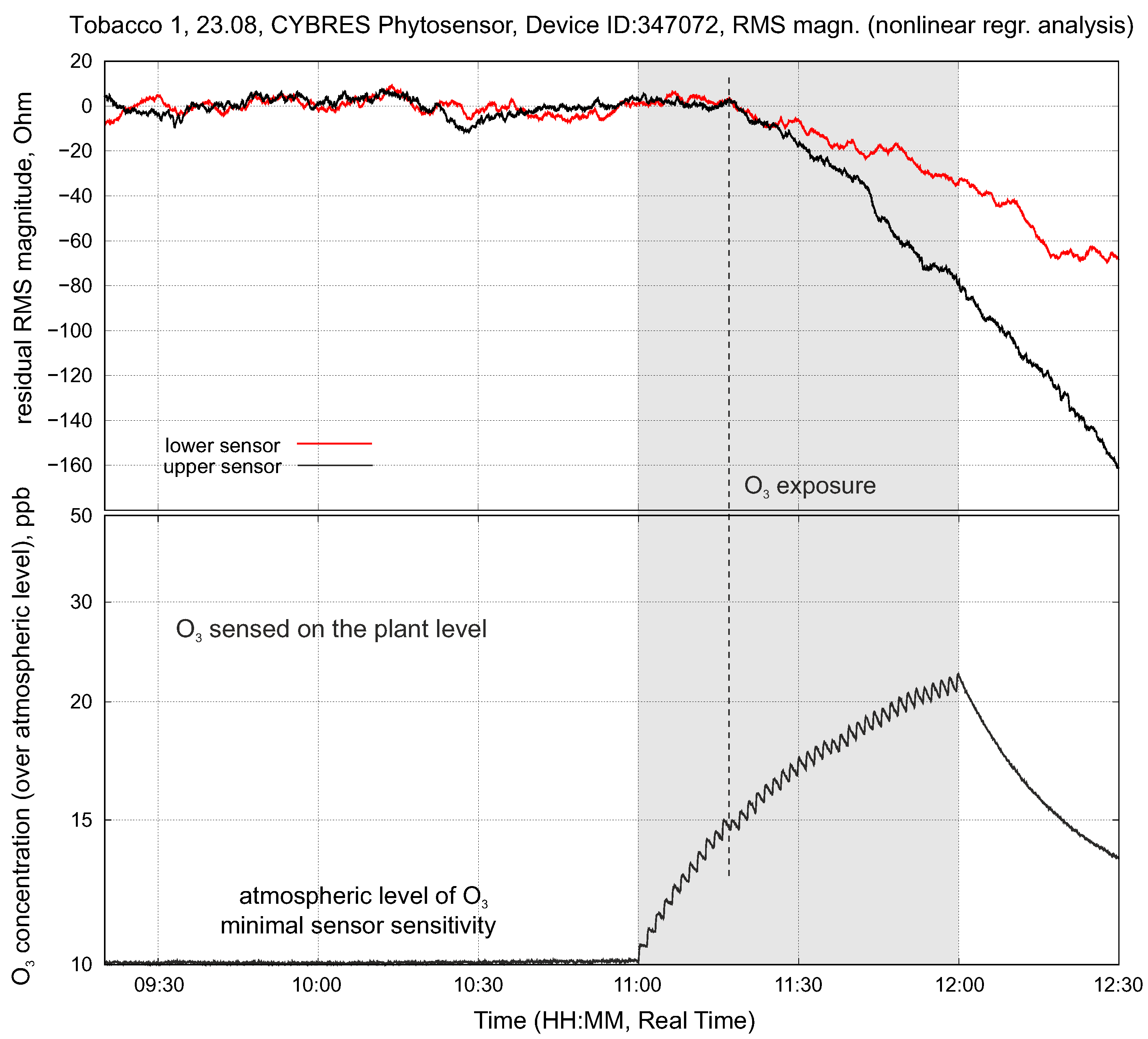
4.3. Stress Detection: Water Deficit and Ozone
5. Discussion
6. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Menciassi, A.; Takeuchi, S.; Kamm, R. Biohybrid systems: Borrowing from nature to make better machines. APL Bioeng. 2020, 4, 020401. [Google Scholar] [CrossRef] [PubMed]
- Webster-Wood, V.A.; Guix, M.; Xu, N.W.; Behkam, B.; Sato, H.; Sarkar, D.; Sanchez, S.; Shimizu, M.; Parker, K.K. Biohybrid robots: Recent progress, challenges, and perspectives. Bioinspir. Biomimetics 2022, 18, 015001. [Google Scholar] [CrossRef] [PubMed]
- Kernbach, S.; Thenius, R.; Kernbach, O.; Schmickl, T. Re-Embodiment of Honeybee Aggregation Behavior in Artificial Micro-Robotic System. Adapt. Behav. 2009, 17, 237–259. [Google Scholar] [CrossRef]
- Kernbach, S.; Haebe, D.; Kernbach, O.; Thenius, R.; Radspieler, G.; Kimura, T.; Schmickl, T. Adaptive Collective Decision Making in Limited Robot Swarms without Communication. Int. J. Robot. Res. 2013, 32, 35–55. [Google Scholar] [CrossRef]
- Schmickl, T.; Bogdan, S.; Correia, L.; Kernbach, S.; Mondada, M.; Bodi, M.; Gribovskiy, A.; Hahshold, S.; Miklic, D.; Szopek, M.; et al. ASSISI: Mixing Animals with Robots in a Hybrid Society. In Proceedings of the Biomimetic and Biohybrid Systems, Second International Conference, Living Machines 2013, London, UK, 29 July–2 August 2013; Springer: Berlin/Heidelberg, Germany, 2013; Volume 9064, pp. 441–443. [Google Scholar] [CrossRef]
- Kernbach, S. Electric-field-coupled oscillators for collective electrochemical perception in biohybrid robotics. Bioinspir. Biomimetics 2022, 17, 065012. [Google Scholar] [CrossRef]
- Hamann, H.; Soorati, M.; Heinrich, M.; Hofstadler, D.; Kuksin, I.; Veenstra, F.; Wahby, M.; Nielsen, S.; Risi, S.; Skrzypczak, T.; et al. Flora robotica—An Architectural System Combining Living Natural Plants and Distributed Robots. In Proceedings of the ECAL 2017: The 14th European Conference on Artificial Life, Lyon, France, 4–8 September 2017. [Google Scholar] [CrossRef]
- Hamann, H.; Wahby, M.; Schmickl, T.; Zahadat, P.; Hofstadler, D.; Stoy, K.; Risi, S.; Faina, A.; Veenstra, F.; Kernbach, S.; et al. Flora Robotica—Mixed Societies of Symbiotic Robot-Plant Bio-Hybrids. In Proceedings of the 2015 IEEE Symposium Series on Computational Intelligence, Cape Town, South Africa, 7–10 December 2015; pp. 1102–1109. [Google Scholar] [CrossRef]
- Buss, E.; Aust, T.; Wahby, M.; Rabbel, T.L.; Kernbach, S.; Hamann, H. Stimuli classification with electrical potential and impedance of living plants: Comparing discriminant analysis and deep learning methods. Bioinspir. Biomimetics 2023, 18, 025003. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Qian, J.; Di, B.; Zhang, G.; Ren, Z. Electrical impedance spectroscopy (EIS) in plant roots research: A review. Plant Methods 2021, 17, 118. [Google Scholar] [CrossRef]
- Jin, K.; Shen, J.; Ashton, R.; White, R.; Dodd, I.; Phillips, A.; Parry, M.; Whalley, R. The effect of impedance to root growth on plant architecture in wheat. Plant Soil 2015, 392, 323–332. [Google Scholar] [CrossRef]
- Windt, C.W.; Nabel, M.; Kochs, J.; Jahnke, S.; Schurr, U. A Mobile NMR Sensor and Relaxometric Method to Non-destructively Monitor Water and Dry Matter Content in Plants. Front. Plant Sci. 2021, 12, 617768. [Google Scholar] [CrossRef]
- Murata, K.; Mitsuoka, K.; Hirai, T.; Walz, T.; Agre, P.; Heymann, J.; Engel, A.; Fujiyoshi, Y. Structural determinants of water permeation through aquaporin-1. Nature 2000, 407, 599–605. [Google Scholar] [CrossRef]
- Pershin, S.; Bunkin, A.; Anisimov, N.; Pirogov, Y.A. Water enrichment by H2O ortho-isomer: Four-photon and NMR spectroscopy. Laser Phys. 2009, 19, 410–413. [Google Scholar] [CrossRef]
- Kernbach, S. Electrochemical characterisation of ionic dynamics resulting from spin conversion of water isomers. J. Electrochem. Soc. 2022, 169, 067504. [Google Scholar] [CrossRef]
- Van Haeverbeke, M.; De Baets, B.; Stock, M. Plant impedance spectroscopy: A review of modeling approaches and applications. Front. Plant Sci. 2023, 14, 1187573. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Poyatos, R.; Aguade, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef]
- Smith, D.; Allen, S. Measurement of sap flow in plant stems. J. Exp. Bot. 1996, 47, 1833–1844. [Google Scholar] [CrossRef]
- Clearwater, M.J.; Luo, Z.; Mazzeo, M.; Dichio, B. An external heat pulse method for measurement of sap flow through fruit pedicels, leaf petioles and other small-diameter stems. Plant Cell Environ. 2009, 32, 1652–1663. [Google Scholar] [CrossRef] [PubMed]
- Kernbach, S.; Kuksin, I.; Kernbach, O. On Accurate Differential Measurements with Electrochemical Impedance Spectroscopy. Water 2017, 8, 136–155. [Google Scholar] [CrossRef]
- Kernbach, S.; Kernbach, O.; Kuksin, I.; Kernbach, A.; Nepomnyashchiy, Y.; Dochow, T.; Bobrov, A. The Biosensor based on electrochemical dynamics of fermentation in yeast Saccharomyces cerevisiae. Environ. Res. 2022, 213, 113535. [Google Scholar] [CrossRef]
- Hamann, H.; Bogdan, S.; Diaz-Espejo, A.; Garcia Carmona, L.; Hernandez-Santana, V.; Kernbach, S.; Kernbach, A.; Quijano-Lopez, A.; Salamat, B.; Wahby, M. WatchPlant: Networked Bio-hybrid Systems for Pollution Monitoring of Urban Areas. In Proceedings of the ALIFE 2021: The 2021 Conference on Artificial Life, Virtual, 19–23 July 2021. [Google Scholar] [CrossRef]
- Laosuntisuk, K.; Elorriaga, E.; Doherty, C.J. The Game of Timing: Circadian Rhythms Intersect with Changing Environments. Annu. Rev. Plant Biol. 2023, 74, 511–538. [Google Scholar] [CrossRef]
- Gagliano, M.; Vyazovskiy, V.V.; Borbely, A.A.; Grimonprez, M.; Depczynski, M. Learning by association in plants. Sci. Rep. 2016, 6, 38427. [Google Scholar] [CrossRef]
- CYBRES. Differential Impedance Spectrometer for Electrochemical and Electrophysiological Analysis of Fluids and Organic Tissues. Handbook and User Manual; CYBRES GmbH: Stuttgart, Germany, 2024. [Google Scholar]
- Fernandez, J.; Alcon, F.; Diaz-Espejo, A.; Hernandez-Santana, V.; Cuevas, M. Water productivity and economic analyses for super high density olive orchards. Acta Hortic. 2022, 395–402. [Google Scholar] [CrossRef]
- Wegner, L.H. Root pressure and beyond: Energetically uphill water transport into xylem vessels? J. Exp. Bot. 2013, 65, 381–393. [Google Scholar] [CrossRef]
- CYBRES. Application Note 24. Analysis of Electrochemical Fluctuations for Fast Impedance Spectroscopy; CYBRES GmbH: Stuttgart, Germany, 2018. [Google Scholar] [CrossRef]
- Boškoski, P.; Debenjak, A.; Boshkoska, B. Fast Electrochemical Impedance Spectroscopy: As a Statistical Condition Monitoring Tool; Springer Briefs in Applied Sciences and Technology; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Pawlowska, B.; Wojtala, D.; Biczak, R. Ionic Liquids as Environmental Pollutants—Analysis of the Effect of Tetrabutylammonium Chloride on the Growth and Development of Wheat and Cucumber. Toxics 2023, 11, 522. [Google Scholar] [CrossRef]
- Tränkner, M.; Tavakol, E.; Jakli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef]
- Ferreira, P.; Ceretta, C.; Tiecher, T.; Basso Facco, D.; Garlet, L.; Soares, C.; Soriani, H.; Nicoloso, F.; Giachini, A.; Brunetto, G.; et al. Rhizophagus Clarus and Phosphorus in Crotalaria juncea: Growth, Glomalin Content and Acid Phosphatase Activity in a Copper-Contaminated Soil. Rev. Bras. Ciênc. Solo 2018, 42, e01702451. [Google Scholar] [CrossRef]
- Hoshika, Y.; Katata, G.; Deushi, M.; Watanabe, M.; Koike, T. Ozone-induced stomatal sluggishness changes carbon and water balance of temperate deciduous forests. Sci. Rep. 2015, 5, 9871. [Google Scholar] [CrossRef]
- Chen, B.; Song, Q.; Pan, Q. Study on Transpiration Water Consumption and Photosynthetic Characteristics of Landscape Tree Species under Ozone Stress. Atmosphere 2022, 13, 1139. [Google Scholar] [CrossRef]
- Kernbach, S.; Pershin, S. Dynamics of capillary effects in spin conversion of water isomers. Pre-Print 2023. [Google Scholar] [CrossRef]
- Chan, A.M.; Bowling, D.R. Assessing the thermal dissipation sap flux density method for monitoring cold season water transport in seasonally snow-covered forests. Tree Physiol. 2017, 37, 984–995. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Vandegehuchte, M.; Steppe, K. Sap flux density measurements based on the heat field deformation method. Trees—Struct. Funct. 2012, 26, 1439–1448. [Google Scholar] [CrossRef]
- Vandegehuchte, M.; Steppe, K. Sap-flux density measurement methods: Working principles and applicability. Funct. Plant Biol. 2013, in press. [Google Scholar] [CrossRef]
- Dennis, J.; Schnabel, R. Numerical Methods for Unconstrained Optimization and Nonlinear Equations; Prentice-Hall Series in Computational Mathematics; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1983. [Google Scholar] [CrossRef]
- Xin, P.; Li, B.; Zhang, H.; Hu, J. Optimization and control of the light environment for greenhouse crop production. Sci. Rep. 2019, 9, 8650. [Google Scholar] [CrossRef]
- Hubbard, R.; Bond, B.; Ryan, M. Evidence that hydraulic conductance limits photosynthesis in old Pinus poderosa trees. Tree Physiol. 1999, 19, 165–172. [Google Scholar] [CrossRef]
- Lee, S.; Rasaiah, J. Proton transfer and the mobilities of the H+ and OH− ions from studies of a dissociating model for water. J. Chem. Phys. 2011, 135, 124505. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Z.; Wang, X.; Sun, S.; Zhang, Y.; Wang, S.; Yang, M.; Ji, F.; Ji, C.; Xiang, D.; et al. Sap velocity, transpiration and water use efficiency of drip-irrigated cotton in response to chemical topping and row spacing. Agric. Water Manag. 2022, 267, 107611. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Z.; Yang, X.; Cai, G.; Shen, Y. Effect of simulated precipitation regimes on sap flow and water use efficiency for xerophytic Caragana korshinskii. Ecol. Indic. 2022, 143, 109309. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, Y.; Yin, L.; Wenninger, J.; Zhang, J.; Hou, G.; Zhang, E.; Uhlenbrook, S. Climatic controls on sap flow dynamics and water use of Salix (psammophila) bush in a semi-arid environment in northwest China. Environ. Earth Sci. 2014, 73, 289–301. [Google Scholar] [CrossRef]
- Vinterhalter, D.; Vinterhalter, B.; Motyka, V. Periodicity and Spectral Composition of Light in the Regulation of Hypocotyl Elongation of Sunflower Seedlings. Plants 2022, 11, 1982. [Google Scholar] [CrossRef] [PubMed]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef]
- Guedes, W.A.; Nobre, R.G.; Soares, L.A.D.A.; de Lima, G.S.; Gheyi, H.R.; Fernandes, P.D.; Ferreira, A.P.N.; da Silva, A.A.R.; de Azevedo, C.A.V.; Silva, D.V.; et al. Irrigation Strategies with Controlled Water Deficit in Two Production Cycles of Cotton. Plants 2023, 12, 2892. [Google Scholar] [CrossRef]
- Geerts, S.; Raes, D. Deficit irrigation as an on-farm strategy to maximize crop water productivity in dry areas. Agric. Water Manag. 2009, 96, 1275–1284. [Google Scholar] [CrossRef]
- Karmakar, S.; Baul Das, A.; Gurung, C.; Ghosh, C. Effects of Ozone on Plant Health and Environment: A Mini Review. Res. J. Agric. Sci. 2022, 13, 612–619. [Google Scholar]
- Hasan, M.M.; Rahman, M.; Skalicky, M.; Alabdallah, N.M.; Waseem, M.; Shah Jahan, M.; Ahammed, G.J.; Fang, X.; Ibrahim, M.; Abou El-Yazied, A. Ozone Induced Stomatal Regulations, MAPK and Phytohormone Signaling in Plants. Int. J. Mol. Sci. 2021, 22, 6304. [Google Scholar] [CrossRef] [PubMed]
- Kernbach, S. In-situ biological ozone detection by measuring electrochemical impedances of plant tissues. 2024; submitted, under review. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Parameter | Description |
|---|---|
| tissue impedance | 4× Ag99 electrodes, 1V excitation, time–frequency EIS |
| electrochemical spectroscopy | time–frequency EIS, fast EIS for in situ sap analysis |
| biopotentials | 4× Ag99 electrodes, input impedance Ohm, input bias current ±70 pA |
| leaf transpiration | differential air–humidity-based method, CYBRES |
| leaf temperature | precision LM35 sensor |
| thermal sap flow | heat balance and heat pulse methods, 3× t-sensing, PID stabilized, CYBRES |
| fluid content of tissue | (electrochemical) 4× electrode method, CYBRES |
| Sensed Environm. Param. | Description |
| light, humidity, temperature | APDS-9008-020, HIH-5031-001, LM35CA |
| EM emission, magnetometer | 450 Mhz–2.5 Ghz RF power meter, MAX2204 chip; 3-axis, LIS3MDL |
| soil humidity, temperature | capacitive-based sensor, CYBRES |
| water parameters | conductivity, pH, temperature, etc. |
| CO2, PM1-2.5-10, O3 | SCD4x, accuracy ±(40 ppm + 5%); SPS30, accuracy 10%, CENSIRION |
| Environmental Stimuli | Description |
| light, irrigation, temperature | full-spectrum light, IR/UV supplementary light, automatic irrigation system, heater |
| EM emission, O3, PMx | mobile phones (GSM 890-1.805 MHz) WIFI routers (2.4 GHz), weak magnetic fields and O3 generators |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kernbach, S. Biofeedback-Based Closed-Loop Phytoactuation in Vertical Farming and Controlled-Environment Agriculture. Biomimetics 2024, 9, 640. https://doi.org/10.3390/biomimetics9100640
Kernbach S. Biofeedback-Based Closed-Loop Phytoactuation in Vertical Farming and Controlled-Environment Agriculture. Biomimetics. 2024; 9(10):640. https://doi.org/10.3390/biomimetics9100640
Chicago/Turabian StyleKernbach, Serge. 2024. "Biofeedback-Based Closed-Loop Phytoactuation in Vertical Farming and Controlled-Environment Agriculture" Biomimetics 9, no. 10: 640. https://doi.org/10.3390/biomimetics9100640
APA StyleKernbach, S. (2024). Biofeedback-Based Closed-Loop Phytoactuation in Vertical Farming and Controlled-Environment Agriculture. Biomimetics, 9(10), 640. https://doi.org/10.3390/biomimetics9100640