Maternal Lineages during the Roman Empire, in the Ancient City of Gadir (Cádiz, Spain): The Search for a Phoenician Identity

 ,
,  , , , and
, , , and
Abstract
1. Introduction
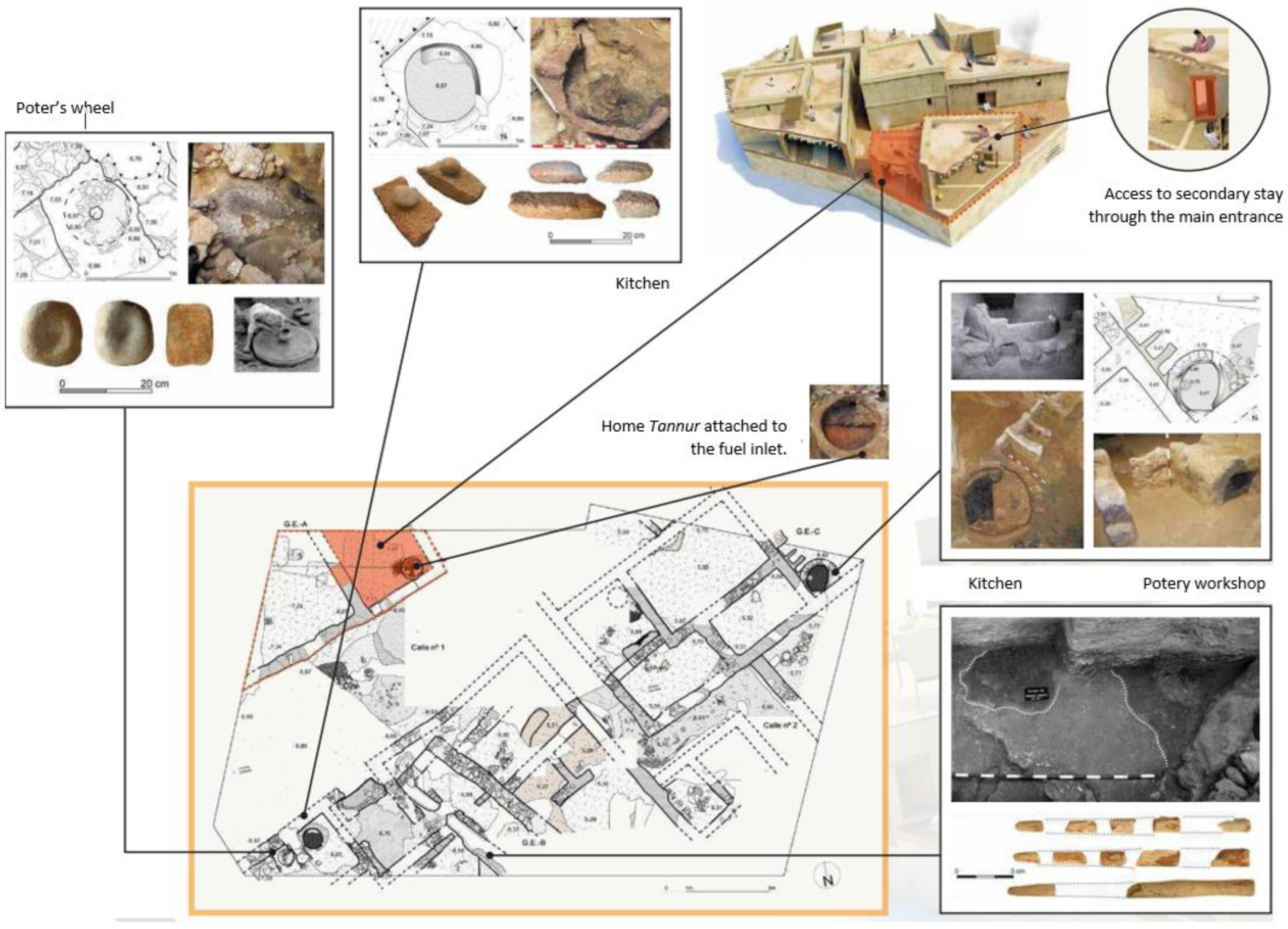
Archaeological Data
2. Materials and Methods
2.1. Anthropological Records
A Complex Burial: The Case of Individuals 8i and 8ii
2.2. Ancient DNA Analysis
2.2.1. Sampling Process
2.2.2. DNA Extraction
Method A: Destructive Procedure
Method B: Non-Destructive Technique
2.2.3. mtDNA Amplification
2.2.4. mtDNA Haplogroup Estimation
2.2.5. Specific Haplogroup SNP Amplification
2.3. Statistical Analysis
3. Results
3.1. Result Quality
3.2. Biogeographical Ancestry
Haplogroup-Specific SNP Results, Haplogroup Assignment, and Phylogenetic Analysis
3.3. Statistical Analysis
4. Discussion
4.1. Results of the Quality and Authenticity
4.2. Biogeographical Ancestry
4.2.1. The HV0 Lineage
4.2.2. The H Lineage
4.2.3. The L3 Lineage
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amadori, Maria Dvc, Paola Fermo, and Paloma Pallante. 2016. Archaeometric researches on the provenance of Mediterranean Archaic Phoenician and Punic Pottery. Environmental Science and Pollution Research 24: 13921–49. [Google Scholar] [CrossRef] [PubMed]
- Arruda, ana Margarida. 2009. Phoenician Characterization on the Atlantic Coast of the Iberian Peninsula. Chicago: The University of Chicago Press. [Google Scholar]
- Aubet, Maria Eugenia. 2001. The Phoenicians and the West: Politics, Colonies and Trade. Cambridge: Cambridge University Press. [Google Scholar]
- Barral-Arca, Ruth, Sara Pischedda, Alberto Gómez-Carballa, Ana Pastoriza, Ana Mosquera-Miguel, Manuel López-Soto, Federico Martinón-Torres, Vanesa Álvarez-Iglesias, and Antonio Salas. 2016. Meta-Analysis of Mitochondrial DNA Variation in the Iberian Peninsula. PLoS ONE 11: e0159735. [Google Scholar] [CrossRef] [PubMed]
- Cavalli-Sforza, Luca, and Marcus Feldman. 2003. The application of molecular genetic approaches to the study of human evolution. Nature Genetics 33: 266–75. [Google Scholar] [CrossRef]
- De Fanti, Sara, Chiara Barbieri, Stefania Sarno, Federica Sevini, Dario Vianello, Erika Tamm, Ene Metspalu, Mannis van Oven, Alexander Hübner, Marco Sazzini, and et al. 2015. Fine Dissection of Human Mitochondrial DNA Haplogroup HV Lineages Reveals Paleolithic Signatures from European Glacial Refugia. PLoS ONE 10: e0144391. [Google Scholar] [CrossRef] [PubMed]
- Di Bernardo, Giovanni, Stefania Del Gaudio, Umberto Galderisi, Antonio Cascino, and Marilena Cipollaro. 2009. Ancient DNA and family relationships in a Pompeian house. Annals of Human Genetics 73: 429–37. [Google Scholar]
- Di Giacomo, Fabio, Luis Popa, Nejat Akar, Nicholas Anagnou, Juraj Banyko, Radim Brdicka, Guido Barbujani, Franco Papola, Giovanni Ciavarella, Franceso Cucci, and et al. 2004. Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe. Human Genetics 115: 357–71. [Google Scholar] [CrossRef] [PubMed]
- Dietler, Michael, and Carolina López-Ruiz. 2009. Colonial Encounters in Ancient Iberia: Phoenicians, Greek, and Indigenous Relations. Chicago: Press UoC. [Google Scholar]
- Emery, Leslie, Kevin Magnaye, Abigail Bigham, Joshua Akey, and Michael Bamshad. 2015. Estimates of continental ancestry vary widely among individuals with the same mtDNA haplogroup. American Journal of Human Genetics 96: 183–93. [Google Scholar] [CrossRef]
- Fernández Uriel, Pilar, Carlos González Wagner, and Fernando López Pardo. 2000. Editor Intercambio y comercio preclásico en el Mediterráneo. In I coloquio del Centro de Estudios Fenicios y Púnicos (CEFYP). Madrid: Actas del I coloquio del CEFYP. [Google Scholar]
- Friedrich-Schiller-Universitaet Jena. 2012. Oldest Jewish Archaeological Evidence on the Iberian Peninsula. Available online: http://www.uni-jena.de/en/News/PM120525_Schrifttafel.html (accessed on 14 October 2022).
- Gamba, Cristina, Eva Fernández, Alexis Oliver, Miriam Tirado, Carlos Baeza, ana María López-Parra, and Eduardo Arroyo-Pardo. 2008. Population genetics and DNA preservation in ancient human remains from Eastern Spain. Forensic Science International: Genetics Supplement Series 1: 462–64. [Google Scholar] [CrossRef]
- Gamba, Cristina, Eva Fernandez, Miriam Tirado, M. F. Deguilloux, M. H. Pemonge, P. Utrilla, M. Edo, M. Molist, R. Rasteiro, L. Chikhi, and et al. 2012. Ancient DNA from an Early Neolithic Iberian population supports a pioneer colonization by first farmers. Molecular Ecology 21: 45–56. [Google Scholar] [CrossRef] [PubMed]
- Gener, osé María, and Carlos Núñez. 2015. Gadir Archaeological Site. In The Phoenician Origin—Making off. Cádiz: Ayuntamiento de Cádiz. [Google Scholar]
- Gomes, Cláudia, María Fondevila, Concepción Magaña-Loarte, Juan Fernández-Jiménez, José Fernández-Serrano, Sara Palomo-Díez, Carlos Baeza-Richer, Ana María López-Parra, and Eduardo Arroyo-Pardo. 2019. An unusual kinship case from the Spanish Civil War (1936–1939) Ancient versus degraded sample’s investigation. Forensic Science International: Genetics Supplement Series 7: 690–91. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Sara Palomo-Díez, ana María López-Parra, and Eduardo Arroyo-Pardo. 2021. Genealogy: The Tree Where History Meets Genetics. Genealogy 5: 98. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Sara Palomo-Díez, Jordi Roig, ana María López-Parra, Carlos Baeza-Richer, Angel Esparza-Arroyo, Juan Gibaja, and Eduardo Arroyo-Pardo. 2015. Nondestructive extraction DNA method from bones or teeth, true or false? Forensic Science International: Genetics Supplement Series 5: e279–82. [Google Scholar] [CrossRef]
- Groot, Beatrijs de. 2012. Testing Hybridity in Early Iron Age Iberia: Using Ceramics and Theory to Explain the Significance of Phoenician Colonialism. Leiden: Leiden University. [Google Scholar]
- Harich, Nourdin, Marta D. Costa, Verónica Fernandes, Mostafa Kandil, Joana B. Pereira, Nuno M. Silva, and Luísa Pereira. 2010. The trans-Saharan slave trade—Clues from interpolation analyses and high-resolution characterization of mitochondrial DNA lineages. BMC Evolutionary Biology 10: 138. [Google Scholar] [CrossRef] [PubMed]
- Kloss-Brandstatter, Anita, Dominic Pacher, Sebastian Schönherr, Hansi Weissensteiner, Robert Binna, Günther Specht, and Florian Kronenberg. 2011. HaploGrep: A fast and reliable algorithm for automatic classification of mitochondrial DNA haplogroups. Human Mutation 32: 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kujanova, Martina, Luisa Pereira, Verónica Fernandes, Joana Pereira, and Viktor Cerny. 2009. Near eastern neolithic genetic input in a small oasis of the Egyptian Western Desert. American Journal of Physical Anthropology 140: 336–46. [Google Scholar] [CrossRef]
- Levy-Coffman, Ellen. 2005. A mosaic of people: The Jewish story and reassessment of the DNA evidence. Journal of Genetic Genealogy 1: 12–33. [Google Scholar]
- Maca-Meyer, Nicole, Ana González, José Larruga, Carlos Flores, and Vicente M. Cabrera. 2001. Major genomic mitochondrial lineages delineate early human expansions. BMC Genetics 2: 13. [Google Scholar] [CrossRef] [PubMed]
- Mairal, Quim, Cristina Santos, Marina Silva, Sofia Marques, Amanda Ramos, Maria Pilar Aluja, Antonio Amorim, Maria João Prata, and Luis Alvarez. 2013. Linguistic isolates in Portugal: Insights from the mitochondrial DNA pattern. Forensic Science International: Genetics 7: 618–23. [Google Scholar] [CrossRef]
- Martiniano, Rui, Anwen Caffell, Malin Holst, Kurt Hunter-Mann, Janet Montgomery, Gundula Müldner, Russell L. McLaughlin, Matthew D. Teasdale, Wouter van Rheenen, Jan H. Veldink, and et al. 2016. Genomic signals of migration and continuity in Britain before the Anglo-Saxons. Nature Communications 7: 10326. [Google Scholar] [CrossRef]
- Matisoo-Smith, Elizabeth A., Anna L. Gosling, James Boocock, Olga Kardailsky, Yara Kurumilian, Sihem Roudesli-Chebbi, Leila Badre, Jean-Paul Morel, Leïla Ladjimi Sebaï, and Pierre A. Zalloua. 2016. A European Mito-chondrial Haplotype Identified in Ancient Phoenician Remains from Carthage, North Africa. PLoS ONE 11: e0155046. [Google Scholar] [CrossRef]
- Nogueiro, Inês, João Teixeira, António Amorim, Leonor Gusmao, and Luis Alvarez. 2015. Echoes from Sepharad: Signatures on the maternal gene pool of crypto-Jewish descendants. European Journal of Human Genetics 23: 693–99. [Google Scholar] [CrossRef][Green Version]
- Ottoni, Claudio, Giuseppina Primativo, Baharak Hooshiar Kashani, Alessandro Achilli, Cristina Martínez-Labarga, Gianfranco Biondi, Antonio Torroni, and Olga Rickards. 2010. Mitochondrial haplogroup H1 in North Africa: An early Holocene arrival from Iberia. PLoS ONE 5: e13378. [Google Scholar] [CrossRef] [PubMed]
- Pääbo, Svante, Hendrik Poinar, David Serre, Viviane Jaenicke-Despres, Juliane Hebler, Nadin Rohland, Melanie Kuch, Johannes Krause, Linda Vigilant, and Michael Hofreiter. 2004. Genetic analyses from ancient DNA. Annual Review of Genetics 38: 645–79. [Google Scholar] [CrossRef] [PubMed]
- Padró, Josep. 2012. Os Fenícios. In Grande História Universal—Os Grandes Impérios Orientais, 4. 1ª ed. Edited by C. M. Abón. Madrid: Ediclube, pp. 47–63. [Google Scholar]
- Parson, Walther, and Arne Dur. 2007. EMPOP—A forensic mtDNA database. Forensic Science International: Genetics 1: 88–92. [Google Scholar] [CrossRef]
- Prieto, Lourdes, Bettina Zimmermann, Ana Goios, Ana María Rodriguez-Monge, Greiciane Paneto, Cíntia Alves, Antonio Alonso, Cíntia Fridman, Sergio Cardoso, Grabiela Lima, and et al. 2011. The GHEP-EMPOP collaboration on mtDNA population data—A new resource for forensic casework. Forensic Science International: Genetics 5: 146–51. [Google Scholar] [CrossRef] [PubMed]
- Rohland, Nadin, and Michael Hofreiter. 2007. Ancient DNA extraction from bones and teeth. Nature Protocols 2: 1756–62. [Google Scholar] [CrossRef] [PubMed]
- Romanini, Carola, Magdalena Romero, Mercedes Salado Puerto, Laura Catelli, Christopher Phillips, Rui Pereira, Leonor Gusmão, and Carlos Vullo. 2015. Ancestry informative markers: Inference of ancestry in aged bone samples using an autosomal AIM-Indel multiplex. Forensic Science International: Genetics 16: 58–63. [Google Scholar] [CrossRef]
- Roostalu, Urmas, Ildus Kutuev, E.-L. Loogväli, Ene Metspalu, Kristiina Tambets, M. Reidla, Elza K. Khusnutdinova, Esien Usanga, Toomas Kivisild, and Richard Villems. 2007. Origin and expansion of haplogroup H, the dominant human mitochondrial DNA lineage in West Eurasia: The Near Eastern and Caucasian perspective. Molecular Biology and Evolution 24: 436–48. [Google Scholar] [CrossRef]
- Shriver, Mark, and Rick A. Kittles. 2004. Genetic ancestry and the search for personalized genetic histories. Nature Reviews Genetics 5: 611–18. [Google Scholar] [CrossRef]
- Töpf Ana, Thomas Gilbert, John Dumbacher, and Rus Hoelzel. 2006. Tracing the phylogeography of human populations in Britain based on 4th-11th century mtDNA genotypes. Molecular Biology and Evolution 23: 152–61. [Google Scholar] [CrossRef][Green Version]
- Töpf, Ana L., Thomas Gilbert, Robert Fleischer, and Rus Hoelzel. 2007. Ancient human mtDNA genotypes from England reveal lost variation over the last millennium. Biology Letters 3: 550–53. [Google Scholar] [CrossRef][Green Version]
- Tsirkin, Juli. 1997. The Phoenicians and Tartessos. Gerión Revista de Historia Antigua 15: 243. [Google Scholar]
- Valério, Pedro, Rui Silva, Maria Fátima Araújo, António Soares, and Luis Barros. 2012. A multianalytical approach to study the Phoenician Bronze technology in the Iberia Peninsula—A view from the Quinta do Almaraz. Materials Characterization 67: 74–82. [Google Scholar] [CrossRef]
- van Oven, Mannis. 2015. PhyloTree Build 17: Growing the human mitochondrial DNA tree. Forensic Science International: Genetics Supplement Series 5: e392–94. [Google Scholar] [CrossRef]
- van Oven, Mannis, and Manfred Kayser. 2009. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Human Mutation 30: E386–94. [Google Scholar] [CrossRef] [PubMed]
- Vijande, Eduardo, Antonio Sáez, Salvador Domínguez-Bella, Juan Jesús Cantillo, Antonio Sáez, Milagros Macías, Ana Juliá, Ésther Sánchez, Antonio Barrena, and Lourdes Lorenzo. 2007. Anuario arqueológico de Andalucia. Junta de Andalucía: Conserjería de Cultura. [Google Scholar]
- Wallace, Douglas C., Michael D. Brown, and Marie Lott. 1999. Mitochondrial DNA variation in human evolution and disease. Gene 38: 211–30. [Google Scholar] [CrossRef]
- Zalloua, Pierre, Catherine Collins, Anna Gosling, Simone Biagini, Benjamí Costa, Olga Kardailsky, Lorenzo Nigro, Wissam Khalil, Francesc Calafell, and Elizabeth Matisoo-Smith. 2018. Ancient DNA of Phoenician remains indicates discontinuity in the settlement history of Ibiza. Scientific Reports 8: 17567. [Google Scholar] [CrossRef]
- Zalloua, Pierre A., Daniel E. Platt, Mirvat El Sibai, Jade Khalife, Nadine Makhoul, Marc Haber, Yali Xue, Hassan Izaabel, Elena Bosch, Susan M. Adams, and et al. 2008. Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean. The American Journal of Human Genetics 83: 633–42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual | mtDNA Position Range | mtDNA Haplotype | HAPLOGREP | EMPOP Frequency (17 March 2023) | Haplogroup-Specific SNP Result | Assigned Haplogroup | Consensus Haplogroup |
|---|---|---|---|---|---|---|---|
| 1G | --- | No result | --- | --- | --- | --- | --- |
| 2G | --- | No result | --- | --- | --- | --- | --- |
| 3G | --- | Without consensus | --- | --- | --- | --- | --- |
| 4G | --- | Without consensus | --- | --- | --- | --- | --- |
| 5G | --- | Without consensus | --- | --- | --- | --- | --- |
| 6G | --- | Without consensus | --- | --- | --- | --- | --- |
| 7G | 123–320; 16,105–16,399 | 195C 198T 263G 309.1C 16269G 16298C | HV0b 86.18% | HV0b 1/42,210 | 14766C | HV0 | HV0 |
| 7028T | HV0 | ||||||
| 8iG | 8–128; 16,105–16,280 | 72C | HV0b 100% | HV0b 413/40,258 | 14766C | HV0 | HV0 |
| 7028T | HV0 | ||||||
| 8iiG | 10–390; 16,105–16,399 | 72C 195C 198T 263G 309.1C 16269G 16298C | HV0b 89.02% | HV0b 0/39,355 | 14766C | HV0 | HV0 |
| 7028T | HV0 | ||||||
| 9G | 8–73; 127–391; 16,105–16,399 | 72C 195C 198T 263G 315.1C 16269G 16298C | HV0b 89.02% | HV0b 1/39,355 | 7028T | HV0 | HV0 |
| 10G | 55–127; 135–378; 740–769; 16,231–16,399 | 73G 263G 315.1C 750G 16278T 16362C | L3b 100% | L3b 17/4300 | 10873C | L3 | L3 |
| 10398A | No result | ||||||
| 8701A | No result | ||||||
| 11G | 8–390; 16,105–16,399 | 152C 263G 315.1C | H1c | H 290/39,355 | 7028C | H | H |
| 12G | 126–390; 16,105–16,399 | 195C 198T 263G 309.1C 16269G 16278T 16298C 16362C | HV0b 80.1% | HV0b 0/41,708 | 7028T | HV0 | HV0 |
| 13G | ---- | Without consensus | --- | --- | --- | --- | --- |
| 14G | ---- | No result | --- | --- | --- | --- | --- |
| 15G | 8–390; 16,105–16,399 | 263G 315.1C | H2a2a 100% | H 1237/39,355 | 7028C | H | H |
| Researcher C. | 8–390; 16,105–16,399 | T152C, A263G, 309.1C, 315.1C, T16304C | H5 | H5 | --- | --- | --- |
| Researcher S. | 8–390; 16,105–16,399 | A55G, C150T, T239C, A263G, 309.1C, 309.2C, 315.1C, T16362C | H6a | H6a | --- | --- | --- |
| H1 | H2 | LR |
|---|---|---|
| African | European | 10,561.3604 |
| Eastern | African | 11.8762 |
| Eastern | European | 125,428.8424 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, C.; González Wagner, C.; Calero-Fresneda, M.; Palomo-Díez, S.; López-Matayoshi, C.; Nogueiro, I.; López-Parra, A.M.; González, E.L.; Pérez, B.P.; Basallote, J.M.G.; et al. Maternal Lineages during the Roman Empire, in the Ancient City of Gadir (Cádiz, Spain): The Search for a Phoenician Identity. Genealogy 2023, 7, 27. https://doi.org/10.3390/genealogy7020027
Gomes C, González Wagner C, Calero-Fresneda M, Palomo-Díez S, López-Matayoshi C, Nogueiro I, López-Parra AM, González EL, Pérez BP, Basallote JMG, et al. Maternal Lineages during the Roman Empire, in the Ancient City of Gadir (Cádiz, Spain): The Search for a Phoenician Identity. Genealogy. 2023; 7(2):27. https://doi.org/10.3390/genealogy7020027
Chicago/Turabian StyleGomes, Cláudia, Carlos González Wagner, Manuel Calero-Fresneda, Sara Palomo-Díez, César López-Matayoshi, Inês Nogueiro, Ana María López-Parra, Elena Labajo González, Bernardo Perea Pérez, José María Gener Basallote, and et al. 2023. "Maternal Lineages during the Roman Empire, in the Ancient City of Gadir (Cádiz, Spain): The Search for a Phoenician Identity" Genealogy 7, no. 2: 27. https://doi.org/10.3390/genealogy7020027
APA StyleGomes, C., González Wagner, C., Calero-Fresneda, M., Palomo-Díez, S., López-Matayoshi, C., Nogueiro, I., López-Parra, A. M., González, E. L., Pérez, B. P., Basallote, J. M. G., Pajuelo, J. M., & Arroyo Pardo, E. (2023). Maternal Lineages during the Roman Empire, in the Ancient City of Gadir (Cádiz, Spain): The Search for a Phoenician Identity. Genealogy, 7(2), 27. https://doi.org/10.3390/genealogy7020027