
Plant Production with Microalgal Biostimulants



Abstract
1. Introduction
2. Crop Production and Quality Can Be Improved with the Use of Microalgae and Algal–Bacterial Consortia
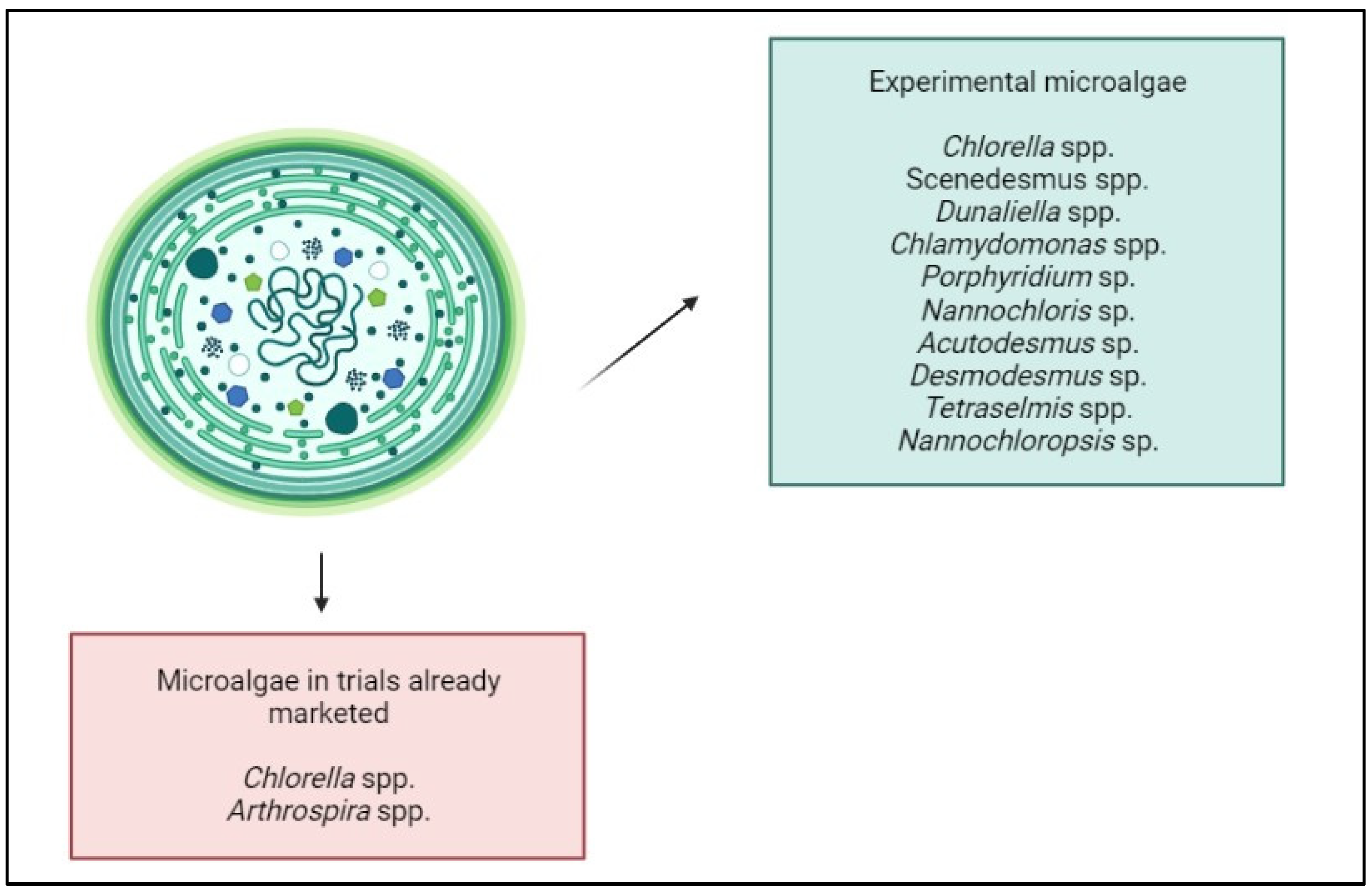
3. Microalgal Biomass Production
4. Microalgae as a New Source of Biostimulants
5. Processes and Applications of Biostimulating Algal Biomass
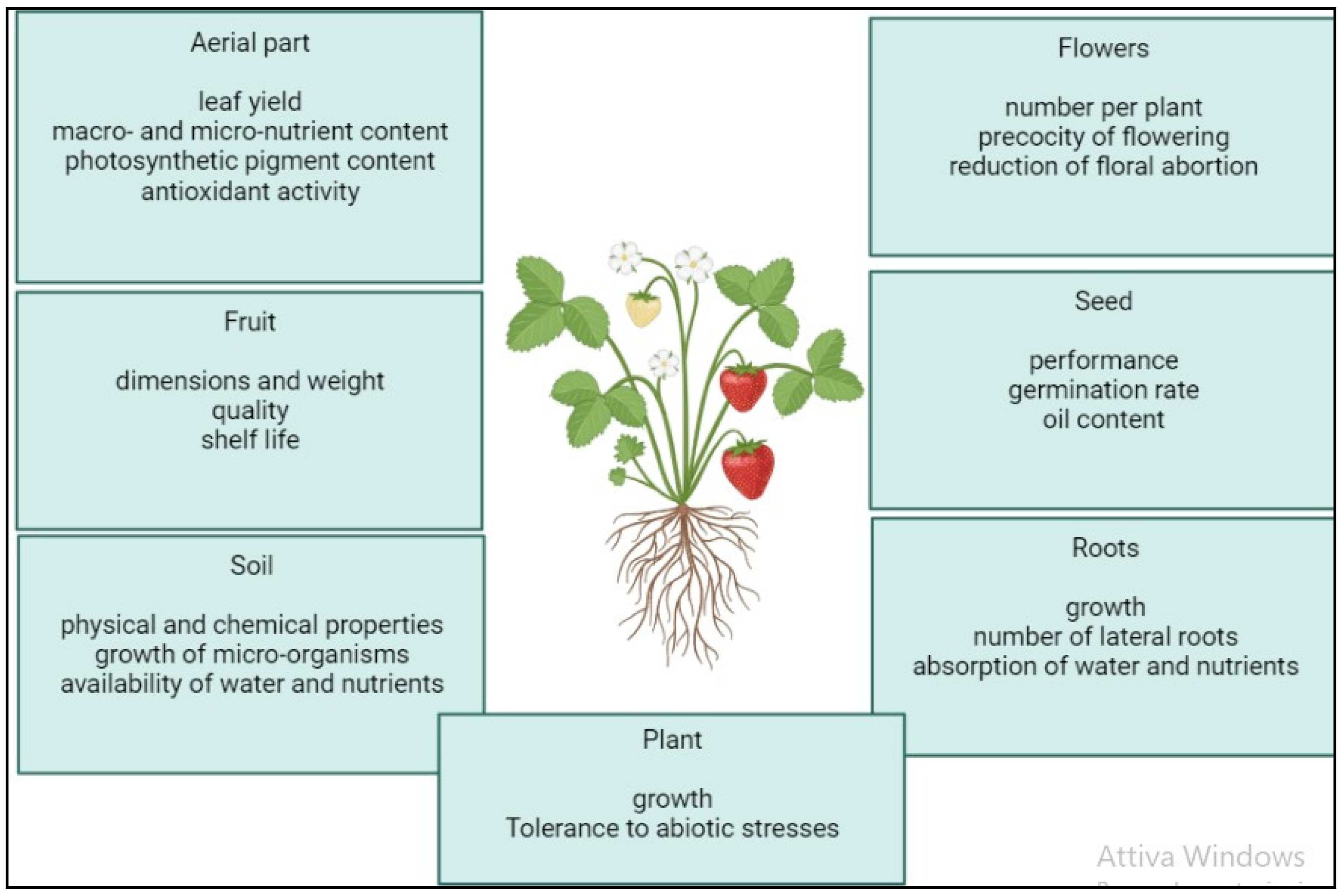
6. Main Biostimulating Effects of Microalgae on Plants
7. Use of Microalgal Biostimulants as a Contribution to Sustainable Agricultural Practices
8. Advantages and Critical Issues in the Use of Microalgae for Biostimulants
9. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable sources of plant biostimulation: Microalgae as a sustainable means to improve crop performance. Front. Plant Sci. 2018, 9, 1782. [Google Scholar] [CrossRef] [PubMed]
- Dmytryk, A.; Chojnacka, K. Algae as fertilizers, biostimulants, and regulators of plant growth. In Algae Biomass: Characteristics and Applications; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 115–122. [Google Scholar]
- Mzibra, A.; Aasfar, A.; Benhima, R.; Khouloud, M.; Boulif, R.; Douira, A.; Bamouh, A.; Meftah Kadmiri, I. Biostimulants derived from moroccan seaweeds: Seed germination metabolomics and growth promotion of tomato plant. J. Plant Growth Regul. 2020, 40, 353–370. [Google Scholar] [CrossRef]
- Povero, G.; Mejia, J.F.; di Tommaso, D.; Piaggesi, A.; Warrior, P. A systematic approach to discover and characterize natural plant biostimulants. Front. Plant Sci. 2016, 7, 435. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from red seaweeds as promoters of growth and elicitors of defense response in plants. Front. Mar. Sci. 2016, 3, 81. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Win, T.T.; Barone, G.D.; Secundo, F.; Fu, P. Algal biofertilizers and plant growth stimulants for sustainable agriculture. Ind. Biotechnol. 2018, 14, 203–211. [Google Scholar] [CrossRef]
- Arnau, L.; Richard, H. Techno-Economic Feasibility Study for the Production of Microalgae Based Plant Biostimulant; KTH Royal Institute of Technology: Stockholm, Sweden, 2016; pp. 300–365. [Google Scholar]
- Barone, V.; Baglieri, A.; Stevanato, P.; Broccanello, C.; Bertoldo, G.; Bertaggia, M.; Cagnin, M.; Pizzeghello, D.; Moliterni, V.M.C.; Mandolino, G.; et al. Root morphological and molecular responses induced by microalgae extracts in sugar beet (Beta vulgaris L.). J. Appl. Phycol. 2018, 30, 1061–1071. [Google Scholar] [CrossRef]
- Kopta, T.; Pavlíkova, M.; Sękara, A.; Pokluda, R.; Maršálek, B. Effect of bacterial-algal biostimulant on the yield and internal quality of Lettuce (Lactuca sativa L.) produced for spring and summer crop. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 615–621. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress—A review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Chanda, M.J.; Merghoub, N.; El Arroussi, H. Microalgae polysaccharides: The new sustainable bioactive products for the development of plant bio-stimulants? World J. Microbiol. Biotechnol. 2019, 35, 411–442. [Google Scholar] [CrossRef]
- Chakdar, H.; Jadhav, S.D.; Dhar, D.W.; Pabbi, S. Potential applications of blue green algae. J. Sci. Ind. Res. 2012, 71, 13–20. [Google Scholar]
- Dasgan, H.Y.; Aydoner, G.; Akyol, M. Use of some microorganisms as bio-fertilizers in soilless grown squash for saving chemical nutrients. Acta Hortic. Int. Soc. Hortic. Sci. 2012, 927, 155–162. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, J.; Sommerfeld, M. Biofertilizer and biostimulant properties of the microalga Acutodesmus dimorphus. J. Appl. Phycol. 2016, 28, 1051–1061. [Google Scholar] [CrossRef]
- Renuka, N.; Prasanna, R.; Sood, A.; Bansal, R.; Bidyarani, N.; Singh, R.; Shivay, Y.S.; Nain, L.; Ahluwalia, A.S. Wastewater grown microalgal biomass as inoculants for improving micronutrient availability in wheat. Rhizosphere 2017, 3, 150–159. [Google Scholar] [CrossRef]
- Stiles, W.A.V.; Styles, D.; Chapman, S.P.; Esteves, S.; Bywater, A.; Melville, L.; Silkina, A.; Lupatsch, I.; Grünewald, C.F.; Lovitt, R. Using microalgae in the circular economy to valorise anaerobic digestate: Challenges and opportunities. Bioresour. Technol. 2018, 267, 732–742. [Google Scholar] [CrossRef]
- Rumin, J.; Nicolau, E.; Gonçalves de Oliveira, R., Jr.; Fuentes-Grünewald, C.; Flynn, K.J.; Picot, L. European commission. Mar. Drugs 2020, 18, 79. [Google Scholar] [CrossRef]
- La Torre, A.; Battaglia, V.; Caradonia, F. An overview of the current plant biostimulant legislations in different European Member States. J. Sci. Food Agric. 2016, 96, 727–734. [Google Scholar] [CrossRef]
- Coppens, J.; Grunert, O.; van den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; de Gelder, L. The use of microalgae as a high-value organic slow-release fertiliser results in tomatoes with increased carotenoid and sugar levels. J. Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]
- Mulbry, W.; Kondrad, S.; Pisarro, C. Biofertilisers from algal treatment of dairy and swine manure effluents. J. Veg. Sci. 2007, 12, 107–125. [Google Scholar]
- Shaaban, M.M.; El-Fouly, M.M.; Abdel-Maguid, A.A. Zinc-Boron relationship in wheat plants grown under low or high levels of calcium carbonate in the soil. Pak. J. Biol. Sci. 2004, 7, 633–639. [Google Scholar]
- Shaaban, M.M.; Loehnertz, O.; El-Fouly, M.M. Grapevine genotypic tolerance to lime and possibility of chlorosis recovery through micronutrients foliar application. Int. J. Bot. 2007, 3, 179–187. [Google Scholar] [CrossRef]
- Shaaban, M.M.; Hussein, M.M.; El Saady, A.M. Nutritional status in shoots of barley genotypes as affected by salinity of irrigation water. Am. J. Plant Physiol. 2008, 3, 89–95. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: Boston, MA, USA, 1995. [Google Scholar]
- El-Fouly, M.M.; Shaaban, M.M.; El-Khdraa, T.F. Soil and plant nutritional status in fruit orchards in Syria. Acta Agron. Hung. 2008, 56, 363–370. [Google Scholar] [CrossRef]
- Shaaban, M.M. Nutritional status and growth of maise plants as affected by green microalgae as soil additives. J. Biol. Sci. 2001, 6, 475–479. [Google Scholar] [CrossRef]
- Shaaban, M.M. Green microalgae water extract as foliar feeding to wheat plants. Pak. J. Biol. Sci. 2001, 4, 628–632. [Google Scholar]
- Faheed, F.A.; Fattah, Z.A. Effect of Chlorella vulgaris as Biofertiliser on Growth Parameters and Metabolic Aspects of Lettuce Plant. J. Agric. Soc. Sci. 2008, 4, 165–169. [Google Scholar]
- Tarraf, S.A.; Talaat, I.M.; El-Sayed, A.E.K.B.; Balbaa, L.K. Influence of foliar application of algae extract and amino acids mixture on fenugreek plants in sandy and clay soils. Amino Acids 2015, 16, 19–58. [Google Scholar] [CrossRef]
- Paudel, Y.P.; Pradhan, S.; Pant, B.; Prasad, B.N. Role of blue green algae in rice productivity. Agric. Biol. J. N. Am. 2012, 3, 332–335. [Google Scholar] [CrossRef]
- Shalaby, T.A.; El-Ramady, H. Effect of foliar application of bio-stimulants on growth, yield, components, and storability of garlic (Allium sativum L.). Aust. J. Crop Sci. 2014, 8, 271. [Google Scholar]
- Gonzalez, L.E.; Bashan, Y. Increased growth of the microalga chlorella vulgariswhen coimmobilized and cocultured in alginate beads with the plant-growth-promoting bacterium Azospirillum brasilense. Appl. Environ. Microbiol. 2000, 66, 1527–1531. [Google Scholar] [CrossRef]
- Babu, N.R.; Devaraj, V.R. High temperature and salt stress response in French bean (Phaseolus vulgaris). Aust. J. Crop Sci. 2008, 2, 40–48. [Google Scholar]
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; Bux, F. Microalgae as multi-functional options in modern agriculture: Current trends, prospects and challenges. Biotechnol. Adv. 2018, 36, 1255–1273. [Google Scholar] [CrossRef]
- Xue, C.; Wang, L.; Wu, T.; Zhang, S.; Tang, T.; Wang, L.; Zhao, Q.; Sun, Y. Characterization of co-cultivation of cyanobacteria on growth, productions of polysaccharides and extracellular proteins, nitrogenase activity, and photosynthetic activity. Appl. Biochem. Biotechnol. 2017, 181, 340–349. [Google Scholar] [CrossRef]
- Padmaperuma, G.; Kapoore, R.V.; Gilmour, D.J.; Vaidyanathan, S. Microbial consortia: A critical look at microalgae co-cultures for enhanced biomanufacturing. Crit. Rev. Biotechnol. 2018, 38, 690–703. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Hultberg, M.; Carlsson, A.S.; Gustafsson, S. Treatment of drainage solution from hydroponic greenhouse production with microalgae. Bioresour. Technol. 2013, 136, 401–406. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Meireles, L.A.; Malcata, X. Microalgal reactors: A review of enclosed system designs and performances. Biotechnol. Prog. 2006, 22, 1490–1506. [Google Scholar] [CrossRef]
- Ugwu, C.U.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef]
- Moheimani, N.R.; Borowitzka, M.A. The long-term culture of the coccolithophore Pleurochrysis carterae (Haptophyta) in outdoor raceway ponds. J. Appl. Phycol. 2006, 18, 703–712. [Google Scholar] [CrossRef]
- Pushparaj, B.; Pelosi, E.; Tredici, M.R.; Pinzani, E.; Materassi, R. As integrated culture system for outdoor production of microalgae and cyanobacteria. J. Appl. Phycol. 1997, 9, 113–119. [Google Scholar] [CrossRef]
- Lee, Y.K. Microalgal mass culture systems and methods: Their limitation and potential. J. Appl. Phycol. 2001, 13, 307–315. [Google Scholar] [CrossRef]
- Gong, Y.; Jiang, M. Biodiesel production with microalgae as feedstock: From strains to biodiesel. Biotechnol. Lett. 2011, 33, 1269–1284. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Zhou, Q. Co-cultivation of Chlorella spp. and tomato in a hydroponic system. Biomass Bioenerg. 2017, 97, 132–138. [Google Scholar] [CrossRef]
- Barone, V.; Puglisi, I.; Fragalà, F.; Piero, A.R.L.; Giuffrida, F.; Baglieri, A. Novel bioprocess for the cultivation of microalgae in hydroponic growing system of tomato plants. J. Appl. Phycol. 2019, 31, 465–470. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W.; Volkman, J.K.; Dunstan, G.A. Nutritional properties of microalgae for mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Kaplan, D.; Richmond, A.E.; Dubinsky, Z.; Aaronson, S. Algal nutrition. In Handbook of Microalgal Mass Culture; Richmond, A., Ed.; CRC Press: Boca Raton, FL, USA, 1986; pp. 147–198. [Google Scholar]
- Levasseur, M.; Thompson, P.A.; Harrison, P.J. Physiological acclimation of marine phytoplankton to different nitrogen sources†. J. Phycol. 1993, 29, 587–595. [Google Scholar] [CrossRef]
- Danesi, E.D.G.; Rangel-Yagui, C.O.; de Carvalho, J.C.M.; Sato, S. An investigation of effect of replacing nitrate by urea in the growth and production of chlorophyll by Spirulina platensis. Biomass Bioenerg. 2002, 23, 261–269. [Google Scholar] [CrossRef]
- Colla, L.M.; Reinehr, C.O.; Reichert, C.; Costa, J.A.V. Production of biomass and nutraceutical compounds by Spirulina platensis under different temperature and nitrogen regimes. Bioresour. Technol. 2007, 98, 1489–1493. [Google Scholar] [CrossRef]
- Czerpak, R.; Bajguz, A.; Piotrowska, A.; Dobrogowska, R.; Matejczyk, W.; Wieslawski, W. Biochemical activity of di- and polyamines in the green alga Chlorella vulgaris Beijerinck (Chlorophyceae). Acta Soc. Bot. Pol. 2003, 72, 19–24. [Google Scholar] [CrossRef]
- Hourmant, A.; Mereau, N.; Penot, M.; Cann, C.; Caroff, J. Influence of polyamines on growth and metabolism of Dunaliella primolecta. Acta Bot. Neerl. 1994, 43, 129–136. [Google Scholar] [CrossRef]
- Czerpak, R.; Krotke, A.; Mical, A. Comparison of stimulatory effect of auxins and cytokinins on protein, saccharides and chlorophylls content in Chlorella pyrenoidosa Chick. Pol. Arch. Hydrobiol. 1999, 46, 71–82. [Google Scholar]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing sustainability by improving plant salt tolerance through macro- and microalgal biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal biostimulants and biofertilisers in crop productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Rachidi, F.; Benhima, R.; Sbabou, L.; El Arroussi, H. Microalgae polysaccharides biostiluating effect on tomato plants: Growth and metabolic distribution. Biotechnol. Rep. 2020, 25, e00426. [Google Scholar] [CrossRef]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: A critical review of microalgae as renewable resources for agriculture. Algal Res. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systemes. Italus Hortus 2018, 25, 27–36. [Google Scholar] [CrossRef]
- Puglisi, I.; la Bella, E.; Rovetto, E.I.; lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar]
- Manjunath, M.; Kanchan, A.; Rajan, K.; Venkatachalam, S.; Prasanna, R.; Ramakrishnan, B.; Hossain, F.; Nain, L.; Shivay, Y.S.; Rai, A.B. Beneficial cyanobacteria and eubacteria synergistically enhance bioavailability of soil nutrients and yield of okra. Heliyon 2016, 2, e00066. [Google Scholar] [CrossRef]
- Adessi, A.; de Carvalho, R.C.; de Philippis, R.; Branquinho, C.; da Silva, J.M. Microbial extracellular polymeric substances improve water retention in dryland biological soil crusts. Soil Biol. Biochem. 2018, 116, 67–69. [Google Scholar] [CrossRef]
- Plaza, B.M.; Gomez-Serrano, C.; Acien-Fernandez, F.G.; Jimenez-Becker, S. Effetc of microalgae hydrolysate foliar application (Arthrospira platensis and Scenedesmus sp.) on Petunia x hybrida growth. Environ. Boil. Fishes 2018, 30, 2359–2365. [Google Scholar]
- Navarro, Q.R.; de Oliveira Correa, D.; Behling, A.; Noseda, M.D.; Amano, E.; Mamoru Suzuki, R.; Lopes Fortes Ribas, L. Efficient use of biomass and extract of the microalga Desmodesmus subspicatus (Scenedesmaceae) in asymbotic seed germination and seedling development of the orchid Cattleya warneri. J. Appl. Phycol. 2021, 2, 134–141. [Google Scholar]
- Mutale-Joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; El Arroussi, H. Screening of microalgae liquid extracts for their biostimulant properties on plant growth, nutrient uptake and metabolite profile of Solanum lycopersicum L. Sci. Rep. 2020, 10, 2820. [Google Scholar] [CrossRef]
- Rodrigues, M.; Baptistella, J.L.C.; Horz, D.C.; Bortolato, L.M.; Mazzafera, P. Organic plant biostimulants and fruit quality—A review. Agronomy 2020, 10, 98. [Google Scholar] [CrossRef]
- Shariatmadari, Z.; Riahi, H.; Abdi, M.; Hashtroudi, M.S.; Ghassempour, A.R. Impact of cyanobacterial ectracts on the growth and oil content of the medicinal plant Menthas piperita L. Environ. Boil. Fishes 2015, 27, 2279–2287. [Google Scholar]
- Geries, L.S.M.; Elsadany, A.Y. Maximizing growth and productivity of onion (Allium cepa L.) by Spirulina platensis extract and nitrogen fixing endophyte Pseudomonas stutzeri. Arch. Microbiol. 2021, 203, 169–181. [Google Scholar] [CrossRef]
- Panda, D.; Pramanik, K.; Nayak, B.R. Use of seaweed extracts as plant growth regulators for sustainable agriculture. Int. J. Bioresour. Stress Manag. 2012, 3, 404–411. [Google Scholar]
- De Philippis, R.; Colica, G.; Micheletti, E. Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: Molecular basis and practical applicability of the biosorption process. Appl. Microbial. Biotechnol. 2011, 92, 697–708. [Google Scholar] [CrossRef]
- Arora, M.; Kaushik, A.; Rani, N.; Kaushik, C.P. Effect of cyanobacterial exopolysaccharides on salt stress alleviation and seed germination. J. Environ. Biol. 2010, 31, 701–704. [Google Scholar]
- Seifikalhor, M.; Hassani, S.B.; Aliniaeifard, S. Seed priming by cyanobacteria (Spirulina platensis) and Salep Gum enhances tolerance of Maize plant against Cadmium toxicity. J. Plant Growth Regul. 2019, 39, 1009–1021. [Google Scholar] [CrossRef]
- Godlewska, K.; Michalak, I.; Pacyga, P.; Basladynska, S.; Chojnacka, K. Potential applications of cyanobacteria: Spirulina platensis filtrates and homogenates in agriculture. World J. Microbiol. Biotechnol. 2019, 35, 80. [Google Scholar] [CrossRef]
- Mogor, A.F.; Ordog, V.; Pace Pereira Lima, G.; Molnar, Z.; Mogor, G. Biostimulant properties of cyanobacterial hydrolysate related to polyamines. J. Appl. Phycol. 2018, 30, 453–460. [Google Scholar] [CrossRef]
- Tuhy, L.; Samoraj, M.; Witkowska, Z.; Chojnacka, K. Biofortification of maize with micronutrients by Spirulina. OpenChem 2015, 13, 1119–1126. [Google Scholar] [CrossRef]
- Dias, G.A.; Rocha, R.H.C.; Araujo, J.L.; de Lima, J.F.; Guedes, W.A. Growth, yield, and postharvest quality in eggplant produced under different foliar fertilizer (Spirulina platensis) treatments. Semin. Cienc. Agrar. Londrina 2016, 6, 3893–3902. [Google Scholar] [CrossRef]
- Ertani, A.; Sambo, P.; Nicoletto, C.; Santagata, S.; Schiavon, M.; Nardi, S. The use of organic biostimulants in hot pepper plants to help low input sustainable agriculture. Chem. Biol. Technol. Agric. 2015, 2, 11. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Abo-shady, A.M.; El-Nagar, M.M.F. Cyanobacterial Arthrospira (Spirulina platensis) as safener against harmaful effects of fusilade herbicide on faba bean plant. Rend. Fis. Acc. Lincei. 2016, 27, 455–462. [Google Scholar] [CrossRef]
- Gemin, L.G.; Mogor, A.F.; de Oliveira Amatussi, J.; Mogor, G. Microalgae associated to humic acid as a novel biostimulant improving onion growth and yield. Sci. Horticult. 2019, 256, 108560. [Google Scholar] [CrossRef]
- Roser, M.; Ritchie, H.; Ortiz-Ospina, E. World population growth. Our World Data 2019, 2, 126. [Google Scholar]
- Liu, H. Ammonia synthesis catalyst 100 years: Practice, enlightenment and challenge. Cuihua Xuebao Chin. J. Catal. 2014, 35, 1619–1640. [Google Scholar] [CrossRef]
- Daneshgar, S.; Callegari, A.; Capodoglio, A.; Vaccari, D. The potential phosphorus crisis: Resource conservation and possibile escape technologies: A review. Resources 2018, 7, 37. [Google Scholar] [CrossRef]
- Perin, G.; Yunus, I.S.; Valton, M.; Alobwede, E.; Jones, P.R. Sunlight driven recycling to increase nutrient use efficiency in agriculture. Algal Res. 2019, 41, 101554. [Google Scholar] [CrossRef]
- Schutz, L.; Gattinger, A.; Meier, M.; Muller, A.; Boller, T.; Mader, P.; Mathimaran, N. Improving crop yield and nutrient use efficiency via biofertilization—A global metaanalysis. Front. Plant Sci. 2018, 8, 2204. [Google Scholar] [CrossRef]
- Alobwed, E.; Leak, J.R.; Pandhal, J. Circular economy fertilization: Testing micro and macro algal species as soil improvers and nutrient sources for crop production in greenhouse and field conditions. Geoderma 2019, 7, 49. [Google Scholar] [CrossRef]
- Nisha, R.; Kaushik, A.; Kaushik, C.P. Effect of indigenous cyanobacterial application on structural stability and productivity of an organically poor semi-arid soil. Geoderma 2007, 138, 49–56. [Google Scholar] [CrossRef]
- Stirk, W.A.; Ordog, V.; Novak, O.; Rolcik, J.; Strnad, M.; Balint, P.; van Staden, J. Auxin and cytokinin relationships in 24 microalgal strains. J. Phycol. 2013, 49, 459–467. [Google Scholar] [CrossRef]
- Guzman-Murillo, M.A.; Ascencio, F.; Larrinaga-Mayoral, J.A. Germination and ROS detoxification in bell pepper (Capsicum anuum L.) under NaCl stress and treatment with microalgae extracts. Protoplasma 2013, 250, 33–42. [Google Scholar] [CrossRef]
- Tredici, M.R. Mass production of microalgae: Photobioreactors. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; John Wiley: Hoboken, NJ, USA, 2004; pp. 178–214. [Google Scholar]
- Valderrama, D.; Cai, J.; Hishamunda, N.; Ridler, N.; Neish, I.C.; Hurtado, A.Q.; Msuya, F.E.; Krishnan, M.; Narayanakumar, R.; Kronen, M.; et al. The economics of Kappaphycus seaweed cultivation in developing countries: A comparative analysis of farming systems. Aquac. Econ. Manag. 2015, 19, 251–277. [Google Scholar] [CrossRef]
- Fortser, J.; Radulovich, R. Seaweed and Food Security. Seaweed Sustainability: Food and Non-Food Applications; Elsevier: Amsterdam, The Netherlands, 2015; pp. 289–313. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Genera of Microalgae | Effects | Ref. |
|---|---|---|---|
| Lettuce (Lactuca sativa L.) | Chlorella, Scenedesmus quadricauda, Spirulina platensis | Improved productivity, antioxidant capacity, and carotenoid content and increased dry matter, chlorophyll, and protein in seedlings. | [82] |
| Maize (Zea mays L.) | Spirulina platensis | Increased production of caryopses and micronutrient absorption. | [83] |
| Aubergine (Solanum melongena L.) | Spirulina platensis | Increased vegetative growth and fruit production. | [84] |
| Tomato (Solanum lycopersicum L.) | Acutodesmus dimorphus, Chlorella vulgaris, Scenedesmus quadricauda, Nannochloropsis oculata | Increased seed germination, crop biomass, root development, and dry matter. Increased sugar and carotenoid content in fruit. | [72,85] |
| Pepper (Capsicum annuum L.) | Spirulina platensis, Dunaliella salina | Plant growth stimulation and salt stress mitigation in seed germination. | [86] |
| Cucumber (Cucumis sativus L.) | Spirulina platensis | Improved fresh weight. | [82] |
| Fava (Vicia faba L.) | Spirulina platensis | Improved protein and amino acid levels of roots and sprouts. | [87] |
| Garlic (Allium sativum L.) | Arthrospira fusiformis | Increased plant height. | [88] |
| Onion (Allium cepa L.) | Spirulina platensis, Scenedesmus subspicatus | Increased production, photosynthetic pigments, root development, and sugar and protein content. | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prisa, D.; Spagnuolo, D. Plant Production with Microalgal Biostimulants. Horticulturae 2023, 9, 829. https://doi.org/10.3390/horticulturae9070829
Prisa D, Spagnuolo D. Plant Production with Microalgal Biostimulants. Horticulturae. 2023; 9(7):829. https://doi.org/10.3390/horticulturae9070829
Chicago/Turabian StylePrisa, Domenico, and Damiano Spagnuolo. 2023. "Plant Production with Microalgal Biostimulants" Horticulturae 9, no. 7: 829. https://doi.org/10.3390/horticulturae9070829
APA StylePrisa, D., & Spagnuolo, D. (2023). Plant Production with Microalgal Biostimulants. Horticulturae, 9(7), 829. https://doi.org/10.3390/horticulturae9070829