
Effects of 10 Dwarfing Interstocks on Cold Resistance of ‘Tianhong 2’ Apple

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Test Methods
2.2.1. Materials, Treatment and Sampling
2.2.2. Relative Electrolyte Leakage (REL) Measurement and LT50 Calculation
2.2.3. Determination of Relevant Physiological Indexes
2.2.4. Statistical Analysis
3. Results
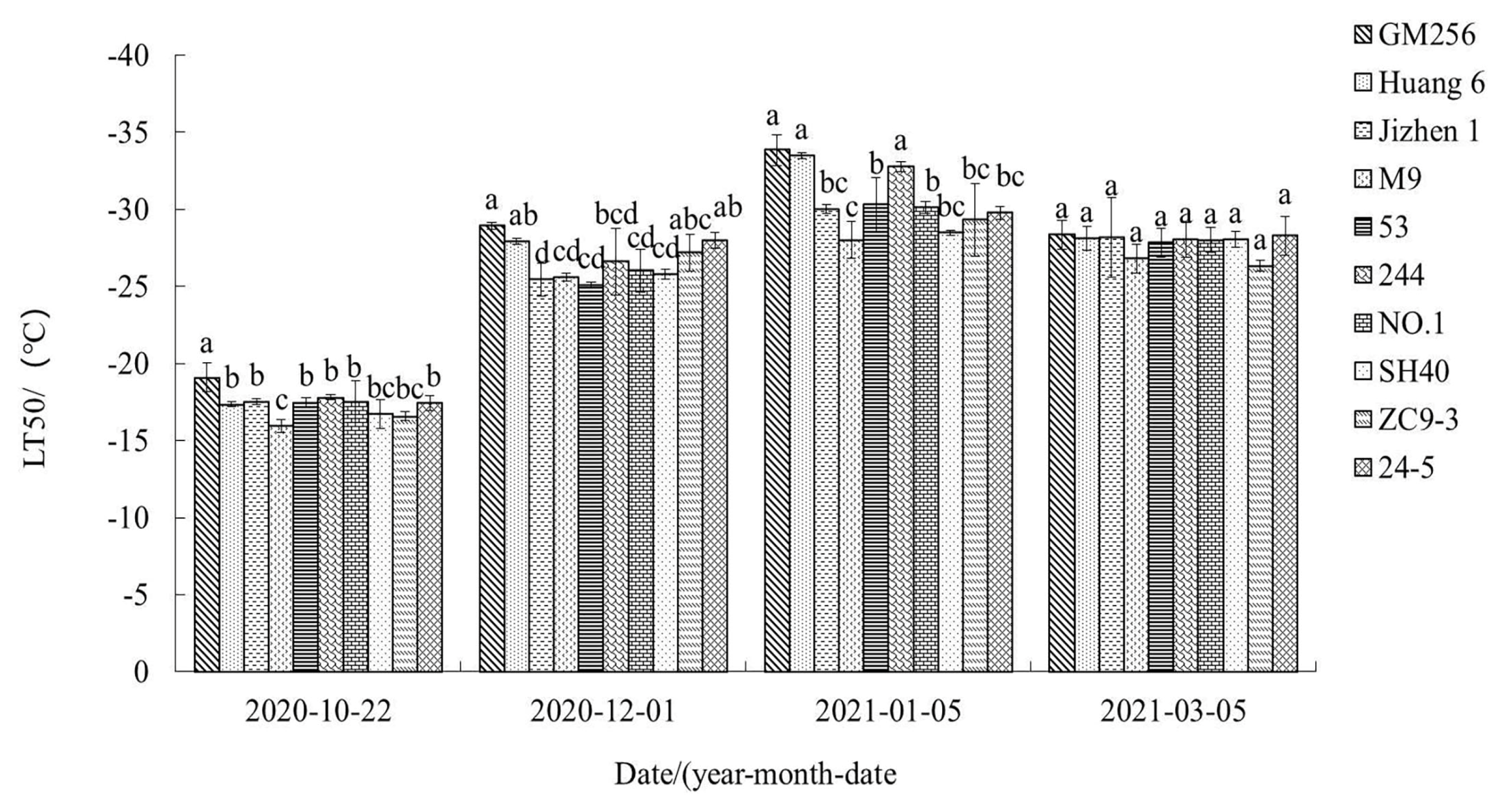
3.1. Changes of LT50 of ‘Tianhong 2’ Apple Branches Grafted on Different Interstocks under Natural Overwintering Process
3.2. Changes of Physiological Indexes of ‘Tianhong 2’ Apple Branches Grafted on Different Interstocks under Natural Overwintering Process
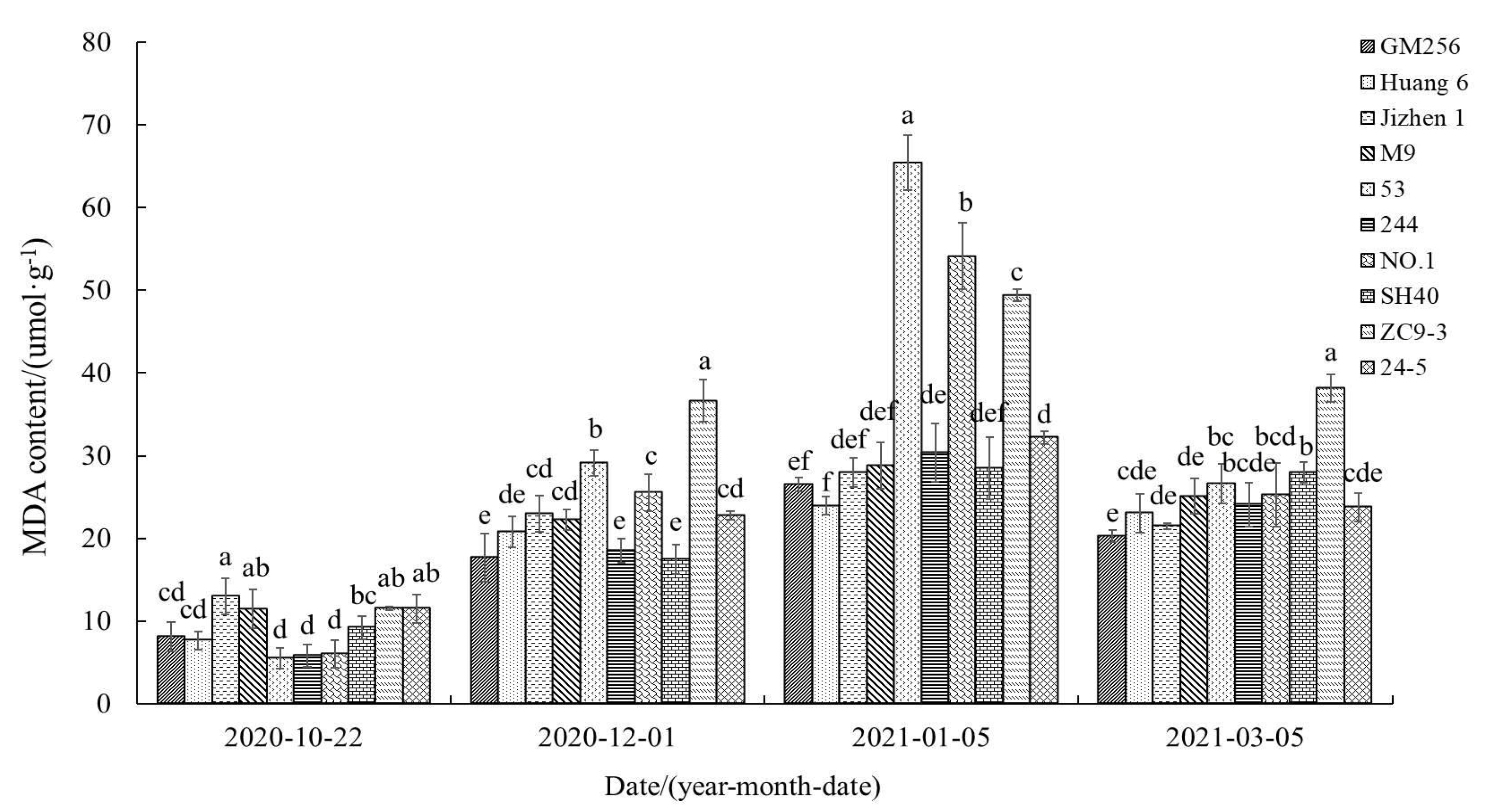
3.2.1. Change of MDA Content
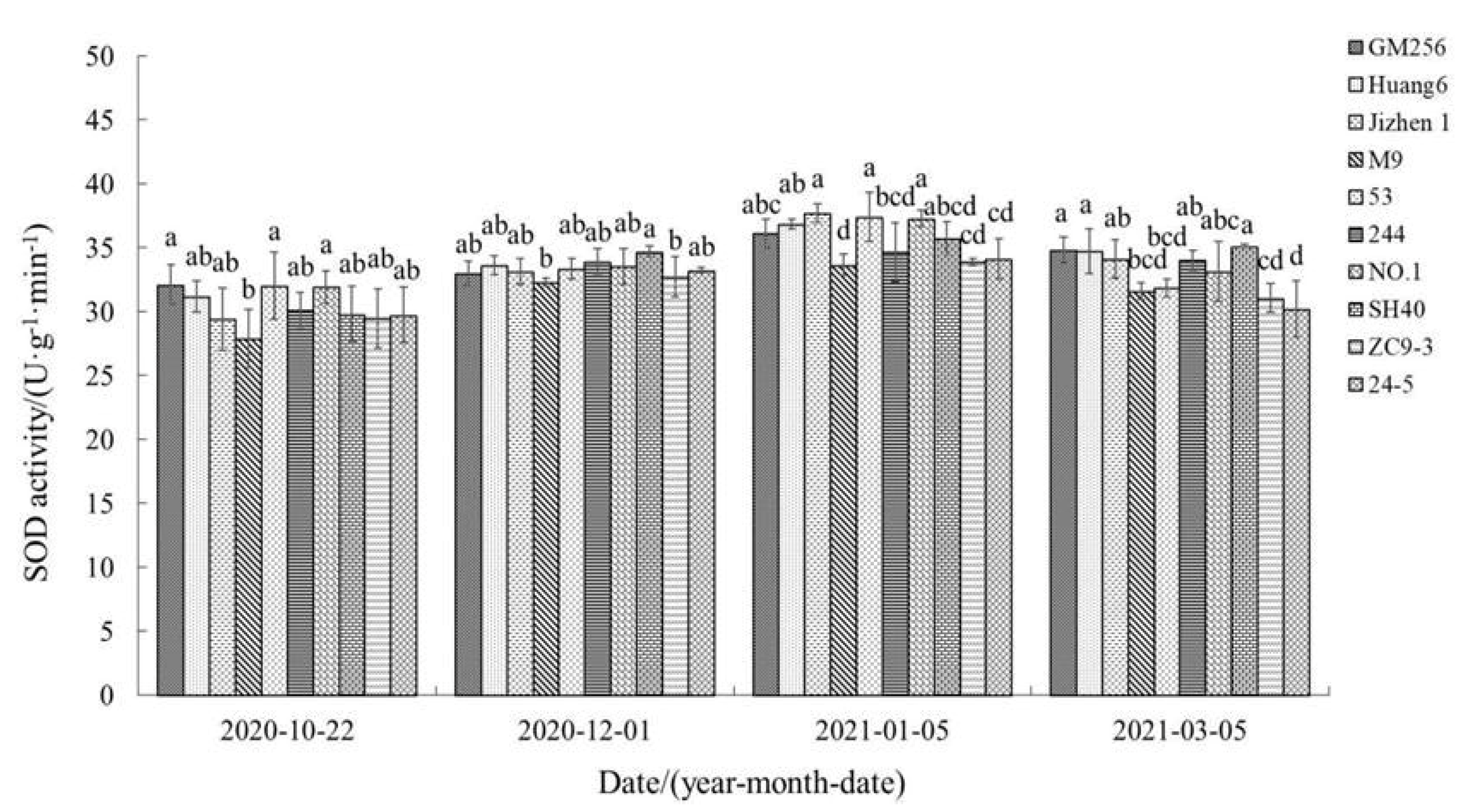
3.2.2. Change of Superoxide Dismutase (SOD) Activity
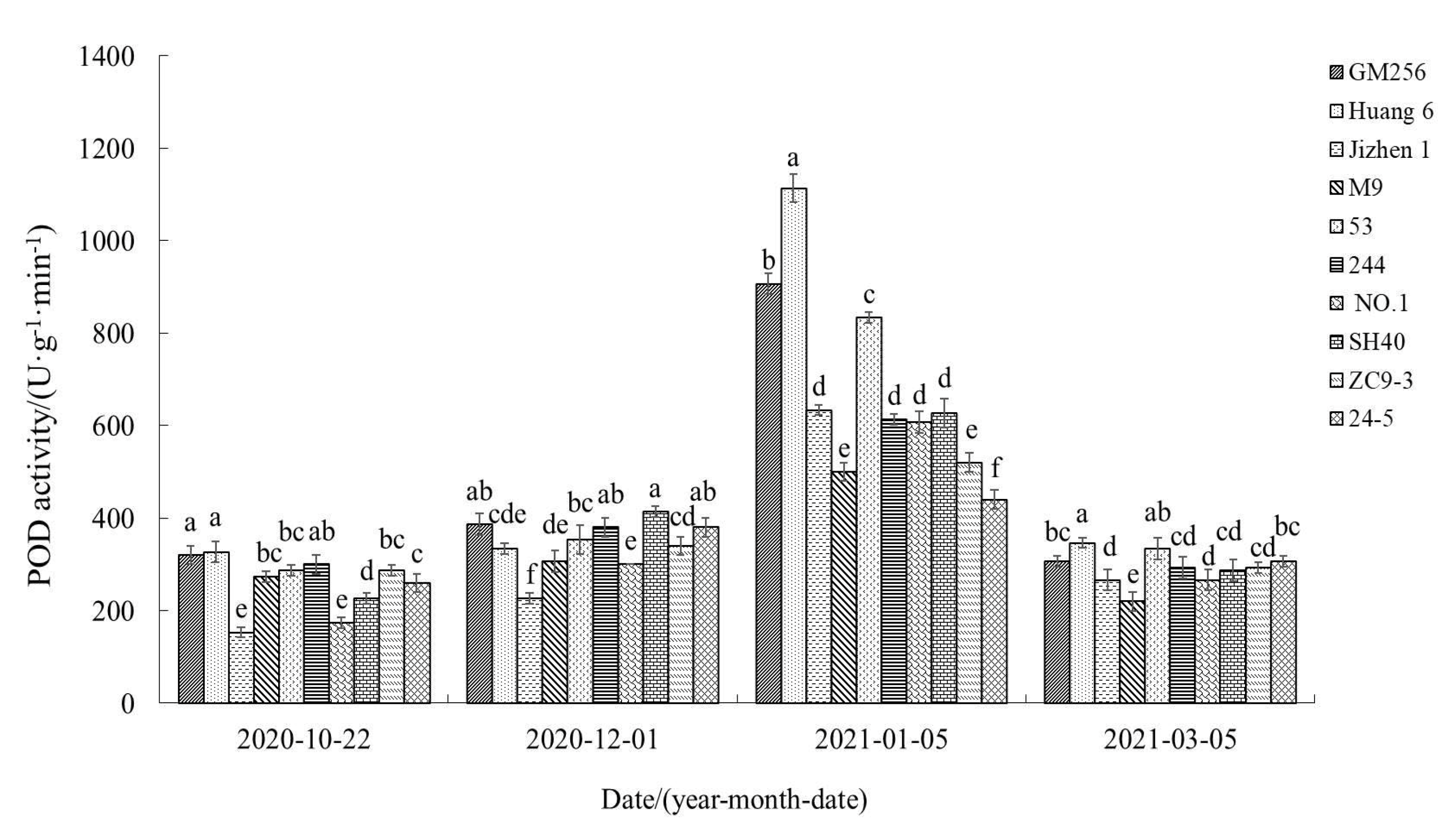
3.2.3. Change of Peroxide Dismutase (POD) Activity
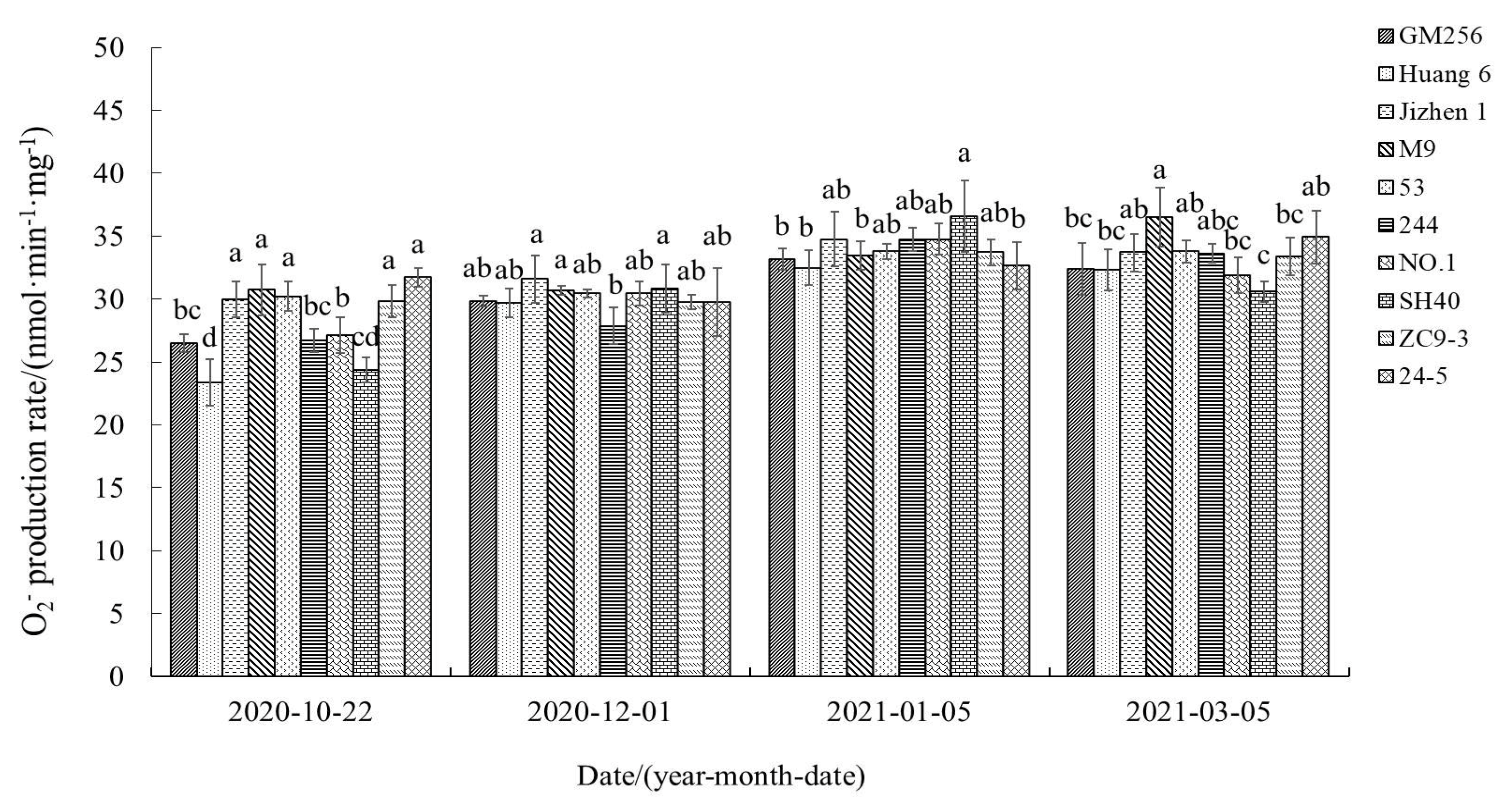
3.2.4. Change of Superoxide Anion (O2−) Production Rate
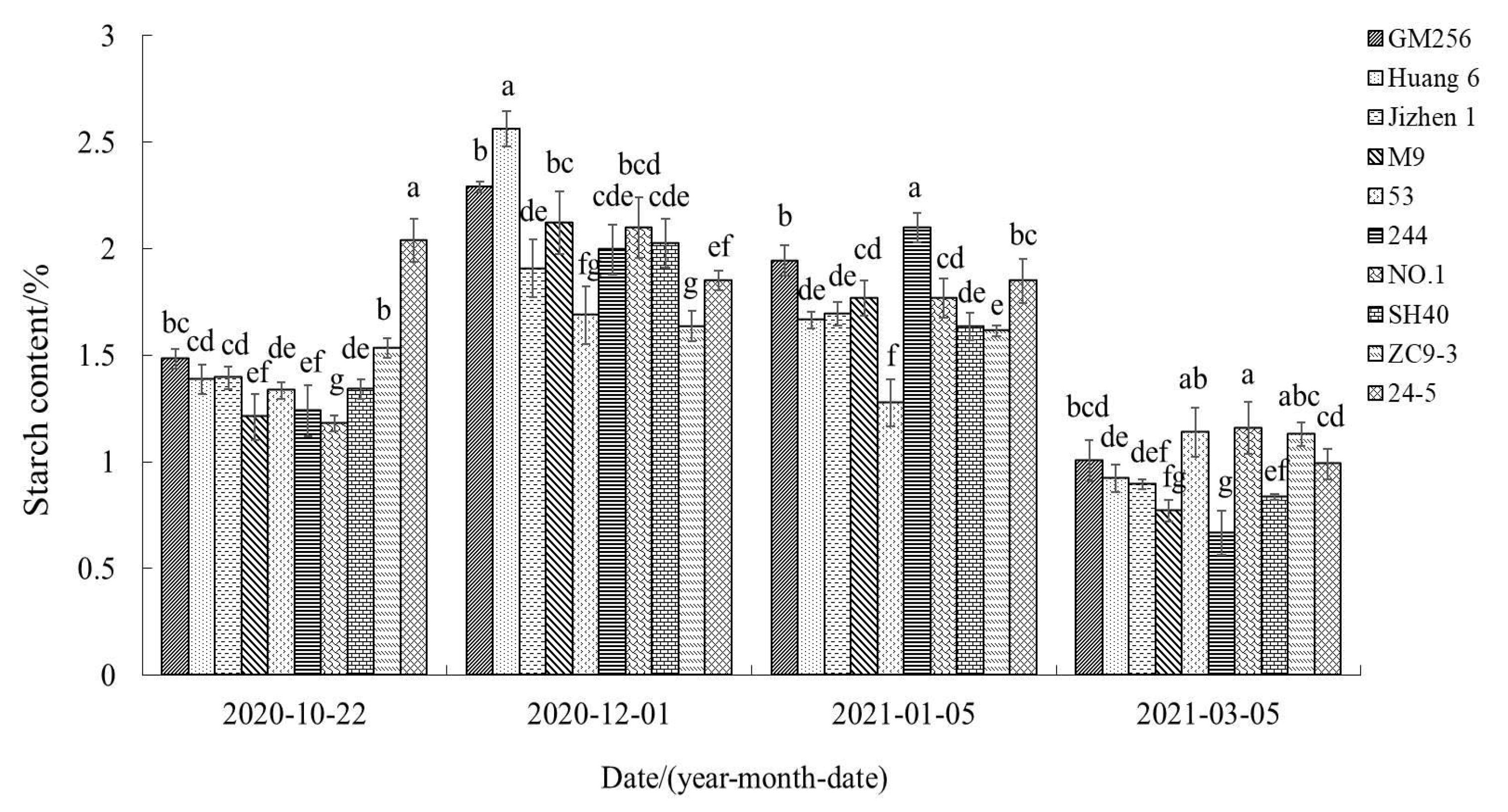
3.2.5. Change of Starch Content
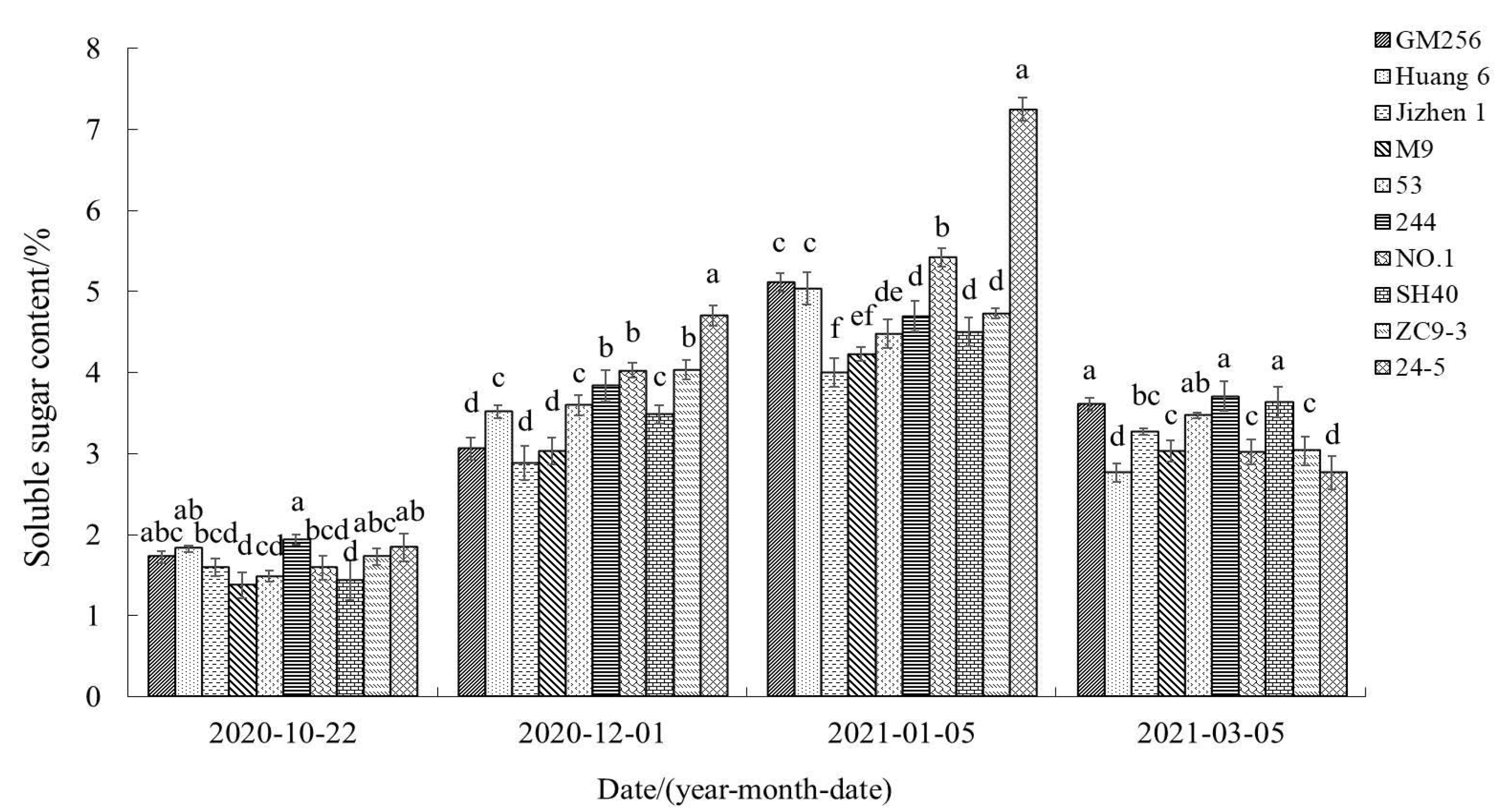
3.2.6. Change of Soluble Sugar Content
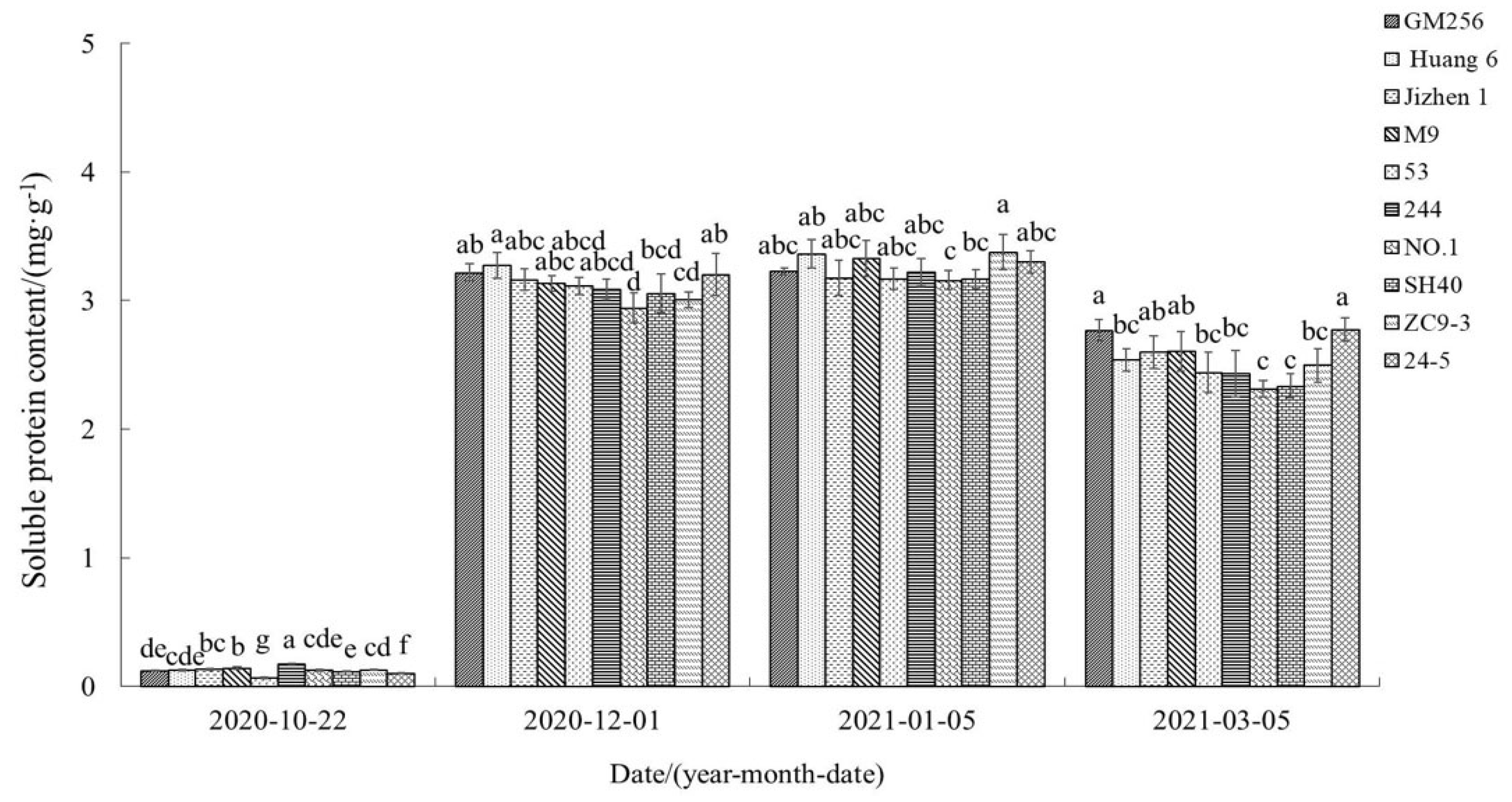
3.2.7. Change of Soluble Protein Content
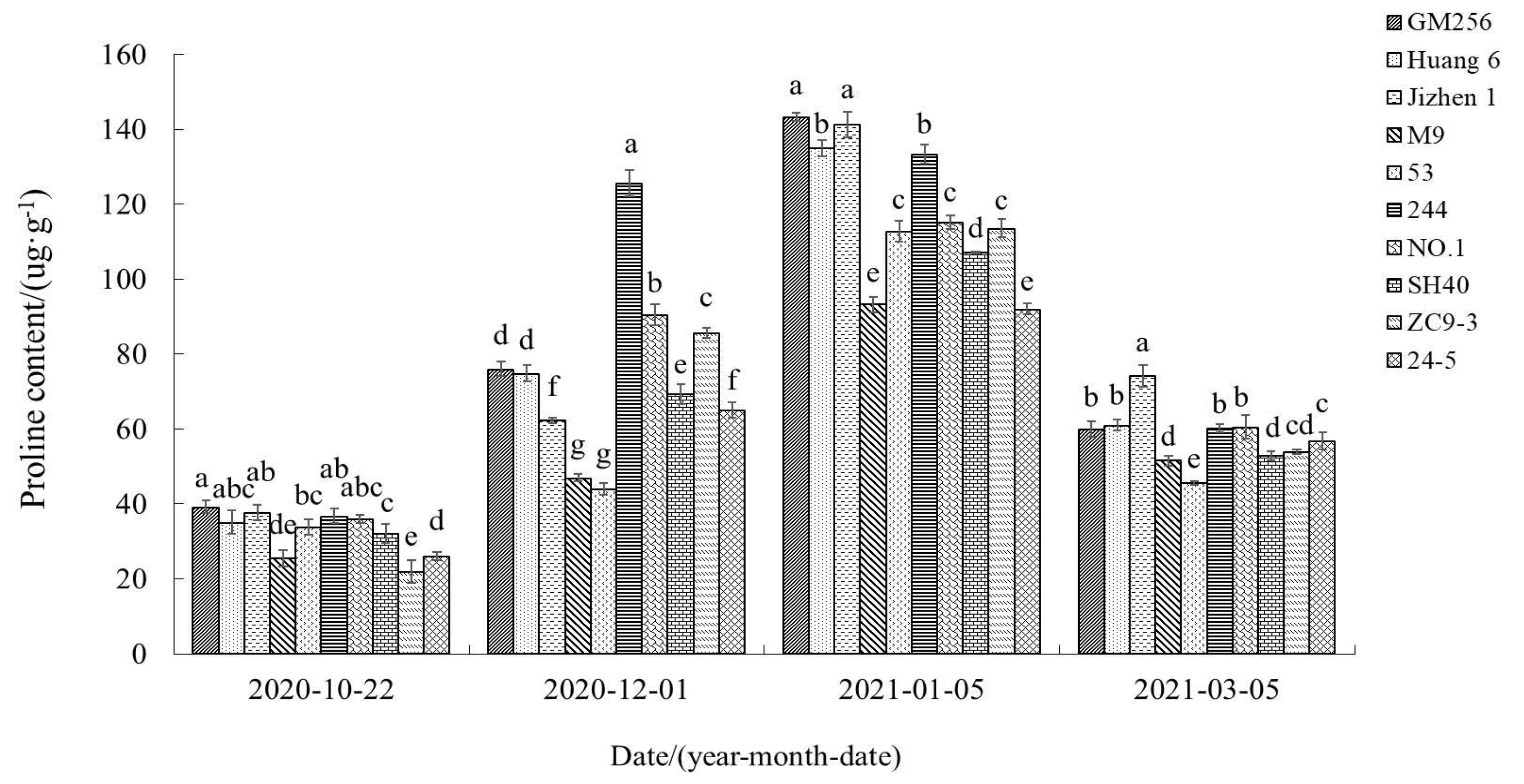
3.2.8. Change of Proline Content
3.3. Comprehensive Evaluation of the Effects of Different Interstocks on the Cold Resistance of ‘Tianhong 2’ Apple Tree
3.3.1. Correlation Analysis of Physiological Indexes of ‘Tianhong 2’ Apple Branches Grafted on Different Interstocks
3.3.2. Principal Component Factor Analysis of Various Indexes of ‘Tianhong 2’ Apple Branches Grafted on Different Interstocks
3.3.3. Comprehensive Evaluation and Classification of Cold Resistance of ‘Tianhong 2’ Apple Branches Grafted on Different Interstocks
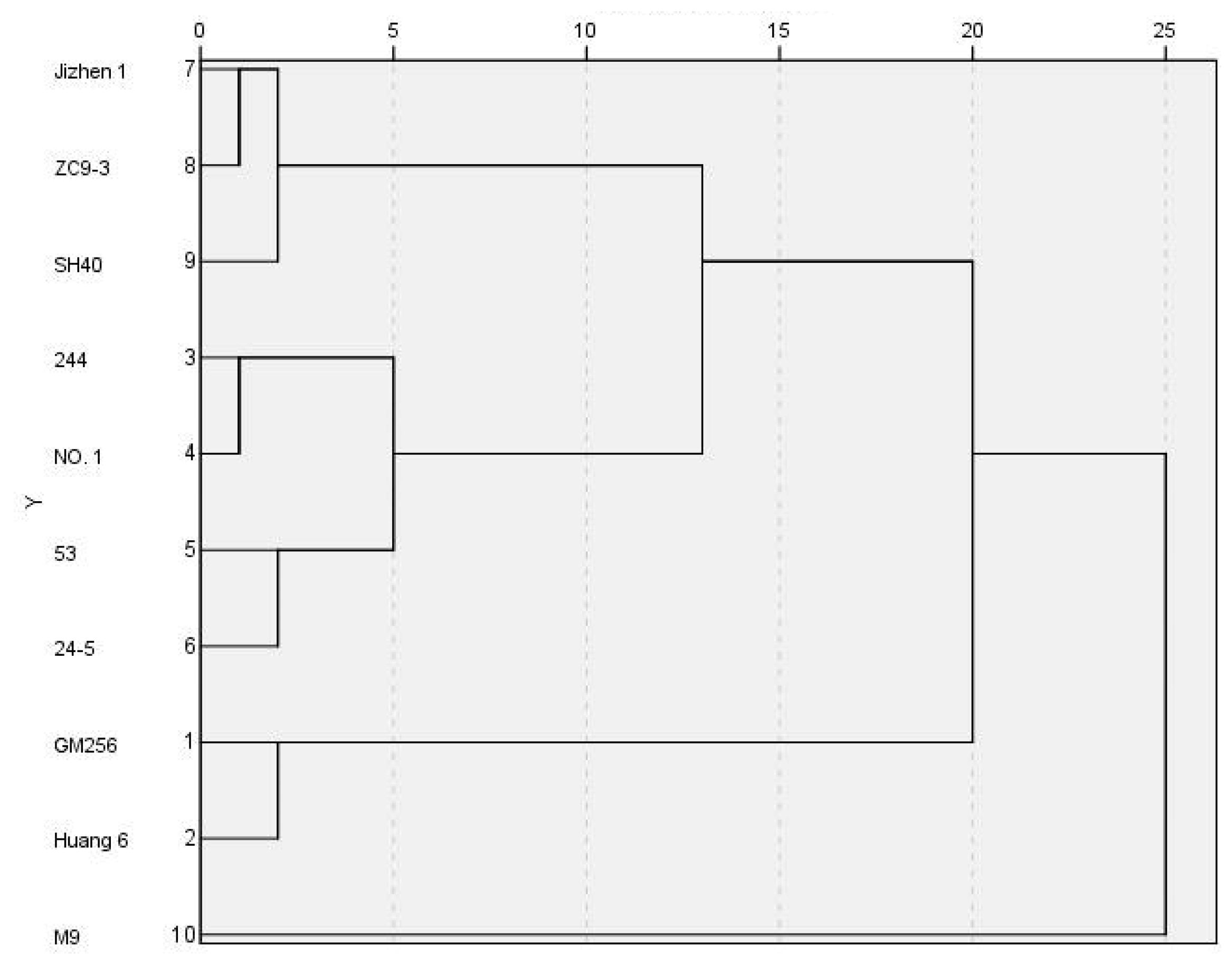
3.4. Cluster Analysis of Cold Resistance of ‘Tianhong 2’ Apple Branches Grafted on Different Interstocks
4. Discussion
4.1. Relationship between LT50 and Cold Resistance
4.2. Relationship between MDA Content and Cold Resistance
4.3. Relationship between Reactive Oxygen Species, Antioxidant System, and Cold Resistance
4.4. Relationship between the Content of Osmotic Adjustment Substancse and Cold Resistance
4.5. Relationship between Starch Content and Cold Resistance
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDA | Malondialdehyde |
| REL | Relative electrolyte leakage |
| LT50 | Semi-lethal temperature |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| ·O2− | Superoxide anion free radical |
| TBA | Thiobarbituric acid |
| NBT | Nitroblue tetrazolium |
References
- Peng, Z.; Fu, D. Effects of 1-methylcyclopropene treatment on the quality of red ‘Fuji’ apples fruit during short-term storage. Food Qual. Saf. 2022, 7, fyac074. [Google Scholar] [CrossRef]
- Wang, N.; Wolf, J.; Zhang, F. Towards sustainable intensification of apple production in China—Yield gaps and nutrient use efficiency in apple farming systems. J. Integr. Agric. 2016, 15, 716–725. [Google Scholar] [CrossRef]
- Shen, X.; Ping, Y.; Bao, C.; Liu, C.; Tahir, M.; Li, X.; Song, Y.; Xu, W.; Ma, F.; Guan, Q. Mdm-miR160-MdARF17-MdWRKY33 module mediates freezing tolerance in apple. Plant J. 2023, 114, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.S.; Büyük, I.; Aras, S. Relationships among lipid peroxidation, sod enzyme activity, and sod gene expression profile in lycopersicum esculentum l. exposed to cold stress. Genet. Mol. Res. 2013, 12, 3220–3229. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, C.; Zhu, J.K. Molecular genetic analysis of cold–regulated gene transcription. Philos. Trans. R. Soc. Ser. B 2002, 357, 877–886. [Google Scholar] [CrossRef]
- Marini, R.P.; Anderson, J.L.; Autio, W.R.; Barritt, B.H.; Unrath, R. Performance of ‘Gala’ apple trees on 18 dwarfing rootstocks: Ten-year summary of the 1994 nc-140 rootstock trial. J. Am. Pomol. Soc. 2006, 60, 69–83. [Google Scholar]
- Wang, Y.X.; Ya, H.U.; Chen, B.H.; Zhu, Y.F.; Dawuda, M.M.; Svetla, S. Physiological mechanisms of resistance to cold stress associated with 10 elite apple rootstocks. J. Integr. Agric. 2018, 17, 857–866. [Google Scholar] [CrossRef]
- Zhou, S.; Shen, Z.; Yin, B.; Liang, B.; Li, Z.; Zhang, X.; Xu, J. Effects of Dwarfifing Interstock Length on the Growth and Fruit of Apple Tree. Horticulturae 2023, 9, 40. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Wu, D.; Hui, M.; Han, X.; Xue, T.; Yao, F.; Gao, F.; Cao, X.; Li, H.; et al. Identification and Regionalization of Cold Resistance of Wine Grape Germplasms (V. vinifera). Agriculture 2021, 11, 1117. [Google Scholar] [CrossRef]
- Melo, P.M.D.; Calvin, F.P.; Sage, R.F. Winter cold-tolerance thresholds in field-grown miscanthus hybrid rhizomes. J. Exp. Bot. 2015, 14, 4415–4425. [Google Scholar]
- Lee, J.; Yu, D.; Lee, J.; Kim, S.; Lee, H. Comparison of mid-Winter cold-hardiness and soluble sugars contents in the shoots of 21 highbush blueberry (Vaccinium corymbosum) cultivars. J. Hortic. Sci. Biotechnol. 2013, 88, 727–734. [Google Scholar] [CrossRef]
- Cansev, A.; Gulen, H.; Eris, A. The activities of catalase and ascorbate peroxidase in olive (Olea europaea L. cv. Gemlik) under low temperature stress. Hortic. Environ. Biotechnol. 2011, 52, 113–120. [Google Scholar] [CrossRef]
- Sheng, L.; Sun, X.; Mo, C.; Hao, M.; Wei, X.; Ma, A. Relationship between antioxidant enzymes and sclerotial formation of Pleurotus tuber-regium under abiotic stress. Appl. Microbiol. Biotechnol. 2023, 107, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Saruhan, N.; Turgut-Terzi, R.; Kadioglu, A. The effects of exogenous polyamines on some biochemical changes during drought stress in Ctenanthe setosa (Rosc.) Eichler. Acta Biol. Hung. 2018, 57, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, Z.; Saleh, A.S.M.; Lu, Y.; Zhang, X.; Ge, X.; Shen, H.; Yu, X.; Li, W. Effects of various microwave intensities collaborated with different cold plasma duration time on structural, physicochemical, and digestive properties of lotus root starch. Food Chem. 2023, 405, 134837. [Google Scholar] [CrossRef]
- Wang, J.W.; Ma, L.Y.; Gómez-Del-Campo, M.; Zhang, D.S.; Deng, Y.; Jia, Z.K. Youth tree behavior of olive (Olea europaea L.) cultivars in wudu, China: Cold and drought resistance, growth, fruit production, and oil quality. Sci. Hortic. 2018, 236, 106–122. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Y.; Zhu, Y.; Chen, B. Multivariate statistical analyses and predictive model of cold resistance associated with eleven crabapples and fuji apple. Cryoletters 2018, 39, 235–244. [Google Scholar]
- Wilner, J.J. Note on an electrolytic procedure for differentiating between frost injury of roots and shoots in woody plants. Can. J. Plant Sci. 1959, 39, 512–513. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Yang, L.T.; Li, Y.R. Physiological and Biochemical characteristics related to cold resistance in sugarcane. Sugar Tech 2015, 17, 49–58. [Google Scholar] [CrossRef]
- Du, F.; Shi, H.; Zhang, X.; Xu, X. Responses of Reactive Oxygen Scavenging Enzymes, Proline and Malondialdehyde to Water Deficits among Six Secondary Successional Seral Species in Loess Plateau. PLoS ONE 2014, 9, e98872. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, X.; Wang, X.; Yan, C.; Ma, C.; Liu, L.; Dong, S. Effects of the Soil Moisture Content on the Superoxide Anion and Proline Contents in Soybean Leaves. Legume Res. 2022, 45, 315–318. [Google Scholar] [CrossRef]
- Qian, J.; Zhou, J.; Di, B.; Liu, Y.; Zhang, G.; Yang, X. Using electrical impedance tomography for rapid determination of starch and soluble sugar contents in Rosa hybrida. Sci. Rep. 2021, 11, 2871. [Google Scholar] [CrossRef]
- Shahi, S.; Katiyar, R.K.; Bhatnagar, A.K.; Singh, A.B. Soluble and nonsoluble protein assay for antigenic extracts from pollen and seeds of mustard (Brassica spp.). Allergy Asthma Proc. 2008, 29, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Li, P.; Su, Z.; Li, M.; Guo, S. Cold-tolerant introgression line construction and low-temperature stress response analysis for bell pepper. Plant Signal. Behav. 2020, 15, e1773097. [Google Scholar] [CrossRef]
- Lyons, J.M. Chilling injury in plants. Annu. Rev. Plant Physiol. 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Campos, P.S.; nia Quartin, V.; chicho Ramalho, J.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Yang, J.; Yang, X.; Chen, S.; Xie, Z.; Zhang, M.; Huang, Y.; Zhang, J.; Huang, X. Genome-Wide Analysis and Expression of Cyclic Nucleotide-Gated Ion Channel (CNGC) Family Genes under Cold Stress in Mango (Mangifera indica). Plants 2023, 12, 592. [Google Scholar] [CrossRef]
- Sato, Y.; Masuta, Y.; Saito, K.; Murayama, S.; Ozawa, K. Enhanced chilling tolerance at the booting stage in rice by transgenic overexpression of the ascorbate peroxidase gene, OsAPXa. Plant Cell Rep. 2011, 30, 399–406. [Google Scholar] [CrossRef]
- Saadati, S.; Baninasab, B.; Mobli, M.; Gholami, M. Cold Tolerance in Olive Leaves of Three Cultivars Related to Some Physiological Parameters during Cold Acclimation and De-Acclimation Stages. J. Agric. Sci. Technol. 2020, 22, 1313–1326. [Google Scholar]
- Boyer, J.S. Grain yields with limited water. J. Exp. Bot. 2004, 55, 2385–2394. [Google Scholar] [CrossRef]
- Javadian, N.; Karimzadeh, G.; Mahfoozi, S.; Ghanati, F. Cold-induced changes of enzymes, proline, carbohydrates, and chlorophyll in wheat. Russ. J. Plant Physiol. 2010, 57, 540–547. [Google Scholar] [CrossRef]
- Türkan, İ.; Bor, M.; Özdemir, F.; Koca, H. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci. 2005, 168, 223–231. [Google Scholar] [CrossRef]
- Sun, B.; Liu, G.; Phan, T.; Yang, L.; Li, Y.; Xing, Y. Effects of Cold Stress on Root Growth and Physiological Metabolisms in Seedlings of Different Sugarcane Varieties. Sugar Tech 2017, 19, 165–175. [Google Scholar] [CrossRef]
- Bao, J.; Zhang, S. Changes in germination, storage materials and antioxidant enzyme activities in pear (Pyrus betulaefolia Bge. and Pyrus calleryana Dcne.) stock seeds during cold stratification. Seed Sci. Technol. 2011, 39, 655–659. [Google Scholar] [CrossRef]
- Albina, A.; Wang, Q.Y.; Ludmila, K.; Peter, N. Is microtubule disassembly a trigger for coldacclimation? Plant Cell Physiol. 2003, 44, 676–686. [Google Scholar]
- Bertrand, A.; Bipfubusa, M.; Claessens, A.; Rocher, S.; Castonguay, Y. Effect of photoperiod prior to cold acclimation on freezing tolerance and carbohydrate metabolism in alfalfa (Medicago sativa L.). Plant Sci. 2017, 264, 122–128. [Google Scholar] [CrossRef]
- Castonguay, Y.; Bertrand, A.; Michaud, R.; Laberge, S. Cold-Induced Biochemical and Molecular Changes in Alfalfa Populations Selectively Improved for Freezing Tolerance. Crop Sci. 2011, 51, 213. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rootstock | Parents | Breeding Institution |
|---|---|---|
| Jizhen 1 | seedling progeny of SH40 | Hebei Agricultural University |
| 53 | seedling progeny of SH40 | |
| 244 | seedling progeny of SH40 | |
| NO. 1 | seedling progeny of SH40 | |
| 24-5 | Malus micromalus × Inner Mongolia11 | |
| Huang 6 | seedling progeny of Malus robusta Rehd | |
| ZC9-3 | seedling progeny of P22 |
| Sampling Date | Natural Wintering Temperature/ °C | ||
|---|---|---|---|
| Maximum Temperature | Minimum Temperature | Average Temperature | |
| 22 October 2020 | 18 | 2 | 9.46 |
| 1 December 2020 | 1 | −5 | −1.08 |
| 5 January 2021 | −2 | −14 | −7.33 |
| 5 March 2021 | 10 | 1 | 4.29 |
| Sampling Date | Temperature Settings/ °C | |||||
|---|---|---|---|---|---|---|
| 22 October 2020 | 4 | −8 | −20 | −26 | −36 | −45 |
| 1 December 2020 | 4 | −8 | −22 | −36 | −50 | −60 |
| 5 January 2021 | 4 | −10 | −22 | −45 | −55 | −65 |
| 5 March 2021 | 4 | −12 | −20 | −28 | −36 | −45 |
| Correlation Coefficient | LT50 | SOD | POD | O2− | Protein | MDA | Starch | Proline | Sugar |
|---|---|---|---|---|---|---|---|---|---|
| LT50 | 1 | ||||||||
| SOD | −0.797 ** | 1 | |||||||
| POD | −0.630 ** | 0.713 ** | 1 | ||||||
| O2− | −0.673 ** | 0.455 ** | 0.387 * | 1 | |||||
| Protein | −0.931** | 0.722 ** | 0.492 ** | 0.608 ** | 1 | ||||
| MDA | −0.692 ** | 0.611 ** | 0.507 ** | 0.614 ** | 0.688 ** | 1 | |||
| Starch | −0.164 | 0.184 | 0.283 | −0.125 | 0.314 * | 0.04 | 1 | ||
| Proline | −0.799 ** | 0.810 ** | 0.807 ** | 0.481 ** | 0.726 ** | 0.605 ** | 0.382 * | 1 | |
| Sugar | −0.847 ** | 0.748 ** | 0.642 ** | 0.541 ** | 0.824 ** | 0.706 ** | 0.308 | 0.790 ** | 1 |
| Principal Component | Initial Eigenvalue | ||
|---|---|---|---|
| Characteristic Root | Contribution Rate (%) | Cumulative Contribution Rate (%) | |
| 1 | 5.83 | 64.773 | 64.773 |
| 2 | 1.241 | 13.789 | 78.562 |
| Load | Eigenvector | |||
|---|---|---|---|---|
| Biochemical Indexes | Component 1 | Component 2 | Component 1 | Component 2 |
| LT50 | −0.942 | 0.121 | −0.390 | 0.109 |
| Sugar | 0.911 | 0.056 | 0.377 | 0.050 |
| Proline | 0.902 | 0.226 | 0.374 | 0.203 |
| Protein | 0.896 | −0.025 | 0.371 | −0.022 |
| SOD | 0.869 | 0.053 | 0.360 | 0.048 |
| MDA | 0.787 | −0.291 | 0.326 | −0.261 |
| POD | 0.768 | 0.232 | 0.318 | 0.208 |
| O2− | 0.672 | −0.538 | 0.278 | −0.483 |
| Starch | 0.287 | 0.861 | 0.119 | 0.773 |
| Interstock | Principal Component Value | Comprehensive Score | Cold Resistance Order (1, Highest to 10, Lowest) | Classification | |
|---|---|---|---|---|---|
| Component 1 | Component 2 | ||||
| GM256 | 1.420503 | 2.516214 | 1.267063 | 1 | Strong tolerance |
| Huang 6 | 0.940823 | 2.989505 | 1.021622 | 2 | Strong tolerance |
| Jizhen1 | −0.28465 | −1.03902 | −0.32764 | 7 | Mean tolerance |
| M9 | −2.81883 | −1.96704 | −2.09708 | 10 | Weak tolerance |
| 53 | 0.784528 | −2.20221 | 0.204499 | 5 | Relatively resistant |
| 244 | 0.554611 | 0.944982 | 0.489542 | 3 | Relatively resistant |
| NO. 1 | 0.660832 | 0.020357 | 0.430848 | 4 | Relatively resistant |
| SH40 | −0.95421 | 0.244786 | −0.58431 | 9 | Mean tolerance |
| ZC9-3 | −0.2112 | −1.60406 | −0.35798 | 8 | Mean tolerance |
| 24-5 | −0.09241 | 0.096477 | −0.04656 | 6 | Relatively resistant |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, J.; Liu, M.; Yin, B.; Liang, B.; Li, Z.; Zhang, X.; Xu, J.; Zhou, S. Effects of 10 Dwarfing Interstocks on Cold Resistance of ‘Tianhong 2’ Apple. Horticulturae 2023, 9, 827. https://doi.org/10.3390/horticulturae9070827
Jing J, Liu M, Yin B, Liang B, Li Z, Zhang X, Xu J, Zhou S. Effects of 10 Dwarfing Interstocks on Cold Resistance of ‘Tianhong 2’ Apple. Horticulturae. 2023; 9(7):827. https://doi.org/10.3390/horticulturae9070827
Chicago/Turabian StyleJing, Junli, Mingxiao Liu, Baoying Yin, Bowen Liang, Zhongyong Li, Xueying Zhang, Jizhong Xu, and Shasha Zhou. 2023. "Effects of 10 Dwarfing Interstocks on Cold Resistance of ‘Tianhong 2’ Apple" Horticulturae 9, no. 7: 827. https://doi.org/10.3390/horticulturae9070827
APA StyleJing, J., Liu, M., Yin, B., Liang, B., Li, Z., Zhang, X., Xu, J., & Zhou, S. (2023). Effects of 10 Dwarfing Interstocks on Cold Resistance of ‘Tianhong 2’ Apple. Horticulturae, 9(7), 827. https://doi.org/10.3390/horticulturae9070827