

Three-Dimensional UAV Path Planning Based on Multi-Strategy Integrated Artificial Protozoa Optimizer


Abstract
1. Introduction
- A population initialization method using the tent map and ROBL is employed, which promotes a uniform distribution of the population, significantly enhances the diversity of the initial population, and thereby improves the algorithm’s exploration capability.
- During the autotrophic foraging phase, a dynamic optimal leadership mechanism is applied by introducing leaders and a nonlinear dynamic adjustment factor, which guides the search process toward the optimal solution, thus accelerating convergence while ensuring robust exploration.
- In the reproduction phase, a Cauchy mutation strategy is utilized to update positions; owing to the heavy-tailed nature of the Cauchy distribution, the algorithm is more likely to escape local optima during exploration, thereby increasing the probability of finding the global optimum.
- In the heterotrophic foraging and dormancy phases, a simulated annealing algorithm is incorporated, allowing the algorithm to accept inferior solutions during the early stages. This not only provides a probabilistic means to escape local optima and expand the search scope but also further enhances convergence efficiency.
- The performance of the IAPO algorithm is validated using the CEC2022 benchmark functions, with comparative experiments conducted against nine other commonly used algorithms. The experimental results confirm the superiority of the IAPO algorithm.
- The application of IAPO to UAV three-dimensional path planning problems, in comparison with other algorithms, demonstrates its adaptability and reliability in UAV 3D path planning scenarios.
2. APO Algorithm Overview
2.1. Traditional Artificial Protozoa Optimizer
2.1.1. Autotrophic Foraging
2.1.2. Heterotrophic Foraging
2.1.3. Dormancy
2.1.4. Reproduction
2.1.5. Algorithm Analysis
3. IAPO Algorithm
3.1. Algorithm Improvement Scheme
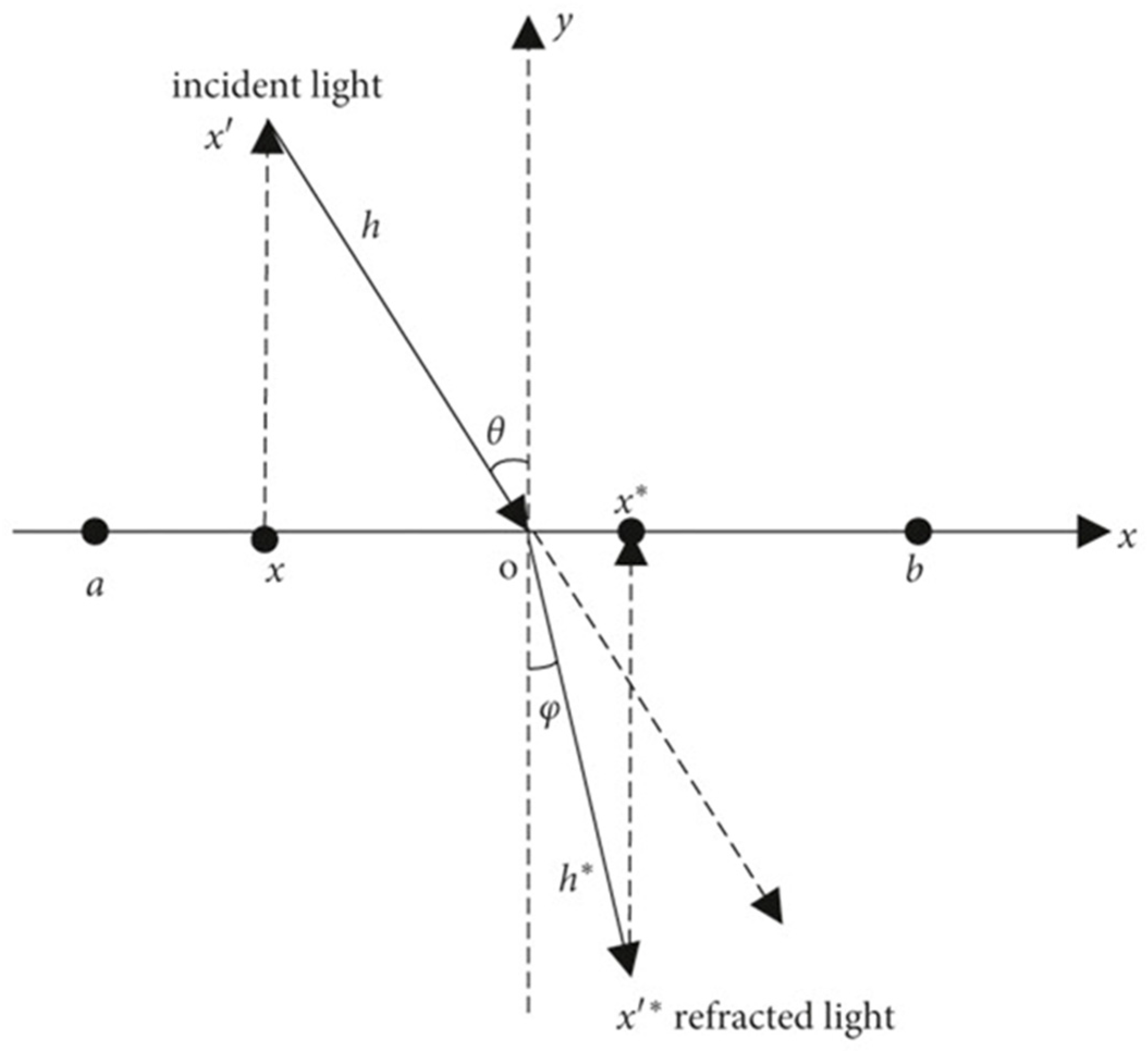
3.1.1. Population Initialization Based on Tent Map and Refractive Opposition-Based Learning
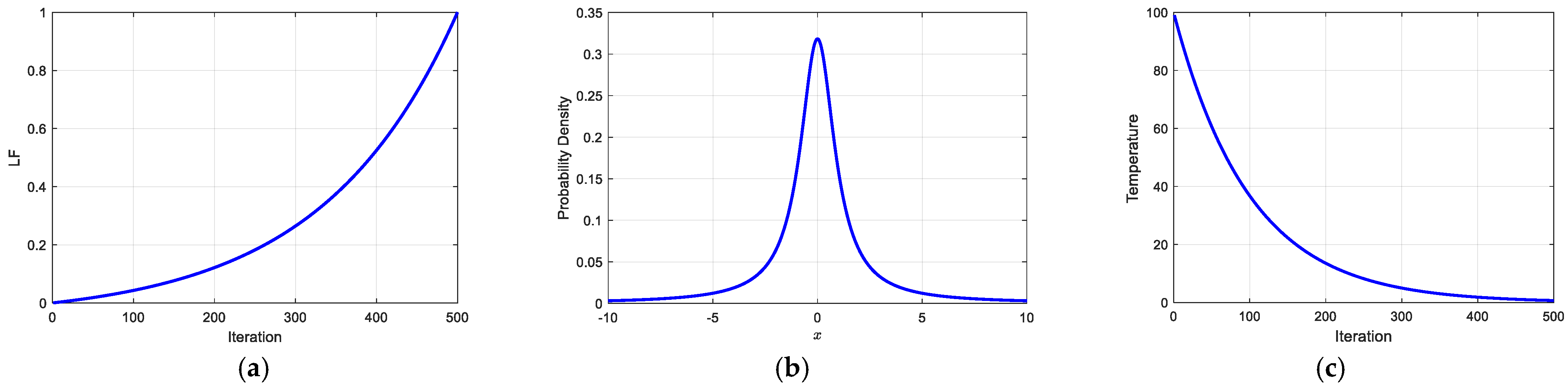
3.1.2. Dynamic Optimal Leadership Mechanism
3.1.3. Cauchy Mutation Strategy
3.1.4. Simulated Annealing Algorithm
3.1.5. Computational Complexity
3.1.6. Algorithm Pseudocode
| Algorithm 1 |
| Input: Initialize parameters ps, dim, np, , , k, , , , and MaxFEs (maximum function evaluations). Output: The global optima and . |
| 1 Initialize population = ,…, 2 for i = 1 → ps do 3 // tent map 4 , j = 1, 2,…,dim // ROBL 5 // initial population consolidation 6 sort(), i = 1, 2,…, 2ps, X , …, // screened populations according to fitness 7 end for 8 while FEs < MaxFEs do // check whether the maximum number of iterations is reached 9 sort(), i = 1, 2,…, ps; 10 ; // proportion fraction 11 12 for i = 1: ps do // ergodic population 13 if i is in then 14 if then 15 Calculate using Equation (10) // dormancy 16 Calculate // calculate the difference in fitness 17 Calculate P using Equation (27) // probability of acceptance 18 if then 19 else 20 end if 21 else 22 23 Calculate using Equation (11) // reproduction 24 Calculate // calculate the difference in fitness 25 Calculate P using Equation (27) // probability of acceptance 26 if then 27 else 28 end if 29 end if 30 else 31 32 if then 33 Calculate using Equation (24) // nonlinear dynamic adjustment factor 34 Calculate using Equation (25) // foraging by an autotroph 35 else 36 Calculate using Equation (24) // calculate the Cauchy factor 37 Calculate using Equation (5) // foraging by a heterotroph 38 end if 39 end if 40 if then 41 else 42 end if 43 end for 44 // update the optimal solution 45 46 end while |
3.2. Experiments and Analyses
3.2.1. Development Environment Settings
3.2.2. CEC2022 Benchmark Function Test Results and Analysis
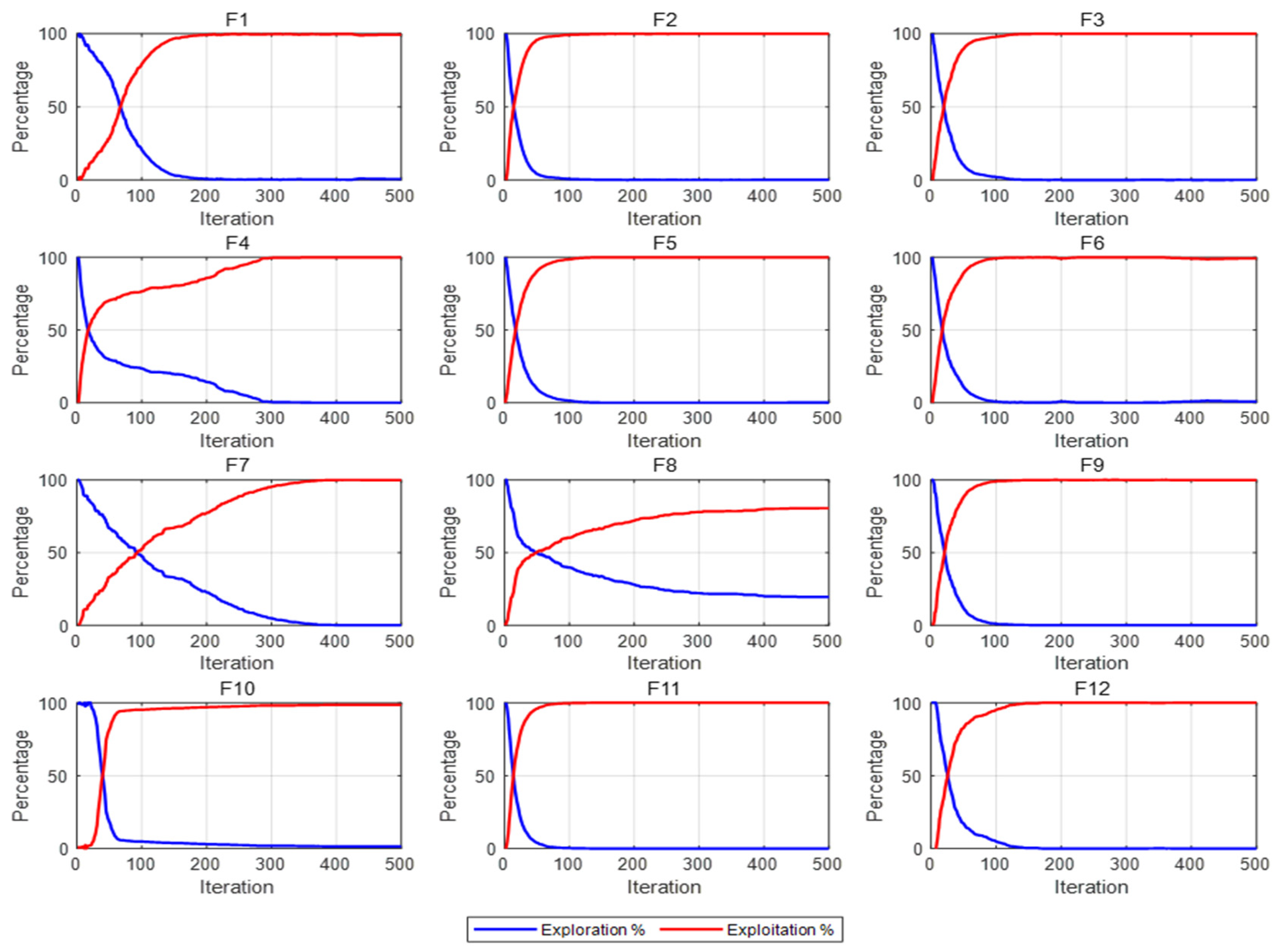
3.2.3. Exploration and Development Analyses
3.2.4. Engineering Applications and Ablation Experiments
- Welding beam design
- 2.
- Tension/compression spring design
4. Overview of UAV Path Planning
4.1. Problem Modeling
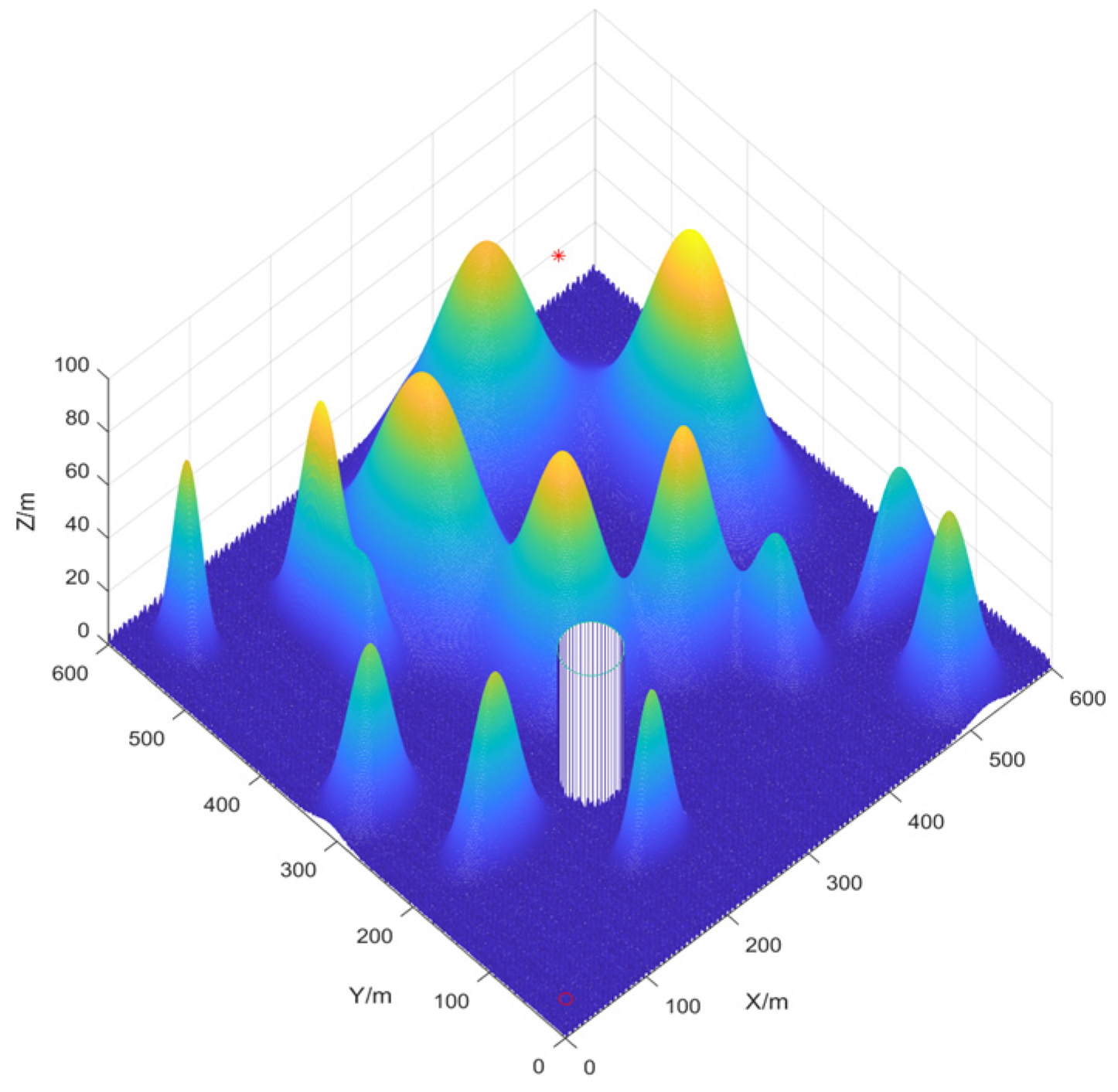
Three-Dimensional Environment Modeling
4.2. UAV Path Planning Modeling
4.2.1. Initial Conditions and Collision Detection
4.2.2. Path Length Costs
4.2.3. Curvature Constraint Costs
4.2.4. Cost of Height Variation
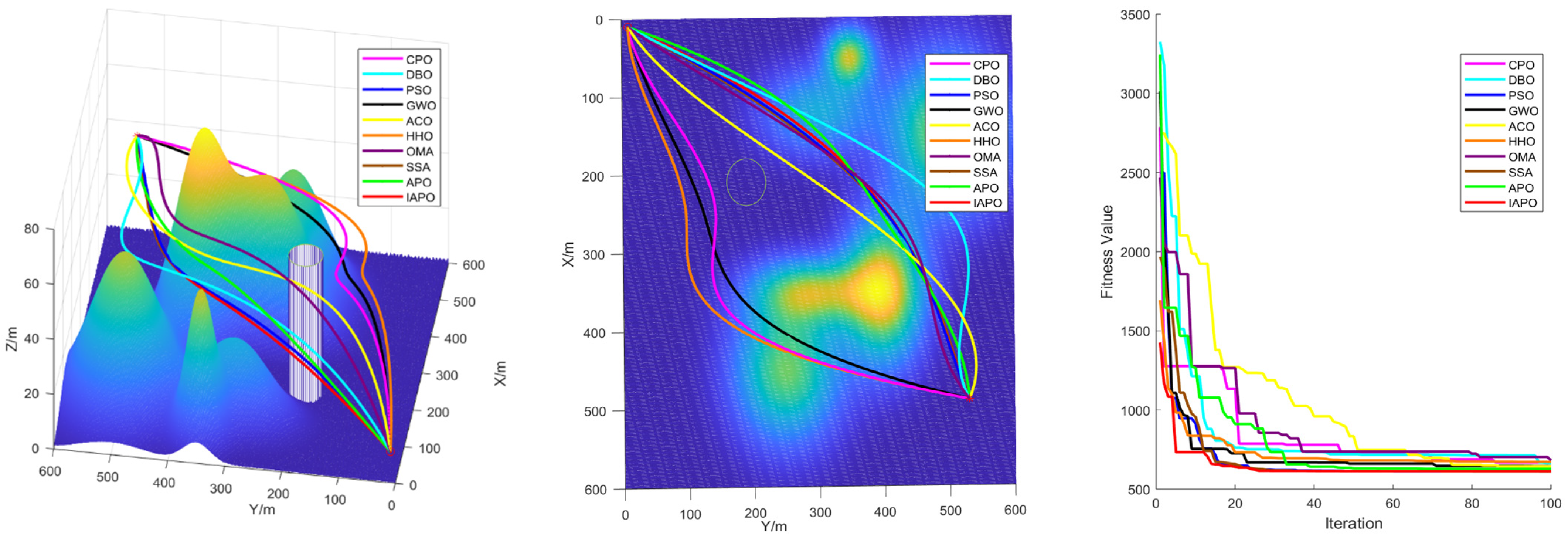
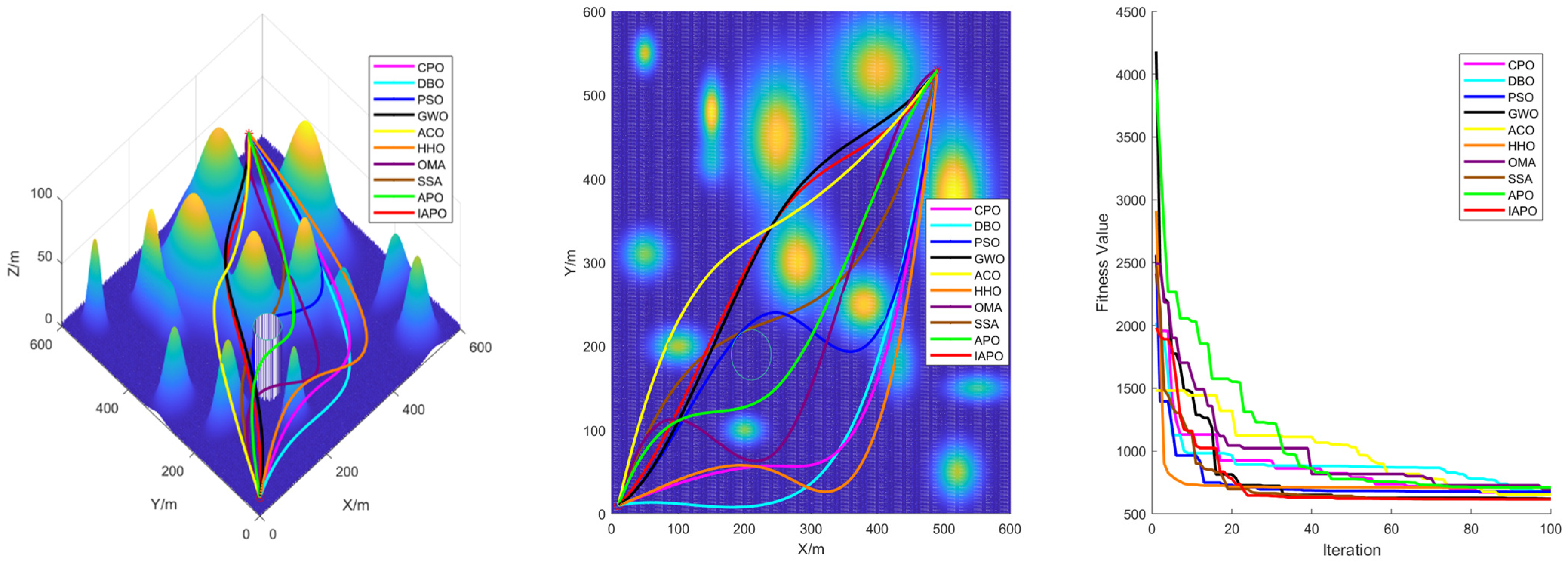
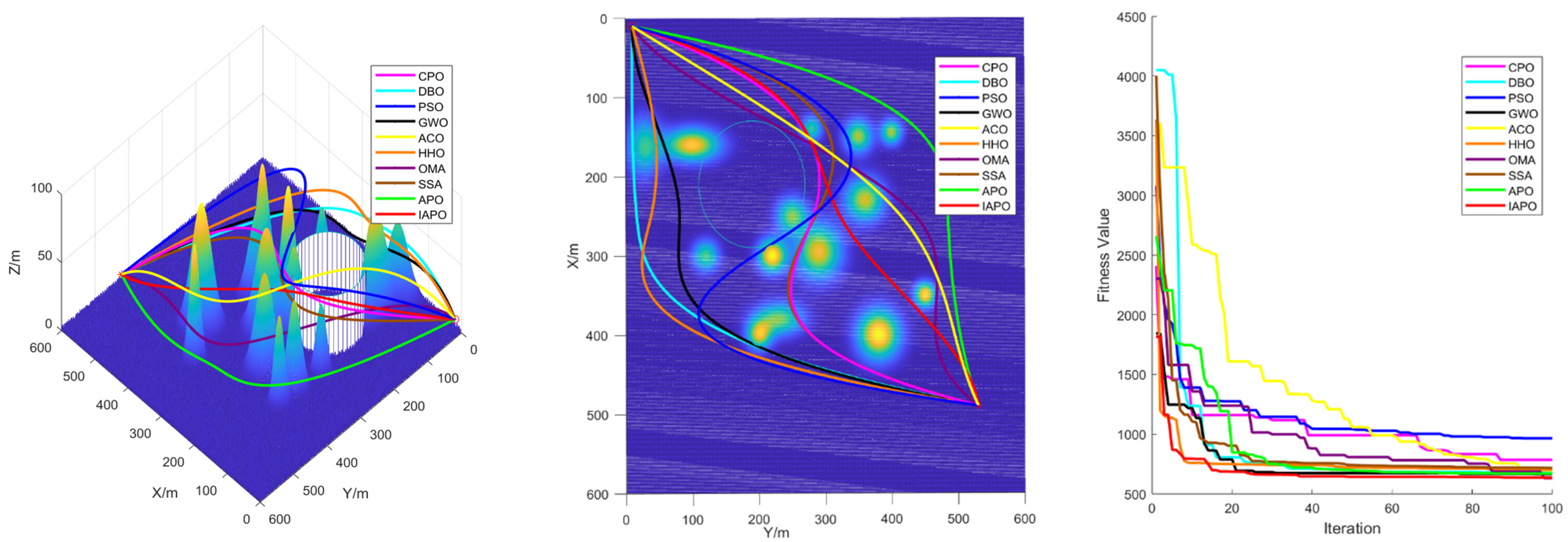
4.3. Comparison of Various Algorithms for UAV Path Planning
4.3.1. Environmental Settings
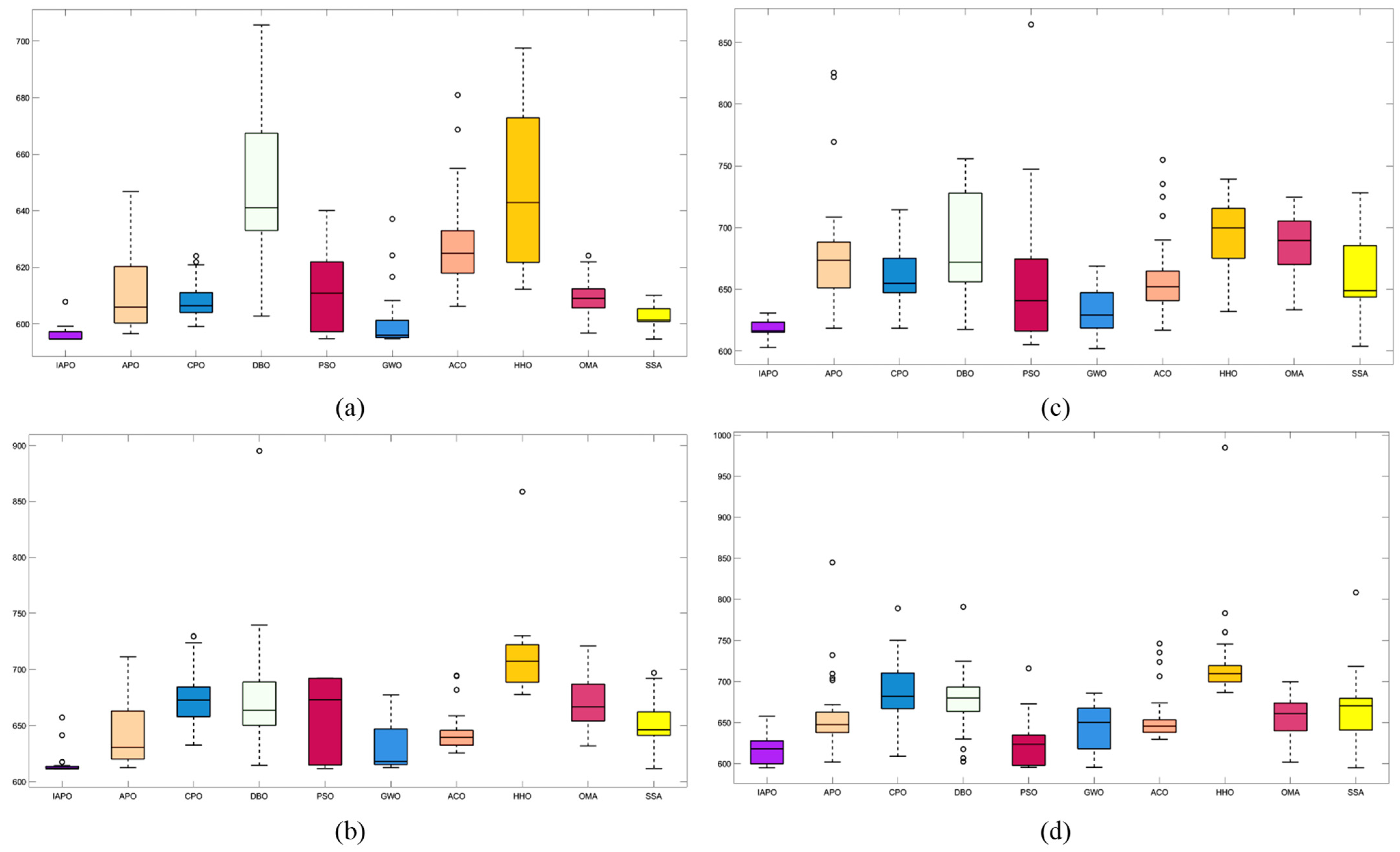
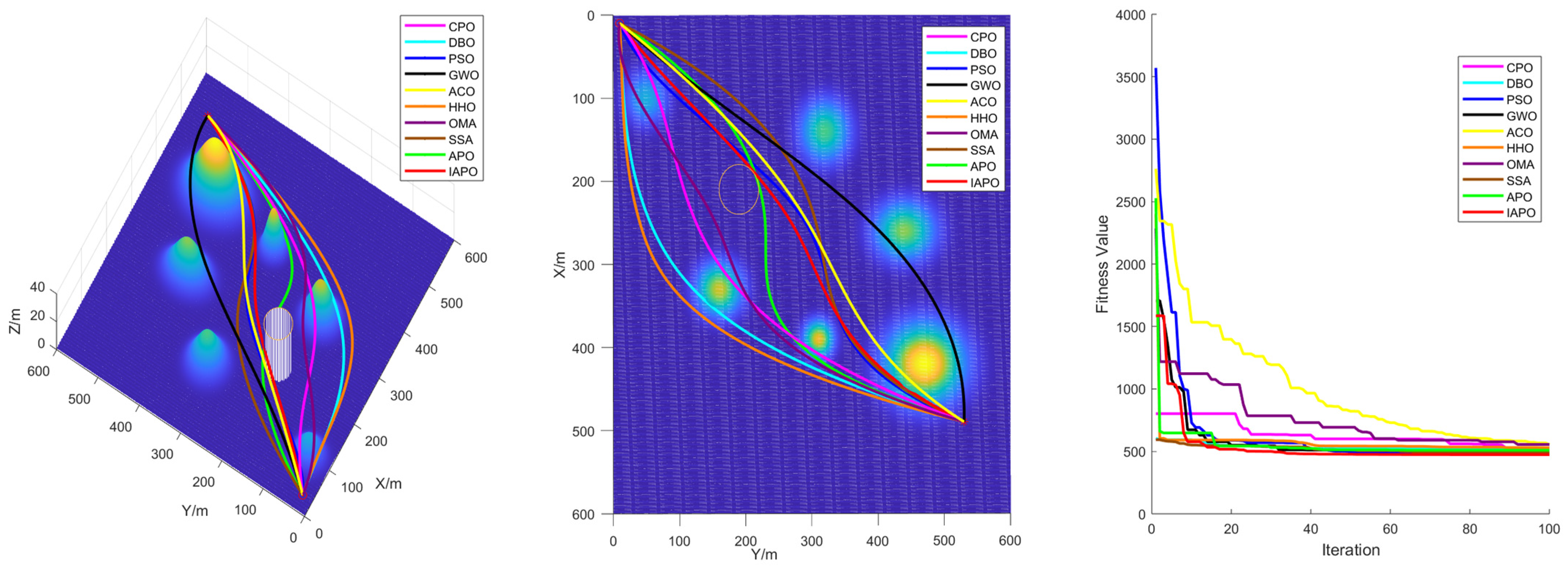
4.3.2. Algorithm Comparison Experiments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Nomenclature |
|---|---|
| Population size. | |
| Number of decision variables. | |
| Dimension index. | |
| Number of neighbor pairs. | |
| Foraging factor. | |
| Weight factor in autotrophs. | |
| Weight factor in heterotrophs. | |
| Proportion fraction of dormancy and reproduction. | |
| Probability of autotrophic and heterotrophic behavior. | |
| Probability of dormancy and reproduction. | |
| Random number in the range [0,1]. | |
| Current iteration. | |
| Maximum iteration. | |
| ith protozoan. | |
| Lower-bound vector. | |
| Upper-bound vector. | |
| Mapping vector in foraging. | |
| Mapping vector in reproduction. | |
| Index vector in dormancy and reproduction. | |
| Random vector with elements in [0,1]. | |
| Ceiling function. | |
| Fitness function. | |
| Ranking based on fitness values from smallest to largest. | |
| Returns a row vector containing l unique integers that are randomly selected between 1 and n. | |
| Hadamard product. |
References
- Liu, H.; Tsang, Y.P.; Lee, C.K.M. UAV trajectory planning via viewpoint resampling for autonomous remote inspection of industrial facilities. IEEE Trans. Ind. Inform. 2024, 20, 7492–7501. [Google Scholar] [CrossRef]
- Subrahmanyam, V.; Sagar, M.; Balram, G. An Efficient Reliable Data Communication For Unmanned Air Vehicles (UAV) Enabled Industry Internet of Things (IIoT). In Proceedings of the 2024 3rd International Conference on Artificial Intelligence For Internet of Things (AIIoT), Vellore, India, 3–4 May 2024; IEEE: Piscataway, NJ, USA, 2024; pp. 1–4. [Google Scholar]
- Velusamy, P.; Rajendran, S.; Mahendran, R.K. Unmanned Aerial Vehicles (UAV) in precision agriculture: Applications and challenges. Energies 2021, 15, 217. [Google Scholar] [CrossRef]
- Rejeb, A.; Abdollahi, A.; Rejeb, K. Drones in agriculture: A review and bibliometric analysis. Comput. Electron. Agric. 2022, 198, 107017. [Google Scholar]
- Betti Sorbelli, F. UAV-Based Delivery Systems: A Systematic Review, Current Trends, and Research Challenges. J. Auton. Transp. Syst. 2024, 1, 1–40. [Google Scholar]
- Mohamed, N.; Al-Jaroodi, J.; Jawhar, I. Unmanned aerial vehicles applications in future smart cities. Technol. Forecast. Soc. Change 2020, 153, 119293. [Google Scholar]
- Motlagh, N.H.; Kortoçi, P.; Su, X. Unmanned aerial vehicles for air pollution monitoring: A survey. IEEE Internet Things J. 2023, 10, 21687–21704. [Google Scholar] [CrossRef]
- Lyu, M.; Zhao, Y.; Huang, C. Unmanned aerial vehicles for search and rescue: A survey. Remote Sens. 2023, 15, 3266. [Google Scholar] [CrossRef]
- Hart, P.E.; Nilsson, N.J.; Raphael, B. A formal basis for the heuristic determination of minimum cost paths. IEEE Trans. Syst. Sci. Cybern. 1968, 4, 100–107. [Google Scholar]
- Dijkstra, E.W. A note on two problems in connexion with graphs. In Edsger Wybe Dijkstra: His Life, Work, and Legacy; Association for Computing Machinery: New York, NY, USA, 2022; pp. 287–290. [Google Scholar]
- Kuffner, J.J.; LaValle, S.M. RRT-connect: An efficient approach to single-query path planning. In Proceedings of the Proceedings 2000 ICRA. Millennium Conference. IEEE International Conference on Robotics and Automation. Symposia Proceedings (Cat. No. 00CH37065), San Francisco, CA, USA, 24–28 April 2000; IEEE: Piscataway, NJ, USA, 2000; Volume 2, pp. 995–1001. [Google Scholar]
- Sonny, A.; Yeduri, S.R.; Cenkeramaddi, L.R. Autonomous UAV path planning using modified PSO for UAV-assisted wireless networks. IEEE Access 2023, 11, 70353–70367. [Google Scholar]
- Yu, Z.; Si, Z.; Li, X. A novel hybrid particle swarm optimization algorithm for path planning of UAVs. IEEE Internet Things J. 2022, 9, 22547–22558. [Google Scholar]
- Deng, L.; Chen, H.; Zhang, X. Three-dimensional path planning of UAV based on improved particle swarm optimization. Mathematics 2023, 11, 1987. [Google Scholar] [CrossRef]
- Chen, J.; Ling, F.; Zhang, Y. Coverage path planning of heterogeneous unmanned aerial vehicles based on ant colony system. Swarm Evol. Comput. 2022, 69, 101005. [Google Scholar] [CrossRef]
- He, Y.; Zeng, Q.; Liu, J. Path planning for indoor UAV based on Ant Colony Optimization. In Proceedings of the2013 25th Chinese Control and Decision Conference (CCDC), Guiyang, China, 25–27 May 2013; IEEE: Piscataway, NJ, USA, 2013; pp. 2919–2923. [Google Scholar]
- Sonmez, A.; Kocyigit, E.; Kugu, E. Optimal path planning for UAVs using genetic algorithm. In Proceedings of the 2015 International Conference on Unmanned Aircraft Systems (ICUAS), Denver, CO, USA, 9–12 June 2015; IEEE: Piscataway, NJ, USA, 2015; pp. 50–55. [Google Scholar]
- Leng, S.; Sun, H. UAV path planning in 3D complex environments using genetic algorithms. In Proceedings of the 2021 33rd Chinese Control and Decision Conference (CCDC), Kunming, China, 22–24 May 2021; IEEE: Piscataway, NJ, USA, 2021; pp. 1324–1330. [Google Scholar]
- Nikolos, I.K.; Valavanis, K.P.; Tsourveloudis, N.C. Evolutionary algorithm based offline/online path planner for UAV navigation. IEEE Trans. Syst. Man Cybern. Part B Cybern. 2003, 33, 898–912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, S.; Wu, F. Path planning of UAV based on improved adaptive grey wolf optimization algorithm. IEEE Access 2021, 9, 89400–89411. [Google Scholar] [CrossRef]
- Qu, C.; Gai, W.; Zhang, J. A novel hybrid grey wolf optimizer algorithm for unmanned aerial vehicle (UAV) path planning. Knowl.-Based Syst. 2020, 194, 105530. [Google Scholar] [CrossRef]
- Lin, S.; Li, F.; Li, X. Improved artificial bee colony algorithm based on multi-strategy synthesis for UAV path planning. IEEE Access 2022, 10, 119269–119282. [Google Scholar] [CrossRef]
- Lei, L.; Shiru, Q. Path planning for unmanned air vehicles using an improved artificial bee colony algorithm. In Proceedings of the 31st Chinese Control Conference, Hefei, China, 25–27 July 2012; IEEE: Piscataway, NJ, USA, 2012; pp. 2486–2491. [Google Scholar]
- Zhang, T.; Yu, L.; Li, S. Unmanned aerial vehicle 3D path planning based on an improved artificial fish swarm algorithm. Drones 2023, 7, 636. [Google Scholar] [CrossRef]
- Tian, D. Particle swarm optimization with chaos-based initialization for numerical optimization. Intell. Autom. Soft Comput. 2018, 24, 331–342. [Google Scholar] [CrossRef]
- Haghighi, H.; Sadati, S.H.; Dehghan, S.M.M. Hybrid form of particle swarm optimization and genetic algorithm for optimal path planning in coverage mission by cooperated unmanned aerial vehicles. J. Aerosp. Technol. Manag. 2020, 12, e4320. [Google Scholar] [CrossRef]
- Tu, B.; Wang, F.; Huo, Y. A hybrid algorithm of grey wolf optimizer and harris hawks optimization for solving global optimization problems with improved convergence performance. Sci. Rep. 2023, 13, 22909. [Google Scholar] [CrossRef]
- Zhou, X.; Gao, F.; Fang, X. Improved bat algorithm for UAV path planning in three-dimensional space. IEEE Access 2021, 9, 20100–20116. [Google Scholar] [CrossRef]
- Li, F.; Fan, X.; Hou, Z. A firefly algorithm with self-adaptive population size for global path planning of mobile robot. IEEE Access 2020, 8, 168951–168964. [Google Scholar]
- Wang, X.; Snášel, V.; Mirjalili, S. Artificial Protozoa Optimizer (APO): A novel bio-inspired metaheuristic algorithm for engineering optimization. Knowl.-Based Syst. 2024, 295, 111737. [Google Scholar] [CrossRef]
- Gokhale, S.S.; Kale, V.S. An application of a tent map initiated Chaotic Firefly algorithm for optimal overcurrent relay coordination. Int. J. Electr. Power Energy Syst. 2016, 78, 336–342. [Google Scholar]
- Zheng-Ming, G.A.O.; Juan, Z.; Yu-Rong, H.U. The improved Harris hawk optimization algorithm with the Tent map. In Proceedings of the2019 3rd International Conference on Electronic Information Technology and Computer Engineering (EITCE), Xiamen, China, 18–20 October 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 336–339. [Google Scholar]
- Shao, P.; Wu, Z.J.; Zhou, X.Y. Improved particle swarm optimization algorithm based on opposite learning of refraction. Acta Electron. Sin. 2015, 43, 2137–2144. [Google Scholar]
- Xiao, Y.; Sun, X.; Guo, Y. An enhanced honey badger algorithm based on Lévy flight and refraction opposition-based learning for engineering design problems. J. Intell. Fuzzy Syst. 2022, 43, 4517–4540. [Google Scholar]
- Sun, L.; Feng, B.; Chen, T. Equalized grey wolf optimizer with refraction opposite learning. Comput. Intell. Neurosci. 2022, 2022, 2721490. [Google Scholar] [CrossRef]
- Tizhoosh, H.R. Opposition-based learning: A new scheme for machine intelligence. In Proceedings of the International Conference on Computational Intelligence for Modelling, Control and Automation and International Conference on Intelligent Agents, Web Technologies and Internet Commerce (CIMCA-IAWTIC’06), Vienna, Austria, 28–30 November 2005; IEEE: Piscataway, NJ, USA, 2005; Volume 1, pp. 695–701. [Google Scholar]
- Zhao, X.; Fang, Y.; Liu, L. A covariance-based Moth–flame optimization algorithm with Cauchy mutation for solving numerical optimization problems. Appl. Soft Comput. 2022, 119, 108538. [Google Scholar]
- Yang, F.; Jiang, H.; Lyu, L. Multi-strategy fusion improved Northern Goshawk optimizer is used for engineering problems and UAV path planning. Sci. Rep. 2024, 14, 23300. [Google Scholar]
- Kirkpatrick, S.; Gelatt, C.D., Jr.; Vecchi, M.P. Optimization by simulated annealing. Science 1983, 220, 671–680. [Google Scholar]
- Kumar, A.; Price, K.V.; Mohamed, A.W.; Hadi, A.A.; Suganthan, P.N. Problem Definitions and Evaluation Criteria for the CEC 2022 Special Session and Competition on Single Objective Bound Constrained Numerical Optimization; Technical Report; Nanyang Technological University: Singapore, 2021. [Google Scholar]
- Abdel-Basset, M.; Mohamed, R.; Abouhawwash, M. Crested Porcupine Optimizer: A new nature-inspired metaheuristic. Knowl.-Based Syst. 2024, 284, 111257. [Google Scholar] [CrossRef]
- Xue, J.; Shen, B. Dung beetle optimizer: A new meta-heuristic algorithm for global optimization. J. Supercomput. 2023, 79, 7305–7336. [Google Scholar] [CrossRef]
- Kennedy, J.; Eberhart, R. Particle swarm optimization. In Proceedings of the ICNN’95-International Conference on Neural Networks, Perth, Australia, 27 November–1 December 1995; IEEE: Piscataway, NJ, USA, 1995; Volume 4, pp. 1942–1948. [Google Scholar]
- Mirjalili, S.; Mirjalili, S.M.; Lewis, A. Grey wolf optimizer. Adv. Eng. Softw. 2014, 69, 46–61. [Google Scholar] [CrossRef]
- Dorigo, M.; Birattari, M.; Stutzle, T. Ant colony optimization. IEEE Comput. Intell. Mag. 2006, 1, 28–39. [Google Scholar] [CrossRef]
- Heidari, A.A.; Mirjalili, S.; Faris, H. Harris hawks optimization: Algorithm and applications. Future Gener. Comput. Syst. 2019, 97, 849–872. [Google Scholar] [CrossRef]
- Cheng, M.Y.; Sholeh, M.N. Optical microscope algorithm: A new metaheuristic inspired by microscope magnification for solving engineering optimization problems. Knowl.-Based Syst. 2023, 279, 110939. [Google Scholar] [CrossRef]
- Gharehchopogh, F.S.; Namazi, M.; Ebrahimi, L. Advances in sparrow search algorithm: A comprehensive survey. Arch. Comput. Methods Eng. 2023, 30, 427–455. [Google Scholar] [CrossRef]
- Liu, C.A.; Wang, X.P.; Liu, C.Y. Three-dimensional trajectory planning for UAV based on an improved grey wolf optimization algorithm. J. Huazhong Univ. Sci. Technol. (Nat. Sci. Ed.) 2017, 45, 38–42. (In Chinese) [Google Scholar]
- Ma, M.; Wu, J.; Shi, Y. Chaotic random opposition-based learning and Cauchy mutation improved moth-flame optimization algorithm for intelligent route planning of multiple UAVs. IEEE Access 2022, 10, 49385–49397. [Google Scholar] [CrossRef]
- Phung, M.D.; Ha, Q.P. Safety-enhanced UAV path planning with spherical vector-based particle swarm optimization. Appl. Soft Comput. 2021, 107, 107376. [Google Scholar] [CrossRef]

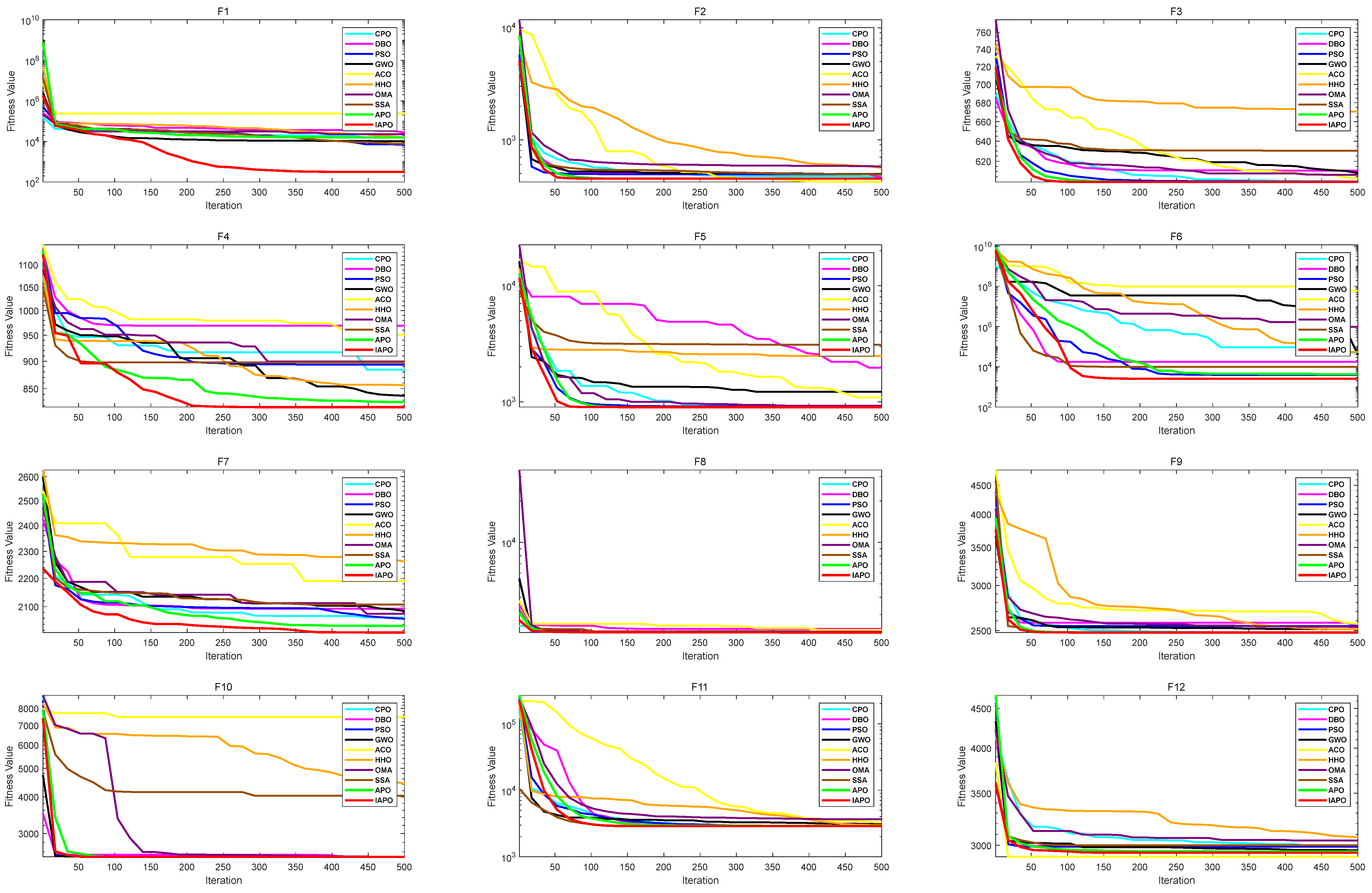
| No. | Functions | ||
|---|---|---|---|
| Unimodal function | F1 | Shifted and full Rotated Zakharov function | 300 |
| Multimodal functions | F2 | Shifted and full Rotated Rosenbrock’s function | 400 |
| F3 | Shifted and full Rotated Expanded Schaffer’s f6 function | 600 | |
| F4 | Shifted and full Rotated Non-Continuous Rastrigin’s function | 800 | |
| F5 | Shifted and full Rotated Levy function | 900 | |
| Hybrid functions | F6 | Hybrid function 1 (N = 3) | 1800 |
| F7 | Hybrid function 2 (N = 6) | 2000 | |
| F8 | Hybrid function 3 (N = 5) | 2200 | |
| Composition functions | F9 | Composition function 1 (N = 5) | 2300 |
| F10 | Composition function 2 (N = 4) | 2400 | |
| F11 | Composition function 3 (N = 5) | 2600 | |
| F12 | Composition function 4 (N = 6) | 2700 | |
| Fun. | Index | IAPO (Ours) | APO | CPO | DBO | PSO | GWO | ACO | HHO | OMA | SSA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| F1 | Std | 1.148 × 102 | 3.523 × 103 | 2.840 × 103 | 8.522 × 103 | 2.632 × 103 | 4.439 × 103 | 1.611 × 105 | 6.307 × 103 | 5.554 × 103 | 4.053 × 103 |
| Mean | 4.429 × 102 | 1.198 × 104 | 1.075× 104 | 2.752× 104 | 2.943× 103 | 1.393× 104 | 2.651× 105 | 1.714 × 104 | 2.609 × 104 | 7.182 × 103 | |
| Rank | 1 | 5 | 4 | 9 | 2 | 6 | 10 | 7 | 8 | 3 | |
| F2 | Std | 1.130 × 101 | 1.068 × 101 | 9.966 × 100 | 7.187× 101 | 3.039× 101 | 2.813× 101 | 2.098× 100 | 4.353× 101 | 1.989× 101 | 1.888× 101 |
| Mean | 4.565 × 102 | 4.572 × 102 | 4.579 × 102 | 4.989 × 102 | 4.609 × 102 | 4.919 × 102 | 4.202 × 102 | 5.267 × 102 | 4.967 × 102 | 4.497 × 102 | |
| Rank | 3 | 4 | 5 | 9 | 6 | 7 | 1 | 10 | 8 | 2 | |
| F3 | Std | 9.299 × 10−2 | 6.304× 10−3 | 8.963 × 10−2 | 1.032 × 101 | 2.809 × 100 | 2.664 × 100 | 1.271 × 100 | 6.892 × 100 | 2.072 × 100 | 1.348 × 101 |
| Mean | 6.000 × 102 | 6.000 × 102 | 6.003 × 102 | 6.223 × 102 | 6.031 × 102 | 6.046 × 102 | 6.048 × 102 | 6.599 × 102 | 6.085 × 102 | 6.338 × 102 | |
| Rank | 2 | 1 | 3 | 8 | 4 | 5 | 6 | 10 | 7 | 9 | |
| F4 | Std | 7.940 × 100 | 6.106 × 100 | 1.160 × 101 | 2.739 × 101 | 1.824 × 101 | 2.728 × 101 | 1.010 × 101 | 1.617 × 101 | 1.162 × 101 | 1.583 × 101 |
| Mean | 8.227 × 102 | 8.228 × 102 | 8.972 × 102 | 9.159 × 102 | 8.529 × 102 | 8.559 × 102 | 9.450 × 102 | 8.875 × 102 | 9.104 × 102 | 8.908 × 102 | |
| Rank | 1 | 2 | 7 | 9 | 3 | 4 | 10 | 5 | 8 | 6 | |
| F5 | Std | 1.493 × 100 | 3.503 × 100 | 4.298 × 100 | 5.115 × 102 | 9.182 × 101 | 2.102 × 102 | 9.114 × 101 | 2.669 × 102 | 5.331 × 101 | 1.569 × 102 |
| Mean | 9.012 × 102 | 9.034 × 102 | 9.025 × 102 | 1.803 × 103 | 9.418 × 102 | 1.156 × 103 | 1.130 × 103 | 2.894 × 103 | 9.508 × 102 | 2.423 × 103 | |
| Rank | 1 | 3 | 2 | 8 | 4 | 7 | 6 | 10 | 5 | 9 | |
| F6 | Std | 1.594 × 103 | 2.309 × 103 | 1.784 × 104 | 6.244× 105 | 4.909 × 103 | 9.110 × 106 | 7.155 × 107 | 7.753 × 104 | 3.647 × 105 | 4.925 × 103 |
| Mean | 3.263 × 103 | 4.073 × 103 | 3.315 × 104 | 2.029 × 105 | 5.566 × 103 | 3.980 × 106 | 1.186 × 108 | 1.428 × 105 | 9.551 × 105 | 6.247 × 103 | |
| Rank | 1 | 2 | 5 | 7 | 3 | 9 | 10 | 6 | 8 | 4 | |
| F7 | Std | 5.309 × 100 | 8.313 × 100 | 8.326 × 100 | 4.684 × 101 | 4.237 × 101 | 5.196 × 101 | 3.396 × 101 | 5.795 × 101 | 1.295 × 101 | 4.980 × 101 |
| Mean | 2.032 × 103 | 2.036 × 103 | 2.064 × 103 | 2.110 × 103 | 2.073 × 103 | 2.091 × 103 | 2.212 × 103 | 2.180 × 103 | 2.113 × 103 | 2.134 × 103 | |
| Rank | 1 | 2 | 3 | 6 | 4 | 5 | 10 | 9 | 7 | 8 | |
| F8 | Std | 5.520 × 10−1 | 1.342 × 100 | 2.011 × 100 | 7.317 × 101 | 6.294 × 101 | 4.799 × 101 | 6.138 × 101 | 1.053 × 102 | 4.272 × 100 | 6.800 × 101 |
| Mean | 2.223 × 103 | 2.223 × 103 | 2.232 × 103 | 2.297 × 103 | 2.262 × 103 | 2.255 × 103 | 2.366 × 103 | 2.312 × 103 | 2.242 × 103 | 2.303 × 103 | |
| Rank | 1 | 2 | 3 | 7 | 6 | 5 | 10 | 9 | 4 | 8 | |
| F9 | Std | 2.801 × 10−3 | 6.192 × 10−1 | 2.324 × 10−1 | 2.817 × 101 | 1.694 × 101 | 1.491 × 101 | 3.732 × 101 | 2.155 × 101 | 9.034 × 100 | 5.788 × 10−4 |
| Mean | 2.481 × 103 | 2.481 × 103 | 2.481 × 103 | 2.505 × 103 | 2.491 × 103 | 2.502 × 103 | 2.558 × 103 | 2.518 × 103 | 2.513 × 103 | 2.481 × 103 | |
| Rank | 2 | 3 | 4 | 7 | 5 | 6 | 10 | 9 | 8 | 1 | |
| F10 | Std | 4.070 × 100 | 2.079 × 102 | 3.497 × 101 | 8.306 × 102 | 3.451 × 102 | 7.538 × 102 | 5.485 × 102 | 7.745 × 102 | 4.840 × 101 | 6.223 × 102 |
| Mean | 2.502 × 103 | 2.602 × 103 | 2.507 × 103 | 3.012 × 103 | 2.919 × 103 | 3.504 × 103 | 7.251 × 103 | 4.001 × 103 | 2.519 × 103 | 3.609 × 103 | |
| Rank | 1 | 4 | 2 | 6 | 5 | 7 | 10 | 9 | 3 | 8 | |
| F11 | Std | 1.114 × 10−1 | 6.076 × 100 | 5.137 × 101 | 3.458 × 101 | 1.770 × 100 | 5.567 × 102 | 8.876 × 101 | 3.792 × 102 | 1.412 × 102 | 4.901 × 101 |
| Mean | 2.900 × 103 | 2.911 × 103 | 2.926 × 103 | 2.913 × 103 | 2.903 × 103 | 3.560 × 103 | 3.360 × 103 | 3.255 × 103 | 3.513 × 103 | 2.937 × 103 | |
| Rank | 1 | 3 | 5 | 4 | 2 | 10 | 8 | 7 | 9 | 6 | |
| F12 | Std | 4.225 × 100 | 4.797 × 100 | 9.071 × 100 | 3.196 × 101 | 3.153 × 101 | 2.728 × 101 | 3.869 × 10−5 | 1.449 × 102 | 1.406 × 101 | 2.801 × 101 |
| Mean | 2.940 × 103 | 2.942 × 103 | 2.990 × 103 | 2.993 × 103 | 2.988 × 103 | 2.977 × 103 | 2.900 × 103 | 3.181 × 103 | 3.031 × 103 | 2.984 × 103 | |
| Rank | 2 | 3 | 7 | 8 | 6 | 4 | 1 | 10 | 9 | 5 |
| Algorithm | Optimal Cost | ||||
|---|---|---|---|---|---|
| IAPO | 0.20535 | 3.2388 | 9.036 | 0.20571 | 1.69248479 |
| IAPO_I | 0.20562 | 3.4759 | 9.0384 | 0.20658 | 1.73218539 |
| IAPO_II | 0.20586 | 3.3658 | 9.0365 | 0.20621 | 1.71439872 |
| IAPO_III | 0.20559 | 3.2675 | 9.036 | 0.20568 | 1.69651910 |
| APO | 0.20598 | 3.4795 | 9.0384 | 0.20671 | 1.73423458 |
| Algorithm | Optimal Cost | |||
|---|---|---|---|---|
| IAPO | 0.051302 | 0.355984 | 11.248914 | 0.0124131 |
| IAPO_I | 0.052247 | 0.369845 | 11.648116 | 0.0137789 |
| IAPO_II | 0.051764 | 0.356974 | 11.588412 | 0.0129975 |
| IAPO_III | 0.051845 | 0.359874 | 11.256841 | 0.0128234 |
| APO | 0.052351 | 0.369845 | 11.648187 | 0.0138339 |
| Map | Index | Std | Mean | Rank | Map | Index | Std | Mean | Rank |
|---|---|---|---|---|---|---|---|---|---|
| I | IAPO | 2.726 | 596.120 | 1 | III | IAPO | 6.462 | 618.747 | 1 |
| APO | 13.842 | 611.741 | 7 | APO | 48.908 | 681.468 | 7 | ||
| CPO | 6.654 | 608.331 | 4 | CPO | 23.871 | 660.606 | 6 | ||
| DBO | 22.047 | 643.105 | 9 | DBO | 34.700 | 682.779 | 8 | ||
| PSO | 13.692 | 611.099 | 6 | PSO | 54.315 | 657.994 | 4 | ||
| GWO | 9.683 | 600.348 | 2 | GWO | 18.703 | 633.465 | 2 | ||
| ACO | 17.137 | 628.844 | 8 | ACO | 32.513 | 660.553 | 5 | ||
| HHO | 28.001 | 647.990 | 10 | HHO | 26.528 | 694.809 | 10 | ||
| OMA | 6.202 | 608.839 | 5 | OMA | 23.519 | 686.937 | 9 | ||
| SSA | 3.529 | 602.106 | 3 | SSA | 34.845 | 657.528 | 3 | ||
| II | IAPO | 10.800 | 615.974 | 1 | IV | IAPO | 18.109 | 618.315 | 1 |
| APO | 29.563 | 643.623 | 3 | APO | 44.660 | 659.520 | 6 | ||
| CPO | 22.185 | 673.490 | 8 | CPO | 40.063 | 685.515 | 9 | ||
| DBO | 54.947 | 680.091 | 9 | DBO | 37.149 | 679.394 | 8 | ||
| PSO | 34.967 | 657.843 | 6 | PSO | 28.245 | 626.163 | 2 | ||
| GWO | 20.417 | 632.021 | 2 | GWO | 30.816 | 641.589 | 3 | ||
| ACO | 17.650 | 643.988 | 4 | ACO | 30.870 | 656.397 | 4 | ||
| HHO | 32.769 | 709.538 | 10 | HHO | 53.868 | 721.477 | 10 | ||
| OMA | 20.673 | 670.385 | 7 | OMA | 23.622 | 658.574 | 5 | ||
| SSA | 25.322 | 648.945 | 5 | SSA | 40.118 | 662.862 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Na, X.; Feng, Z.; Hai, S.; Shi, J. Three-Dimensional UAV Path Planning Based on Multi-Strategy Integrated Artificial Protozoa Optimizer. Biomimetics 2025, 10, 201. https://doi.org/10.3390/biomimetics10040201
Sun Q, Na X, Feng Z, Hai S, Shi J. Three-Dimensional UAV Path Planning Based on Multi-Strategy Integrated Artificial Protozoa Optimizer. Biomimetics. 2025; 10(4):201. https://doi.org/10.3390/biomimetics10040201
Chicago/Turabian StyleSun, Qingbin, Xitai Na, Zhihui Feng, Shiji Hai, and Jinshuo Shi. 2025. "Three-Dimensional UAV Path Planning Based on Multi-Strategy Integrated Artificial Protozoa Optimizer" Biomimetics 10, no. 4: 201. https://doi.org/10.3390/biomimetics10040201
APA StyleSun, Q., Na, X., Feng, Z., Hai, S., & Shi, J. (2025). Three-Dimensional UAV Path Planning Based on Multi-Strategy Integrated Artificial Protozoa Optimizer. Biomimetics, 10(4), 201. https://doi.org/10.3390/biomimetics10040201