1. Introduction
Water in the atmosphere has been an elusive freshwater resource in arid regions of the globe. In their 2001 article, Parker and Lawrence [
1] suggested a biomimetic approach for harvesting water efficiently from the atmosphere. The arguments presented in that work relied on the morphology and wettability of a desert beetle’s back, both presumed to play a critical role in the water-droplet collection process. The article by Parker and Lawrence inspired numerous follow-up studies, many of them using biomimetic principles and arguments to motivate devices in the technological arena [
2,
3,
4]. Subsequent studies showed substantially improved water collection rates on inhomogeneous-wettability surfaces, as compared to flat or uniformly-wettable surfaces [
5,
6]. Similar effects were also observed in studies of dew condensation on desert beetles [
7].
Several comparative studies that evaluated fog basking among the more than 200 Namib Desert tenebrionid species, have found only two species—both Onymacris—to be fog baskers [
8].
Onymacris unguicularis is one of them; its dorsal surface (a.k.a. elytron) features uniform wettability [
9]. Nørgaard and Dacke [
9] reported that boundary layer structure may play a critical role in the fog-collection mechanism, and this particular possibility remains unexplored. Their work pointed to the importance of the boundary layer structure around the body of the beetle and its effects on the transport mechanisms under realistic fog-basking conditions. They observed that during fog basking the
O. ungucularis beetle takes a head stance, so that the ventral side of the animal is at an angle of ∼23 deg to the horizontal. Such posture was consistently observed in experiments conducted with live beetles (see
Figure 1). The significance of such a head stance from the fluid mechanics perspective is intriguing. Here, we carry out a numerical study to characterize the flow properties around a simplified geometry (semi-ellipsoid) resembling that of the
O. unguicularis beetle. Specifically, the objective is to understand whether the posture assumed by the beetle to capture water droplets can be at least partly explained using fluid mechanical arguments, for example, is there a relation between the head stance and the three-dimensional boundary layer developing around the body, which maximizes water collection? To answer these questions, we perform numerical simulations for different inclinations of the ellipsoid that mimics the head stance of the beetle, and monitor the corresponding collection efficiency (number of water droplets collected on the beetle surface) using a three-dimensional Navier–Stokes solver with two-phase flow capabilities to track water droplets.
2. Methods
All simulations were carried out in OpenFOAM (open source field operation and manipulation), an open source C++ toolbox for developing numerical solvers for various problems in the field of computational fluid dynamics [
10]. The OpenFOAM solver used for this study,
reactingParcelFilmFoam, is based on an Eulerian–Lagrangian two-phase flow modeling for the continuous phase (carrier fluid, air) and the dispersed phase (water droplets), respectively (see OpenFOAM documentation)
https://openfoam.org/resources. The two phases are coupled through a two-way coupling procedure in which the carrier fluid and the convected droplets interact with each other via drag forces. The continuous phase behavior is governed by the Navier–Stokes equations for an incompressible fluid,
where
are the fluid velocity vector, density and pressure, respectively,
is the kinematic viscosity of the fluid,
is the gravity acceleration vector, and
the local force exerted by the droplets on the fluid in the two-way coupling setting. We also evaluated the importance of this term in the calculation of the overall mass collected on the body’s surface using a one-way coupled setting (see
Appendix A). Water droplets were tracked individually using the point-particle approximation and their trajectories were obtained by integrating the following set of equations
where
is the droplet position vector,
the droplet velocity,
the droplet mass,
and
are the water density and droplet diameter, respectively, while
is the drag force exerted by the fluid on the droplet, which can be expressed as
where
is the relative velocity of the droplet with respect to the surrounding fluid,
is the droplet cross-sectional area, while
is the drag coefficient approximated by the Stokes drag formula,
, where
is the Reynolds number. This assumption is justified in the present calculations by the small droplet diameter and small relative velocity, which gives
. The governing Equations (
1) and (
2) were solved using the PIMPLE algorithm, which is a combination of two different algorithms, PISO (pressure-implicit split-operator) and SIMPLE (semi-implicit method for pressure linked equations). During each time step, we solved a pressure equation, to enforce mass conservation, within an explicit velocity correction to satisfy momentum conservation. The module
reactingParcelFilmFoam implemented in OpenFOAM was used to resolve Lagrangian particle motion and for the impingement of droplets on the surface of the body. To that end, a special external mesh region was extruded from the body’s surface. For modeling of the film layer, a thin-film approximation was used: the velocity normal to the wall was zero while, along the wall, the tangential diffusion was considered to be negligible compared to normal diffusion. The surface film flow was solved using the continuity and momentum equations. The continuity equation comprised of a source term representing the mass added to the film layer due to the incoming fog droplets (
)
where
is the density of the film fluid and
is the film thickness.
3. Results
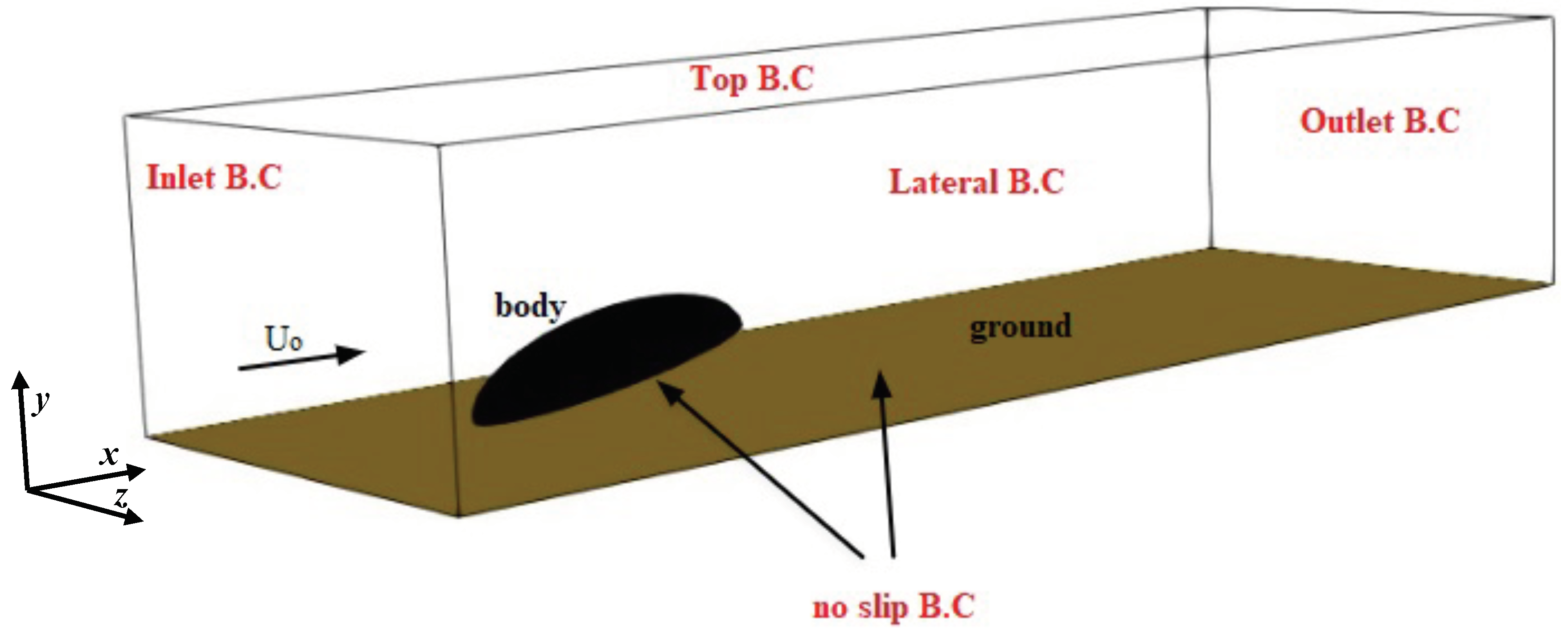
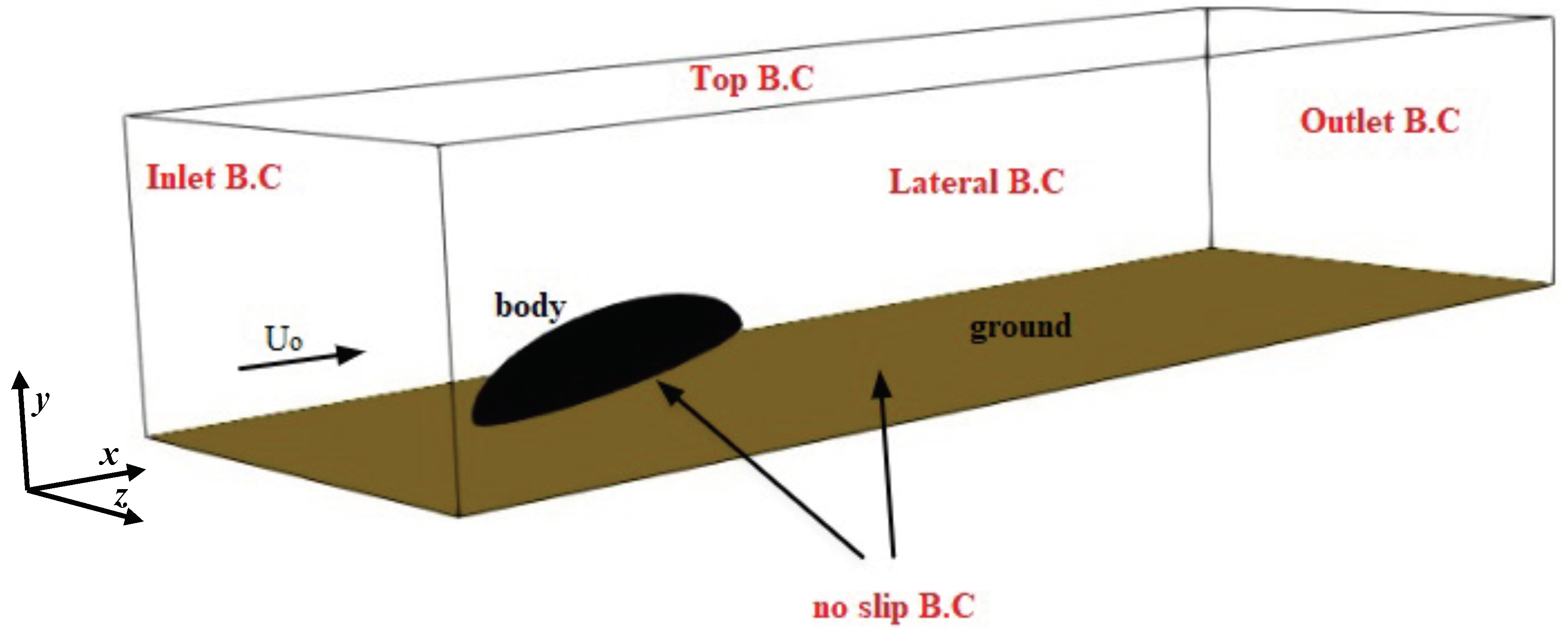
Based on the structure of the beetle’s body, a simplified geometry was chosen consisting of a semi-ellipsoid with axes of 8 mm, 3 mm, and 3 mm, in the horizontal (−x), vertical (−y) and transversal (−z) directions, respectively. The legs were not modeled due to their much lower surface area compared to the rest of the body. The semi-ellipsoid (body hereafter) was placed in a surrounding external box (cuboid shaped with side lengths
, and
) with inclination angle
with respect to the ground (see
Figure 2 for the representative case of
). Several grids were generated for each
with the total number of cells
, kept constant. For each case, we performed a grid independence study to ensure the grid was fine enough to capture the flow characteristics (not shown).
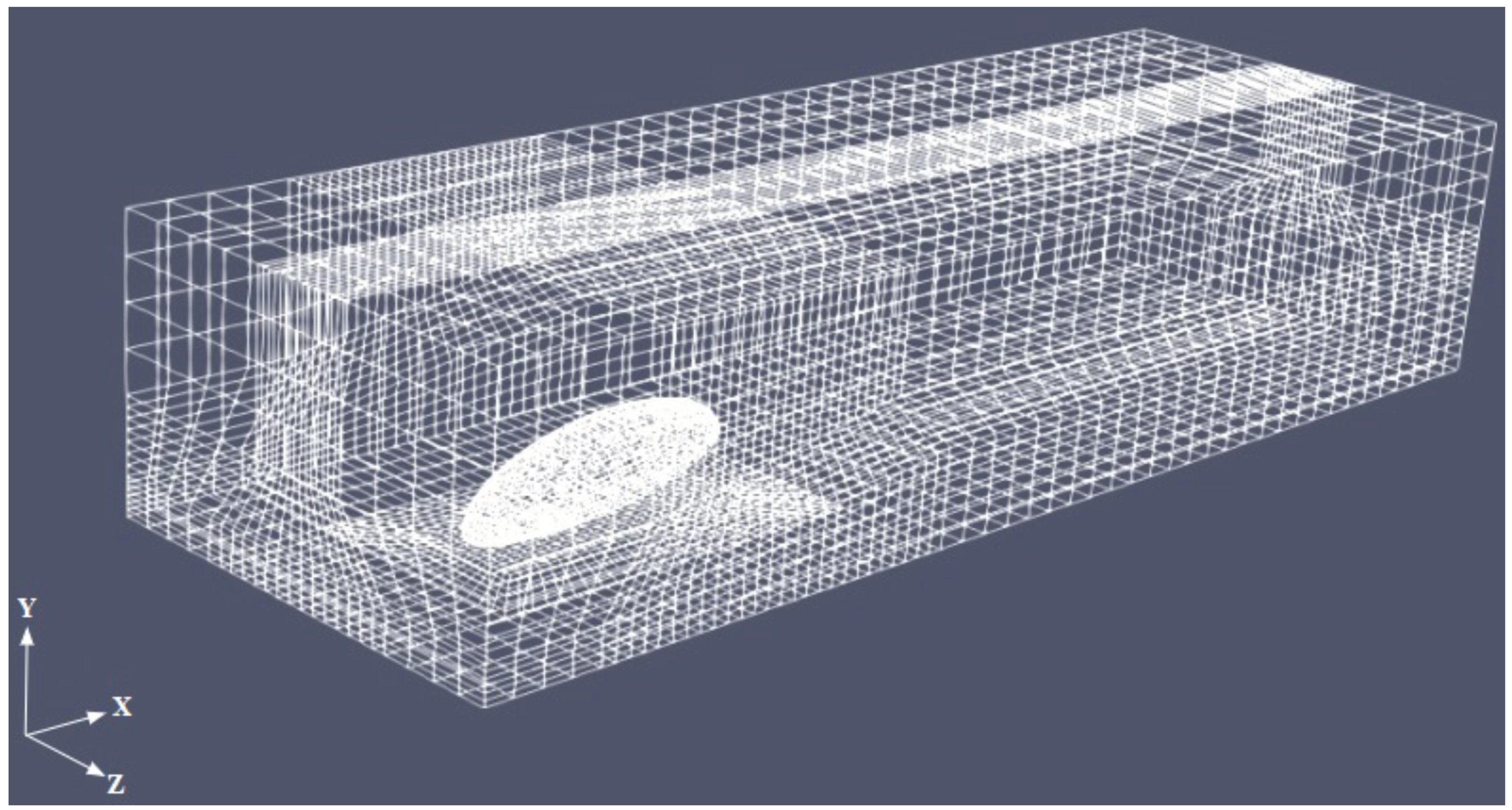
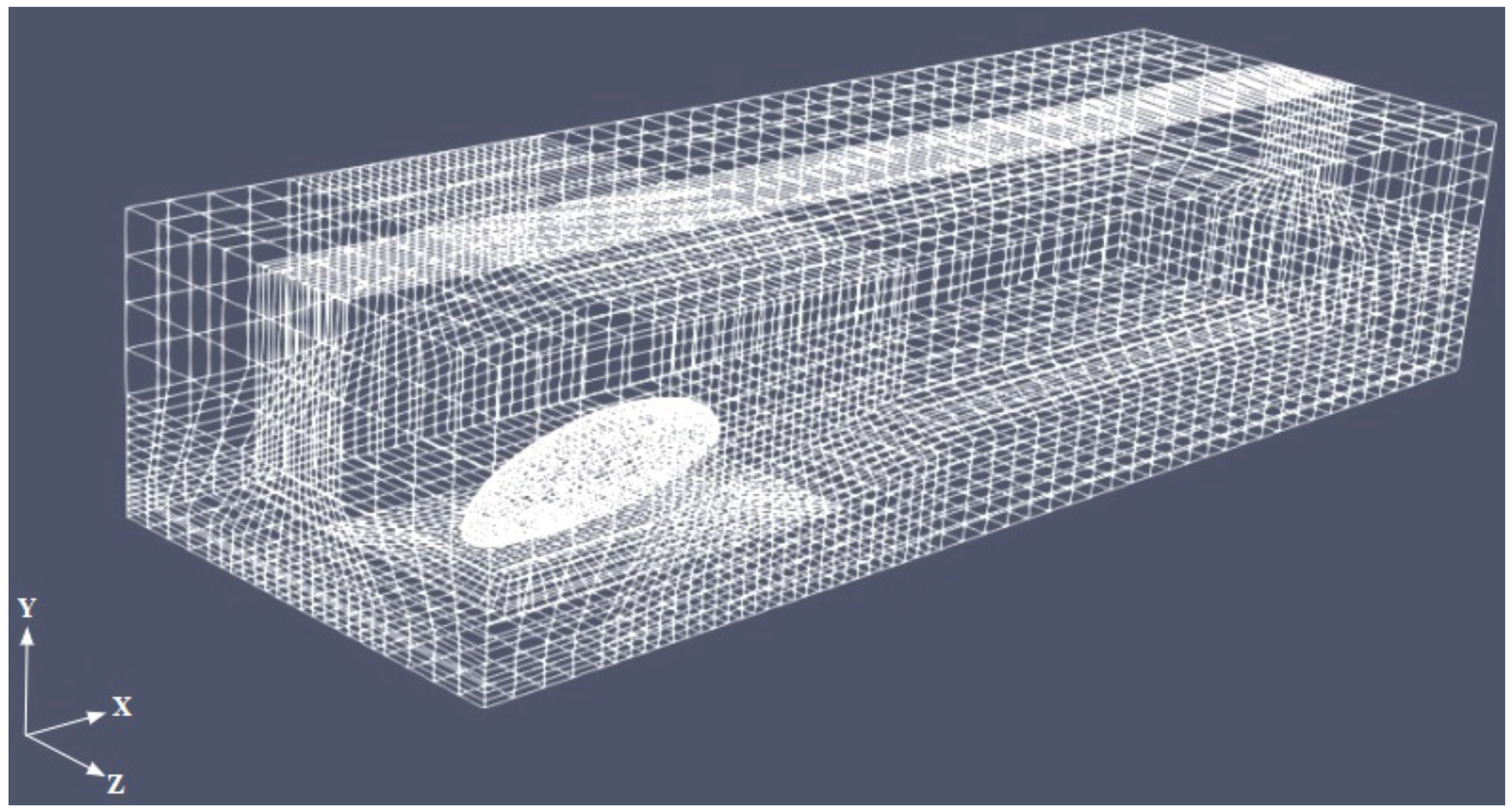
Figure 3 shows the mesh for
. The body is not exactly laying on the ground, but is elevated to mimic the beetle posture due to the presence of the legs (not modeled here). The boundary conditions were assigned in the form of inlet, outlet, no slip at the base wall of the box and the beetle surface, and symmetry at the lateral and top face of the box, as sketched in
Figure 2.
Table 1 summarizes the details of the simulations conducted in this study. In the baseline cases (runs A1–A10) the free-stream air velocity was
and the droplet volume fraction injected in the domain was
. We also considered larger velocities and smaller volume fractions (runs B1–B9 to F1–F10 in
Table 1), so as to explore the sensitivity of the results to these parameters. These values are similar to those used in our laboratory experiments with live beetles, and, in the case of velocity, they represent light to moderate breezes during foggy days in the Namib desert coastal region according to available meteorological data [
11,
12]. Finally, in a last set of simulations (G1–G10), we considered the influence of non-uniform droplet size distribution.
For each body inclination angle
, the numerical simulations were carried out in two steps. First, air was introduced at the inlet of the computational domain with uniform velocity
and Equations (
1) and (
2) were solved with constant time step
until
when the solution converged, i.e., the flow properties did not change any longer throughout the computational domain. The flow Reynolds numbers for
and
are
and 2665, respectively. The first two values are low enough for the flow to be considered laminar throughout the entire domain. For the highest Re, the flow may experience incipient transition to turbulence, even though the impact of turbulence on the droplet collection on the body is likely minor—we do not expect early detachment of the boundary layer induced by low-intensity turbulence, the main effects, if any, being on the wake of the body, which is of less importance in this study.
Figure 4 shows the velocity magnitude contour at
for the representative case of zero incidence angle (run A1), and demonstrates the development of the boundary layers around the body’s surface and over the ground base. Note the confluence of the fluid as it streams inside the gap between the ground and the bottom side of the body.
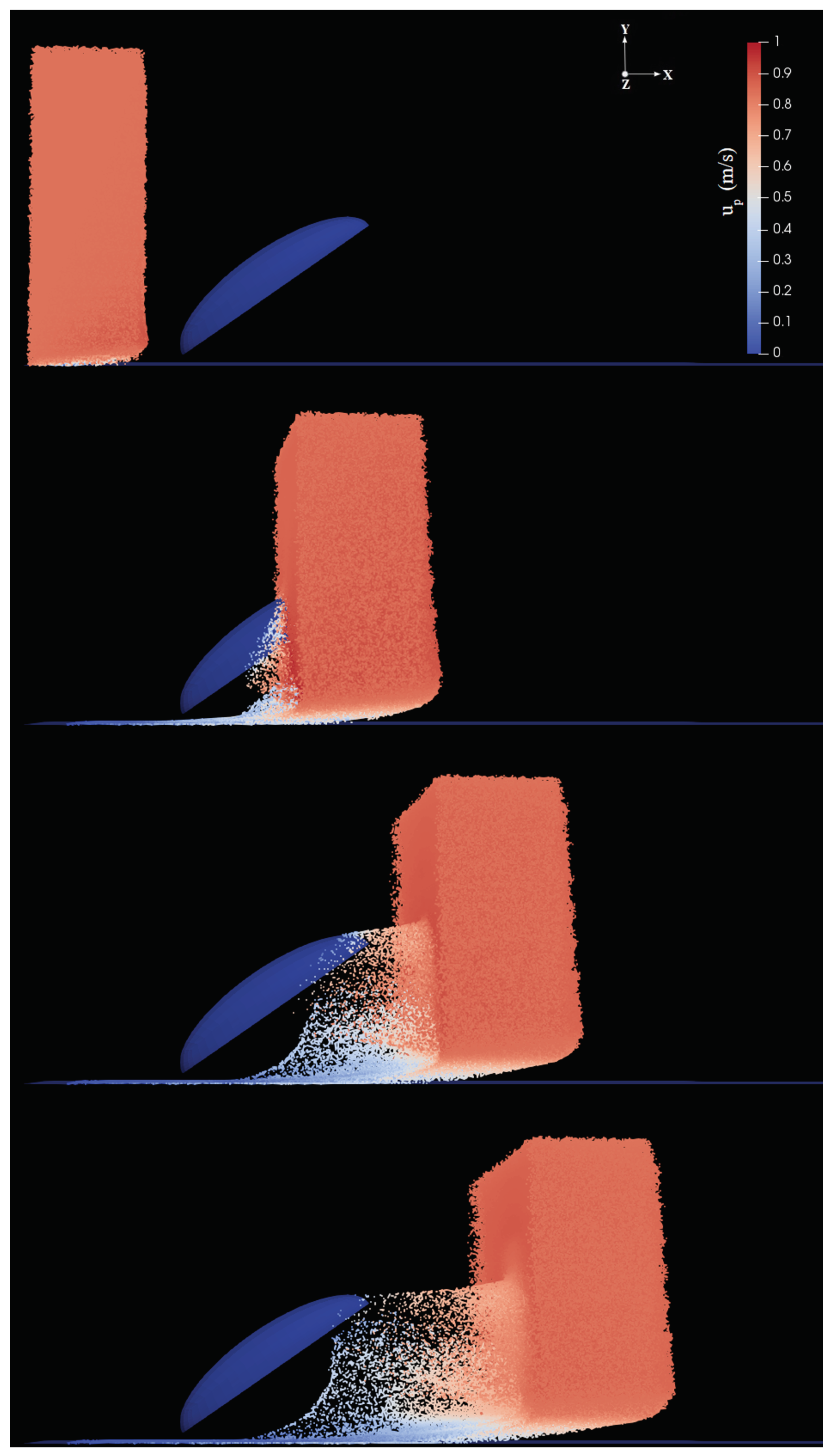
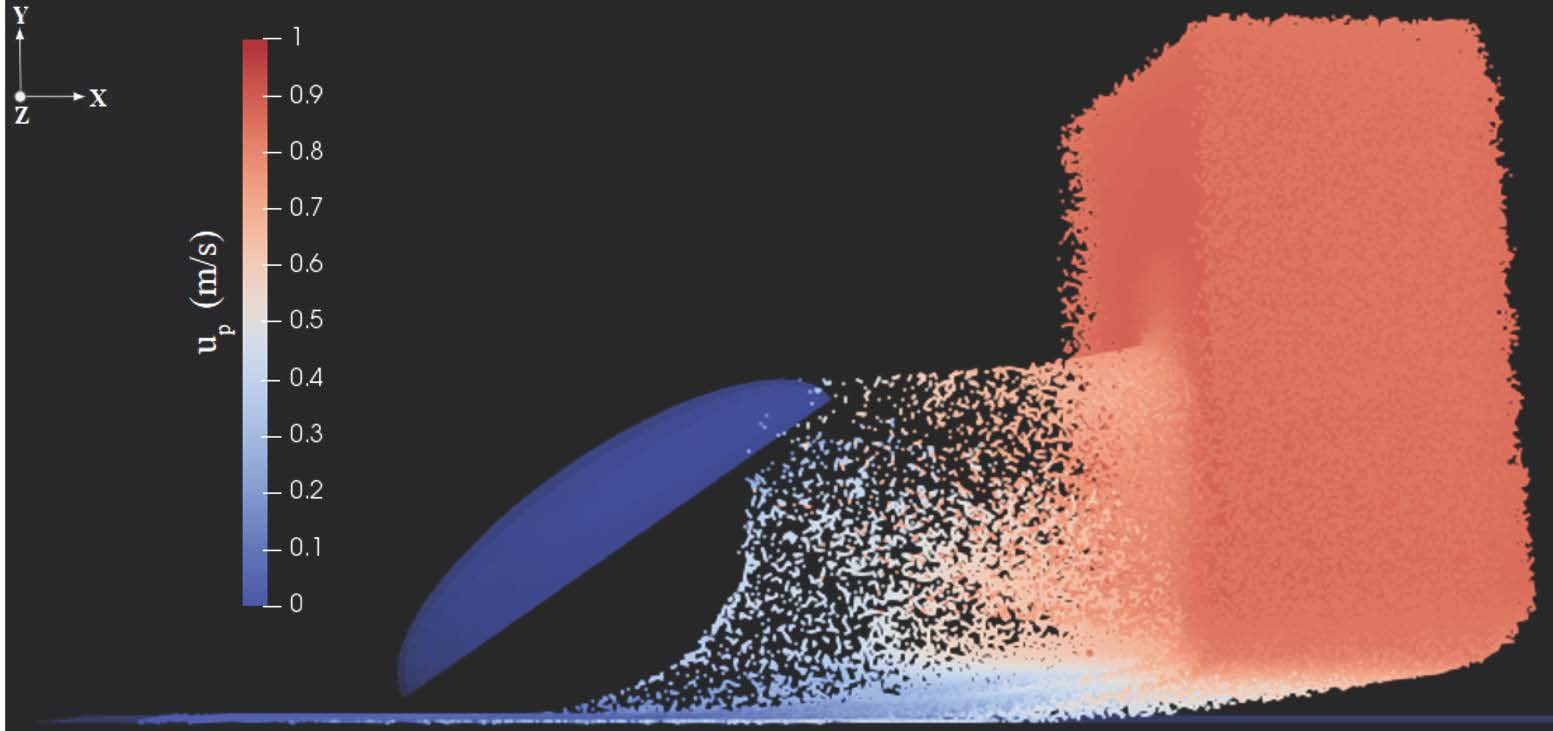
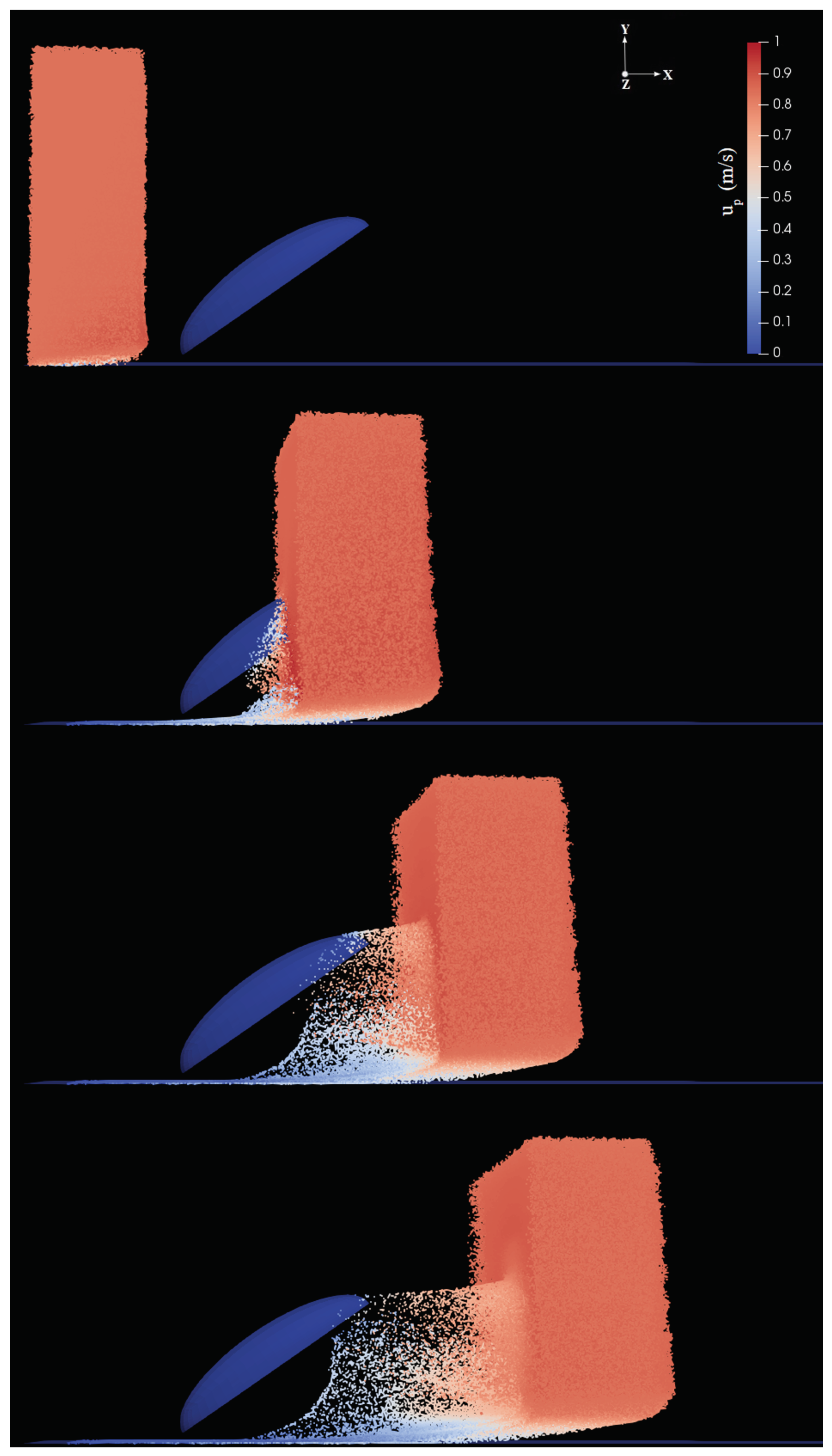
In the second step, Lagrangian fog droplets were injected into the environment with the same free-stream velocity of the carrier air stream (i.e., zero relative velocity with respect to the fluid). In all simulations we used the patchInjection module of OpenFOAM, where
droplets were injected at the inlet. This results in a slab of droplets reaching the body, as shown for example in
Figure 5. The volume of the slab is
where
is the streamwise extension of the slab. Droplets were initially homogeneously distributed inside this slab. In the uniform size distribution cases, droplets were assumed to have diameter
, typical of desert fog [
1], the total volume and mass of water injected in the domain were thus
and
, respectively. The droplet volume fraction in the slab was
(values listed in
Table 1). For the sake of verification of the two-phase flow assumptions, we compared one-way and two-way coupling simulations for the baseline cases; the outcomes were not influenced by the coupling assumption in the most critical case, i.e., larger volume fraction (see
Appendix A). The droplet dynamics can be described qualitatively as follows: droplets are partly intercepted by the body, and partly either continue their trajectory with essentially the same velocity (those above the body) or penetrate the boundary layer developing on the ground (below the body). Among droplets intercepted by the body, some are captured inside the boundary layer and eventually stick to the surface, while some are able to escape (never contacting the body) and continue their trajectory in the wake with reduced velocity (see for example
Figure 5d). In order to determine the droplet collection efficiency, we calculated the ratio of the mass of droplets coming in contact with the surface of the body and the mass of particles injected into the environment, for each inclination
. The data related to this mass is obtained using the surface film modeling of OpenFOAM, Equation (
6).
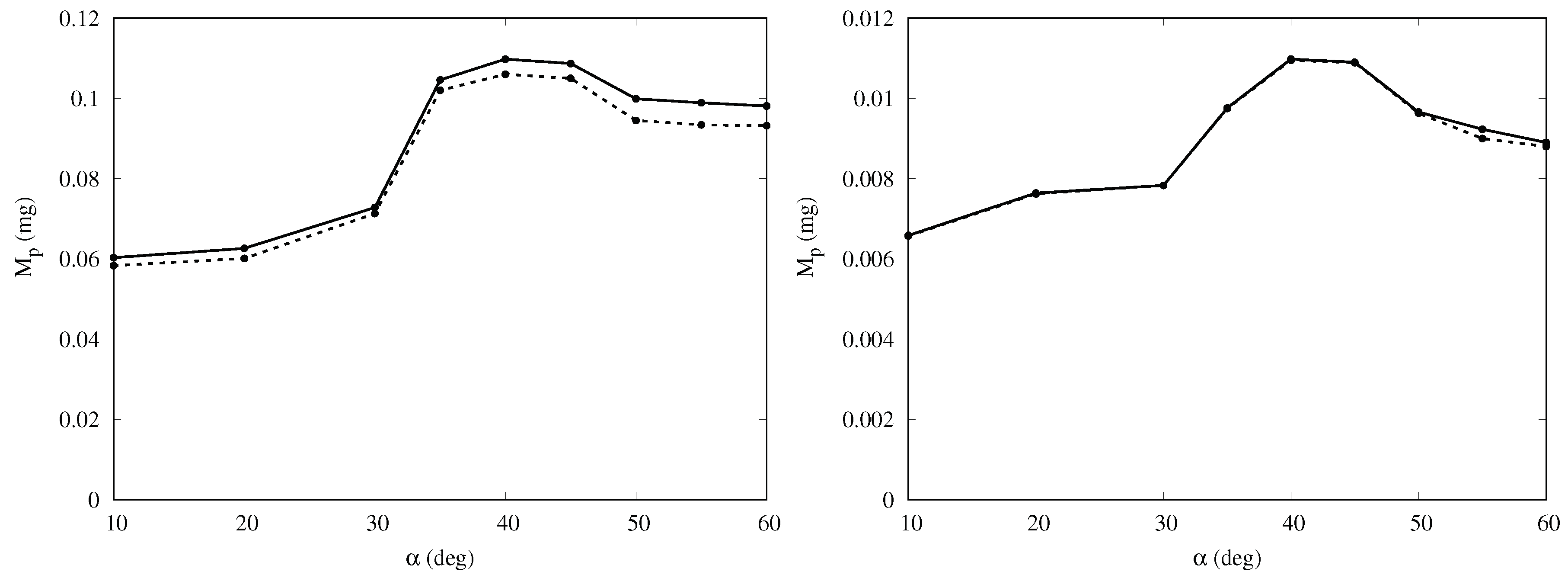
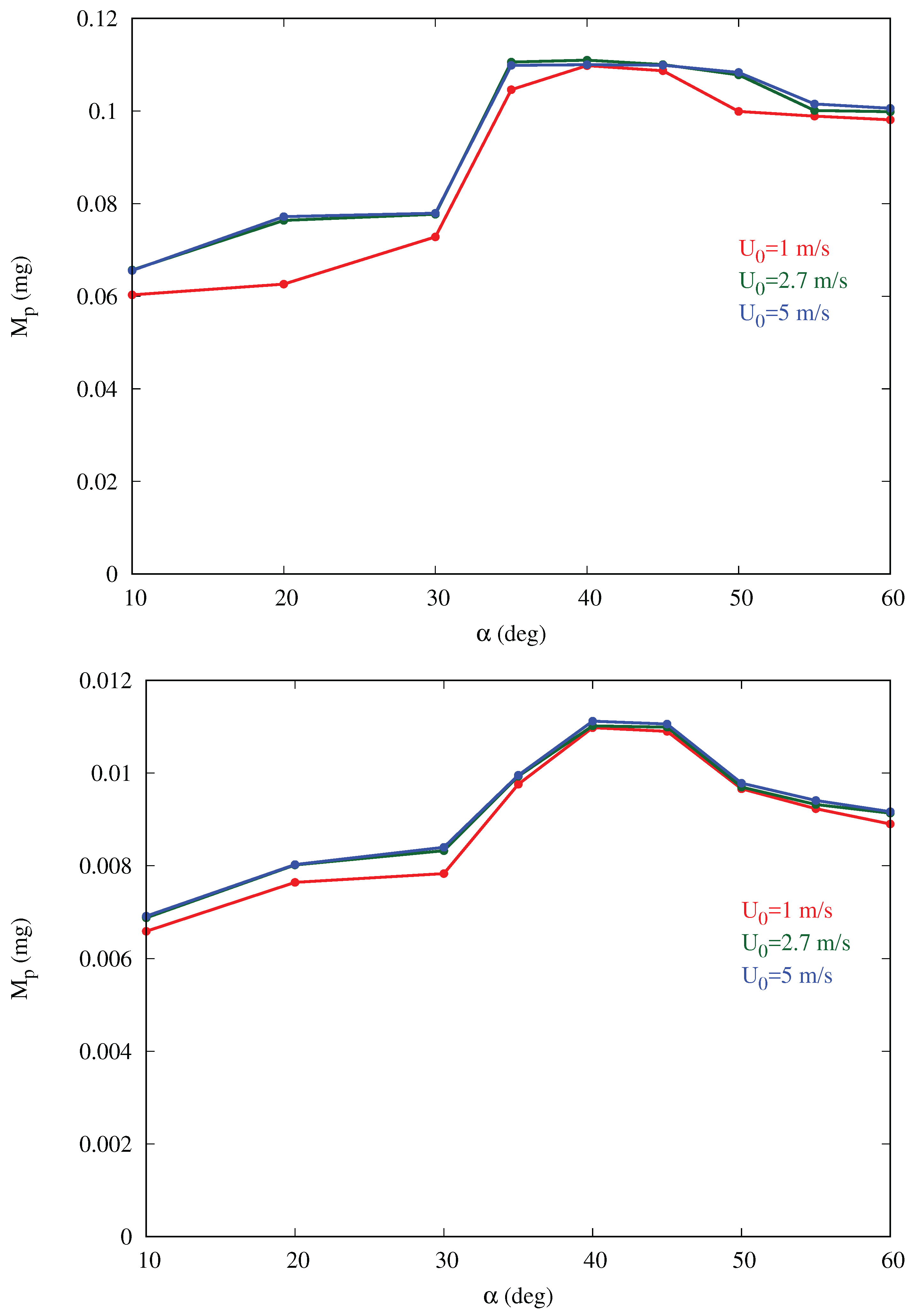
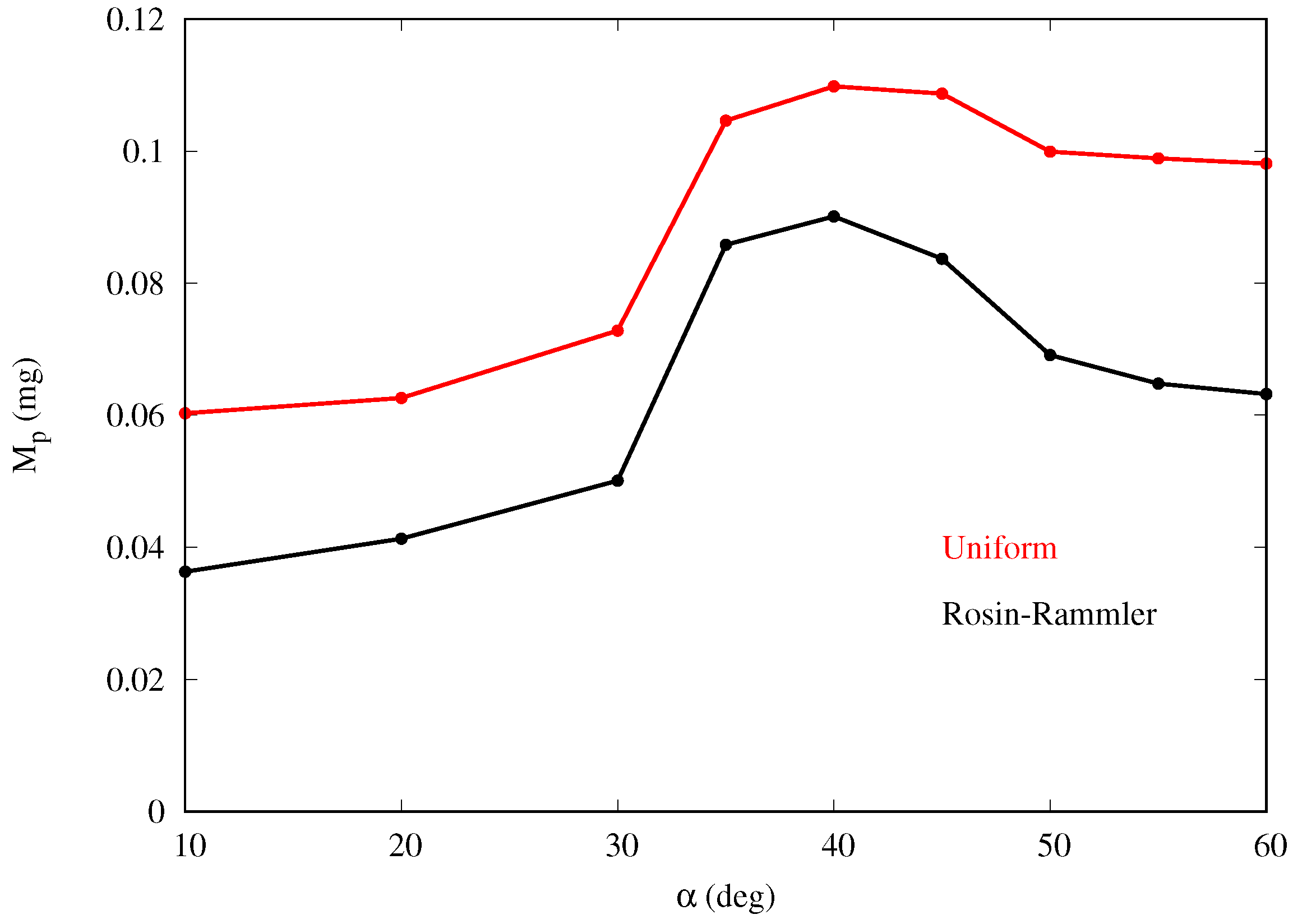
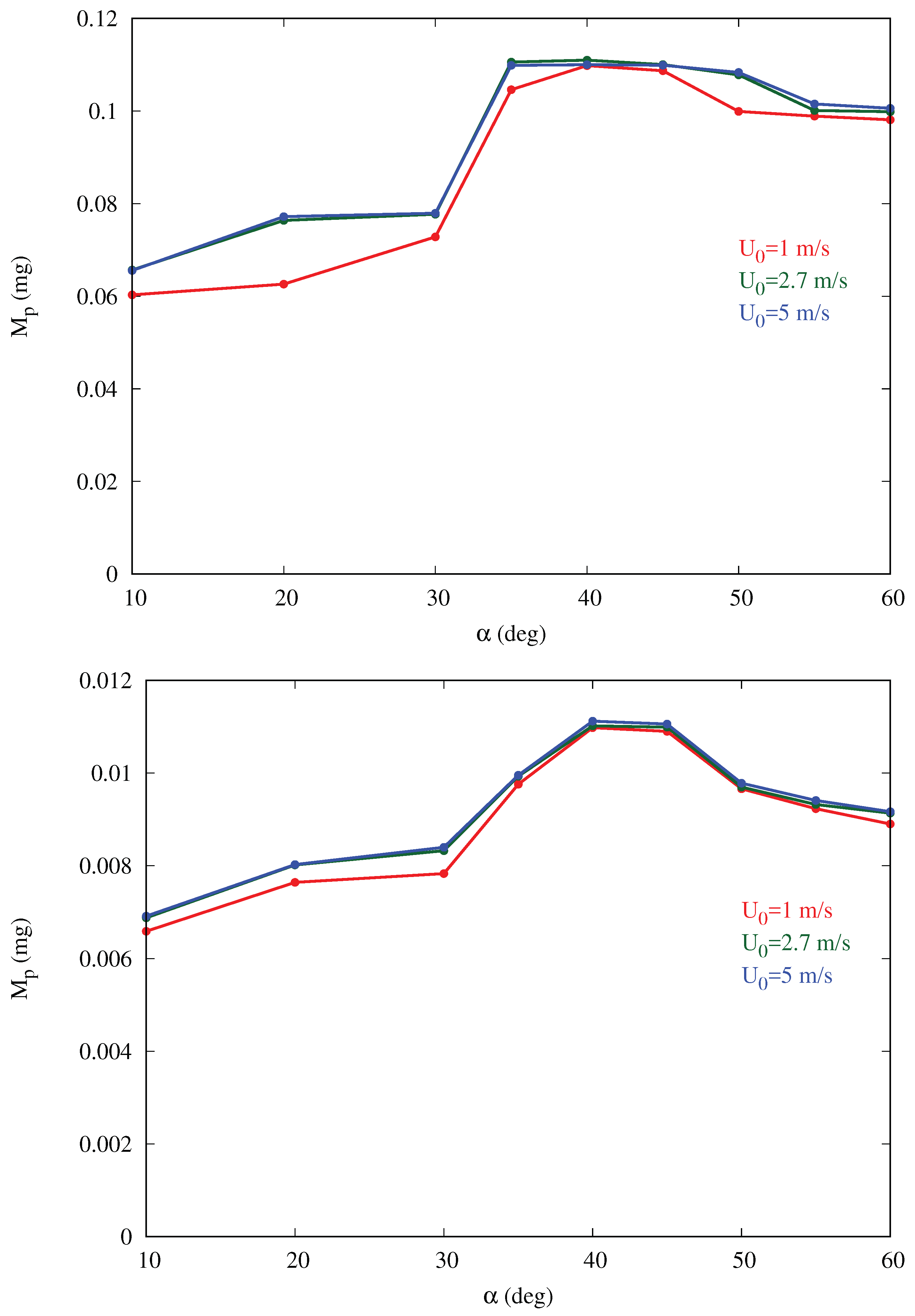
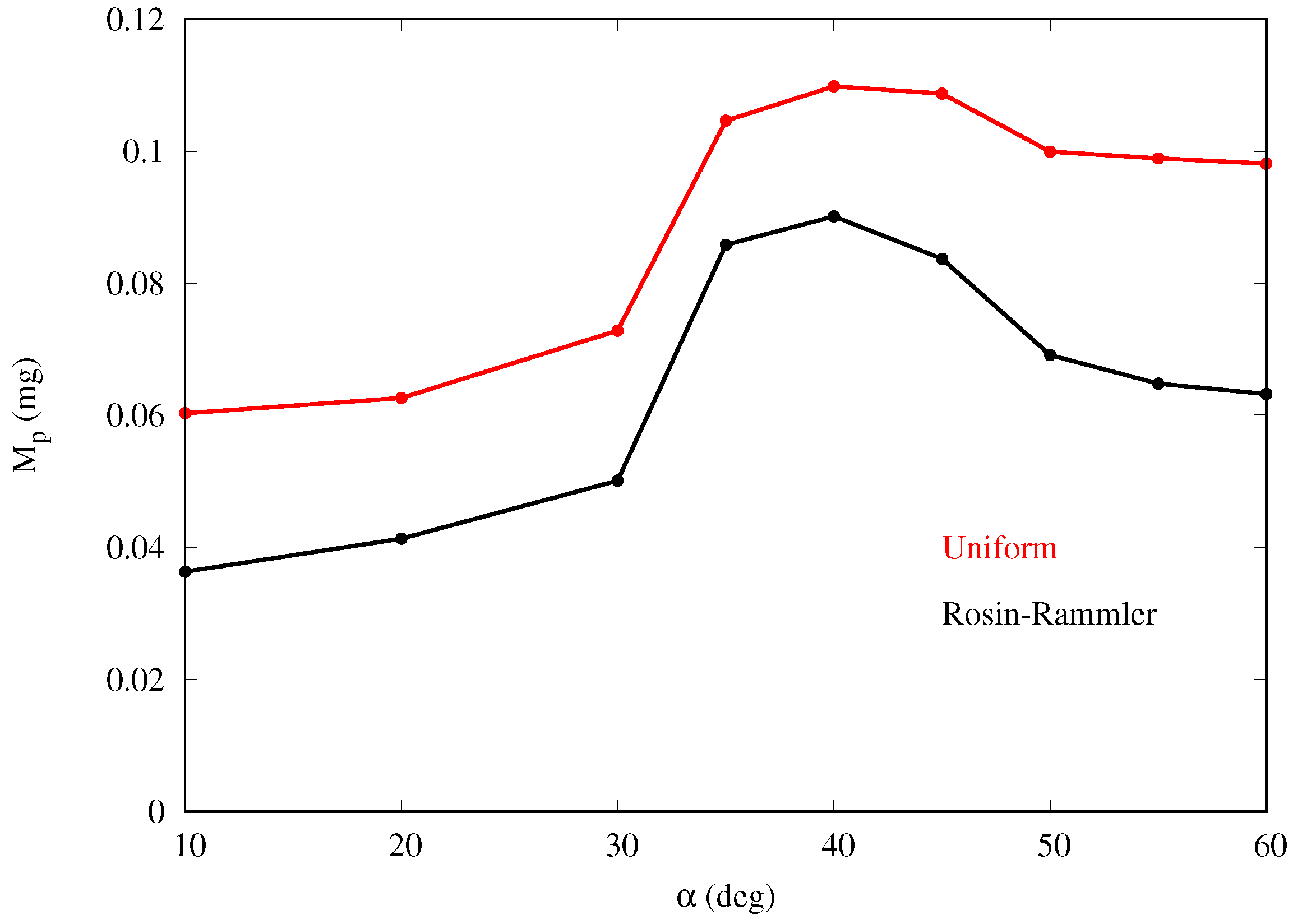
Figure 6 shows that the mass collection is maximum at
, being about 65% more than at
, and 16% more than at
for the baseline cases. This behavior is observed for all cases in
Table 1 with the peak occurring at inclination between 35 and
. For a given inclination angle
, the mass collection tends to increase slightly with the free-stream velocity
(converging for larger values), especially in the cases with larger volume fraction
. This is likely due to the fact that, if velocity increases, more droplets impact the body since they are less likely to circumvent the obstacle due to their higher momentum. In addition, a smaller fraction of droplets is trapped in the thinner boundary layer developing on the ground, without ever reaching the body.
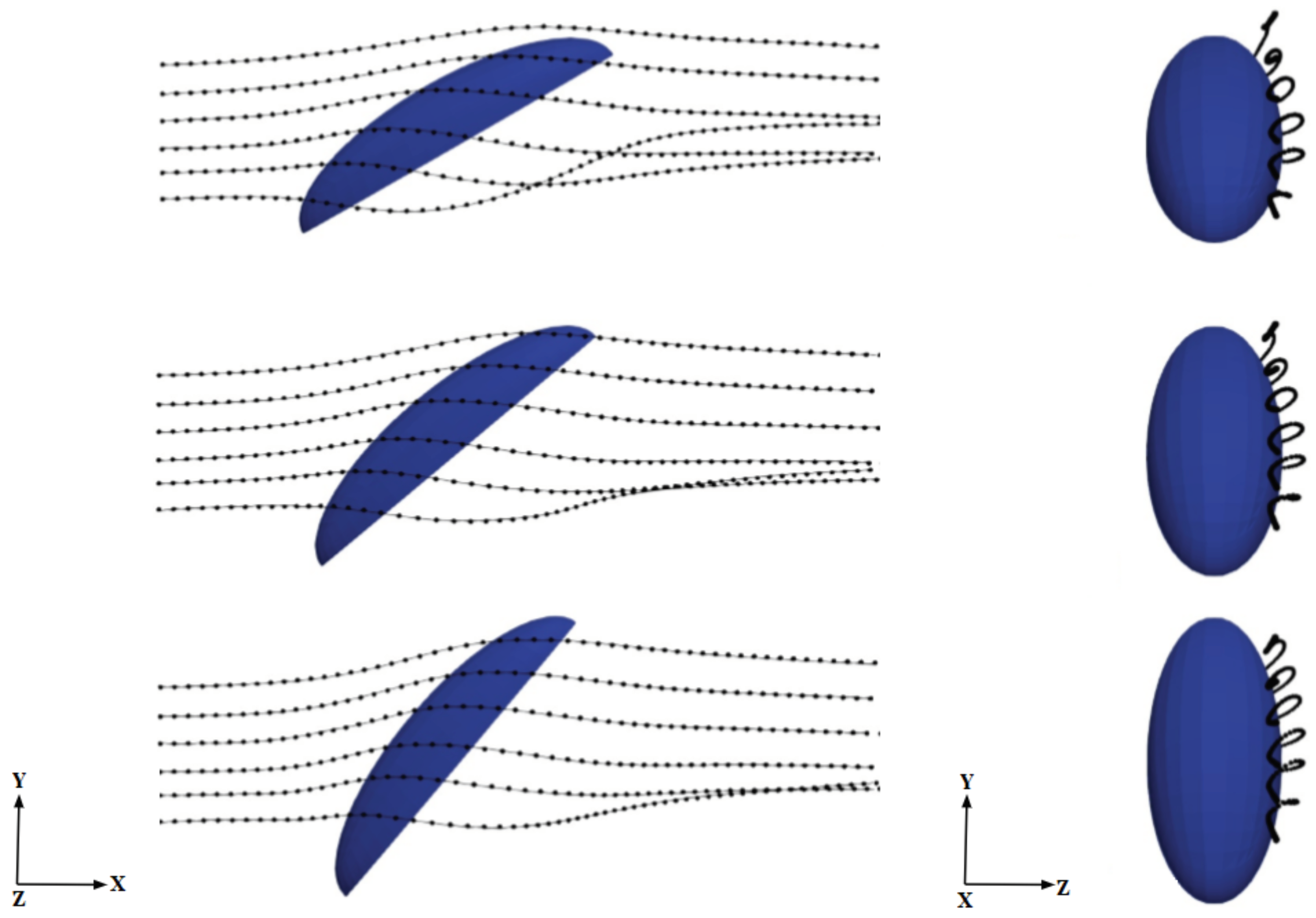
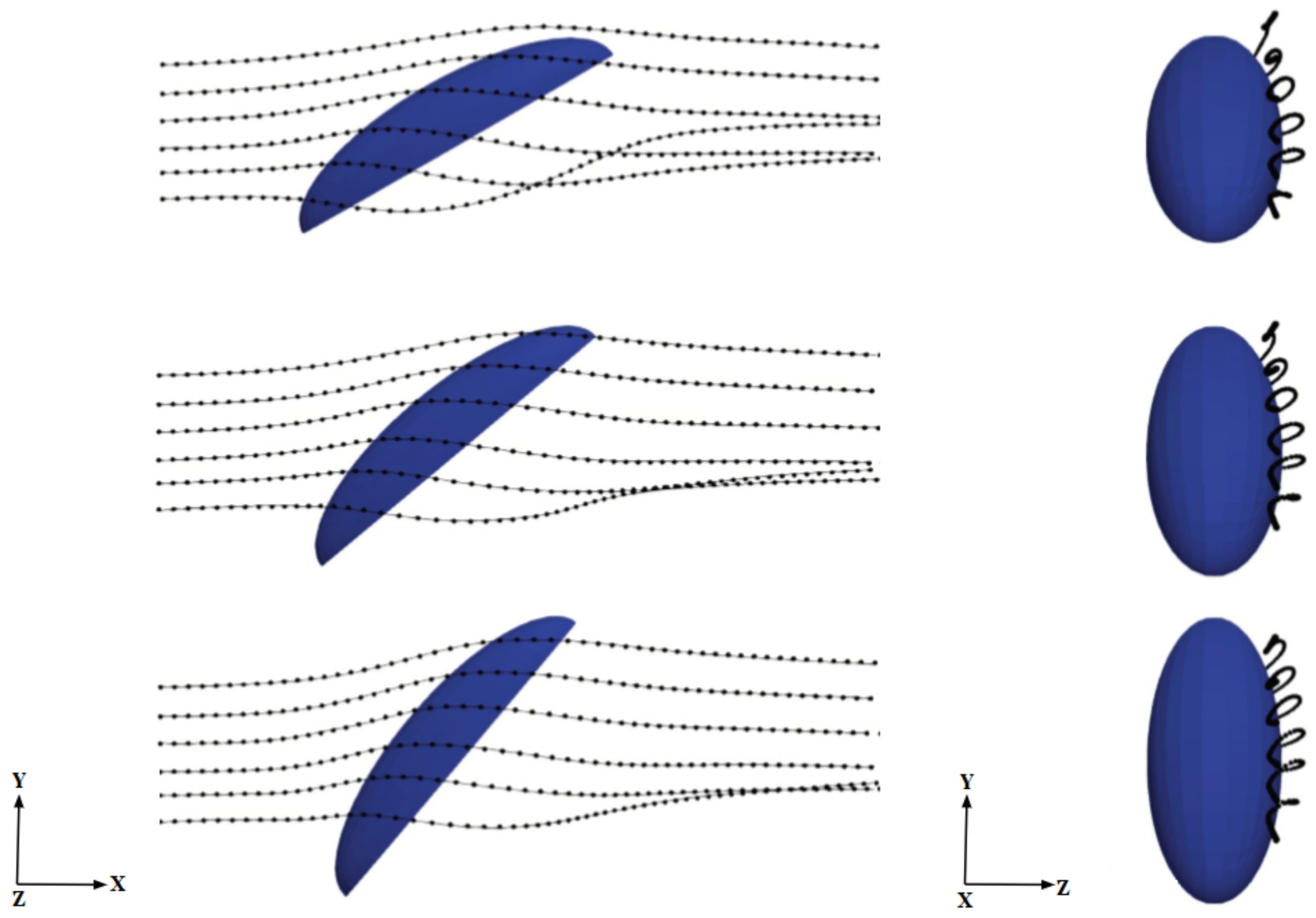
One possible explanation for this intriguing behavior for the mass collection efficiency exhibiting a consistent peak around 40 deg is related to the three-dimensional aerodynamics of the flow around the semi-ellipsoid, which deflects the initially straight droplet trajectories. This, in turn, influences the velocity of droplets entering the boundary layer and depositing on the surface in a complex way that cannot be predicted solely on the basis of geometric arguments. In order to confirm this hypothesis, we analyzed droplet trajectories obtained by connecting the positions of selected droplets at various instants
as they move through the flow field (which also changes in time because of the two-way coupling). We inject particle tracers along lines located at different heights from the ground, each initially separated vertically by 0.2 mm from its nearest neighbor and all located at a distance of 0.87 mm from the central vertical symmetry plane (see
Figure 7). The average arc length
traced by the particles near the surface of the body and the average residence time
are calculated, respectively, as
In the above equations,
represents the differential arc-length along a given droplet trajectory with
and
the initial and final positions (ahead and behind the body, respectively);
is the corresponding velocity magnitude at each location along the arc, whereas brackets denote average over the trajectories.
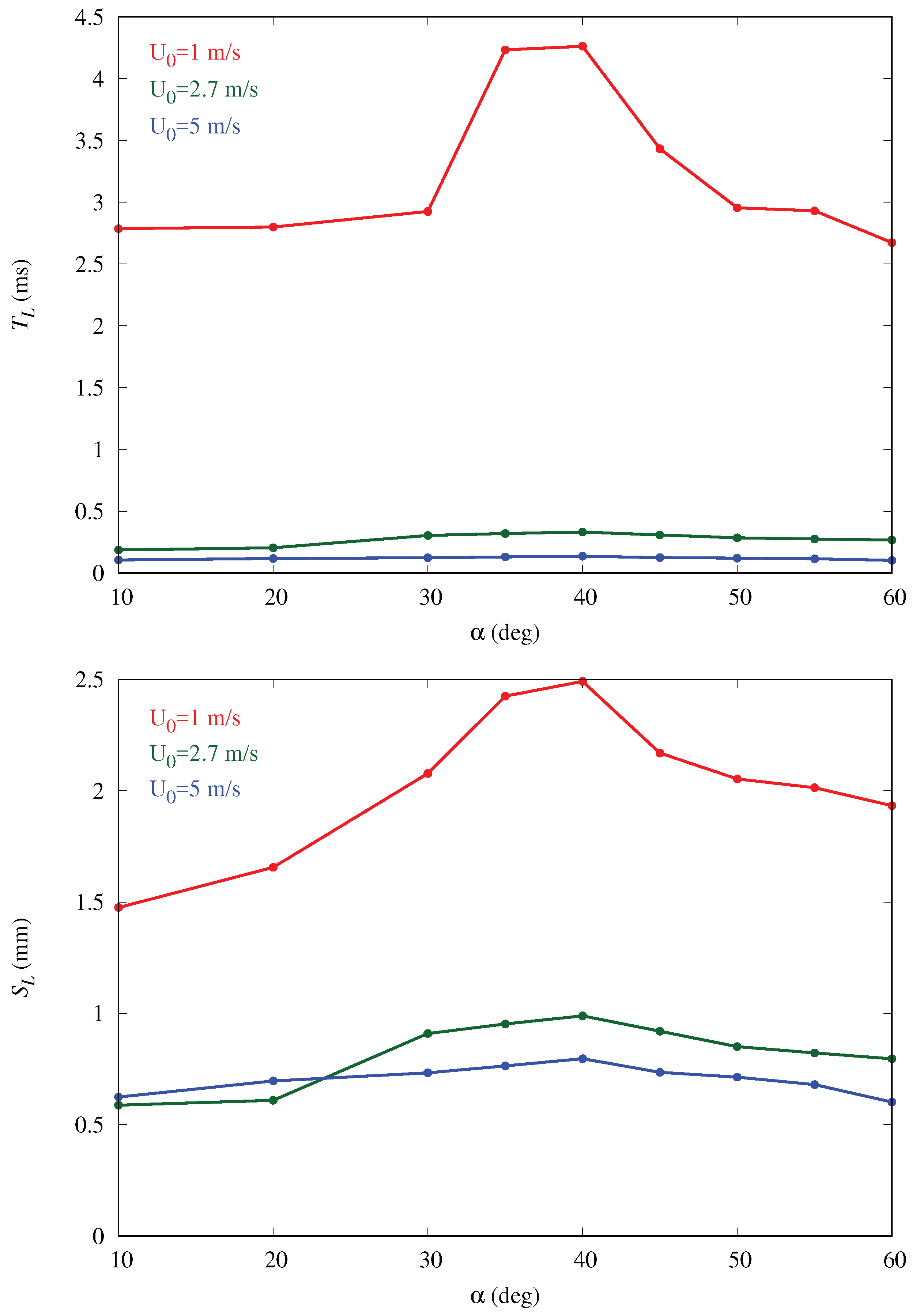
Figure 7 shows seven trajectories at three different body inclination angles; the results are insensitive to this choice, given the smooth character of this laminar flow. Both
and
are plotted for varying inclinations in
Figure 8. We observe that both parameters follow trends similar to those observed for mass collection efficiency with shallower variation with
for higher free-stream velocities. The rationale proposed here for this behavior is that when an approaching droplet spends more time near the surface, the probability for droplet collection on the surface rises. This is an aerodynamically-driven (rather than purely geometrically-driven) process and can only be captured by solving the full Navier–Stokes equations. In particular, the curvature of the body is crucial to explain the residence time trends. This can be appreciated from the trajectories in
Figure 7 and from the three-dimensional distribution of the deposited liquid film thickness determined by Equation (
6) and plotted in
Figure 9 for
. The film thickness is highest around the bottom and in the middle of the body’s surface and decreases at the top as well as on the sides of the semi-ellipsoid. Increasing
would certainly increase the number of particles intercepting the body locally but would also decrease the residence time of particles around the body, thus giving droplets less time to circumvent the body and get captured in the curved boundary layer. Hence, there has to be an optimum inclination (around 40 deg in this simplified geometry) for which the two effects combine to maximize the collection efficiency. Note the difference with a purely two-dimensional geometry like a flat plate at inclination, where all trajectories would eventually impact the plate (as long as
) and so the maximum collection would occur for
(frontal impact), based on purely geometrical reasoning.
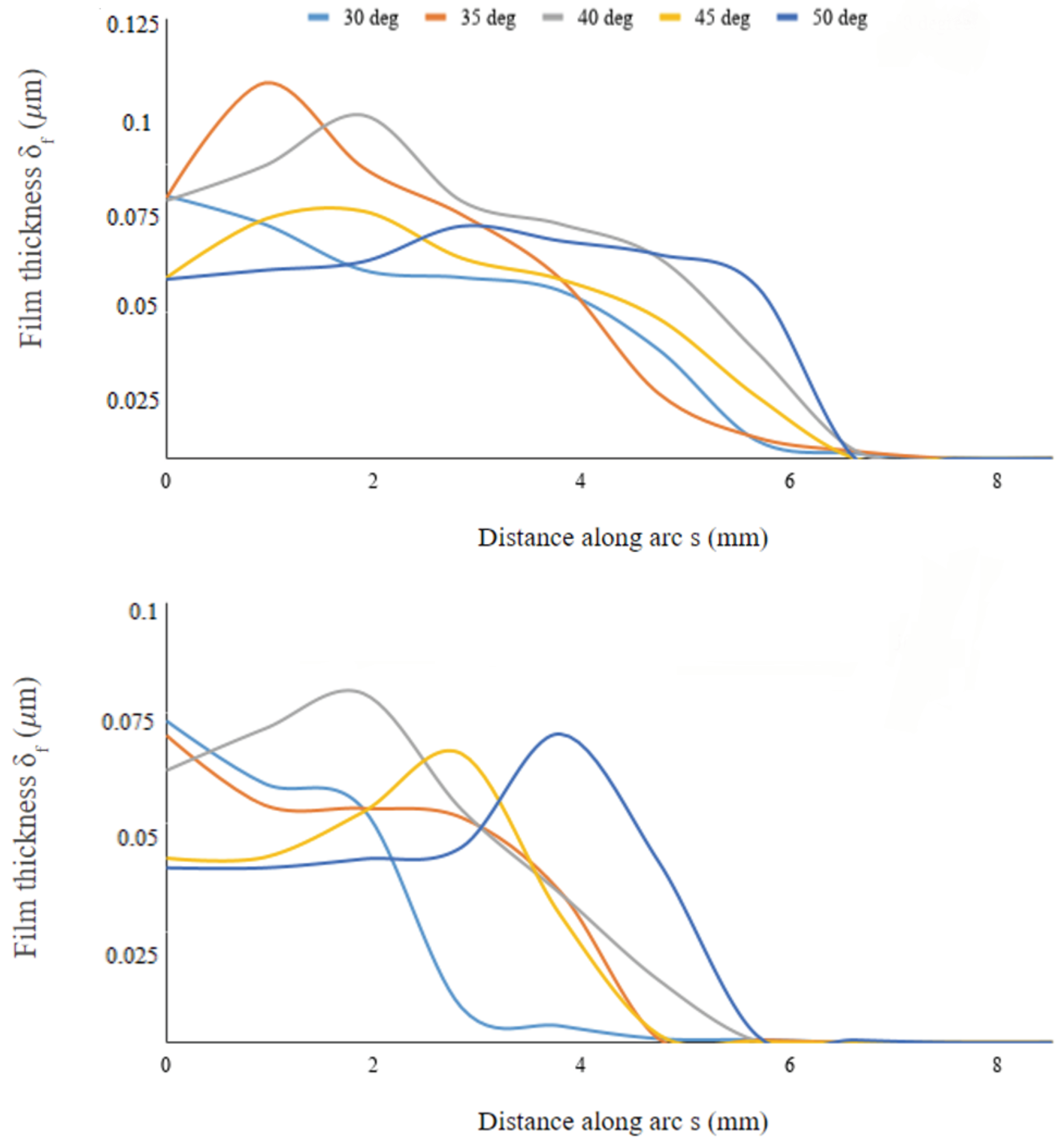
3.1. Water Collection along the Surface
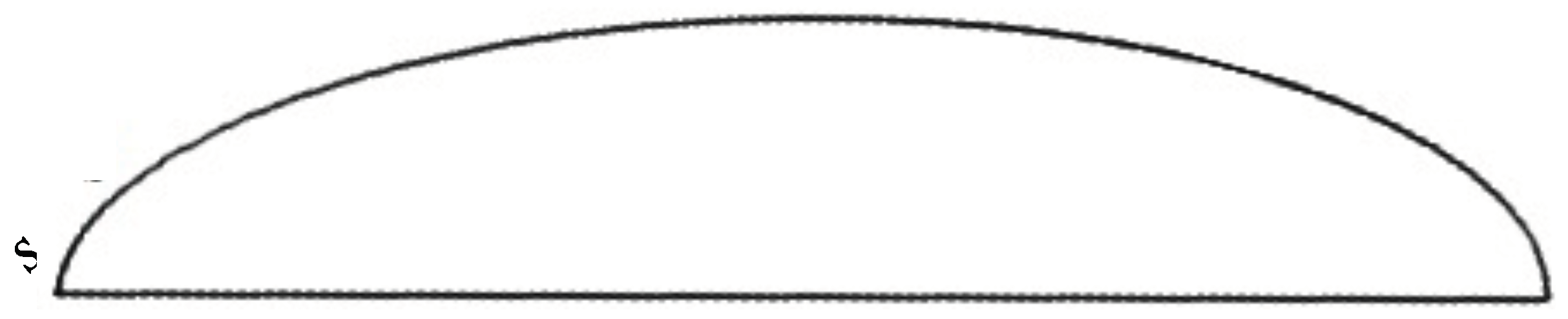
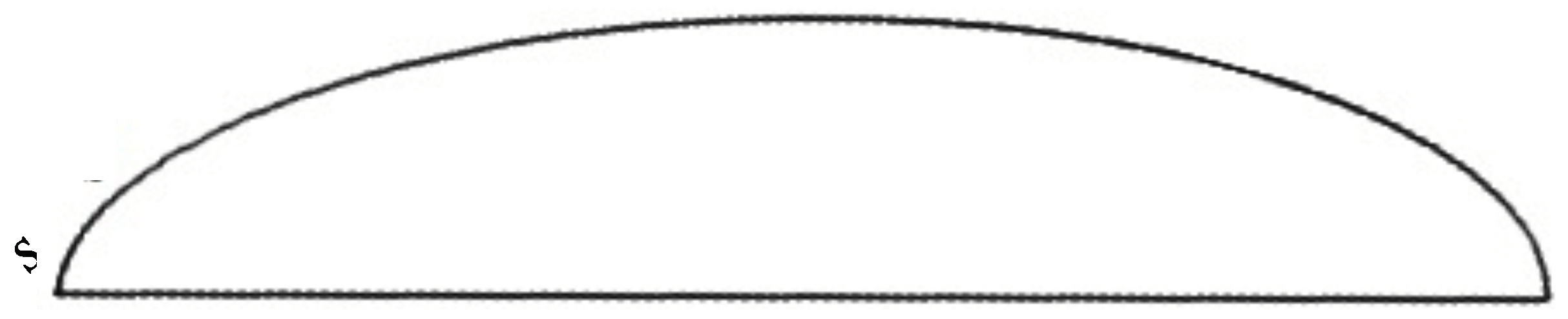
To further investigate the water collection along the body surface, we determined the film thickness
of water collected along an arc defined by the intersection of the vertical symmetry plane and the semi-ellipsoid. We did the same for another arc defined by the intersection of the body and an inclined plane passing through the lengthwise symmetry line at the bottom of the body and turned 45 deg from the symmetry plane. This procedure defines the sections along which the film thickness is measured. The data are then plotted as a function of arc length (see
Figure 10) for varying inclinations of the body.
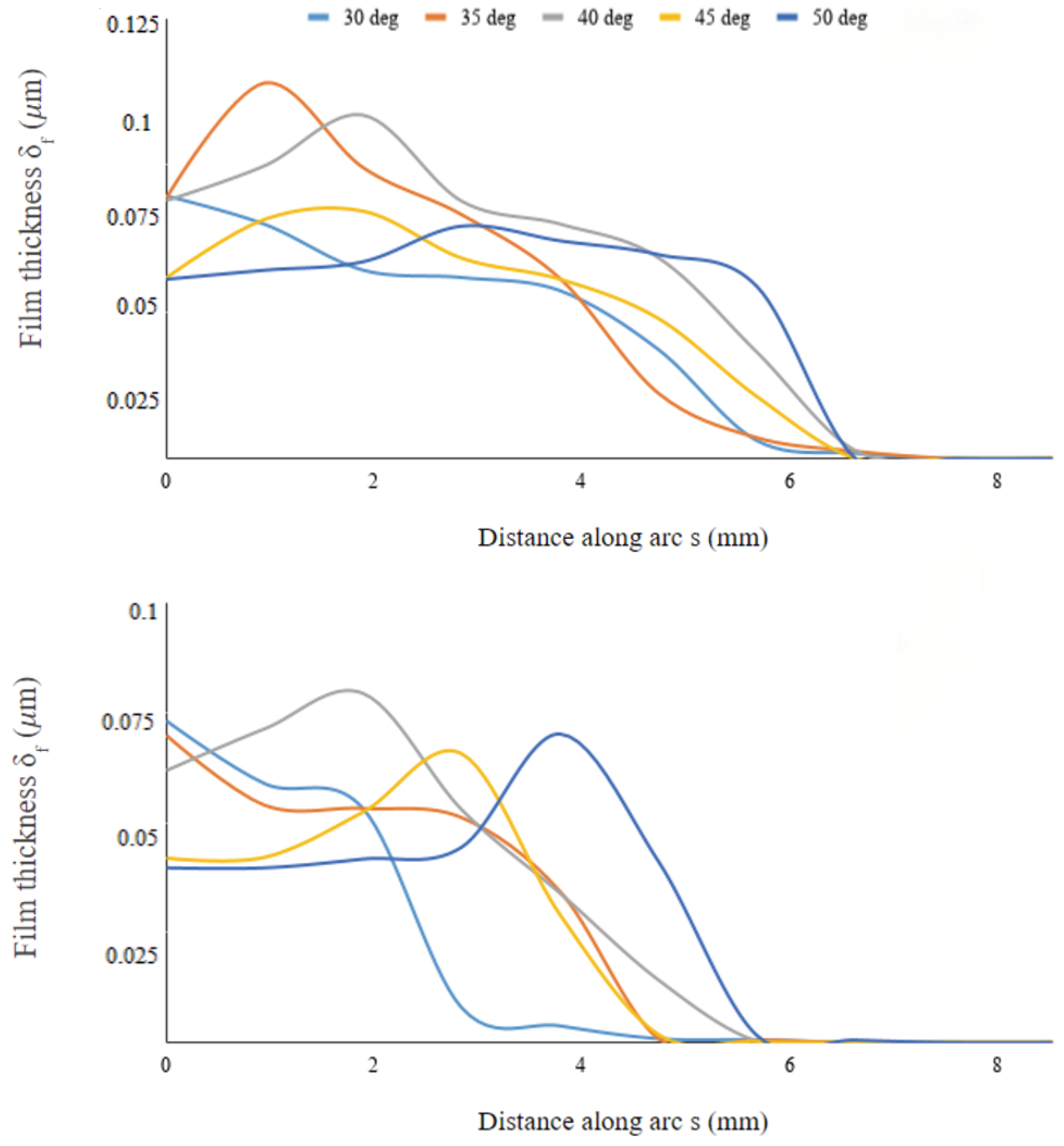
Figure 11 shows the profiles to be wider on the symmetry plane compared to the eccentric plane, thus indicating that the film spreads more effectively lengthwise than laterally along the body’s surface; this is also noticeable from the snapshots of
Figure 9. The film thickness peaks at different stations along the surface arc-length, depending on the inclinations angle
. On the symmetry plane, the arc-length location where
peaks is initially located at the leading edge of the body for
, then it moves downstream as inclination increases to
, moving back upstream for
, and downstream again for
where the distribution of
is more uniform throughout the entire arc-length. Interestingly, this non-monotonic behavior occurs around
where the collected water mass is also maximum (see
Figure 6). The same behavior is observed along the eccentric arc, although the displacement of the peak location is delayed compared to the displacement on the symmetry axis, with film peaking narrowly at the rear of the body when
. Hence, the geometric alignment of the beetle with respect to the incoming fog stream contributes to the high mass collection efficiency at these inclinations.
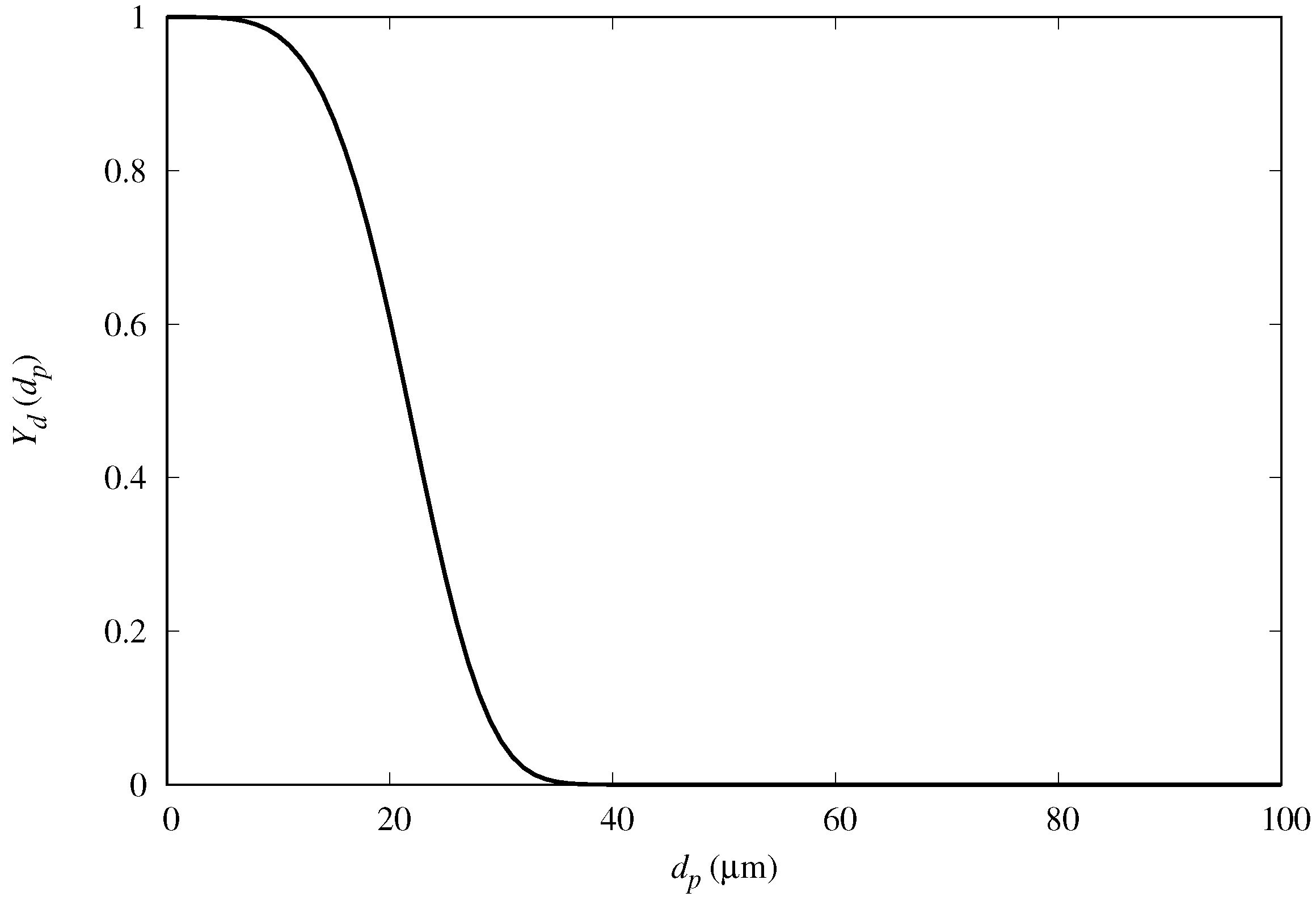
3.2. Results with Rosin–Rammler Droplet Distribution
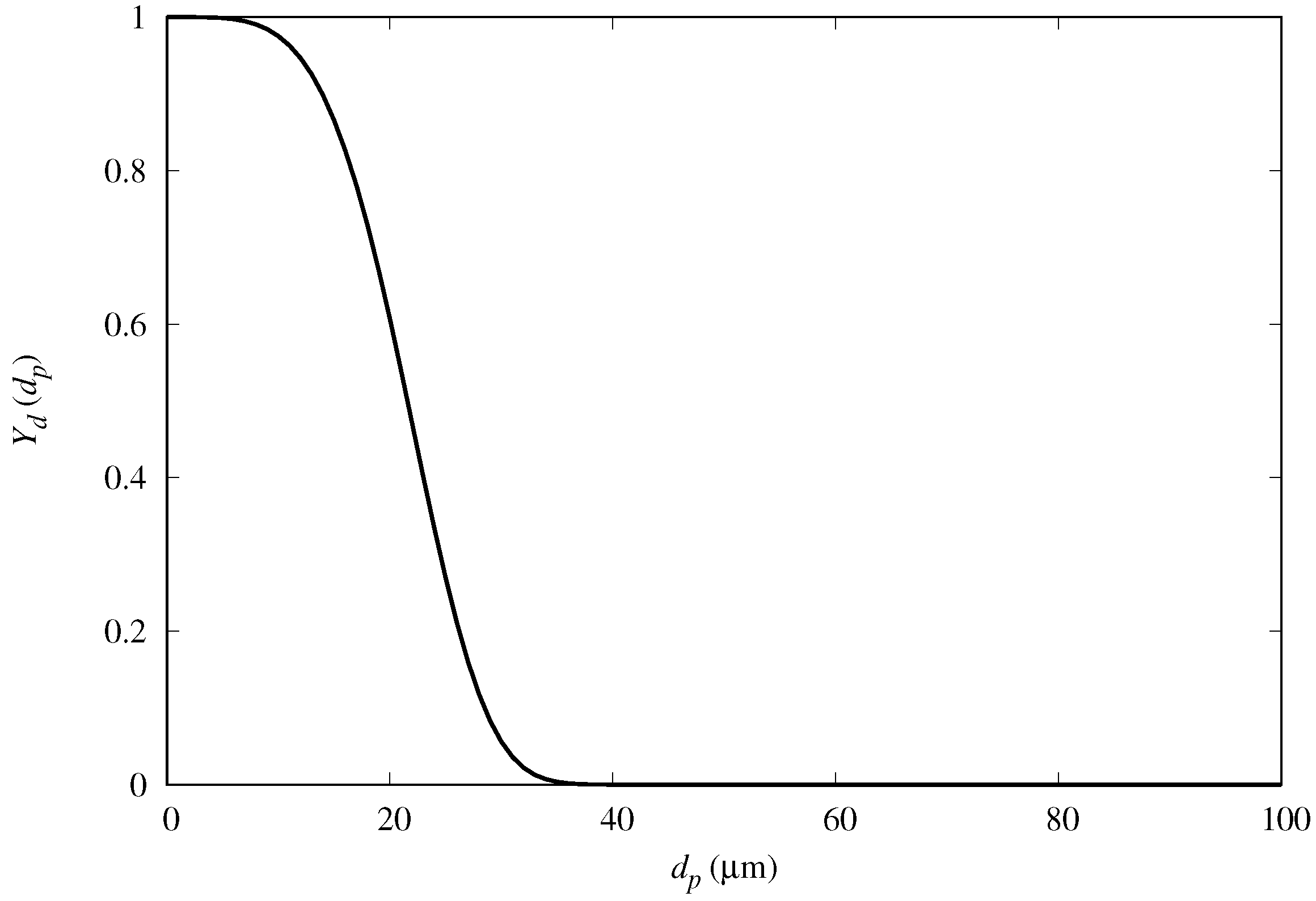
In order to further examine the fog mass collection efficiency of the semi-ellipsoids, we compared the results for predicted mass-collection efficiency for fog with polydisperse droplet size distribution. We cast the diameter of the droplets into a Rosin–Rammler distribution, and repeat the numerical simulations with a polydispersed input flow condition in OpenFOAM (see
Figure 12). Following this distribution, the fraction
of droplets greater than
(complimentary cumulative distribution) is expressed as
where
is the mean diameter and
n is the size distribution parameter. The values considered are
and
, based on fog tests performed in our laboratory with a household type humidifier.
Figure 13 shows that the mass collection efficiency follows a similar trend as for the monodispersed fog cases, with maximum collection also at around
.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}