Biological Practices and Fields, Missing Pieces of the Biomimetics’ Methodological Puzzle




Abstract
1. Introduction
1.1. Positioning of the Article
1.2. State of the Art
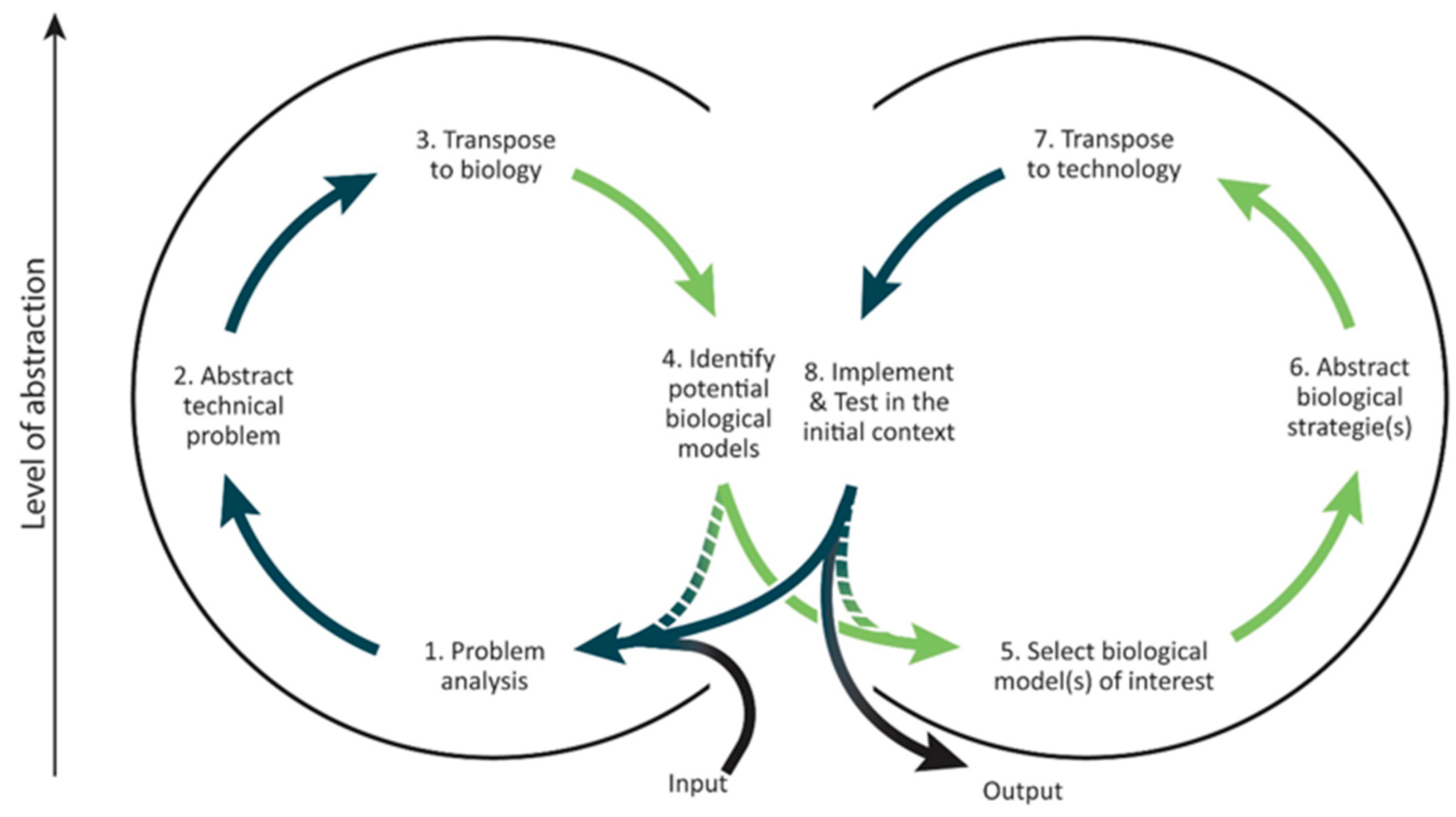
1.2.1. Biomimetics Methodological Framework
1.2.2. Practical Impediments and Current Resolution Strategies
- What to search for? Overall, teams are asked to search for biological models. But again, what is a biological model in biomimetics? Is it a biological process? An organism? A biological system? This ill-define “biological model” represents a first difficulty.
- Where to search for? Most of the scientific findings are stored online on various types of platforms (scientific databases, popular science website, etc.). Facing these various possibilities, the question of where to search, depending on the teams’ expertise and objectives, appears crucial.
- Finally comes the “how”, namely how to identify the proper keywords leading to the searched information?
- How to identify selection criteria and perform evaluation on those criteria to sort the various models that are considered relevant?
- How to assess the quality of the biological data, whether it is enough to formulate a biological model, and its adequacy?
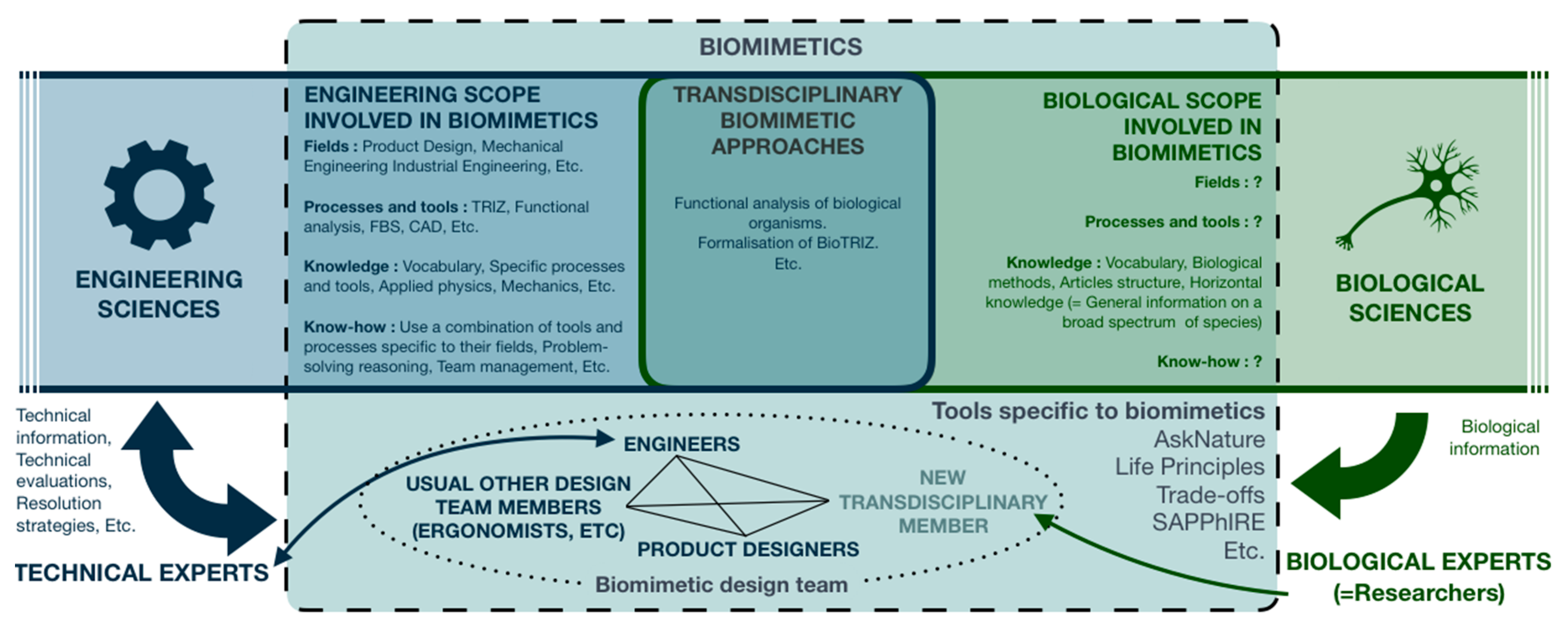
1.2.3. A Biological Approach, another Alternative
1.2.4. Integration of Stakeholders with a Background in Biology
1.3. Synthesis and Research Question
2. Material and Methods
- Research clarification: This phase corresponds to the Section 1.
- Descriptive phase: Based on the literature, we pointed out specific impediments on the targeted steps. This phase is divided between Section 3.1.1 (for the 3rd step) and Section 3.2.1 (for the 4th step).
- Prescriptive phase: We exposed conceptual bridges linking those pitfalls with biological fields and prescribed practical contributions. We also underlined the potential impact on the process to anticipate and better integrate these new practices. This phase is divided between Section 3.1.2 (for the 3rd step) and Section 3.2.2 (for the 4th step).
3. Results
3.1. Findability and the Ill-Defined “Transposition to Biology”
3.1.1. First Descriptive Step: the 3rd Step, the Initial Grain of Sand
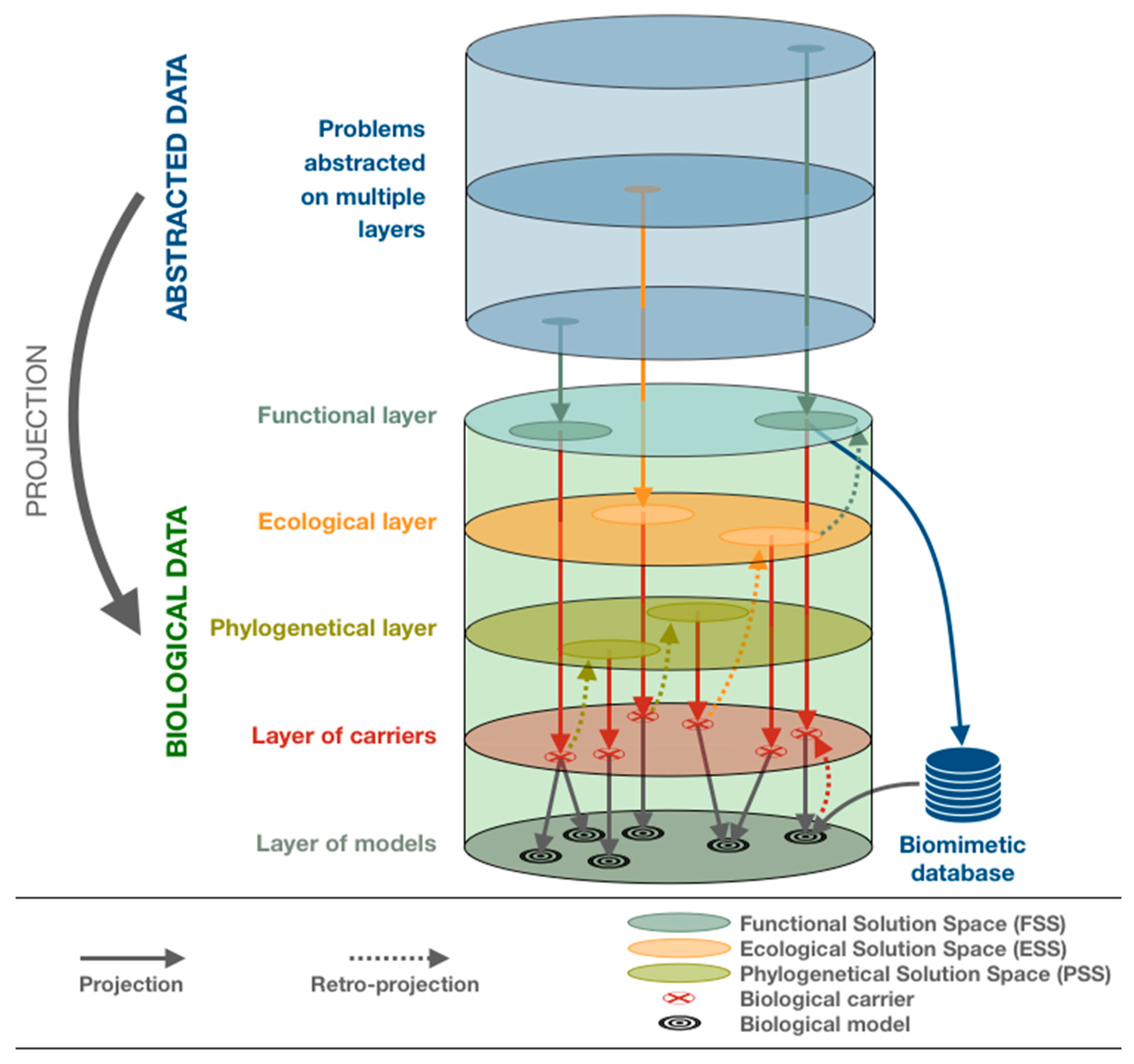
- The required inputs from the second step: generic models and associated functions of interest.
- The goal of the 3rd step: to transpose technical problems and their environments to biology.
- The form of the 3rd step’s outputs: requests to be used as bridges between the abstracted model and the biological solutions.
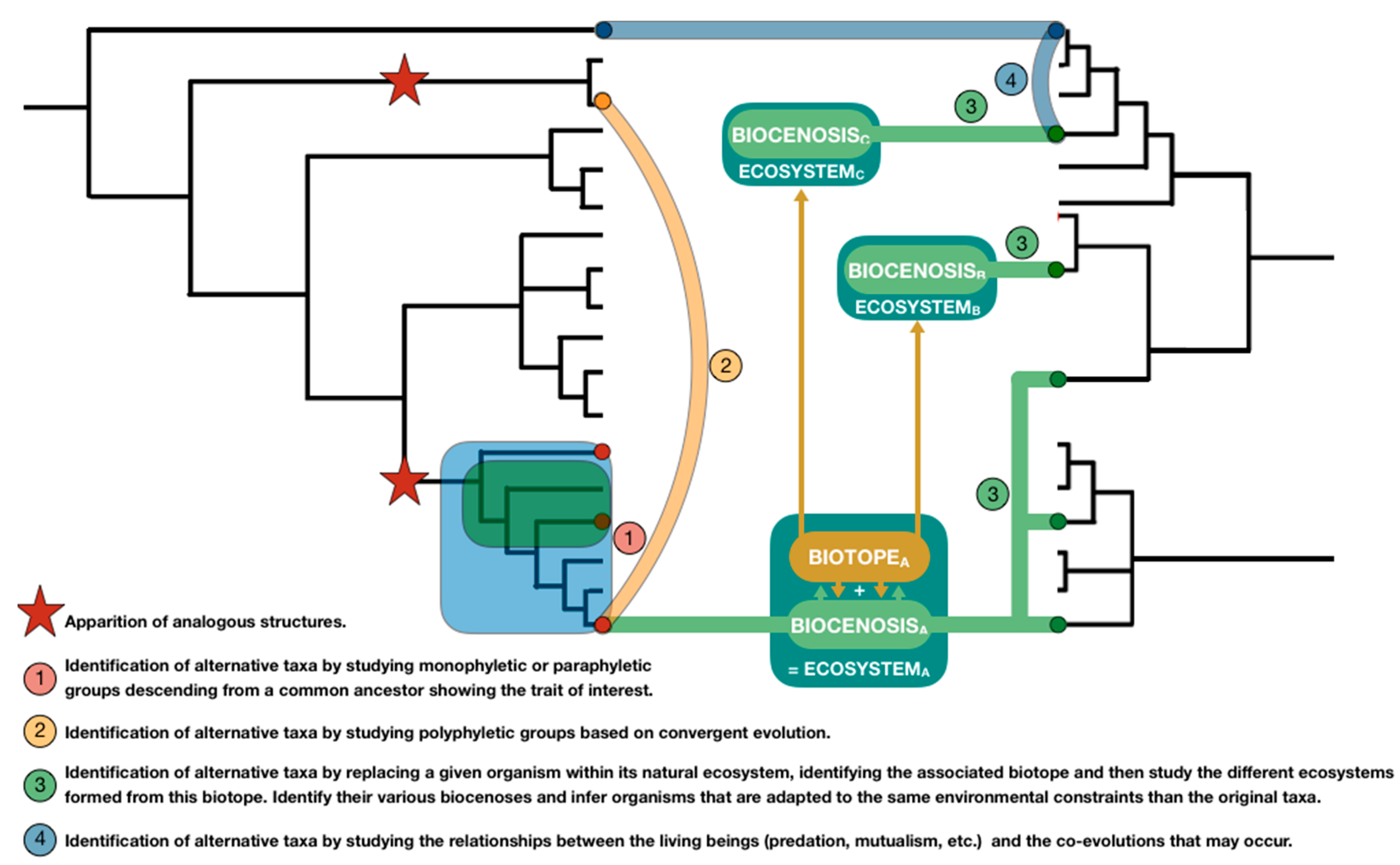
3.1.2. Prescriptive Step: Ecology and Solution Spaces
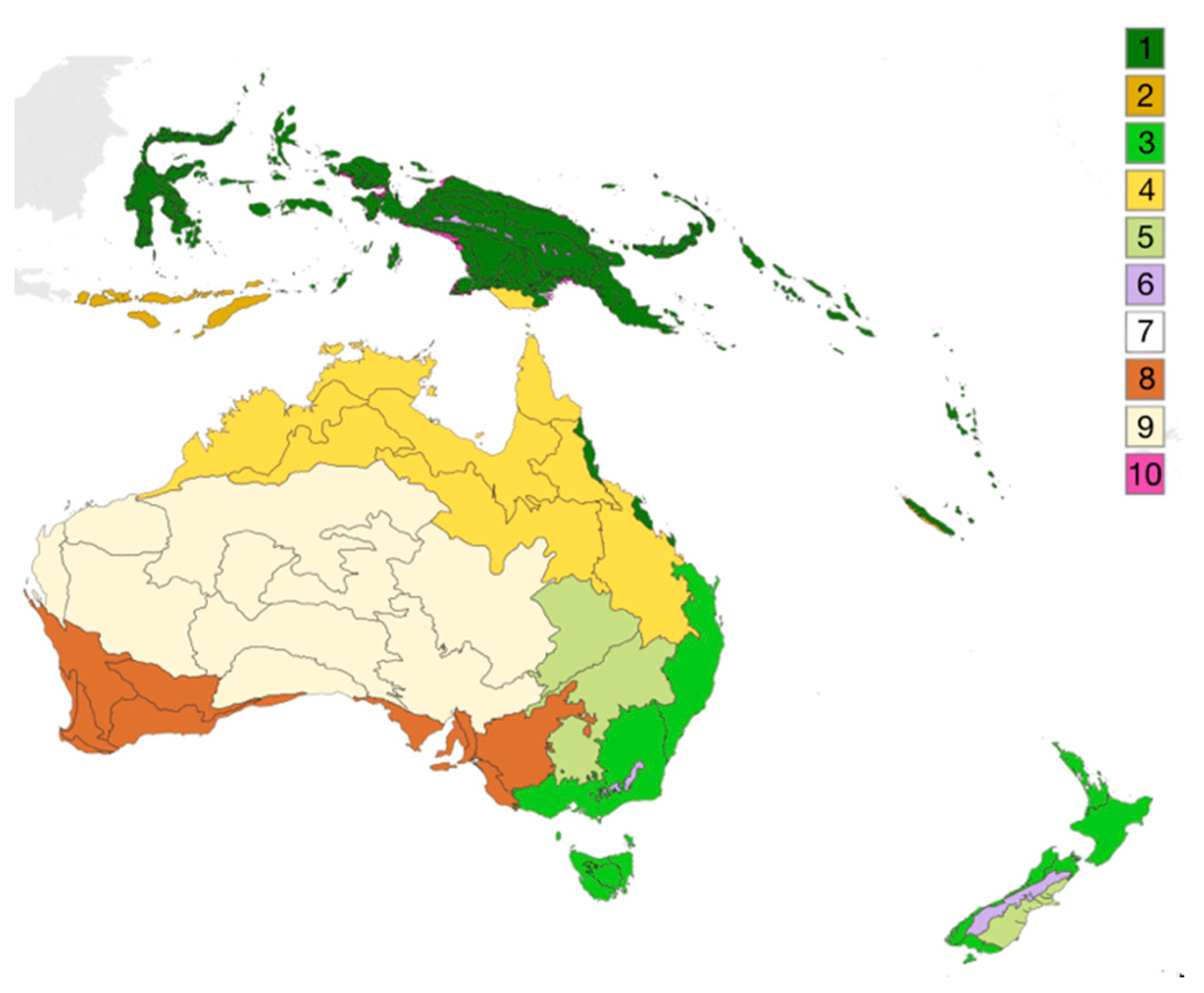
- Ecozones, “represent unique faunas and floras of different continents or ocean basins” [82].
- Biomes are described as “different areas of the world that share similar environmental conditions, habitat structure and patterns of biological complexity (e.g., beta diversity) and that contain communities with similar guild structures and species adaptation” [80].
- A terrestrial ecoregion is defined as “relatively large units of land containing a distinct assemblage of natural communities and species, with boundaries that approximate the original extent of natural communities prior to major land-use change.” [80].
3.2. Findability and the Identification of Biological Models of Interest
3.2.1. First Descriptive Step: Concepts Associated with the 4th Step
3.2.2. Prescriptive Step: From Solution Spaces to Biological Models
- When focusing on Myrmicinae, the search leads to studies on the genus Atta, and brings further information on these species’ ability to ventilate their nest based on the latest research findings [91,92] and associated studies of reference [93,94], as along as alternative solution by other species of the same genus, such as nest relocation under canopee coverage by Atta Sexdens [95]. These articles may also lead biomimetic team to identify other taxonomic groups sharing the same FSS, like the genus Acromyrmex [92] which is also part of the Myrmicinae sub-family or Macrotermes, in a genus of termites, part of a much higher taxonomic group, the Neoptera infra-class. As a result of their greater taxonomic distance, species of these groups, such as Acromyrex heyeri or Macrotermes belicosus, have developed different solution to the problem of thermal regulation: the adaptation of the nest’s thatch thickness and porosity [96].
- When focusing on Vespidae, the search led to no direct solutions.
- When focusing on Hymenoptera, the search leads to the identification of an additional article of interest [97]. Interestingly, it appears after a deeper analysis of the article that the keyword “ventilation” is link to the respiratory system and not directly with thermal regulation. However, along the article several ESS are detected (like the Mojave Desert or the Saharan desert) as part of the biome “Deserts and xeric shrublands” and associated with various species such as Messor pergandei or Cataglyphis bicolor. Article specifically studying these species are also identified, leading to the identification of a new model such as discontinuous ventilation cycles (DVC) to reduce water loss in Cataglyphis bicolor [98].
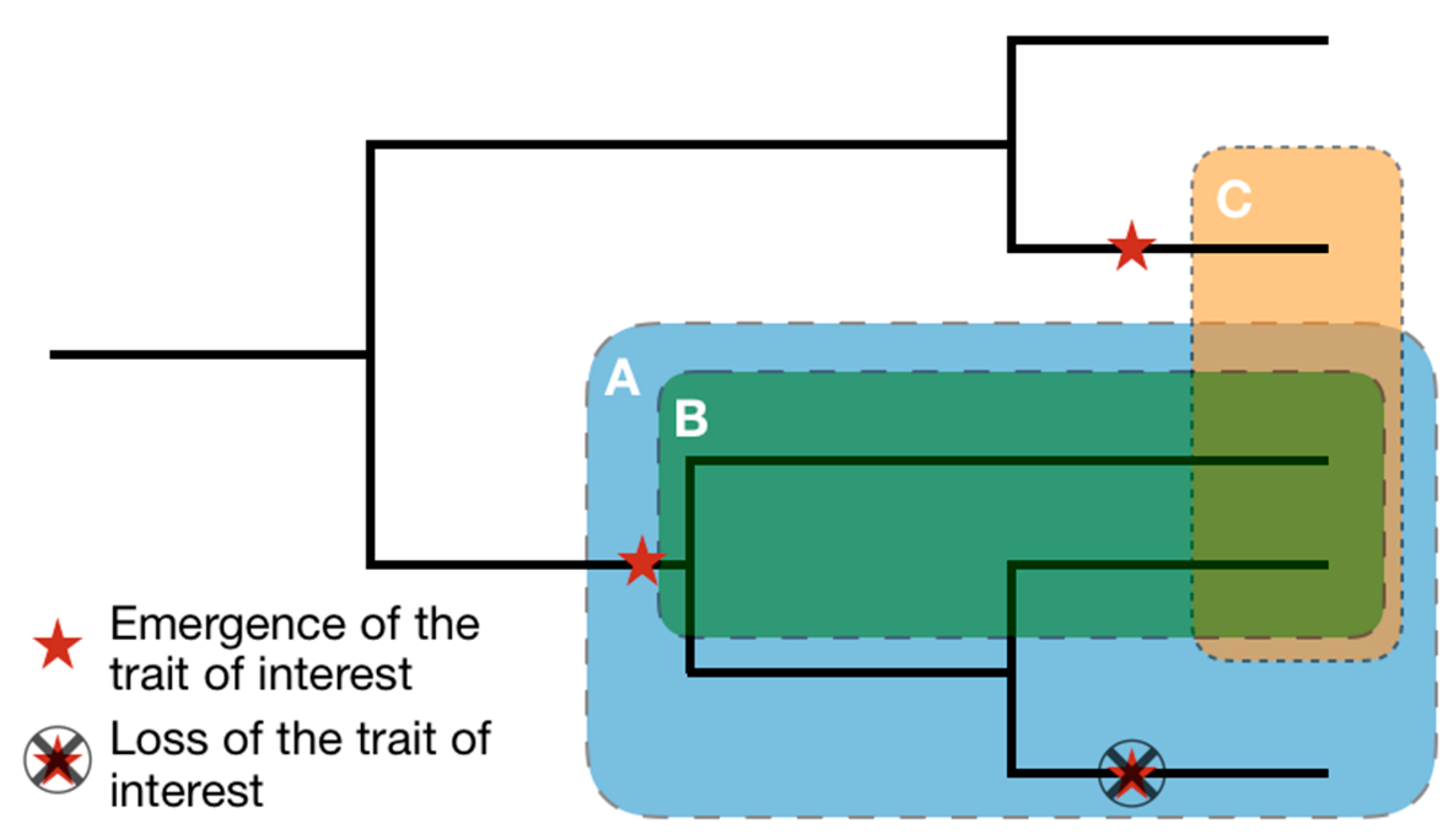
- Biological carriers of reference are organisms which, by themselves, represent biological models directly usable by the team. We suggest four cases that can lead to a biological carrier of reference. Organisms may (1) have already been used in previous biomimetic projects or described in biomimetic articles, (2) have been studied from an interdisciplinary standpoint, more specifically, when chemical, physical or mathematical studies have been performed on biological models, (3) have been extensively studied from a biological standpoint, as model organisms (Mus musculus, Drosophila melanogaster, Escherichia coli, etc.), as key organisms for a given field of application (Homo sapiens for medicine, Bos taurus for agriculture, etc.), as subjects of biological studies specifically focusing on the model of interest, or (4) be particularly well known by a teammate who then ensures the explanation of the associated biological model. Carriers of reference correspond to the final step toward the identification of biological models. Existing databases mostly gather biological carriers of reference, leading them to be the best known by the overall community. Based on the example previously presented, Atta vollenweideri and Macrotermes belicosus can be considered as carriers of reference because of (1) and (3) [99,100].
- Intermediary carriers are often firstly considered as potential carriers of reference before a lack of studies, a lack of resources or their inadequacy with the project requirement, make their associated models too hard to extract or irrelevant. Nevertheless, through literature search and functional, ecological, or phylogenetical tools, these carriers may be used as keywords to identify and better characterize solution spaces. Where biological carriers of reference can be used as intermediary carriers, the reverse is not true. Based on the example previously presented, Cataglyphis bicolor can be considered as an intermediary carrier as it can lead to the identification of other, better-studied, carriers, such as Cataglyphis bombycina [101].
- Carriers of diversification carry alternative versions of the biological models described in the carriers of reference. They might be less studied, farther from the technical context and so potentially harder to transfer, but they represent the biological diversity surrounding the model. Contrarily to intermediary carriers, their models are at least partly extracted, increasing the variability of a given biological model of interest. Based on the example previously presented, some representants of the Atta genus, such as Atta sexdens, can be considered as carriers of diversification with respect to Atta vollenweideri.
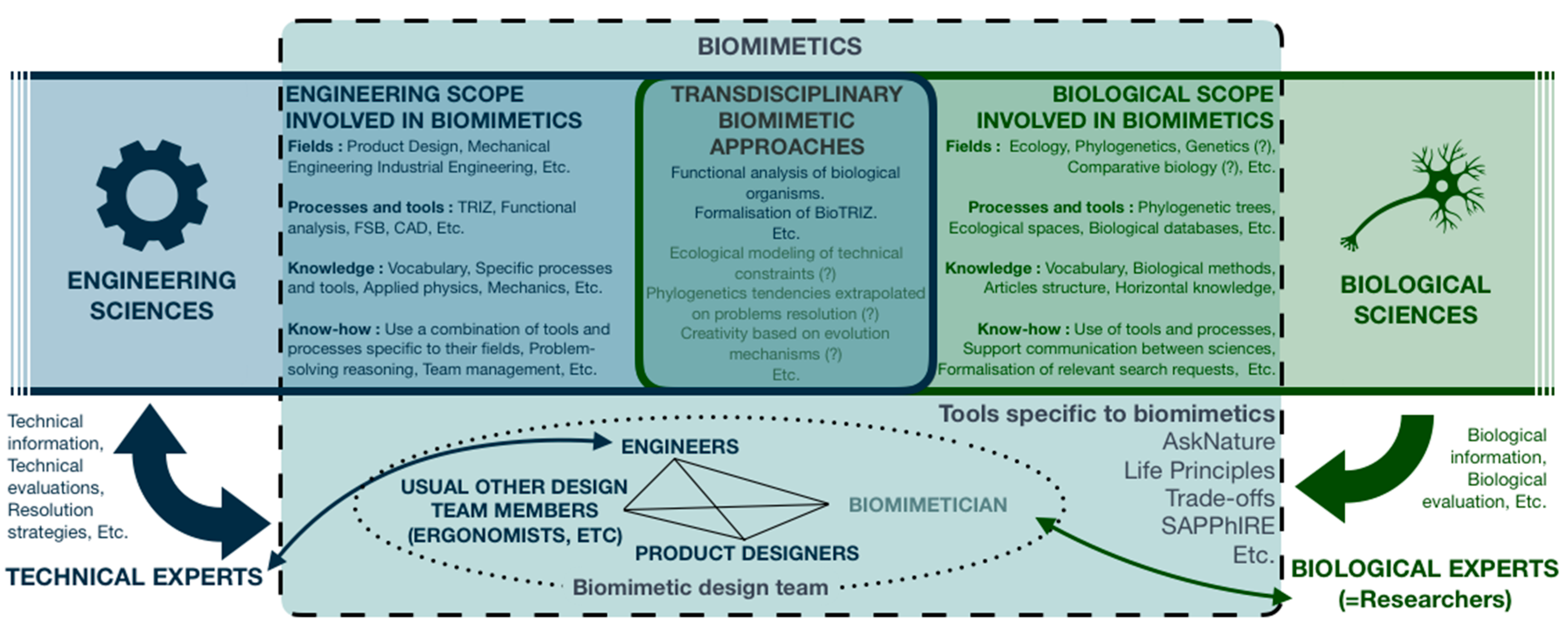
3.3. Impact on Biomimetician’s Definition and Training
- Who are biomimeticians?
- How are they going to be integrated within biomimetic teams?
- What to expect from them?
- How are they going to meet these expectations and so what is their knowledge and know-how?
4. Discussion
- We mainly studied abiotic environmental constraints (constraints that are not due to living beings), living aside ecological approaches investigating the interactions between biological systems (parasites and hosts, predators and preys, mutualistic symbiosis, etc.) which can also represent relevant identification pathways to explore.
- We assumed that biological data is always available in the literature, which isn’t the case. Biological classifications present species regardless of the amount of research performed after they were discovered. This potential lack of information appears as a strong impediment to anticipate. Tools that allow an initial screening of biological organisms based on their associated research effort could be a first step in addressing the problem.
- Since modern phylogenetic trees are mostly based on genetic distance without mentioning the implication on functional traits, new tools combining functional and ecological/phylogenetic classifications should represent a real step forward in their adaptation to biomimetic practices.
- One can argue that using taxonomic tools confines the search for models within a restricted branch of the tree of life. However, as they can bridge species based on their similarity of traits, and so on convergent evolution, polyphyletic groups should allow to expand solution spaces. Once a new species is identified, new mono or paraphyletic groups from other branches can then be described, expanding the searching area. Therefore, tools gathering studies on convergent evolution cross-referenced with functional, ecological, and phylogenetic criteria may be highly valuable.
- Tools from biology, whether they are ecological or phylogenetic tools, aren’t designed to be used in biomimetics. Hence, they aren’t optimized or ergonomics when used as biomimetic tools. A new generation of tools adapting those tools from biology to biomimetic practices should be of great interest.
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Technical Committee: ISO/TC 266 Biomimetics. ISO/TC266 18458:2015: Biomimetics–Terminology, Concepts and Methodology; ISO: Geneva, Switzerland, 2015; Available online: https://www.iso.org/standard/62500.html (accessed on 17 November 2020).
- Ahmed-Kristensen, S.; Christensen, B.T.; Lenau, T.A. Naturally original: Stimulating creative design through biological analogies and Random images. In Proceedings of the International Design Conference, DESIGN, Dubrovnik, Croatia, 19–22 May 2014; Volume 2014, pp. 427–436. [Google Scholar]
- Lenau, T.A.; Pigosso, D.C.A.; McAloone, T.; Lakhtakia, A. Biologically inspired design for environment. In Bioinspiration, Biomimetics, and Bioreplication X; International Society for Optics and Photonics: Bellingham, WA, USA, 2020; Volume 11374, p. 13. [Google Scholar] [CrossRef]
- Rovalo, E.; McCardle, J. Performance Based Abstraction of Biomimicry Design Principles using Prototyping. Designs 2019, 3, 38. [Google Scholar] [CrossRef]
- Fayemi, P.-E.; Wanieck, K.; Zollfrank, C.; Maranzana, N.; Aoussat, A. Biomimetics: Process, tools and practice. Bioinspir. Biomim. 2017, 12, 11002. [Google Scholar] [CrossRef] [PubMed]
- Kruiper, R.; Vincent, J.F.V.; Abraham, E.; Soar, R.; Konstas, I.; Chen-Burger, J.; Desmulliez, M. Towards a Design Process for Computer-Aided Biomimetics. Biomimetics 2018, 3, 14. [Google Scholar] [CrossRef]
- Snell-Rood, E. Interdisciplinarity: Bring biologists into biomimetics. Nature 2016, 529, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Graeff, E.; Maranzana, N.; Aoussat, A. Biomimetics, where are the biologists? J. Eng. Des. 2019, 30, 289–310. [Google Scholar] [CrossRef]
- Hashemi Farzaneh, H. Bio-inspired design: The impact of collaboration between engineers and biologists on analogical transfer and ideation. Res. Eng. Des. 2020, 1–24. [Google Scholar] [CrossRef]
- Yen, J.; Helms, M.E.; Goel, A.K.; Tovey, C.; Weissburg, M. Adaptive Evolution of Teaching Practices in Biologically Inspired Design. In Biologically Inspired Design; Springer: London, UK, 2014; pp. 153–199. [Google Scholar]
- Full, R.J. Using Biological Inspiration to Build Artificial Life That Locomotes. In Evolutionary Robotics; Gomi, T., Ed.; From Intelligent Robotics to Artificial Life. EvoRobots 2001. Lecture Notes in Computer Science; Springer: Berlin/Heidelberg, Germany, 2001; Volume 2217. [Google Scholar] [CrossRef]
- Lindemann, U.; Gramann, J. Engineering Design Using Biological Principles. In Proceedings of the International Design Conference, DESIGN, Dubrovnik, Croatia, 19–22 May 2004; Volume 2014, pp. 355–360. [Google Scholar]
- Lenau, T.A. Biomimetics as a design methodology—Possibilities and challenges. In Proceedings of the International Conference on Engineering Design, Standford, CA, USA, 24–27 August 2009; Volume 5, pp. 121–132. [Google Scholar]
- Goel, A.K.; Vattam, S.; Wiltgen, B.; Helms, M. Information-Processing Theories of Biologically Inspired Design. In Biologically Inspired Design; Springer: London, UK, 2014; pp. 127–152. ISBN 978-1-4471-5247-7. [Google Scholar]
- Fayemi, P.-E.; Maranzana, N.; Aoussat, A.; Bersano, G. Bio-inspired design characterisation and its links with problem solving tools. In Proceedings of the International Design Conference, DESIGN, Dubrovnik, Croatia, 19–22 May 2014; Volume 2014, pp. 173–182. [Google Scholar]
- Wanieck, K.; Fayemi, P.-E.; Maranzana, N.; Zollfrank, C.; Jacobs, S.R. Biomimetics and its tools. Bioinspir. Biomim. Nanobiomater. 2017, 6, 53–66. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Sarkar, P.; Leelavathamma, B.; Nataraju, B.S. A functional representation for aiding biomimetic and artificial inspiration of new ideas. Artif. Intell. Eng. Des. Anal. Manuf. AIEDAM 2005, 19, 113–132. [Google Scholar] [CrossRef]
- Nagel, J.K.S.; Nagel, R.L.; Stone, R.B.; McAdams, D.A. Function-based, biologically inspired concept generation. Artif. Intell. Eng. Des. Anal. Manuf. AIEDAM 2010, 24, 521–535. [Google Scholar] [CrossRef]
- Fayemi, P.-E. Innovation through Bio-Inspired Design: Suggestion of a Structuring Model for Biomimetic Process and Methods; Arts et Métiers Institute of Technology Hesam University: Paris, France, 2016. [Google Scholar]
- Ohno, T. Toyota Production System: Beyond Large-Scale Production, 1st ed.; Productivity Press: Cambridge, MA, USA, 1978; Volume 1. [Google Scholar]
- Altshuller, G.S. Creativity as an Exact Science: The Theory of the Solution of Inventive Problems; Gordon and Breach Science Publishers: New York, NY, USA, 1984; ISBN 0677212305. [Google Scholar]
- Hoagland, M.B.; Dodson, B. The Way Life Works, 1st ed.; Crown: New York, NY, USA, 1995; ISBN 978-0-8129-2020-8. [Google Scholar]
- Tinsley, A.; Midha, P.A.; Nagel, R.L.; McAdams, D.A.; Stone, R.B.; Shu, L.H. Exploring the use of functional models as a foundation for biomimetic conceptual design. In Proceedings of the ASME 2007 International Design Engineering Technical Conferences and Computers and Information in Engineering Conference, Las Vegas, NV, USA, 4–7 September 2007; Volume 3 Pt A, pp. 1–15. [Google Scholar] [CrossRef]
- Vincent, J.F.V.; Bogatyreva, O.A.; Bogatyrev, N.R.; Bowyer, A.; Pahl, A.K. Biomimetics: Its practice and theory. J. R. Soc. Interface 2006, 3, 471–482. [Google Scholar] [CrossRef]
- Vattam, S.S.; Goel, A.K. Seeking bioinspiration online: A descriptive account. In Proceedings of the International Conference on Engineering Design, Seoul, Korea, 19–22 August 2013; Volume 7, pp. 347–356. [Google Scholar]
- Wolff, J.O.; Wells, D.; Reid, C.R.; Blamires, S.J. Clarity of objectives and working principles enhances the success of biomimetic programs. Bioinspir. Biomim. 2017, 12, 051001. [Google Scholar] [CrossRef] [PubMed]
- Chirazi, J.; Wanieck, K.; Fayemi, P.-E.; Zollfrank, C.; Jacobs, S.R. What Do We Learn from Good Practices of Biologically Inspired Design in Innovation? Appl. Sci. 2019, 9, 650. [Google Scholar] [CrossRef]
- Biomimicry Institute AskNature—Innovation Inspired by Nature. Available online: https://asknature.org/ (accessed on 23 November 2018).
- Vattam, S.S.; Wiltgen, B.; Helms, M.E.; Goel, A.K.; Yen, J. DANE: Fostering Creativity in and through Biologically Inspired Design. In Design Creativity 2010; Springer: London, UK, 2011; pp. 115–122. [Google Scholar]
- Sartori, J.; Pal, U.; Chakrabarti, A. A methodology for supporting “transfer” in biomimetic design. Artif. Intell. Eng. Des. Anal. Manuf. AIEDAM 2010, 24, 483–505. [Google Scholar] [CrossRef]
- Vattam, S.S.; Goel, A.K. Foraging for Inspiration: Understanding and Supporting the Online Information Seeking Practices of Biologically Inspired Designers. In Proceedings of the ASME 2011 International Design Engineering Technical Conferences and Computers and Information in Engineering Conference, Washington, DC, USA, 28–31 August 2011; pp. 177–186. [Google Scholar] [CrossRef]
- Vandevenne, D.; Verhaegen, P.-A.; Dewulf, S.; Duflou, J.R. SEABIRD: Scalable search for systematic biologically inspired design. Artif. Intell. Eng. Des. Anal. Manuf. AIEDAM 2016, 30, 78–95. [Google Scholar] [CrossRef]
- Shu, L.H. A natural-language approach to biomimetic design. Artif. Intell. Eng. Des. Anal. Manuf. AIEDAM 2010, 24, 507–519. [Google Scholar] [CrossRef]
- Kruiper, R.; Vincent, J.F.V.; Chen-Burger, J.; Desmulliez, M.P.Y.; Konstas, I. A Scientific Information Extraction Dataset for Nature Inspired Engineering. arXiv 2020, arXiv:2005.07753. [Google Scholar]
- McInerney, S.; Khakipoor, B.; Garner, A.; Houette, T.; Unsworth, C.; Rupp, A.; Weiner, N.; Vincent, J.F.V.; Nagel, J.K.S.; Niewiarowski, P. E2BMO: Facilitating User Interaction with a BioMimetic Ontology via Semantic Translation and Interface Design. Designs 2018, 2, 53. [Google Scholar] [CrossRef]
- Vincent, J.F.V.; Cavallucci, D. Development of an ontology of biomimetics based on altshuller’s matrix. In International TRIZ Future Conference; IFIP Advances in Information and Communication Technology; Springer: Cham, Switzerland, 2018; Volume 541, pp. 14–25. [Google Scholar]
- Deldin, J.-M.; Schuknecht, M. The AskNature Database: Enabling Solutions in Biomimetic Design. In Biologically Inspired Design; Springer: London, UK, 2014; pp. 17–27. ISBN 978-1-4471-5248-4. [Google Scholar]
- Siddharth, L.; Chakrabarti, A. Evaluating the impact of Idea-Inspire 4.0 on analogical transfer of concepts. Artif. Intell. Eng. Des. Anal. Manuf. AIEDAM 2018, 32, 431–448. [Google Scholar] [CrossRef]
- Chiu, I.; Shu, L.H. Biomimetic design through natural language analysis to facilitate cross-domain information retrieval. Artif. Intell. Eng. Des. Anal. Manuf. 2007, 21, 45–59. [Google Scholar] [CrossRef]
- Nagel, J.K.S. A Thesaurus for Bioinspired Engineering Design. In Biologically Inspired Design; Goel, A., McAdams, D., Stone, R., Eds.; Springer: London, UK, 2014; pp. 63–94. [Google Scholar] [CrossRef]
- Stroble, J.K.; Stone, R.B.; McAdams, D.A.; Watkins, S.E. An Engineering-to-Biology Thesaurus To Promote Better Collaboration, Creativity and Discovery. In Proceedings of the 19th CIRP Design Conference–Competitive Design, Cranfield University, Bedford, UK, 30–31 March 2009; p. 355. [Google Scholar]
- Bogatyrev, N.; Bogatyreva, O. TRIZ-based algorithm for Biomimetic design. Procedia Eng. 2015, 131, 377–387. [Google Scholar] [CrossRef]
- Vincent, J.F.V. The trade-off: A central concept for biomimetics. Bioinspir. Biomim. Nanobiomater. 2017, 6, 67–76. [Google Scholar] [CrossRef]
- Kaiser, M.K.; Hashemi Farzaneh, H.; Lindemann, U. Bioscrabble—The role of different types of search terms when searching for biological inspiration in biological research articles. In Proceedings of the International Design Conference, DESIGN, Dubrovnik, Croatia, 19–22 May 2014; Volume 2014, pp. 241–250. [Google Scholar]
- Helms, K.M.; Hashemi Farzaneh, H.; Lindemann, U. Creating bio-inspired solution ideas using biological research articles. In Multidisciplinary Contributions to the Science of Creative Thinking; Springer: Singapore, 2015; pp. 215–232. ISBN 9789812876188. [Google Scholar]
- Helms, M.; Goel, A.K. The Four-Box Method of Problem Specification and Analogy Evaluation in Biologically Inspired Design. In Proceedings of the ASME 2014 International Design Engineering Technical Conferences and Computers and Information in Engineering Conference (ASME), Buffalo, NY, USA, 17–20 August 2014. [Google Scholar] [CrossRef]
- Badarnah, L.; Kadri, U. A methodology for the generation of biomimetic design concepts. Archit. Sci. Rev. 2015, 58, 120–133. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, J.H. A study on metadata structure and recommenders of biological systems to support bio-inspired design. Eng. Appl. Artif. Intell. 2017, 57, 16–41. [Google Scholar] [CrossRef]
- Helms, M.E.; Vattam, S.S.; Goel, A.K. Biologically inspired design: Process and products. Des. Stud. 2009, 30, 606–622. [Google Scholar] [CrossRef]
- Kennedy, E.; Niewiarowski, P. Biomimicry: Do Frames of Inquiry Support Search and Identification of Biological Models? Designs 2018, 2, 27. [Google Scholar] [CrossRef]
- Fayemi, P.-E.; Maranzana, N.; Aoussat, A.; Chekchak, T.; Bersano, G. Modeling biological systems to facilitate their selection during a bio-inspired design process. In Proceedings of the International Conference on Engineering Design, ICED, Milan, Italy, 27–30 July 2015; Volume 2, pp. 225–234. [Google Scholar]
- Bar-Cohen, Y. Biomimetics - Using nature to inspire human innovation. Bioinspir. Biomim. 2006, 1, P1–P12. [Google Scholar] [CrossRef]
- Vincent, J.F.V. Biomimetics—A review. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2009, 223, 919–939. [Google Scholar] [CrossRef]
- Fish, F.E.; Beneski, J.T. Evolution and Bio-Inspired Design: Natural Limitations. In Biologically Inspired Design; Goel, A., McAdams, D., Stone, R., Eds.; Springer: London, UK, 2014; pp. 287–312. [Google Scholar] [CrossRef]
- Gebeshuber, I.C.; Drack, M. An attempt to reveal synergies between biology and mechanical engineering. Proc. Inst. Mech. Eng. Part C J. Mech. Eng. Sci. 2008, 222, 1281–1287. [Google Scholar] [CrossRef]
- Drack, M.; Limpinsel, M.; De Bruyn, G.; Nebelsick, J.H.; Betz, O. Towards a theoretical clarification of biomimetics using conceptual tools from engineering design. Bioinspir. Biomim. 2018, 13, 016007. [Google Scholar] [CrossRef]
- Baumeister, D.; Tocke, R.; Dwyer, J.; Ritter, S.; Benyus, J. Biomimicry Resource Handbook: A Seed Bank of Knowledge and Best Practices, 2013rd ed.; CreateSpace Independent Publishing Platform: Missoula, MT, USA, 2013; ISBN 978-1505634648. [Google Scholar]
- McGregor, S.L.T. Transdisciplinarity and Biomimicry. Transdiscipl. J. Eng. Sci. 2013, 4. [Google Scholar] [CrossRef]
- Dallwitz, M.J. A general system for coding taxonomic descriptions. Taxon 1980, 29, 41–46. [Google Scholar] [CrossRef]
- Dallwitz, M.J.; Paine, T.A.; Zurcher, E.J. 1999 Onwards. User’s Guide to the DELTA Editor. Version: 22nd August 2020. Available online: https://www.delta-intkey.com/www/uguide.htm (accessed on 24 July 2020).
- Fauth, J.E.; Bernardo, J.; Camara, M.; Resetarits, W.J.; Van Buskirk, J.; McCollum, S.A. Simplifying the jargon of community ecology: A conceptual approach. Am. Nat. 1996, 147, 282–286. [Google Scholar] [CrossRef]
- Vincent, J.F.V. Smart by name, smart by nature. Smart Mater. Struct. 2000, 9, 255. [Google Scholar] [CrossRef]
- Yen, J.; Weissburg, M.J.; Helms, M.; Goel, A.K. Biologically inspired design: A tool for interdisciplinary education. In Biomimetics: Nature-Based Innovation; Bar-Cohen, Y., Ed.; Taylor & Francis: New York, NY, USA, 2011. [Google Scholar]
- Darwin, C. On the Origin of the Species, 1st ed.; John Murray: London, UK, 1859. [Google Scholar]
- Schöfer, M.; Maranzana, N.; Aoussat, A.; Bersano, G.; Buisine, S. Distinct and combined effects of disciplinary composition and methodological support on problem solving in groups. Creat. Innov. Manag. 2018, 27, 102–115. [Google Scholar] [CrossRef]
- Hashemi Farzaneh, H.; Helms, K.M.; Lindemann, U. Visual representations as a bridge for engineers and biologists in bio-inspired design collaborations. In Proceedings of the International Conference on Engineering Design, ICED, Milan, Italy, 27–30 July 2015; Volume 2, pp. 215–224. [Google Scholar]
- Graeff, E.; Maranzana, N.; Aoussat, A. Conception biomimétique: Quels acteurs pour quelles attentes? (Biomimetic Design: Which actors for what expectations?). In Proceedings of the CONFERE Conference, Budapest, Hungary, 5–6 July 2018. [Google Scholar]
- Graeff, E.; Maranzana, N.; Aoussat, A. Engineers’ and Biologists’ Roles during Biomimetic Design Processes, Towards a Methodological Symbiosis. In Proceedings of the International Conference on Engineering Design, ICED, Delft, The Netherlands, 5–8 August 2019; Volume 1, pp. 319–328. [Google Scholar]
- Keshwani, S.; Lenau, T.A.; Ahmed-Kristensen, S.; Chakrabarti, A. Comparing novelty of designs from biological-inspiration with those from brainstorming. J. Eng. Des. 2017, 28, 654–680. [Google Scholar] [CrossRef]
- Sane, S.P. Bioinspiration and biomimicry: What can engineers learn from biologists? J. Appl. Sci. Eng. 2016. [Google Scholar] [CrossRef]
- Blessing, L.T.M.; Chakrabarti, A. DRM, a Design Research Methodology; Springer: London, UK, 2009; pp. 1–397. ISBN 9781848825864. [Google Scholar]
- Fu, K.; Moreno, D.; Yang, M.; Wood, K.L. Bio-inspired design: An overview investigating open questions from the broader field of design-by-analogy. J. Mech. Des. Trans. ASME 2014, 136, 111102. [Google Scholar] [CrossRef]
- British Ecological Society Ecology Is a Science That Matters. Available online: https://www.britishecologicalsociety.org/about/what-is-ecology/ (accessed on 24 June 2019).
- Tansley, A.G. The Use and Abuse of Vegetational Concepts and Terms. Ecology 1935, 16, 284–307. [Google Scholar] [CrossRef]
- Design Council What Is the Framework for Innovation? Design Council’s Evolved Double Diamond|Design Council. Available online: https://www.designcouncil.org.uk/news-opinion/what-framework-innovation-design-councils-evolved-double-diamond (accessed on 26 February 2020).
- Kutschera, U.; Niklas, K.J. The modern theory of biological evolution: An expanded synthesis. Naturwissenschaften 2004, 91, 255–276. [Google Scholar] [CrossRef]
- Kutschera, U. Charles Darwin’s Origin of Species, directional selection, and the evolutionary sciences today. Naturwissenschaften 2009, 96, 1247–1263. [Google Scholar] [CrossRef]
- Climate Change & Infectious Diseases Group World Maps of Köppen-Geiger Climate Classification. Available online: http://koeppen-geiger.vu-wien.ac.at/ (accessed on 24 October 2019).
- Holdridge, L.R. Determination of world plant formations from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. Bioscience 2001, 51, 933. [Google Scholar] [CrossRef]
- Shattuck, J. Geography (Study of Science); Rosen Education Service: New York, NY, USA, 2016; p. 32. ISBN 150810428X. [Google Scholar]
- Olson, D.M.; Dinerstein, E. The Global 200: A Representation Approach to Conserving the Earth’s Most Biologically. Conserv. Biol. 1998, 12, 502–515. [Google Scholar] [CrossRef]
- Terpsichores Australasian Biomes. Available online: https://commons.wikimedia.org/wiki/File:Australasian_biomes.svg (accessed on 6 March 2020).
- Terpsichores Deserts and Xeric Shrublands. Available online: https://commons.wikimedia.org/wiki/File:Biome_map_13.svg (accessed on 6 March 2020).
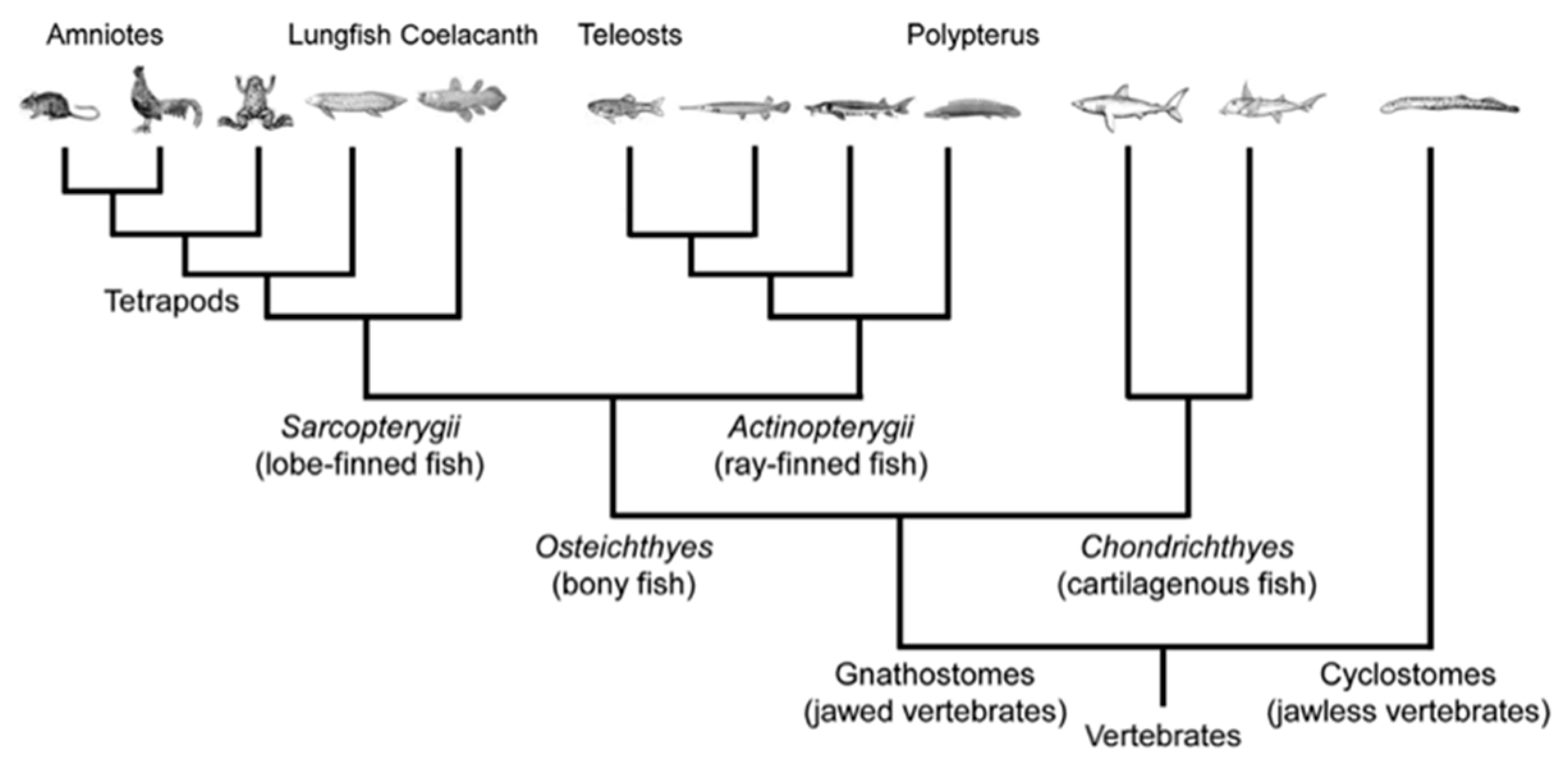
- Lecointre, G.; Le Guyader, H. The Tree of Life: A Phylogenetic Classification; Belknap Press of Harvard University Press: Cambridge, MA, USA, 2006; ISBN 9780674021839. [Google Scholar]
- Yamamoto, K.; Bloch, S.; Vernier, P. New perspective on the regionalization of the anterior forebrain in Osteichthyes. Dev. Growth Differ. 2017, 59, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Avibase Avibase—The World Bird Database. Available online: https://avibase.bsc-eoc.org/search.jsp?lang=EN&isadv=yes (accessed on 29 February 2020).
- SCAR Biodiversity Database. Available online: https://www.scar.org/data-products/bio-database/ (accessed on 29 February 2020).
- GBIF GBIF—Home Page. Available online: https://www.gbif.org/ (accessed on 1 July 2019).
- Linsey, J.S.; Viswanathan, V.K. Overcoming Cognitive Challenges in Bioinspired Design and Analogy. In Biologically Inspired Design; Springer: London, UK, 2014; pp. 221–244. [Google Scholar]
- Swanson, A.C.; Schwendenmann, L.; Allen, M.F.; Aronson, E.L.; Artavia-León, A.; Dierick, D.; Fernandez-Bou, A.S.; Harmon, T.C.; Murillo-Cruz, C.; Oberbauer, S.F.; et al. Welcome to the Atta world: A framework for understanding the effects of leaf-cutter ants on ecosystem functions. Funct. Ecol. 2019, 33, 1386–1399. [Google Scholar] [CrossRef]
- Fernandez-Bou, A.S.; Dierick, D.; Harmon, T.C. Diel pattern driven by free convection controls leaf-cutter ant nest ventilation and greenhouse gas emissions in a Neotropical rain forest. Oecologia 2020, 192, 591–601. [Google Scholar] [CrossRef]
- Bollazzi, M.; Forti, L.C.; Roces, F. Ventilation of the giant nests of Atta leaf-cutting ants: Does underground circulating air enter the fungus chambers? Insectes Soc. 2012, 59, 487–498. [Google Scholar] [CrossRef]
- Jones, J.C.; Oldroyd, B.P. Nest Thermoregulation in Social Insects. Adv. Insect Phys. 2006, 33, 153–191. [Google Scholar]
- Van Gils, H.A.J.A.; Vanderwoude, C. Leafcutter Ant (Atta sexdens) (Hymenoptera: Formicidae) Nest Distribution Responds to Canopy Removal and Changes in Micro-Climate in the Southern Colombian Amazon. Fla. Entomol 2012, 95, 914–921. [Google Scholar] [CrossRef]
- Bollazzi, M.; Roces, F. Leaf-cutting ant workers (Acromyrmex heyeri) trade off nest thermoregulation for humidity control. J. Ethol. 2010, 28, 399–403. [Google Scholar] [CrossRef]
- Lighton, J.; Berrigan, D. Questioning paradigms: Caste-specific ventilation in harvester ants, Messor pergandei and M. julianus (Hymenoptera: Formicidae). J. Exp. Biol. 1995, 198, 521–530. [Google Scholar]
- Lighton, J.R.B.; Wehner, R. Ventilation and respiratory metabolism in the thermophilic desert ant, Cataglyphis bicolor (Hymenoptera, Formicidae). J. Comp. Physiol. B 1993, 163, 11–17. [Google Scholar] [CrossRef]
- Korb, J. The architecture of termite mounds: A result of a trade-off between thermoregulation and gas exchange? Behav. Ecol. 1999, 10, 312–316. [Google Scholar] [CrossRef]
- Korb, J. Thermoregulation and ventilation of termite mounds. Naturwissenschaften 2003, 90, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Willot, Q.; Simonis, P.; Vigneron, J.P.; Aron, S. Total internal reflection accounts for the bright color of the saharan silver ant. PLoS ONE 2016, 11, e0152325. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Current Solutions | Advantages | Drawbacks | References |
|---|---|---|---|
| Databases & associated approaches | Functionally structured information Pre-identified content Ergonomics & ease of use | Low number of models Unbalanced number of models depending on the functional fields or the kingdom of life | [29,32,37,38] |
| Natural language web search (AI is still under development) | Based on the total amount of published biological information Very precise and scientific content can be found Already daily used tools (Google, etc.) | Difficulty of designing relevant requests Ill-structured data Very unprecise and erroneous content can be found A strong sorting step is required | [6,33,39,40,41] |
| Approaches based on highly abstracted principles | Bridge engineering and biology Ergonomics & ease of findability by changing the reasoning scale (highly abstracted solutions) | Difficulty of formulation of an abstracted request Difficulty of using raw abstracted solutions to generate technological innovations Potential fixations | [24,35,36,42,43] |
| Type of Carriers | Amount of Data | Use during the Process | Input for |
|---|---|---|---|
| Intermediary | Low | Identify solution spaces | Step 4 |
| Reference | High | Extract biological models of reference | Step 4 & 5 |
| Diversification | Intermediate | Supplement the models of reference | Step 4 & 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graeff, E.; Maranzana, N.; Aoussat, A. Biological Practices and Fields, Missing Pieces of the Biomimetics’ Methodological Puzzle. Biomimetics 2020, 5, 62. https://doi.org/10.3390/biomimetics5040062
Graeff E, Maranzana N, Aoussat A. Biological Practices and Fields, Missing Pieces of the Biomimetics’ Methodological Puzzle. Biomimetics. 2020; 5(4):62. https://doi.org/10.3390/biomimetics5040062
Chicago/Turabian StyleGraeff, Eliot, Nicolas Maranzana, and Améziane Aoussat. 2020. "Biological Practices and Fields, Missing Pieces of the Biomimetics’ Methodological Puzzle" Biomimetics 5, no. 4: 62. https://doi.org/10.3390/biomimetics5040062
APA StyleGraeff, E., Maranzana, N., & Aoussat, A. (2020). Biological Practices and Fields, Missing Pieces of the Biomimetics’ Methodological Puzzle. Biomimetics, 5(4), 62. https://doi.org/10.3390/biomimetics5040062