Oxidation of L-Ascorbic Acid in the Presence of the Copper-Binding Compound from Methanotrophic Bacteria Methylococcus capsulatus (M)


Abstract
1. Introduction
2. Materials and Methods
2.1. Cells Growth
2.2. Copper-Binding Compound Isolation
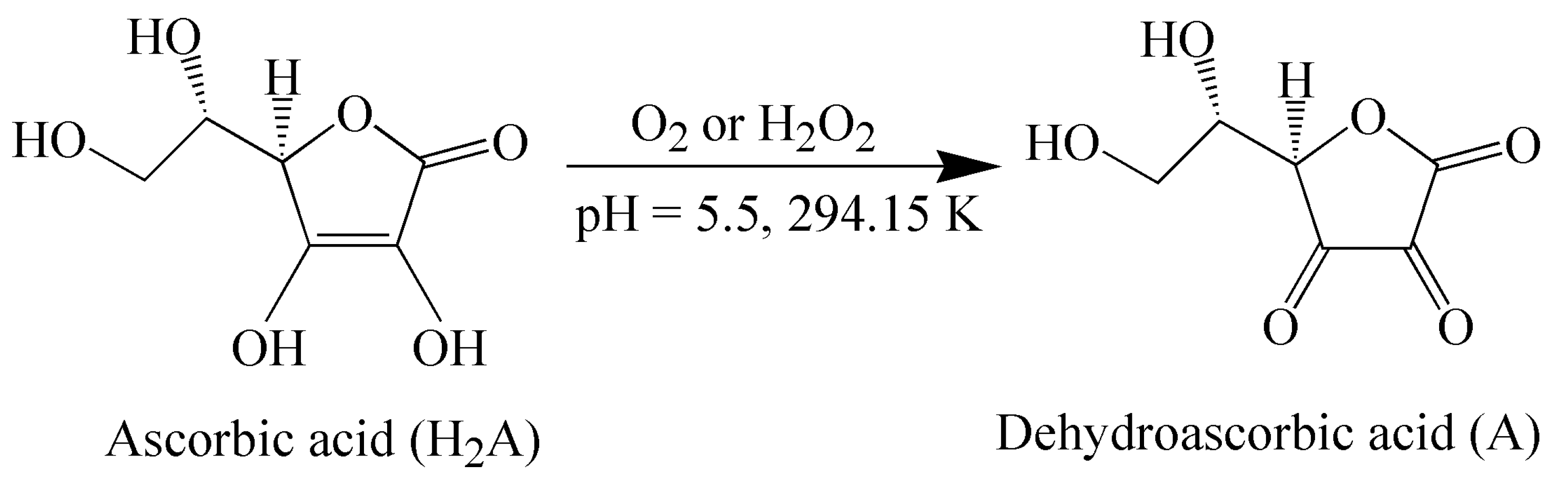
2.3. Oxidation of L-Ascorbic Acid
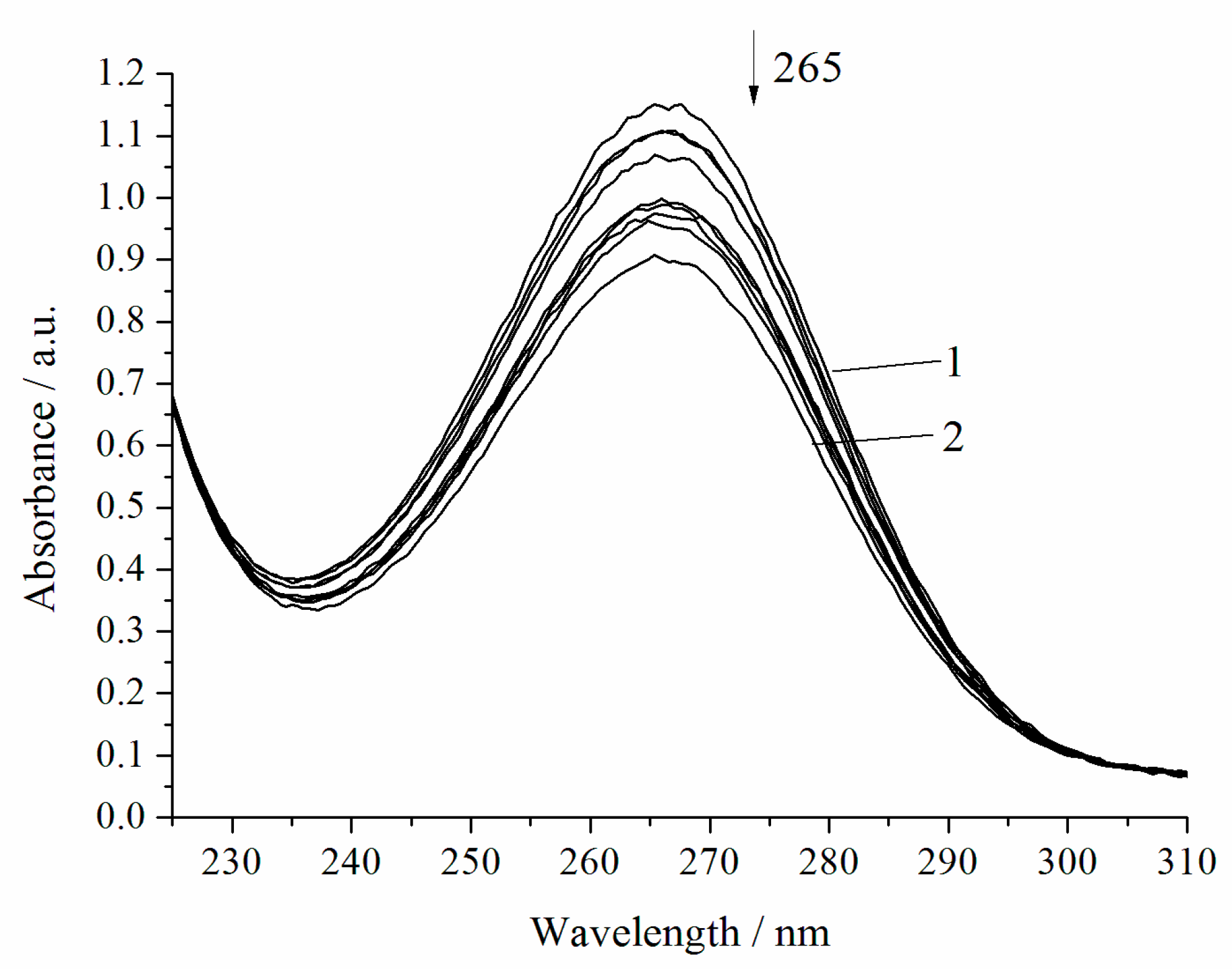
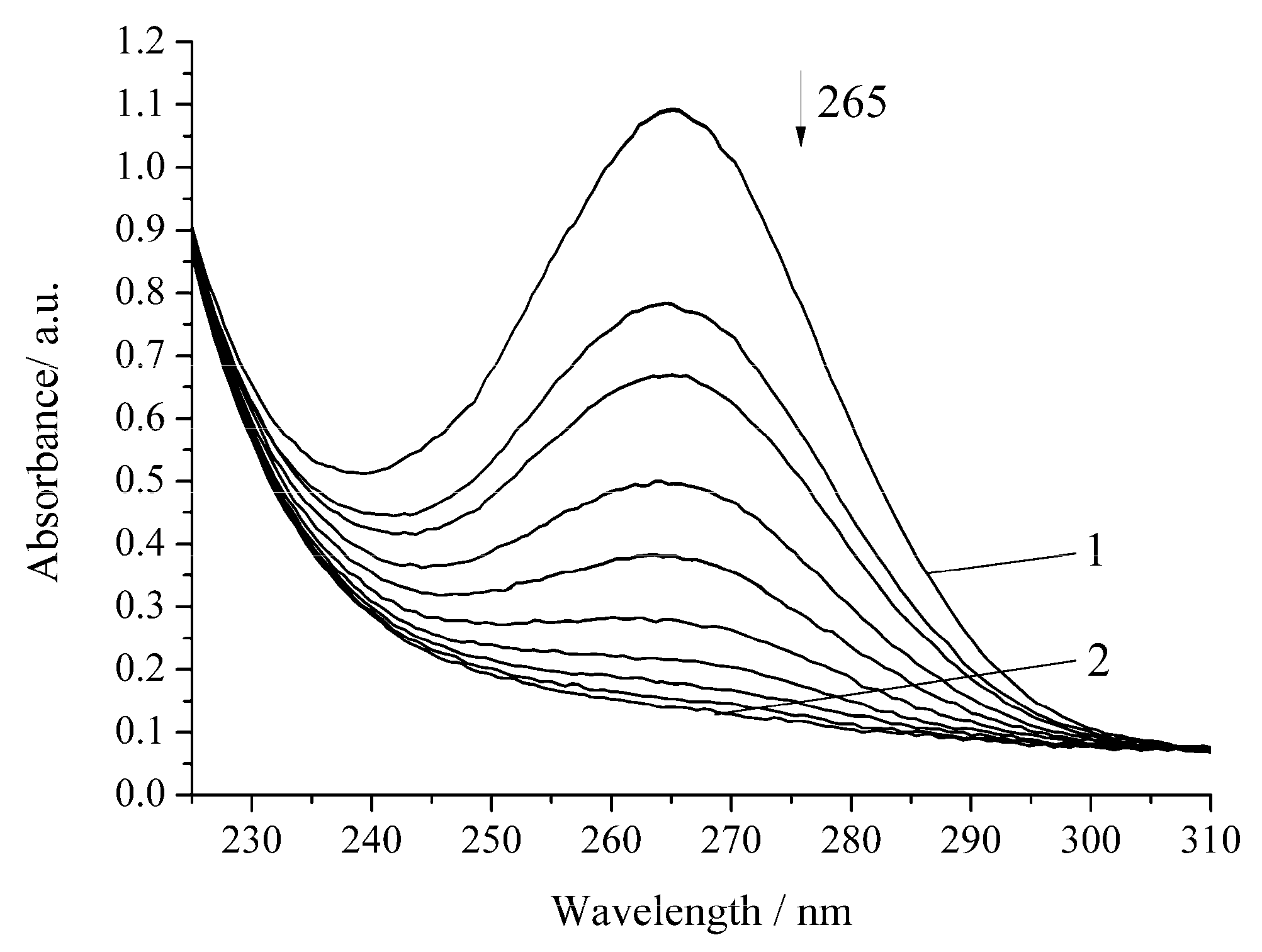
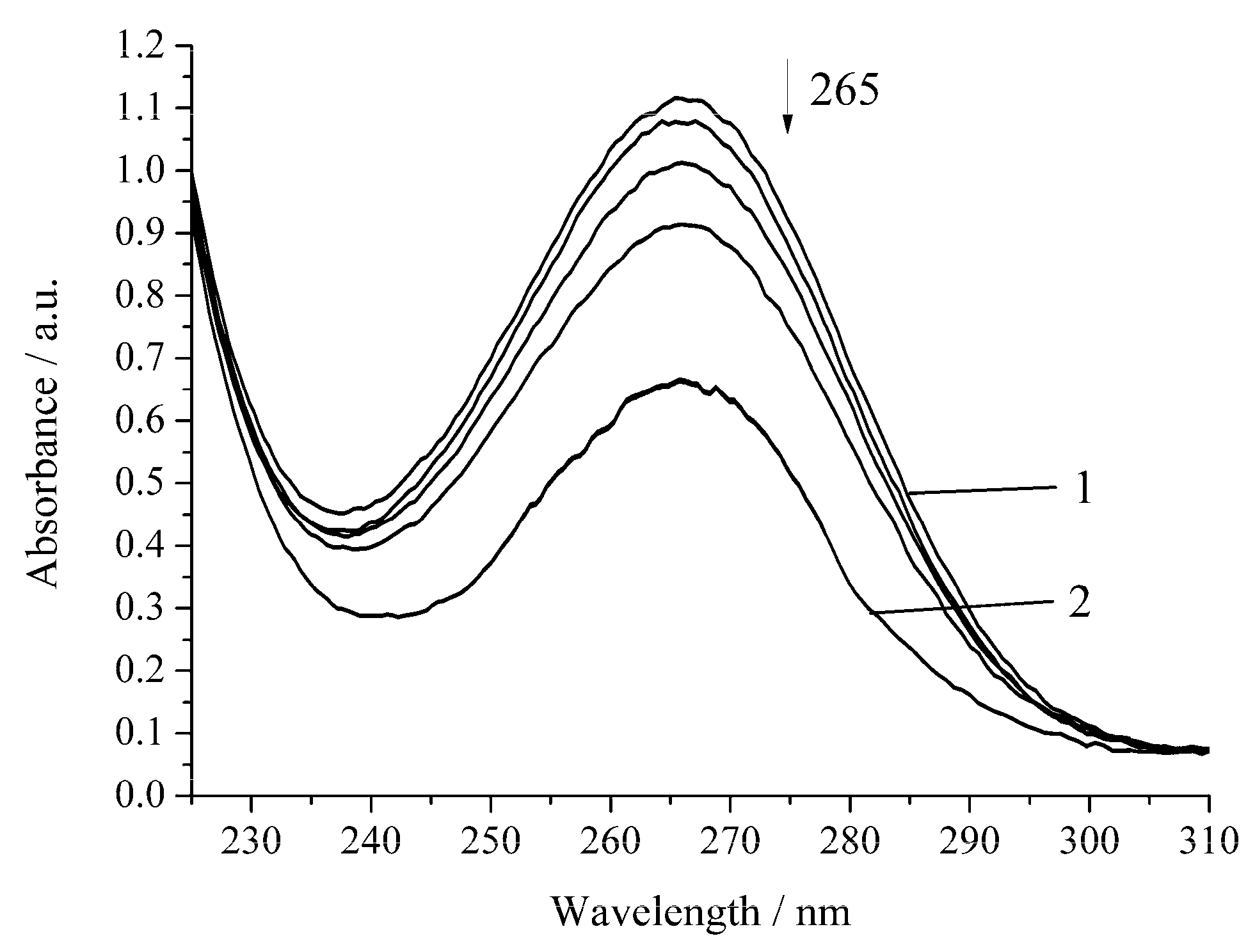
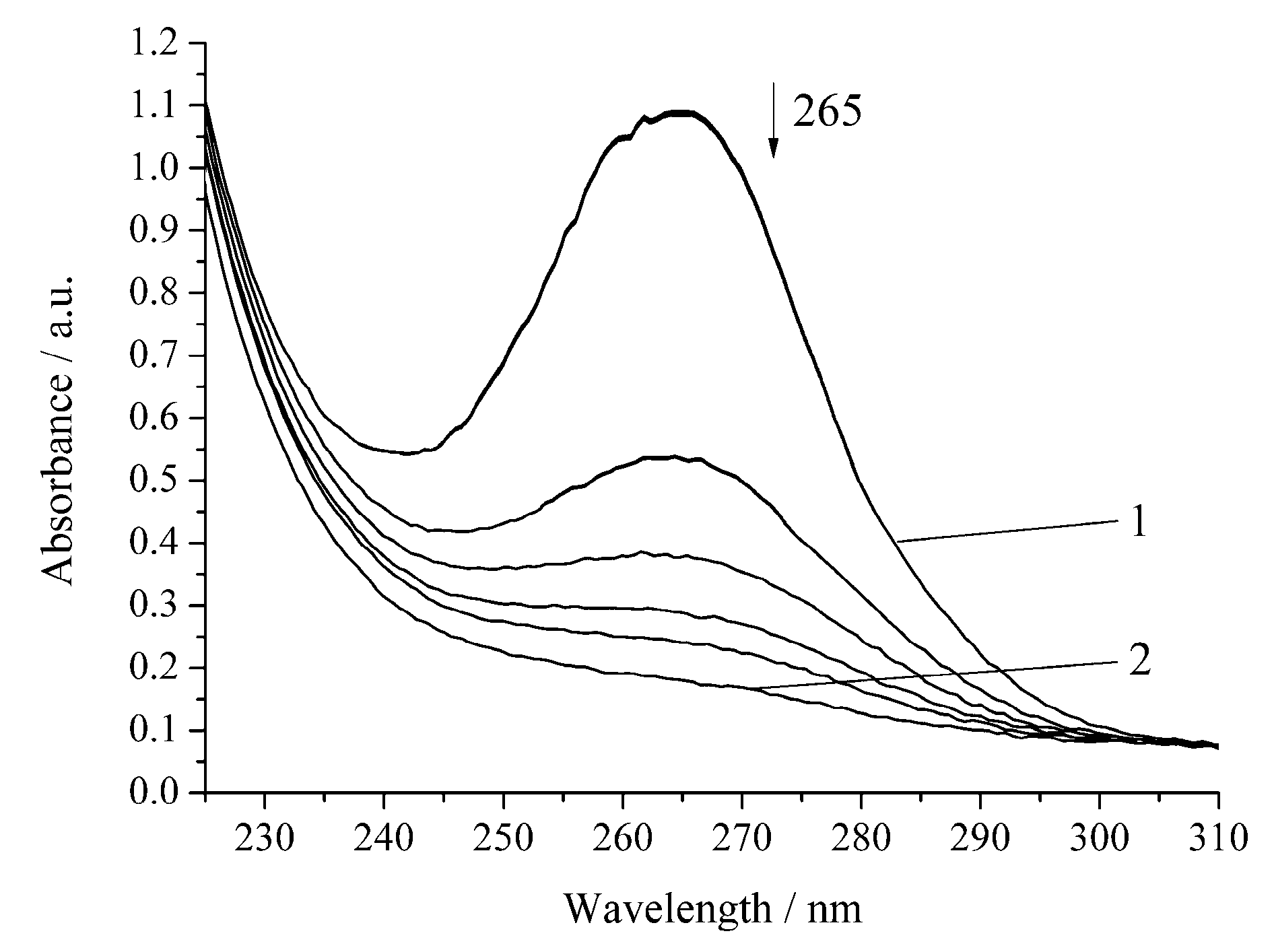
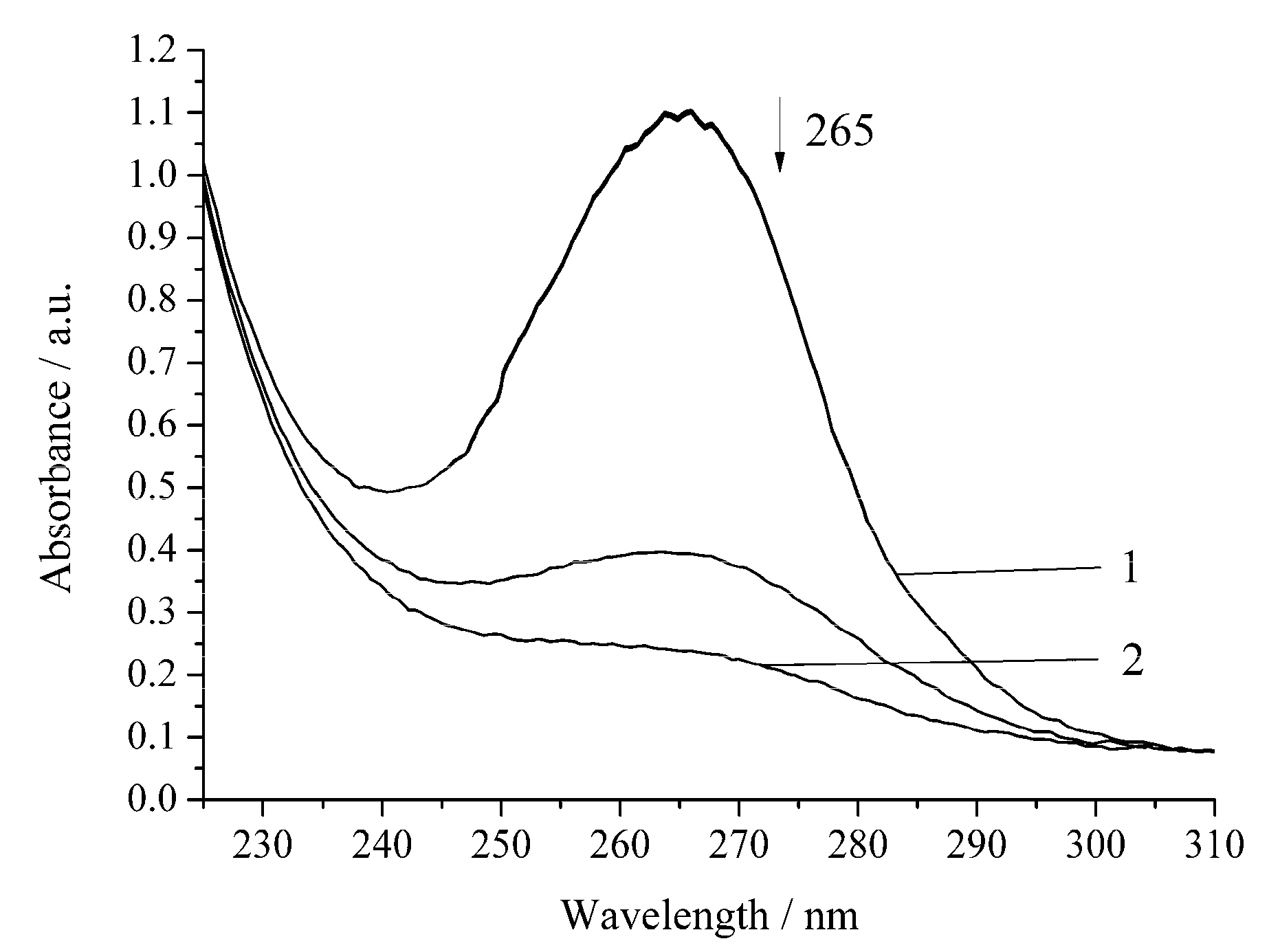
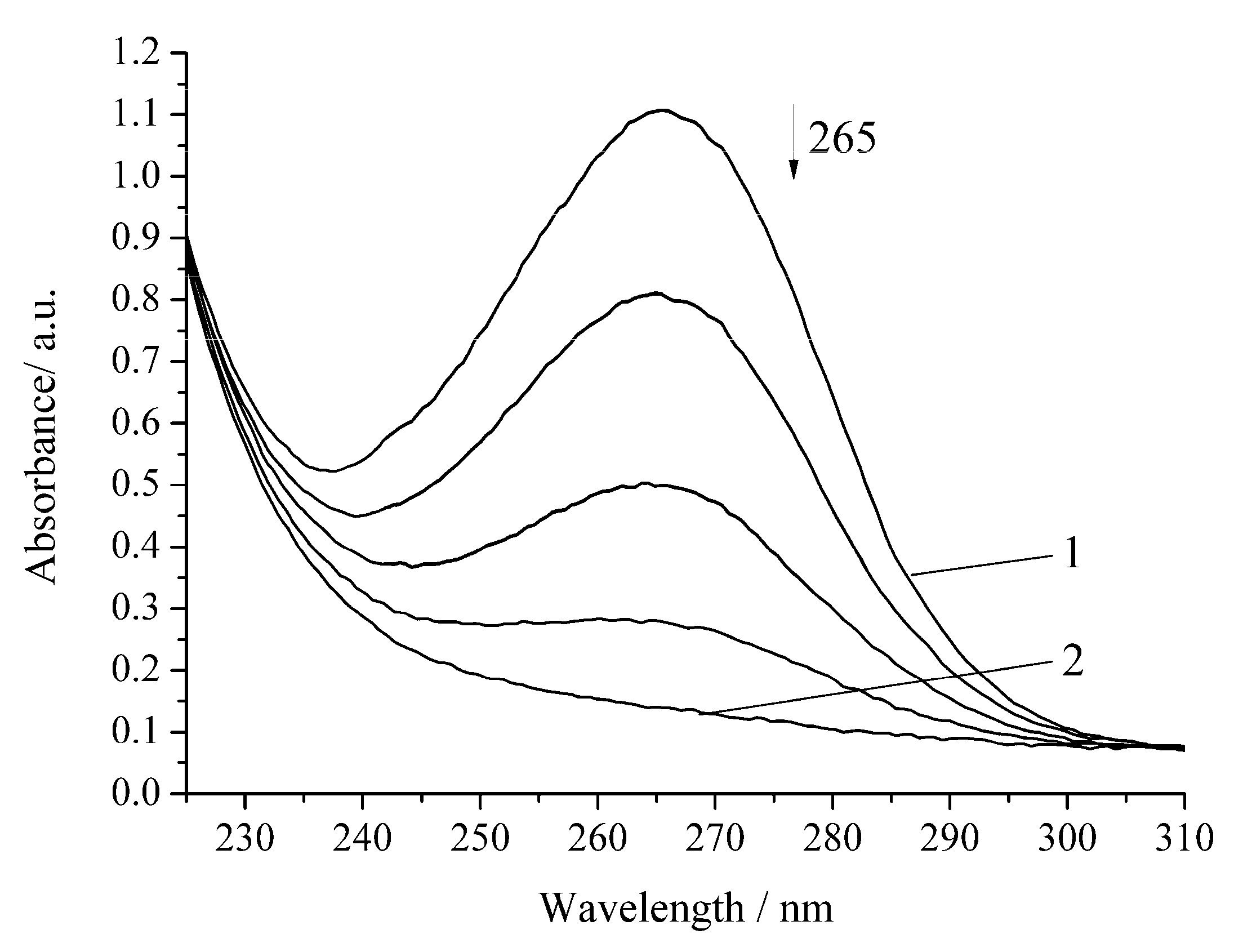
3. Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- DiSpirito, A.A.; Semrau, J.D.; Murrell, J.C.; Gallagher, W.H.; Dennison, C.; Vuilleumier, S. Methanobactin and the link between copper and bacterial methane oxidation. Microbiol. Mol. Biol. Rev. 2016, 80, 387–409. [Google Scholar] [CrossRef] [PubMed]
- Miyaji, A.; Suzuki, M.; Baba, T.; Kamachi, T.; Okura, I. Hydrogen peroxide as an effecter on the inactivation of particulate methane monooxygenase under aerobic conditions. J. Mol. Catal. B-Enzym. 2009, 57, 211–215. [Google Scholar] [CrossRef]
- Medvedkova, K.A.; Khmelenina, V.N.; Suzina, N.E.; Trotsenko, Y.A. Antioxidant systems of moderately thermophilic methanotrophs Methylocaldum szegediense and Methylococcus capsulatus. Microbiology 2009, 78, 670–677. [Google Scholar] [CrossRef]
- Kim, K.; Portis, A.R., Jr. Oxygen-dependent H2O2 production by Rubisco. FEBS Lett. 2004, 571, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Medvedkova, K.A.; Khmelenina, V.N.; Baskunov, B.P.; Trotsenko, Y.A. Synthesis of melanin by a moderately thermophilic methanotroph Methylocaldum szegediense depends on cultivation temperature. Microbiology 2008, 77, 112–114. [Google Scholar] [CrossRef]
- Choi, D.W.; Semrau, J.D.; Antholine, W.E.; Hartsel, S.C.; Anderson, R.C.; Carey, J.N.; Dreis, A.M.; Kenseth, E.M.; Renstrom, J.M.; Scardino, L.L.; et al. Oxidase, superoxide dismutase, and hydrogen peroxide reductase activities of methanobactin from types I and II methanotrophs. J. Inorg. Biochem. 2008, 102, 1571–1580. [Google Scholar] [CrossRef]
- Semrau, J.D.; DiSpirito, A.A.; Gu, W.; Yoon, S. Metals and methanotrophy. Appl. Environ. Microbiol. 2018, 84, e02289-17. [Google Scholar] [CrossRef]
- Khmelenina, V.N.; Rozova, O.N.; But, C.Y.; Mustakhimov, I.I.; Reshetnikov, A.S.; Beschastnyi, A.P.; Trotsenko, Y.A. Biosynthesis of secondary metabolites in methanotrophs: Biochemical and genetic aspects (Review). Appl. Biochem. Microbiol. 2015, 51, 150–158. [Google Scholar] [CrossRef]
- Choi, D.W.; Bandow, N.L.; McEllistrem, M.T.; Semrau, J.D.; Antholine, W.E.; Hartsel, S.C.; Gallagher, W.; Zea, C.J.; Pohl, N.L.; Zahn, J.A.; et al. Spectral and thermodynamic properties of methanobactin from γ-proteobacterial methane oxidizing bacteria: A case for copper competition on a molecular level. J. Inorg. Biochem. 2010, 104, 1240–1247. [Google Scholar] [CrossRef]
- Lu, X.; Gu, W.; Zhao, L.; Haque, M.F.U.; DiSpirito, A.A.; Semrau, J.D.; Gu, B. Methylmercury uptake and degradation by methanotrophs. Sci. Adv. 2017, 3, e1700041. [Google Scholar] [CrossRef]
- Zahn, J.A.; DiSpirito, A.A. Membrane-associated methane monooxygenase from Methylococcus capsulatus (Bath). J. Bacteriol. 1996, 178, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Antholine, W.E.; Do, Y.S.; Semrau, J.D.; Kisting, C.J.; Kunz, R.C.; Campbell, D.; Rao, V.; Hartsel, S.C.; DiSpirito, A.A. Effect of methanobactin on the activity and electron paramagnetic resonance spectra of the membrane-associated methane monooxygenase in Methylococcus capsulatus Bath. Microbiology 2005, 151, 3417–3426. [Google Scholar] [CrossRef] [PubMed]
- Avdeeva, L.V.; Tukhvatullin, I.A.; Gvozdev, R.I. Optimization cultivation of bacteria Methylococcus capsulatus (M) for production methanobactin. Nat. Tech. Sci. 2015, 11, 132–136. (In Russian) [Google Scholar]
- Avdeeva, L.; Tukhvatullin, I.; Gvozdev, R. Isolation and purification of methanobactin from Methylococcus capsulatus (M). FEBS J. 2013, 280, 101–102. [Google Scholar]
- Avdeeva, L.V.; Goryachev, N.S.; Gvozdev, R.I. Spectral-fluorescent properties of methanobactin from Methylococcus capsulatus (strain M). Acta Nat. 2016, 2, 95. (In Russian) [Google Scholar]
- Pozin, M.E. Hydrogen Peroxide and Peroxide Compounds; Goskhimizdat: Moscow, Russia, 1951; p. 475. (In Russian) [Google Scholar]
- Solomonov, A.V.; Rumyantsev, E.V.; Kochergin, B.A.; Antina, E.V. The kinetics of oxidation of bilirubin and ascorbic acid in solution. Russ. J. Phys. Chem. A 2012, 86, 1048–1052. [Google Scholar] [CrossRef]
- Dawson, R.; Elliot, D.; Elliot, W.; Jones, K. Data for Biochemical Research; Clarendon: Oxford, MS, USA, 1986; p. 429. [Google Scholar]
- Yatsimirsky, V.K.; Geraseva, V.G.; Bezuglaya, T.N.; Diyuk, V.E. Oxidation of ascorbic acid by molecular oxygen in the presence of heterobimetallic complexes of copper. Ukr. Chem. J. 2009, 75, 43–48. (In Russian) [Google Scholar]
- Miyake, N.; Kim, M.; Kurata, T. Formation mechanism of monodehydro-l-ascorbic acid and superoxide anion in the autoxidation of l-ascorbic acid. Biosci. Biotechnol. Biochem. 1997, 61, 1693–1695. [Google Scholar] [CrossRef][Green Version]
- Khan, M.M.T.; Martell, A.E. Metal ion and metal chelate catalyzed oxidation of ascorbic acid by molecular oxygen. I. Cupric and ferric ion catalyzed oxidation. J. Am. Chem. Soc. 1967, 89, 4176–4185. [Google Scholar] [CrossRef]
- Moya, H.D.; Coichev, N. Kinetic studies of the oxidation of L-ascorbic acid by tris(oxalate)cobaltate in the presence of CDTA metal ion complexes. J. Braz. Chem. Soc. 2006, 17, 364–368. [Google Scholar] [CrossRef]
- Khan, M.N.; Sarwar, A. The influence of transition metal ions on the kinetics of ascorbic acid oxidation by methylene blue in strongly acidic media. Turk. J. Chem. 2001, 25, 433–440. [Google Scholar]
- Sailani, R.; Dubey, S.; Khandelwal, C.L.; Sharma, P.D.; Khan, P. Kinetics and mechanism of oxidation of L-ascorbic acid by peroxomonosulphate in acid perchlorate medium. Role of copper (II) as a trace metal-ion catalyst. CR Chim. 2011, 14, 1088–1094. [Google Scholar] [CrossRef]
- Mishra, D.K.; Dhas, T.P.A.; Sharma, P.D.; Bhargava, A.P.; Gupta, Y.K. Role of trace metal ions. Kinetics and mechanism of the copper(II)-catalysed oxidation of ascorbic acid with peroxodiphosphate in acetate buffers. J. Chem. Soc. Dalton Trans. 1990, 4, 1265–1270. [Google Scholar] [CrossRef]
- Zhang, G.-F.; Chen, H.-Y. Chemiluminescence studies of the oxidation of ascorbic acid with copper(II) catalyzed by halide anions and its application to the determination of halide anions and ascorbic acid. Anal. Sci. 2000, 16, 1317–1321. [Google Scholar] [CrossRef]
- Bayr Hülya, M.D. Reactive oxygen species. Crit. Care Med. 2005, 33, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.B. Reactions of L-ascorbic acid with transition metal complexes. Polyhedron 1992, 11, 285–321. [Google Scholar] [CrossRef]
- Maksimov, A.L.; Kardasheva, Y.S.; Predeina, V.V.; Kluev, M.V.; Ramazanov, D.N.; Talanova, M.Y.; Karakhanov, E.A. Iron and copper complexes with nitrogen-containing ligands as catalysts for cyclohexane oxidation with hydrogen peroxide under mild reaction conditions. Petrol. Chem. 2012, 52, 318–326. [Google Scholar] [CrossRef]
- Shul’pina, L.S.; Takaki, K.; Strelkova, T.V.; Shul’pin, G.B. Benzene oxidation with hydrogen peroxide catalyzed by soluble and heterogenized copper compounds. Petrol. Chem. 2008, 48, 219–222. [Google Scholar] [CrossRef]
- Avdeeva, L.; Gvozdev, R. Effect of copper concentration on the growth of Methylococcus capsulatus (strain M). Chem. J. Mold. 2017, 12, 110–114. [Google Scholar] [CrossRef]
- Ward, N.; Larsen, Q.; Sakwa, J.; and 39 coauthors. Genomic insights into methanotrophy: The complete genome sequence of Methylococcus capsulatus (Bath). PLoS Biol. 2004, 2, 1616–1628. [Google Scholar] [CrossRef]
- Zahn, J.A.; Arciero, D.M.; Hooper, A.B.; Coats, J.R.; Dispirito, A.A. Cytochrome c peroxidase from Methylococcus capsulatus Bath. Arch. Microbiol. 1997, 168, 362–372. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | kobs × 10−4 (s−1) | t1/2 (s) | wH2A × 10−8 (mol L−1 s−1) |
|---|---|---|---|
| H2A + Air | 1.99 ± 0.14 | 3497.49 | 1.34 ± 0.09 |
| H2A + Air + Cu-cbc | 3.03 ± 0.15 | 2287.13 | 1.72 ± 0.08 |
| H2A + Air +Cu(II) | 48.93 ± 4.91 | 141.63 | 14.41 ± 1.45 |
| H2A + H2O2 | 18.47 ± 1.29 | 375.20 | 4.75 ± 0.12 |
| H2A + H2O2 + Cu-cbc | 11.57 ± 0.57 | 598.96 | 3.96 ± 0.19 |
| H2A + H2O2 + Cu(II) | 98.35 ± 9.87 | 70.46 | 14.90 ± 1.51 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avdeeva, L.V.; Gvozdev, R.I. Oxidation of L-Ascorbic Acid in the Presence of the Copper-Binding Compound from Methanotrophic Bacteria Methylococcus capsulatus (M). Biomimetics 2020, 5, 48. https://doi.org/10.3390/biomimetics5040048
Avdeeva LV, Gvozdev RI. Oxidation of L-Ascorbic Acid in the Presence of the Copper-Binding Compound from Methanotrophic Bacteria Methylococcus capsulatus (M). Biomimetics. 2020; 5(4):48. https://doi.org/10.3390/biomimetics5040048
Chicago/Turabian StyleAvdeeva, Lidia V., and Rudolf I. Gvozdev. 2020. "Oxidation of L-Ascorbic Acid in the Presence of the Copper-Binding Compound from Methanotrophic Bacteria Methylococcus capsulatus (M)" Biomimetics 5, no. 4: 48. https://doi.org/10.3390/biomimetics5040048
APA StyleAvdeeva, L. V., & Gvozdev, R. I. (2020). Oxidation of L-Ascorbic Acid in the Presence of the Copper-Binding Compound from Methanotrophic Bacteria Methylococcus capsulatus (M). Biomimetics, 5(4), 48. https://doi.org/10.3390/biomimetics5040048