
Effect of Dopamine on Growth, Some Biochemical Attributes, and the Yield of Crisphead Lettuce under Nitrogen Deficiency

 ,
,  ,
, 
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Layout
- T1, 100% RN without DA
- T2, 100% RN with 50 µM DA
- T3, 100% RN with 100 µM DA
- T4, 50% RN without DA
- T5, 50% RN with 50 µM DA
- T6, 50% TN with 100 µM DA
2.2. Sampling
2.2.1. Vegetative Growth and Yield Attributes
2.2.2. Photosynthetic Pigments and Chemical Constituents
2.2.3. Malondialdehyde, Hydrogen Peroxide, and Superoxide Anion
2.2.4. Antioxidants Enzymes
2.2.5. Head Quality
2.3. Statistical Analysis
3. Results
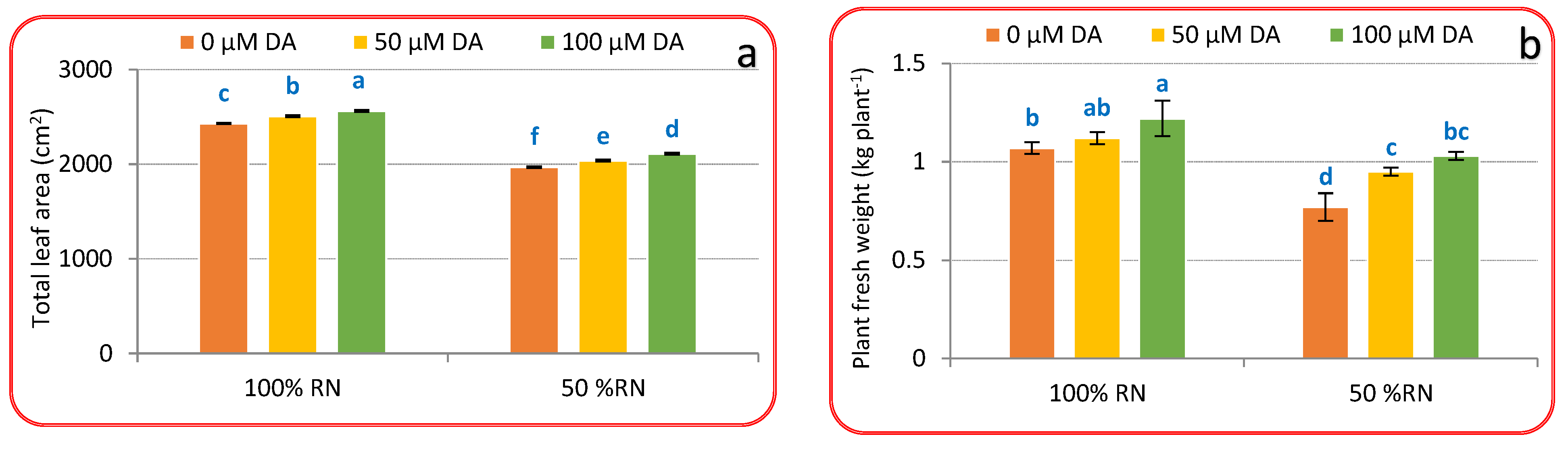
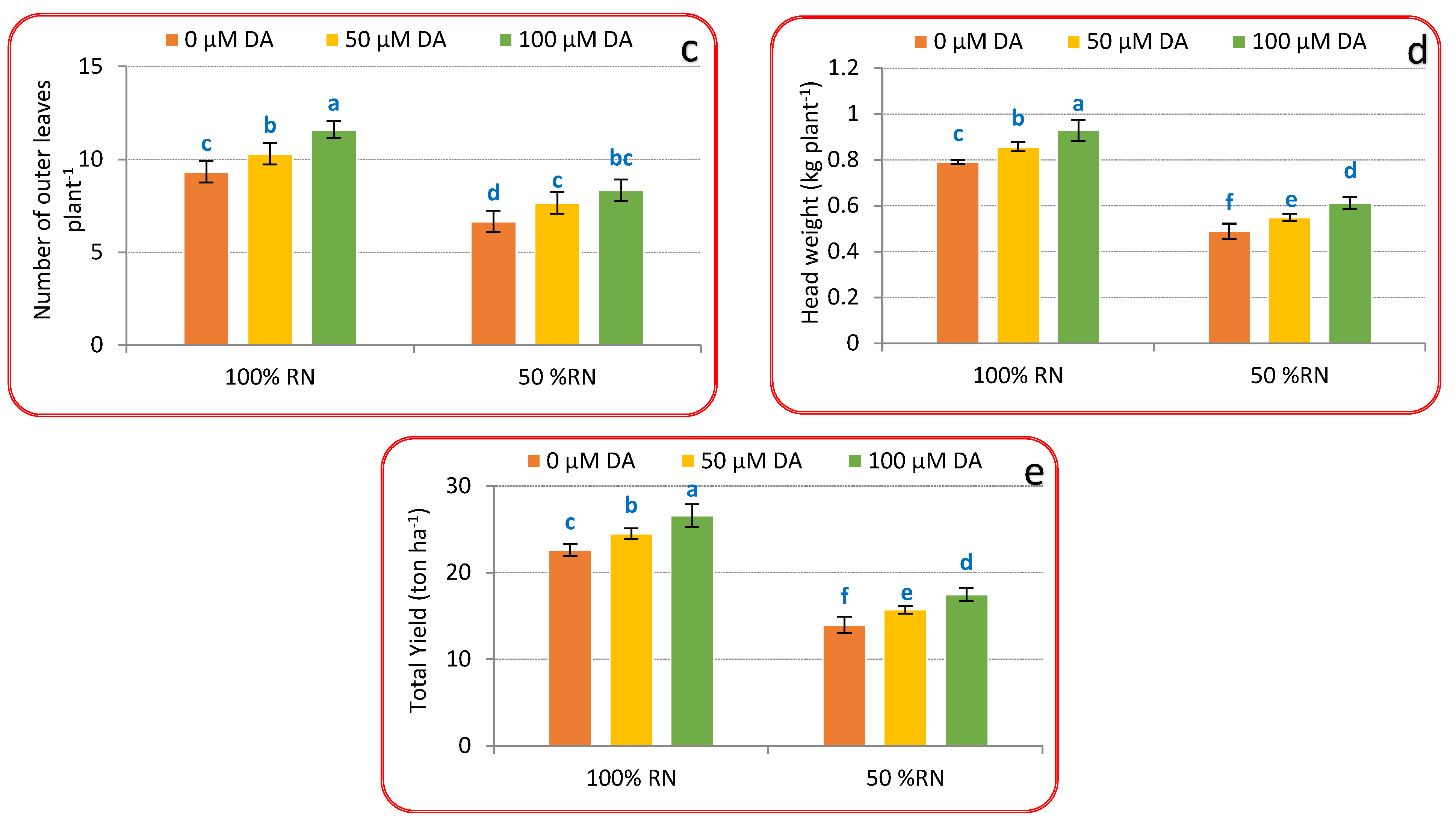
3.1. Vegetative Growth Criteria
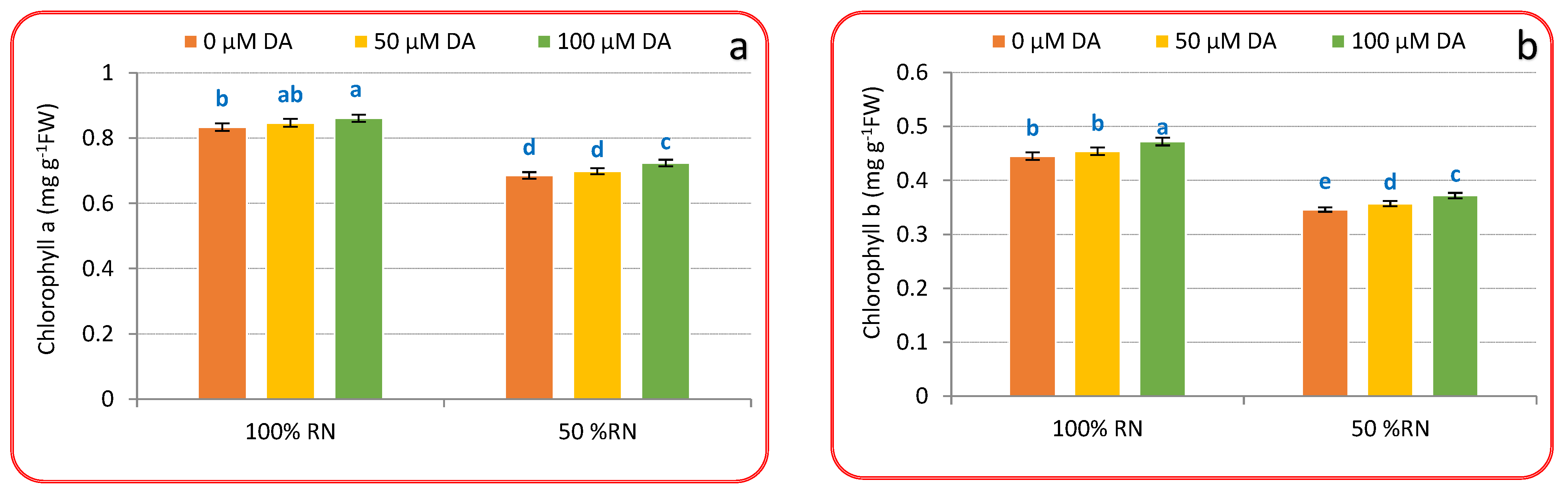
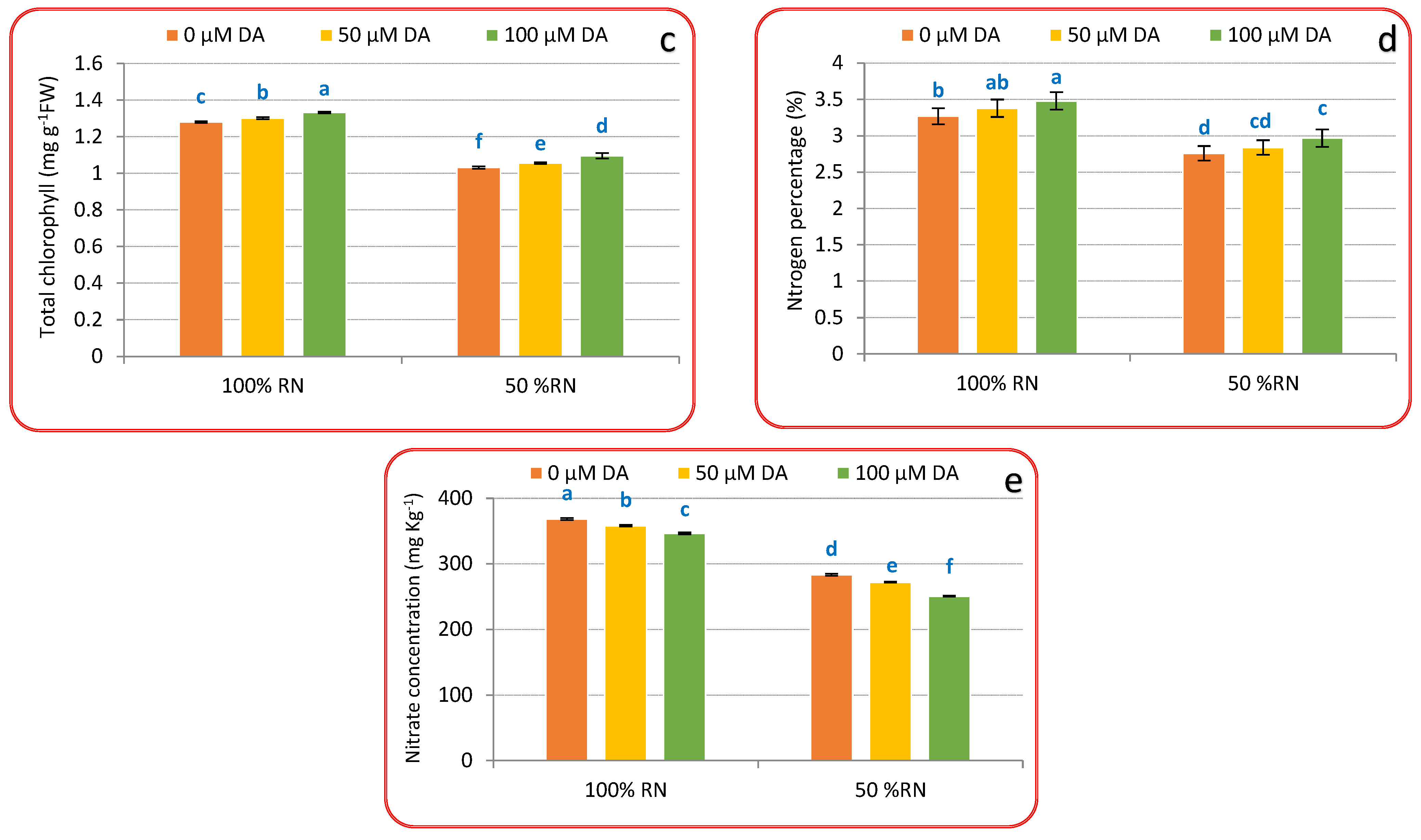
3.2. Photosynthetic Pigments, Nitrogen% and Nitrate Concentration
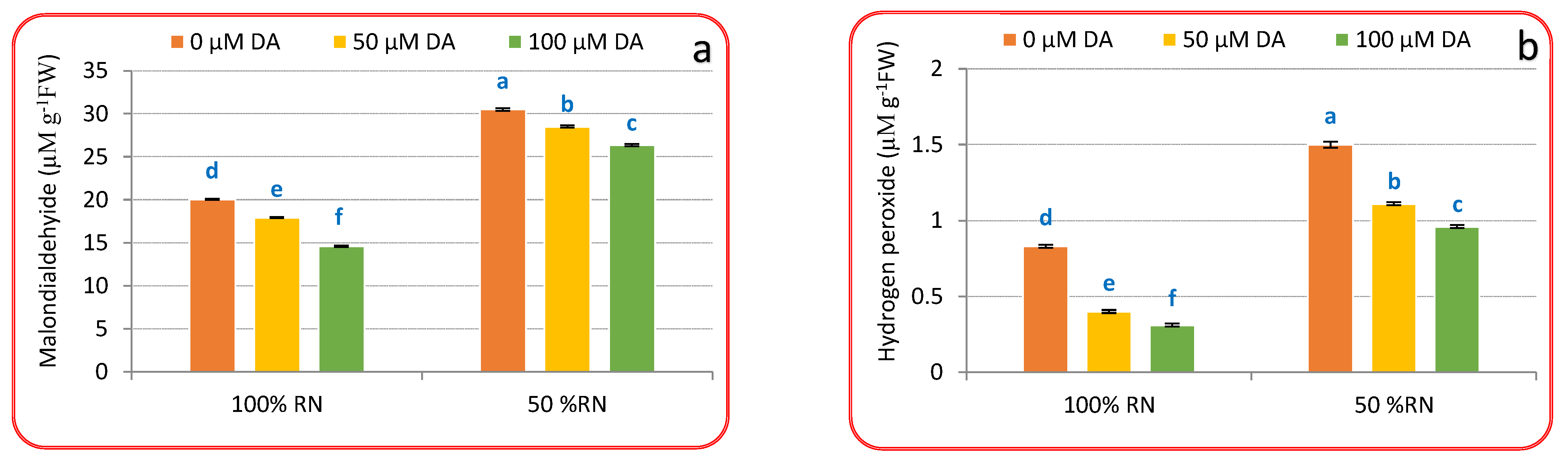
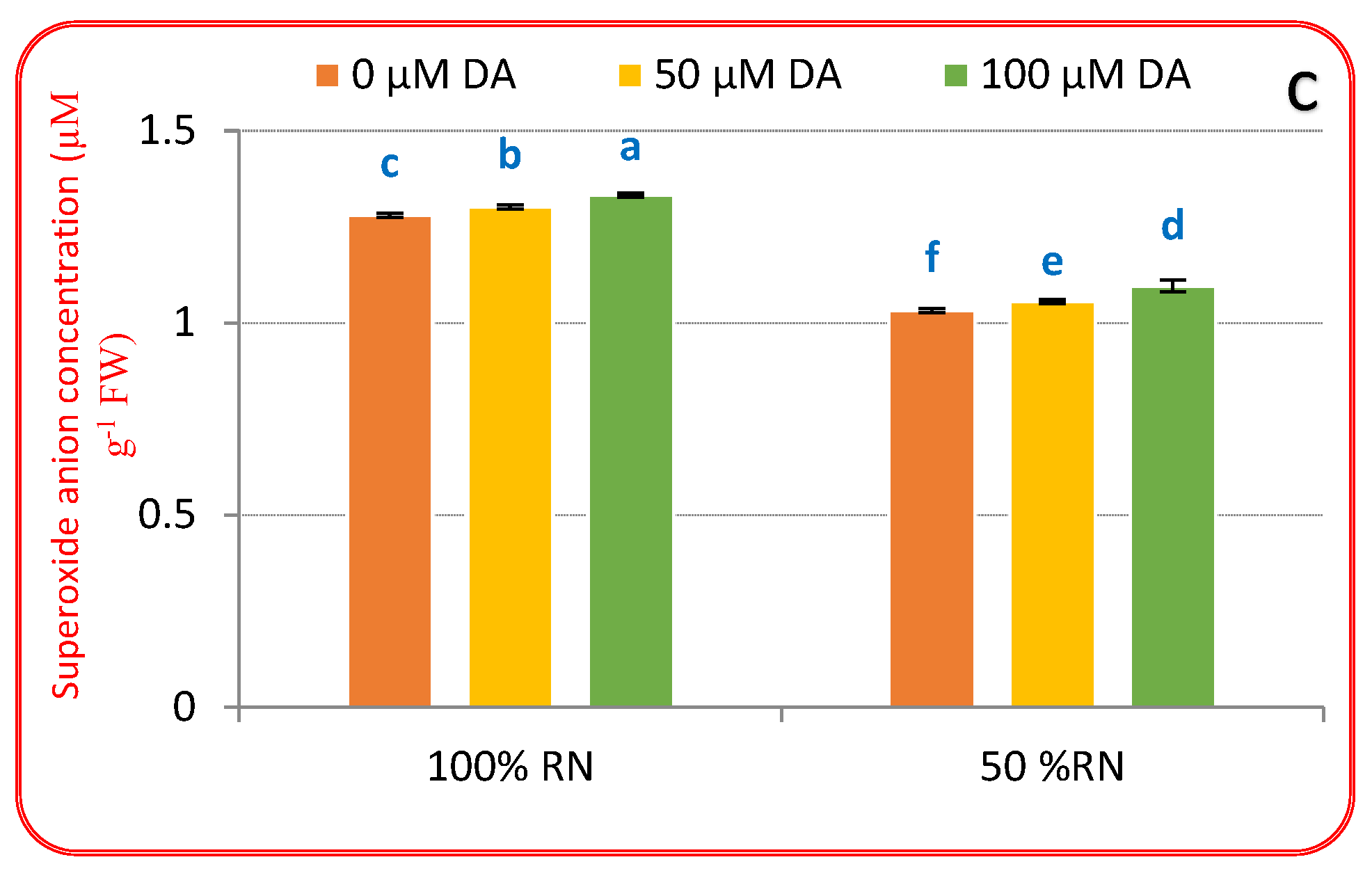
3.3. Oxidative Biomarkers
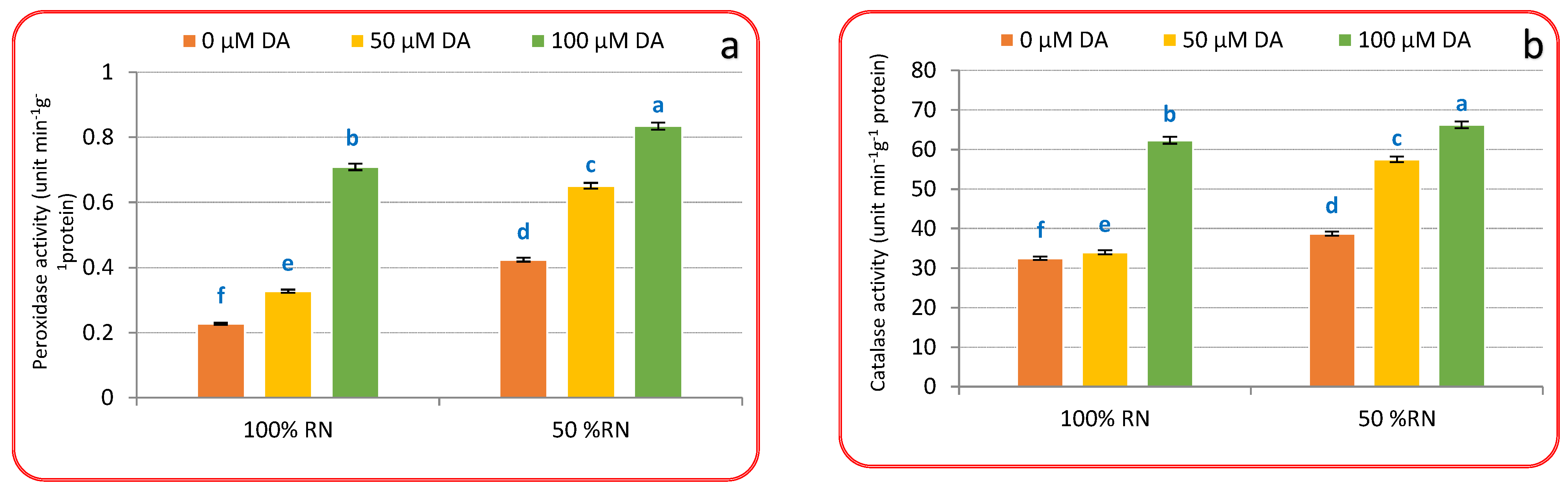
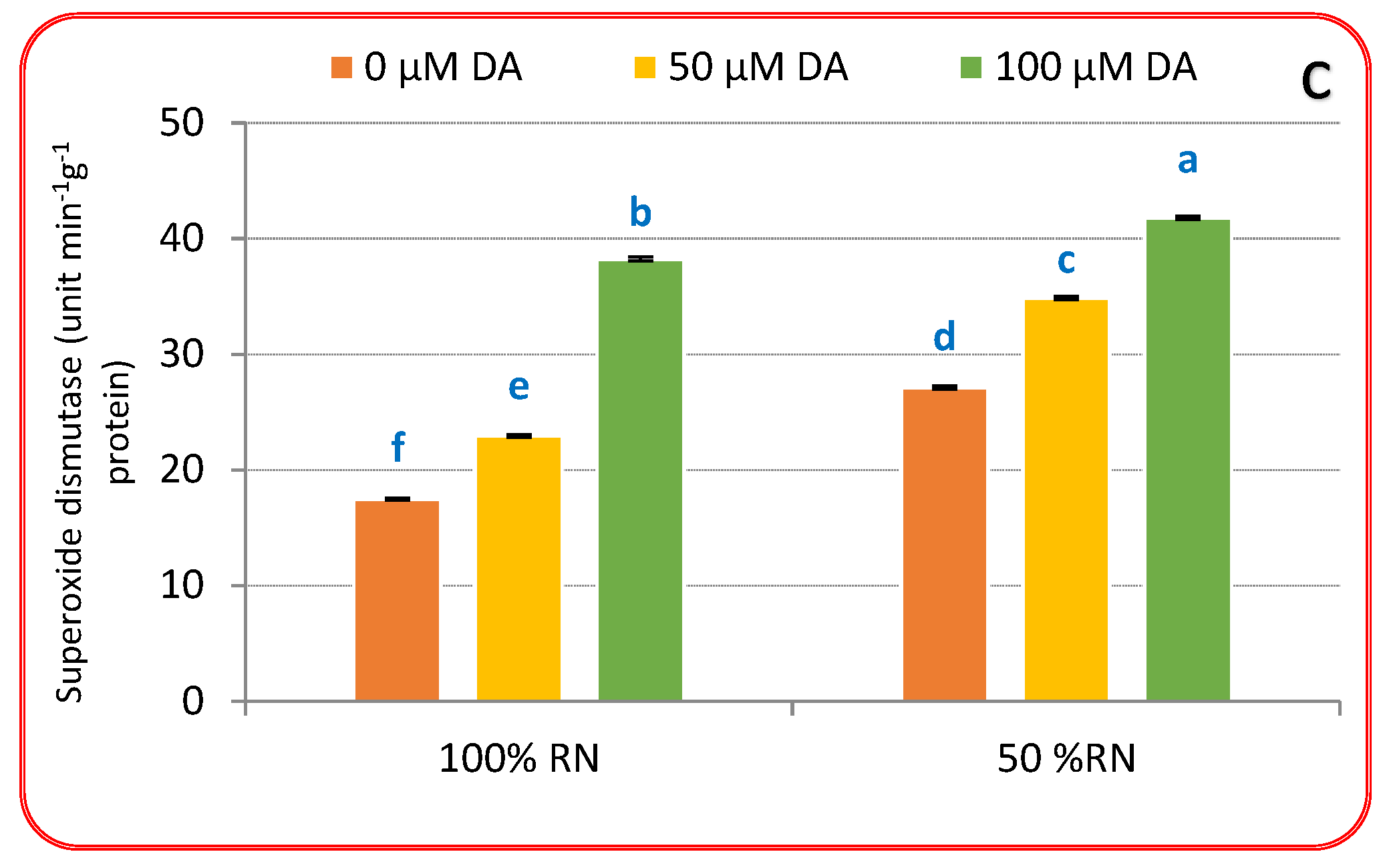
3.4. Antioxidants Enzymes
3.5. Head Quality
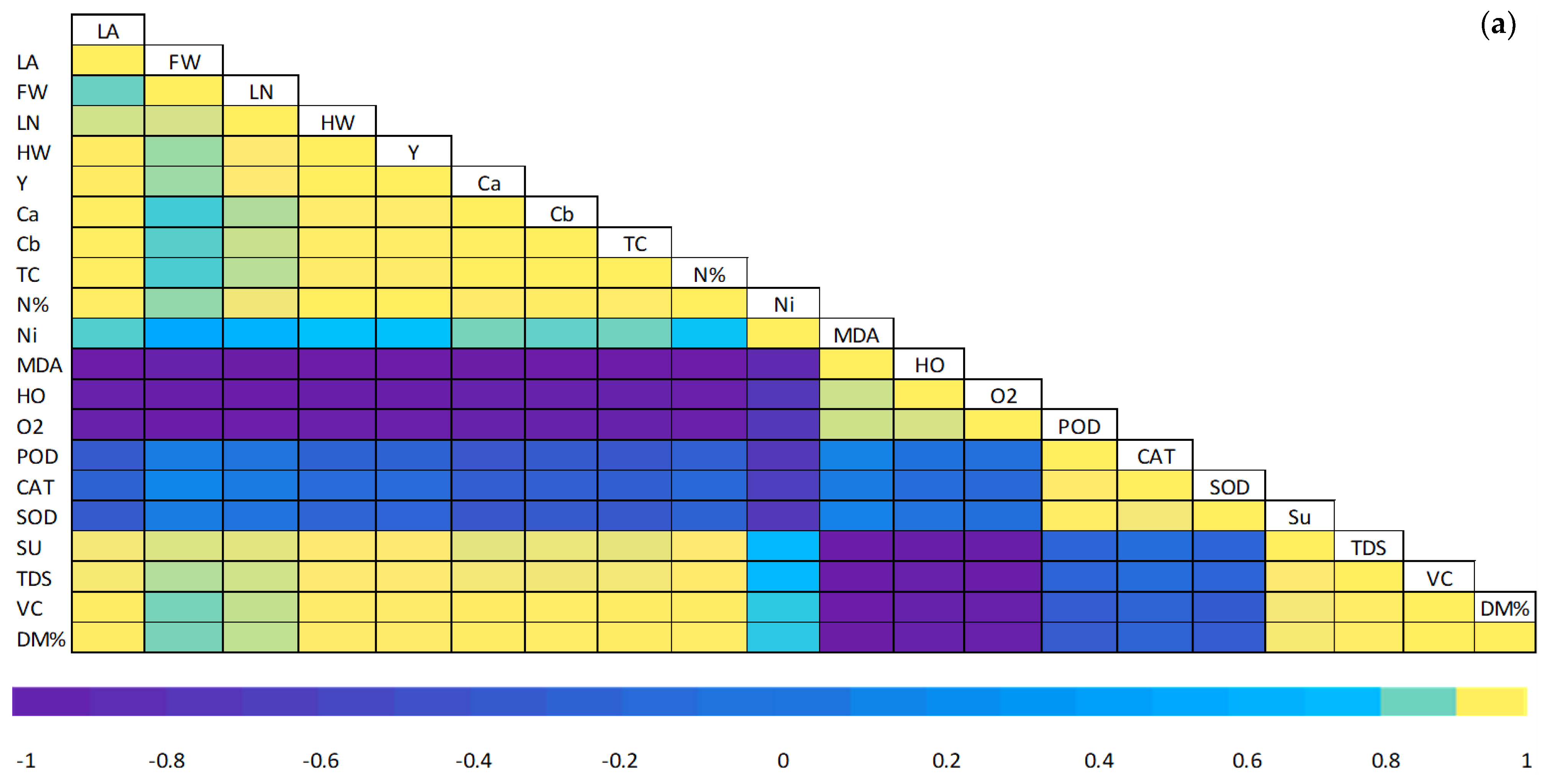
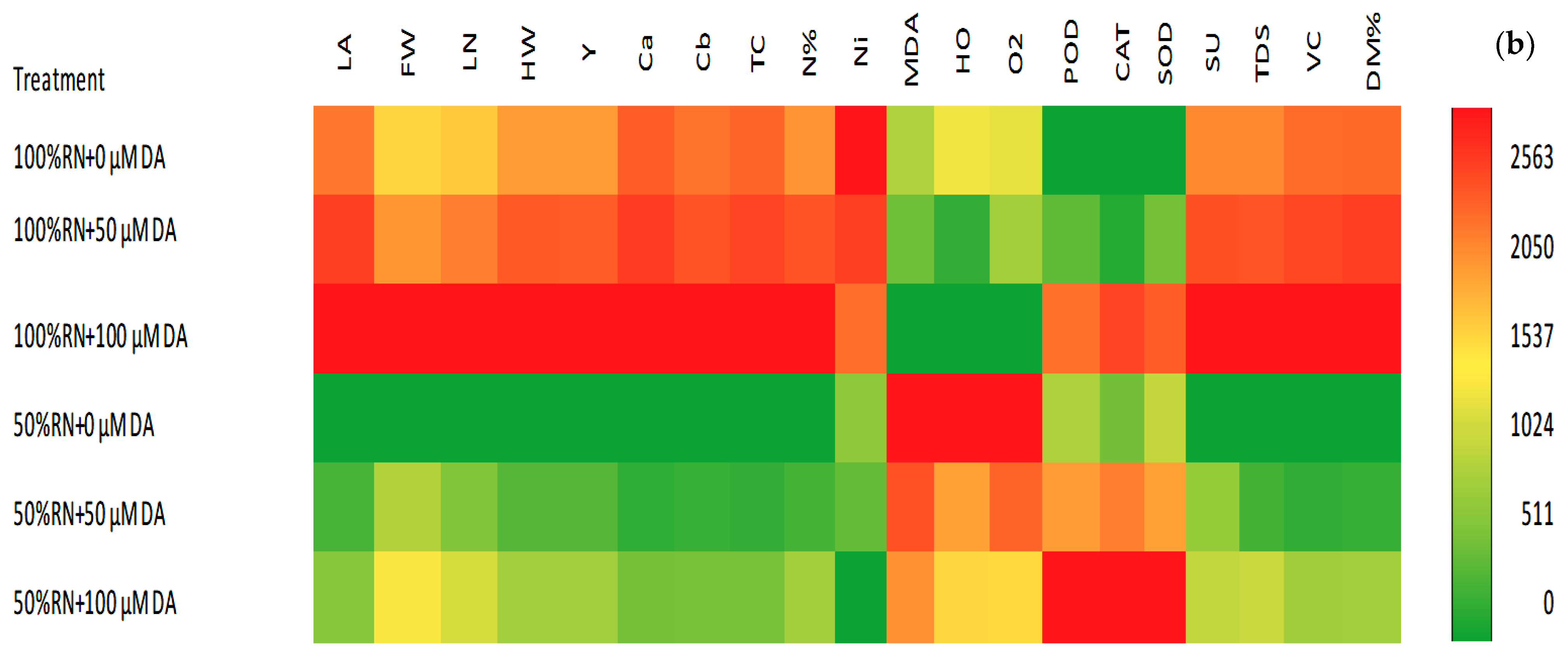
3.6. Pearson Correlations and a Heat Map
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsieh, P.H.; Kan, C.C.; Wu, H.Y.; Yang, H.C.; Hsieh, M.H. Early molecular events associated with nitrogen deficiency in rice seedling roots. Sci. Rep. 2018, 8, 12207. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Li, Q.; Zeng, X.P.; Liu, Y.; Li, Y.R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Yin, B.; Cao, Y.; Han, R.; Ji, J.; He, X.; Liang, B.; Xu, J. Beneficial effects of exogenous melatonin and dopamine on low nitrate stress in Malus hupehensis. Front. Plant Sci. 2022, 12, 807472. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.; Saliba-Colombani, V.; Loudet, O.; Belluomo, P.; Moreau, L.; DanielVedele, F.; Morot-Gaudry, J.F.; Masclaux-Daubresse, C. Leaf yellowing and anthocyanin accumulation are two genetically independent strategies in response to nitrogen limitation in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 74–83. [Google Scholar] [CrossRef]
- Liu, X.-M.; Gao, T.-T.; Zhang, Z.-J.; Tan, K.-X.; Jin, Y.-B.; Zhao, Y.-J.; Ma, F.-W.; Li, C. The mitigation effects of exogenous dopamine on low nitrogen stress in Malus hupehensis. J. Integr. Agric. 2020, 19, 2709–2724. [Google Scholar] [CrossRef]
- Huang, X.; Wei, L.; Xia, Y.; Wang, Y.; Feng, D.; Zhang, H.; Sun, X. Growth and physiological changes of Juglans regia L. seedlings under nitrogen deficiency stress. Pol. J. Environ. Stud. 2023, 32, 567–577. [Google Scholar] [CrossRef]
- Zakari, S.A.; Mau, A.; Han, Z.; Zhao, Q.; Cheng, F. Relationship of nitrogen defciency-induced leaf senescence with ROS generation and ABA concentration in rice fag leaves. J. Plant Growth Regul. 2020, 39, 1503–1517. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef]
- Farouk, S.; Omar, M.M. Sweet basil growth, physiological and ultrastructural modification and oxidative defense system under water deficit and silicon forms treatment. J. Plant Growth Regul. 2020, 39, 1307–1331. [Google Scholar] [CrossRef]
- Farouk, S.; AL-Huqail, A.A. Sustainable biochar and/or melatonin improve salinity tolerance in borage plants by modulating osmotic adjustment, antioxidants, and ion homeostasis. Plants 2022, 11, 765. [Google Scholar] [CrossRef]
- Farouk, S.; AL-Huqail, A.A.; El-Gamal, S.M.A. Potential role of biochar and silicon in improving physio-biochemical and yield characteristics of borage plants under different irrigation regimes. Plants 2023, 12, 1605. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Gao, T.; Liu, W.; Liu, Y.; Zhao, Y.; Liu, Y.; Li, W.; Ding, K.; Ma, F.; Li, C. Functions of dopamine in plants: A review. Plant Signal. Behav. 2020, 5, 1827782. [Google Scholar] [CrossRef] [PubMed]
- Kulma, A.; Szopa, J. Catecholamines are active compounds in plants. Plant Sci. 2007, 172, 433–440. [Google Scholar] [CrossRef]
- Jiao, C.; Lan, G.; Sun, Y.; Wang, G.; Sun, Y. Dopamine alleviates chilling stress in watermelon seedlings via modulation of proline content, antioxidant enzyme activity, and polyamine metabolism. J. Plant Growth Regul. 2021, 40, 277–292. [Google Scholar] [CrossRef]
- Lan, G.; Jiao, C.; Wang, G.; Sun, Y.; Sun, Y. Effects of dopamine on growth, carbon metabolism, and nitrogen metabolism in cucumber under nitrate stress. Sci. Hortic. 2020, 260, 108790. [Google Scholar] [CrossRef]
- Wang, W.; Yang, Y.; Ma, X.; He, Y.; Ren, Q.; Huang, Y.; Wang, J.; Xue, Y.; Yang, R.; Guo, Y.; et al. New insight into the function of dopamine (DA) during Cd stress in duckweed (Lemna turionifera). Plants 2023, 12, 1996. [Google Scholar] [CrossRef]
- Shi, M.; Gu, J.; Wu, H.; Rauf, A.; Emran, T.B.; Khan, Z.; Mitra, S.; Aljohani, A.S.M.; Alhumaydhi, F.A.; Al-Awthan, Y.S.; et al. Phytochemicals, nutrition, metabolism, bioavailability, and health benefits in lettuce-A comprehensive review. Antioxidants 2022, 11, 1158. [Google Scholar] [CrossRef]
- Medina-Lozano, I.; Bertolín, J.R.; Díaz, A. Nutritional value of commercial and traditional lettuce (Lactuca sativa L.) and wild relatives: Vitamin C and anthocyanin content. Food Chem. 2021, 359, 129864. [Google Scholar] [CrossRef]
- Yang, X.; Gil, M.I.; Yang, Q.; Tomás-Barberán, F.A. Bioactive compounds in lettuce: Highlighting the benefits to human health and impacts of preharvest and postharvest practices. Compr. Rev. Food Sci. Food Saf. 2022, 21, 4–45. [Google Scholar] [CrossRef]
- Lafarga, T.; Villaró, S.; Rivera, A.; Bobo, G.; Aguiló-Aguayo, I. Bioaccessibility of polyphenols and antioxidant capacity of fresh or minimally processed modern or traditional lettuce (Lactuca sativa L.) varieties. J. Food Sci. Technol. 2020, 57, 754–763. [Google Scholar] [CrossRef]
- Sepehri, N.Z.; Parvizi, M.M.; Habibzadeh, S.; Handjani, F. Lettuce as an effective remedy in uremic pruritus: Review of the literature supplemented by an in silico study. Evid. Based Complement. Altern. Med. 2022, 2022, 4231854. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Sun, Y.; Hou, W.; Zhao, W.; Wang, P.; Zhao, S.; Zhao, X.; Wang, D. Infection behavior of Listeria monocytogenes on iceberg lettuce (Lactuca sativa var. capitata L). Food Res. Int. 2023, 165, 112487. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, G.; Santos, J.; Freire, M.S.; Antorrena, G.; González-Álvarez, J. Extraction of antioxidants from eucalyptus (Eucalyptus globulus) bark. Wood Sci. Technol. 2012, 46, 443–457. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. “Methods of Soil Analysis”, Part 3: Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Koller, H.R.C. Leaf area-leaf weight relationship in soybean canopy. Crop Sci. 1972, 12, 216–220. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Motsara, M.R.; Roy, R.N. Guide to Laboratory Establishment for Plant Nutrient Analysis, FAO Fertilizer and Plant Nutrition Bulletin; Food and Agriculture Organization: Rome, Italy, 2008; 219p. [Google Scholar]
- Singh, J.P. A rapid method for determination of nitrate in soil and plant extracts. Plant Soil 1988, 110, 137–139. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Tariq, A.; Masroor, M.; Khan, A.; Jaime, A.; da Teixeira, S.; Mohd, I.; Naeem, M. Role of salicylic acid in promoting salt stress tolerance and enhanced artemisinin production in Artemisia annua L. J. Plant Growth Regul. 2011, 30, 425–435. [Google Scholar] [CrossRef]
- Mohammadi, M.; Karr, A.L. Superoxide anion generation in effective and ineffective soybean root nodules. J. Plant Physiol. 2001, 158, 1023–1029. [Google Scholar] [CrossRef]
- Elavarthi, S.; Martin, B. Spectrophotometric assays for antioxidant enzymes in plants. In Plant Stress Tolerance, Methods and Protocols; Ramanjulu, S., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 273–280. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists, Inc.: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Liang, B.; Li, C.; Ma, C.; Wei, Z.; Wang, Q.; Huang, D.; Chen, Q.; Li, C.; Ma, F. Dopamine alleviates nutrient defciency-induced stress in Malus hupehensis. Plant Physiol. Biochem. 2017, 119, 346. [Google Scholar] [CrossRef]
- Taiz, L.; Moller, I.M.; Murphy, A.; Zeiger, E. Plant Physiology and Development; Oxford University Press: New York, NY, USA, 2022; p. 864. [Google Scholar]
- Wen, B.; Li, C.; Fua, X.; Li, D.; Li, L.; Chen, X.; Wu, H.; Cui, X.; Zhang, X.; Shen, H.; et al. Effects of nitrate deficiency on nitrate assimilation and chlorophyll synthesis of detached apple leaves. Plant Physiol. Biochem. 2019, 142, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Halpern, M.; Yermiyahu, U.; Bar-Tal, A.; Gendler, T.; Rachmilevitch, S. CO2 and nitrogen interaction alters root anatomy, morphology, nitrogen partitioning and photosynthetic acclimation of tomato plants. Planta 2019, 250, 14231432. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Li, Y.; Zhang, X.; Liu, C.; Liang, W.; Li, C.; Ma, F.; Li, C. Exogenous dopamine application promotes alkali tolerance of apple seedlings. Plants 2019, 8, 580. [Google Scholar] [CrossRef]
- Protacio, C.M.; Dai, Y.R.; Lewis, E.F.; Flores, H.E. Growth stimulation by catecholamines in plant tissue/organ cultures. Plant Physiol. 1992, 98, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Kamisaka, S. Catecholamine stimulation of the gibberellin action that induces lettuce hypocotyl elongation. Plant Cell Physiol. 1979, 20, 1199–1207. [Google Scholar] [CrossRef]
- Chen, M. Chlorophyll modifications and their spectral extension in oxygenic photosynthesis. Annu. Rev. Biochem. 2014, 83, 317–340. [Google Scholar] [CrossRef]
- Ni, J.; Wang, Q.; Shah, F.A.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous melatonin confers cadmium tolerance by counterbalancing the hydrogen peroxide homeostasis in wheat seedlings. Molecules 2018, 23, 799. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, X. Dopamine-induced abiotic stress tolerance in horticultural plants. Sci. Hortic. 2023, 307, 111506. [Google Scholar] [CrossRef]
- Plett, D.; Baumann, U.; Schreiber, A.W.; Holtham, L.; Kalashyan, E.; Toubia, J.; Nau, J.; Beatty, M.; Rafalski, A.; Dhugga, K.S.; et al. Maize maintains growth in response to decreased nitrate supply through a highly dynamic and developmental stage-specific transcriptional response. Plant Biotechnol. J. 2016, 14, 342–353. [Google Scholar] [CrossRef]
- Balotf, S.; Kavoosi, G.K.; Kholdebarin, B. Nitrate reductase, nitrite reductase, glutamine synthetase, and glutamate synthase expression and activity in response to different nitrogen sources in nitrogen-starved wheat seedlings. Biotechnol. Appl. Biochem. 2016, 63, 220–229. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.K.; Kumar, P.; Sharma, P.N. Oxidative stress and antioxidant responses in young leaves of mulberry plants grown under nitrogen, phosphorus or potassium deficiency. J. Integr. Plant Biol. 2007, 49, 313322. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen alleviates salinity toxicity in Solanum lycopersicum seedlings by regulating ROS homeostasis. Plant Physiol. Biochem. 2019, 141, 466476. [Google Scholar] [CrossRef] [PubMed]
- Macri, F.; Braidot, E.; Petrussa, E.; Vianello, A. Lipoxygenase activity associated to isolated soybean plasma membranes. Biochim. Biophys. Acta 1994, 17, 109–114. [Google Scholar] [CrossRef]
- Jodko-Piórecka, K.; Sikora, B.; Kluzek, M.; Przybylski, P.; Litwinienko, G. Antiradical activity of dopamine, L-DOPA, adrenaline, and noradrenaline in water/methanol and in liposomal systems. J. Org. Chem. 2022, 87, 1791–1804. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef]
- Sohag, A.A.; Tahijb-Ul-Arif, M.D.; Brestic, M.; Afrin, S.; Sakil, M.D.A.; Hossain, T.M.D.; Hassain, M.A.; Hossain, A.M.D. Exogenous salicylic acid and hydrogen peroxide attenuate drought stress in rice. Plant Soil Environ. 2020, 66, 7–13. [Google Scholar] [CrossRef]
- Antunes, F.; Nunes, C.S.; Laranjinha, J.; Cadenas, E. Redox interactions of nitric oxide with dopamine and its derivatives. Toxicology 2005, 208, 207–212. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Parameters | Electric Conductivity (EC) | pH | Organic Matter (OM) |
| 3.49 (dSm−1) | 8.00 | 1.39 (%) | |
| Available nutrients (mg kg−1) | Nitrogen | Phosphorus | Potassium |
| 48.0 | 7.99 | 235.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farouk, S.; El-Hady, M.A.M.A.; El-Sherpiny, M.A.; Hassan, M.M.; Alamer, K.H.; Al-Robai, S.A.; Ali, E.F.; El-Bauome, H.A. Effect of Dopamine on Growth, Some Biochemical Attributes, and the Yield of Crisphead Lettuce under Nitrogen Deficiency. Horticulturae 2023, 9, 945. https://doi.org/10.3390/horticulturae9080945
Farouk S, El-Hady MAMA, El-Sherpiny MA, Hassan MM, Alamer KH, Al-Robai SA, Ali EF, El-Bauome HA. Effect of Dopamine on Growth, Some Biochemical Attributes, and the Yield of Crisphead Lettuce under Nitrogen Deficiency. Horticulturae. 2023; 9(8):945. https://doi.org/10.3390/horticulturae9080945
Chicago/Turabian StyleFarouk, Saad, Mahmoud A. M. Abd El-Hady, Mohamed A. El-Sherpiny, Mohamed M. Hassan, Khalid H. Alamer, Sami Asir Al-Robai, Esmat F. Ali, and Hemat A. El-Bauome. 2023. "Effect of Dopamine on Growth, Some Biochemical Attributes, and the Yield of Crisphead Lettuce under Nitrogen Deficiency" Horticulturae 9, no. 8: 945. https://doi.org/10.3390/horticulturae9080945
APA StyleFarouk, S., El-Hady, M. A. M. A., El-Sherpiny, M. A., Hassan, M. M., Alamer, K. H., Al-Robai, S. A., Ali, E. F., & El-Bauome, H. A. (2023). Effect of Dopamine on Growth, Some Biochemical Attributes, and the Yield of Crisphead Lettuce under Nitrogen Deficiency. Horticulturae, 9(8), 945. https://doi.org/10.3390/horticulturae9080945