


Foliar Fertilization with Molybdate and Nitrate Up-Regulated Activity of Nitrate Reductase in Lemon Balm Leaves

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lemon Balm Cultivation
2.2. Foliar Fertilization
2.3. Determination of the Amount of Nitrate
2.4. Determination of NR Activity In Vivo
2.5. Preparation of Samples for Analysis of NR Activity and Determination of Mo-Co Content
2.6. Determination of NR Activity In Vitro
2.7. Determination of Mo-Co Content
2.8. Statistical Data Analysis
3. Results
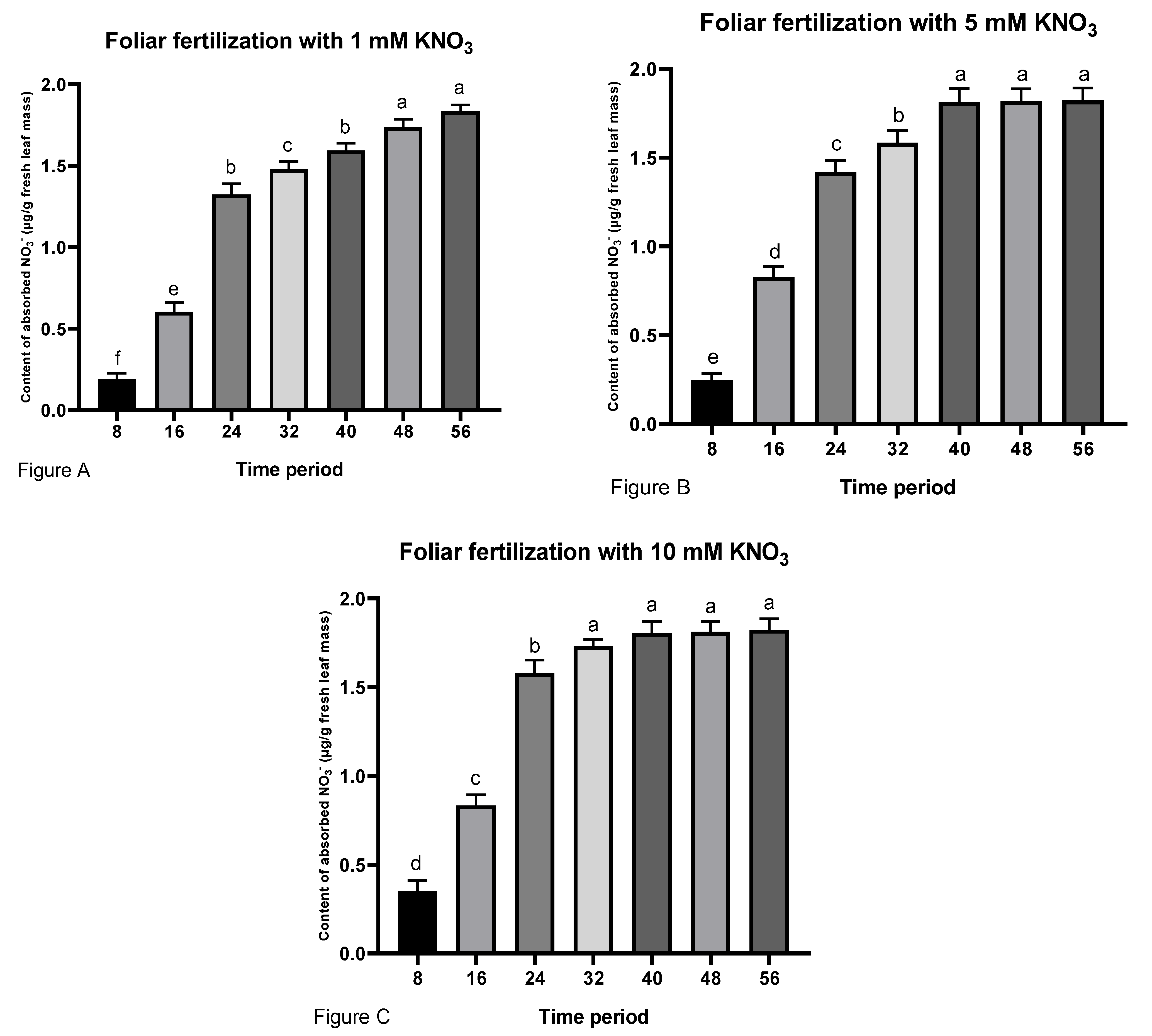
3.1. Effect of Foliar Fertilization with Potassium Nitrate on Nitrate Absorbtion and NR Activity within 56 h after Spraying
3.2. Effect of Foliar Fertilization with Sodium Molybdate on NR Activity within 56 h after Spraying
3.3. Mo-Co Content in the Lemon Balm Leaves within 56 h after Potassium Nitrate Spraying
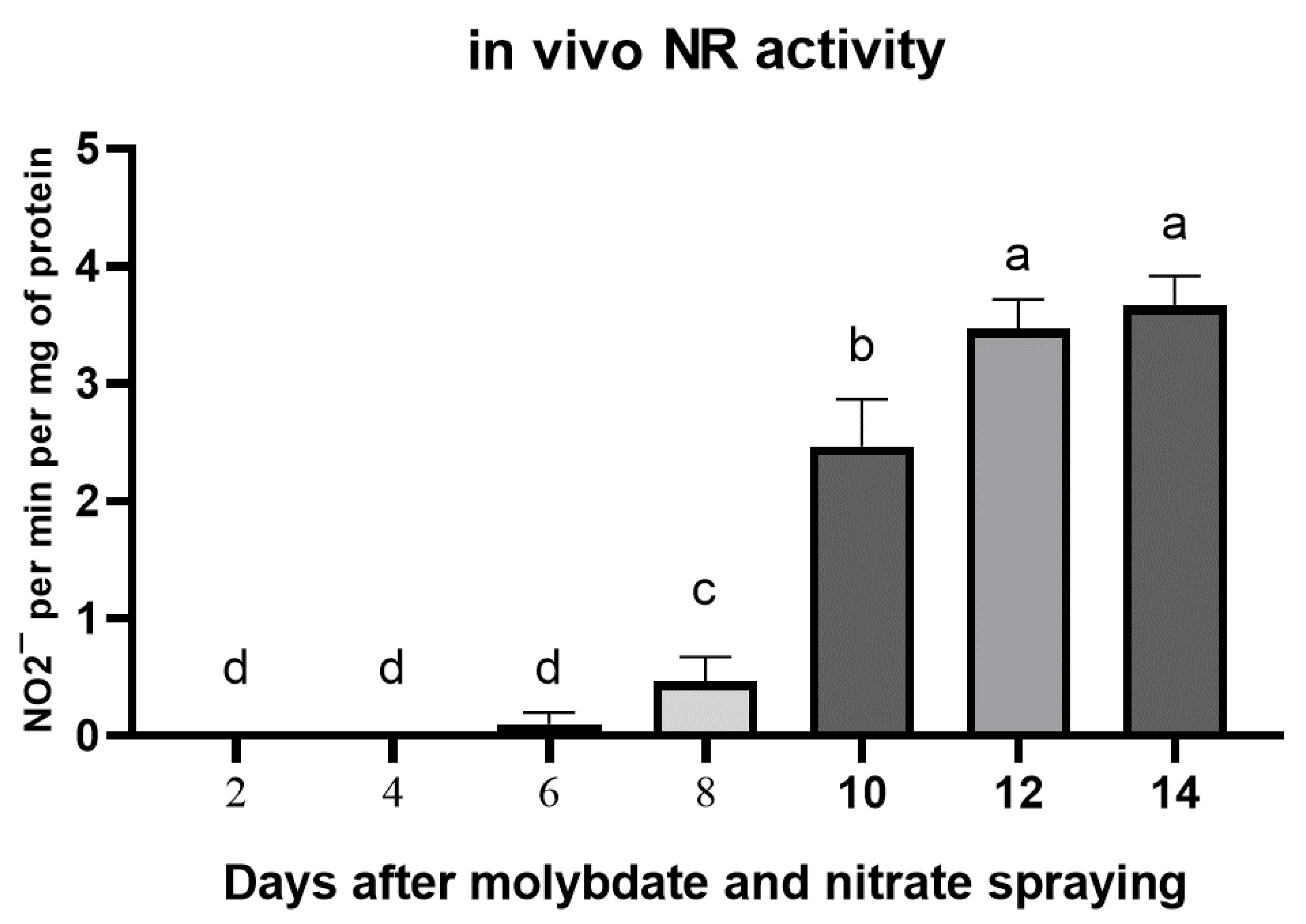
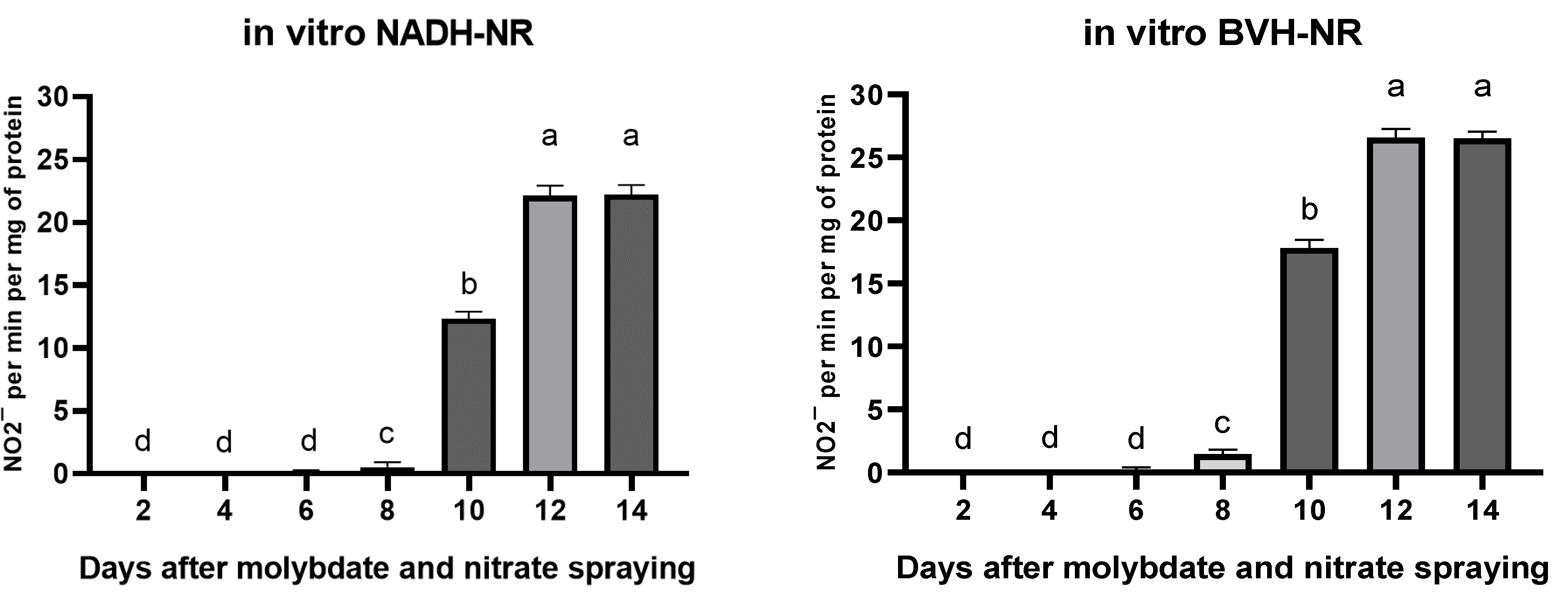
3.4. Effect of Foliar Fertilization with Sodium Molybdate and Nitrate on NR Activity within 15 Days after Spraying
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aulakh, S.K.; Varma, S.J.; Ralser, M. Metal Ion Availability and Homeostasis as Drivers of Metabolic Evolution and Enzyme Function. Curr. Opin. Genet. Dev. 2022, 77, 101987. [Google Scholar] [CrossRef]
- Cordas, C.M.; Moura, J.J.G. Molybdenum and Tungsten Enzymes Redox Properties—A Brief Overview. Coord. Chem. Rev. 2019, 394, 53–64. [Google Scholar] [CrossRef]
- Hille, R.; Hall, J.; Basu, P. The Mononuclear Molybdenum Enzymes. Chem. Rev. 2014, 114, 3963–4038. [Google Scholar] [CrossRef]
- Tejada-Jimenez, M.; Chamizo-Ampudia, A.; Calatrava, V.; Galvan, A.; Fernandez, E.; Llamas, A. From the Eukaryotic Molybdenum Cofactor Biosynthesis to the Moonlighting Enzyme mARC. Molecules 2018, 23, 3287. [Google Scholar] [CrossRef]
- Zhang, W.; Ni, K.; Long, L.; Ruan, J. Nitrogen Transport and Assimilation in Tea Plant (Camellia Sinensis): A Review. Front. Plant Sci. 2023, 14, 1249202. [Google Scholar] [CrossRef]
- Calatrava, V.; Chamizo-Ampudia, A.; Sanz-Luque, E.; Ocaña-Calahorro, F.; Llamas, A.; Fernandez, E.; Galvan, A. How Chlamydomonas Handles Nitrate and the Nitric Oxide Cycle. J. Exp. Bot. 2017, 68, 2593–2602. [Google Scholar] [CrossRef] [PubMed]
- Aubakirova, K.; Satkanov, M.; Kulataeva, M.; Assylbekova, G.; Kambarbekova, A.; Alikulov, Z. Molybdoenzymes Isolated from S. Glanis Liver Can Produce Nitric Oxide from Nitrates and Nitrites. Czech J. Anim. Sci. 2023, 68, 222–230. [Google Scholar] [CrossRef]
- Kaur, R.; Chandra, J.; Varghese, B.; Keshavkant, S. Allantoin: A Potential Compound for the Mitigation of Adverse Effects of Abiotic Stresses in Plants. Plants 2023, 12, 3059. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kamanga, B.M.; Zhang, W.; Xu, Y.; Xu, L. Research Progress of Aldehyde Oxidases in Plants. PeerJ 2022, 10, e13119. [Google Scholar] [CrossRef]
- Nurbekova, Z.; Srivastava, S.; Standing, D.; Kurmanbayeva, A.; Bekturova, A.; Soltabayeva, A.; Oshanova, D.; Turečková, V.; Strand, M.; Biswas, M.S.; et al. Arabidopsis Aldehyde Oxidase 3, Known to Oxidize Abscisic Aldehyde to Abscisic Acid, Protects Leaves from Aldehyde Toxicity. Plant J. 2021, 108, 1439–1455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, H.; Zhang, Y.; Wang, Y.; Li, M.; Zhang, J.; Duan, L.; Zhang, M.; Li, Z. Increased Abscisic Acid Levels in Transgenic Maize Overexpressing AtLOS5 Mediated Root Ion Fluxes and Leaf Water Status under Salt Stress. J. Exp. Bot. 2016, 67, 1339–1355. [Google Scholar] [CrossRef] [PubMed]
- Mukhamejanova, A.; Alikulov, Z.; Shapekova, N.; Aubakirova, K.; Mukhtarov, A. The Effect of Antioxidants on Xanthine Oxidase Activity in Fresh Ovine Milk. Potravin. Slovak J. Food Sci. 2021, 15, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Hänsch, R.L.; Lang, C.; Rennenberg, H.; Mendel, R.R. Significance of Plant Sulfite Oxidase. Plant Biol. 2007, 9, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R.; Schwarz, G. The History of Animal and Plant Sulfite Oxidase—A Personal View. Molecules 2023, 28, 6998. [Google Scholar] [CrossRef]
- Llamas, A.; Chamizo-Ampudia, A.; Tejada-Jimenez, M.; Galvan, A.; Fernandez, E. The Molybdenum Cofactor Enzyme mARC: Moonlighting or Promiscuous Enzyme? BioFactors 2017, 43, 486–494. [Google Scholar] [CrossRef]
- Rothery, R.A.; Stein, B.; Solomonson, M.; Kirk, M.L.; Weiner, J.H. Pyranopterin Conformation Defines the Function of Molybdenum and Tungsten Enzymes. Proc. Natl. Acad. Sci. USA 2012, 109, 14773–14778. [Google Scholar] [CrossRef]
- Basu, P.; Burgmayer, S.J.N. Pterin Chemistry and Its Relationship to the Molybdenum Cofactor. Coord. Chem. Rev. 2011, 255, 1016–1038. [Google Scholar] [CrossRef]
- Llamas, A.; Otte, T.; Multhaup, G.; Mendel, R.R.; Schwarz, G. The Mechanism of Nucleotide-Assisted Molybdenum Insertion into Molybdopterin: A Novel Route Toward Metal Cofactor Assembly. J. Biol. Chem. 2006, 281, 18343–18350. [Google Scholar] [CrossRef]
- Kaufholdt, D.; Baillie, C.-K.; Meinen, R.; Mendel, R.R.; Hänsch, R. The Molybdenum Cofactor Biosynthesis Network: In Vivo Protein-Protein Interactions of an Actin Associated Multi-Protein Complex. Front. Plant Sci. 2017, 8, 1946. [Google Scholar] [CrossRef]
- Kruse, T. Moco Carrier and Binding Proteins. Molecules 2022, 27, 6571. [Google Scholar] [CrossRef] [PubMed]
- Kathpalia, R.; Bhatla, S. Plant Mineral Nutrition. In Plant Physiology, Development and Metabolism; Springer: Berlin/Heidelberg, Germany, 2018; pp. 37–81. ISBN 9789811320224. [Google Scholar]
- Ventura, Y.; Wuddineh, W.A.; Ephrath, Y.; Shpigel, M.; Sagi, M. Molybdenum as an Essential Element for Improving Total Yield in Seawater-Grown Salicornia Europaea L. Sci. Hortic. 2010, 126, 395–401. [Google Scholar] [CrossRef]
- Rana, M.S.; Bhantana, P.; Sun, X.; Imran, M.; Shaaban, M.; Moussa, M.G.; Saleem, M.H.; Elyamine, A.M.; Binyamin, R.; Alam, M.; et al. Molybdenum as an Essential Element for Crops: An Overview. BJSTR 2020, 24, 18535–18547. [Google Scholar] [CrossRef]
- Kentelky, E.; Szekely-Varga, Z. Impact of Foliar Fertilization on Growth, Flowering, and Corms Production of Five Gladiolus Varieties. Plants 2021, 10, 1963. [Google Scholar] [CrossRef]
- Zveushe, O.K.; Ling, Q.; Li, X.; Sajid, S.; Dios, V.R.; Nabi, F.; Han, Y.; Dong, F.; Zeng, F.; Zhou, L.; et al. Reduction of Cd Uptake in Rice (Oryza Sativa) Grain Using Different Field Management Practices in Alkaline Soils. Foods 2023, 12, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, A.; Sakthivel, N.; Subramanian, E.; Rajesh, P. Productivity of Field Crops as Influenced by Foliar Spray of Nutrients: A Review. Agric. Rev. 2020, 41, 146–152. [Google Scholar] [CrossRef]
- López-Tejeida, S.; Felix-Cuencas, L.; De León-Ramirez, J.J.; Flores-Tejeida, L.B.; Villegas-Villegas, M.; García-Trejo, J.F. Methods to Measure Nitrogen Content in Fish Culture: A Review. In Proceedings of the XVII International Engineering Congress (CONIIN), Queretaro, Mexico, 14 June 2021; pp. 1–5. [Google Scholar]
- Liu, X.-Y.; Koba, K.; Koyama, L.A.; Hobbie, S.E.; Weiss, M.S.; Inagaki, Y.; Shaver, G.R.; Giblin, A.E.; Hobara, S.; Nadelhoffer, K.J.; et al. Nitrate Is an Important Nitrogen Source for Arctic Tundra Plants. Proc. Natl. Acad. Sci. USA 2018, 115, 3398–3403. [Google Scholar] [CrossRef]
- Awad, R.; Muhammad, A.; Durst, T.; Trudeau, V.L.; Arnason, J.T. Bioassay-Guided Fractionation of Lemon Balm (Melissa Officinalis L.) Using an in Vitro Measure of GABA Transaminase Activity. Phytother. Res. 2009, 23, 1075–1081. [Google Scholar] [CrossRef]
- Shakeri, A.; Sahebkar, A.; Javadi, B. Melissa Officinalis L.—A Review of Its Traditional Uses, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2016, 188, 204–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Mun, B.-G.; Khan, A.L.; Waqas, M.; Kim, H.-H.; Shahzad, R.; Imran, M.; Yun, B.-W.; Lee, I.-J. Regulation of Reactive Oxygen and Nitrogen Species by Salicylic Acid in Rice Plants under Salinity Stress Conditions. PLoS ONE 2018, 13, e0192650. [Google Scholar] [CrossRef] [PubMed]
- Kadyrbaev, M.K.; Golovatskaya, I.F.; Satkanov, M.Z. Features of Regenerants Morphogenesis and Metabolism in Vitro, Obtained from Different Fragments of Potato Shoots. Vestn. Tomsk. Gos. Univ. Biol. 2021, 55, 114–134. [Google Scholar] [CrossRef]
- Nicholas, J.C.; Harper, J.E.; Hageman, R.H. Nitrate Reductase Activity in Soybeans (Glycine Max [L.] Merr.): I. Effects of Light and Temperature. Plant Physiol. 1976, 58, 731–735. [Google Scholar] [CrossRef]
- Sisay, T.A.; Nurbekova, Z.; Oshanova, D.; Dubey, A.K.; Khatri, K.; Mudgal, V.; Mudgal, A.; Neori, A.; Shpigel, M.; Srivastava, R.K.; et al. Effect of Salinity and Nitrogen Fertilization Levels on Growth Parameters of Sarcocornia Fruticosa, Salicornia Brachiata, and Arthrocnemum Macrostachyum. Agronomy 2022, 12, 1749. [Google Scholar] [CrossRef]
- Li, Z.; Liu, W.; Wu, C.; She, D. Effect of Spraying Direction on the Exposure to Handlers with Hand-Pumped Knapsack Sprayer in Maize Field. Ecotoxicol. Environ. Saf. 2019, 170, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Kovács, B.; Puskás-Preszner, A.; Huzsvai, L.; Lévai, L.; Bódi, É. Effect of Molybdenum Treatment on Molybdenum Concentration and Nitrate Reduction in Maize Seedlings. Plant Physiol. Biochem. 2015, 96, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Irving, G.C.J.; Bouma, D. Rapid and Simple Leaf Tissue Test for Nitrate. Commun. Soil Sci. Plant Anal. 1986, 17, 1299–1310. [Google Scholar] [CrossRef]
- Mukhamejanova, A.; Alikulov, Z.; Tuganova, B.; Adamzhanova, Z. The Xanthine Oxidase and Its Associated Activities in the Ovine Milk and Liver: Distinctive in Impact of in Vivo Molybdenum. Potravin. Slovak J. Food Sci. 2021, 15, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Kruse, T.; Gehl, C.; Geisler, M.; Lehrke, M.; Ringel, P.; Hallier, S.; Hänsch, R.; Mendel, R.R. Identification and Biochemical Characterization of Molybdenum Cofactor-Binding Proteins from Arabidopsis Thaliana*. J. Biol. Chem. 2010, 285, 6623–6635. [Google Scholar] [CrossRef]
- Kielkopf, C.L.; Bauer, W.; Urbatsch, I.L. Bradford Assay for Determining Protein Concentration. Cold Spring Harb. Protoc. 2020, 2020, 102269. [Google Scholar] [CrossRef] [PubMed]
- Savidov, N.A.; Alikulov, Z.A.; Lips, S.H. Identification of an Endogenous NADPH-Regenerating System Coupled to Nitrate Reduction in Vitro in Plant and Fungal Crude Extracts. Plant Sci. 1998, 133, 33–45. [Google Scholar] [CrossRef]
- Mendel, R.R.; Alikulov, Z.A.; Lvov, N.P.; Müller, A.J. Presence of the Molybdenum-Cofactor in Nitrate Reductase-Deficient Mutant Cell Lines of Nicotiana Tabacum. Mol. Gen. Genet. MGG 1981, 181, 395–399. [Google Scholar] [CrossRef]
- Mendel, R.R.; Hänsch, R. Molybdoenzymes and Molybdenum Cofactor in Plants. J. Exp. Bot. 2002, 53, 1689–1698. [Google Scholar] [CrossRef]
- Fischer, K.; Barbier, G.G.; Hecht, H.-J.; Mendel, R.R.; Campbell, W.H.; Schwarz, G. Structural Basis of Eukaryotic Nitrate Reduction: Crystal Structures of the Nitrate Reductase Active Site. Plant Cell 2005, 17, 1167–1179. [Google Scholar] [CrossRef]
- Balotf, S.; Islam, S.; Kavoosi, G.; Kholdebarin, B.; Juhasz, A.; Ma, W. How Exogenous Nitric Oxide Regulates Nitrogen Assimilation in Wheat Seedlings under Different Nitrogen Sources and Levels. PLoS ONE 2018, 13, e0190269. [Google Scholar] [CrossRef]
- Campbell, W.H. Structure and Function of Eukaryotic NAD(P)H:Nitrate Reductase. Cell. Mol. Life Sci. CMLS 2001, 58, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Sakihama, Y. Simultaneous Production of Nitric Oxide and Peroxynitrite by Plant Nitrate Reductase: In Vitro Evidence for the NR-Dependent Formation of Active Nitrogen Species. FEBS Lett. 2000, 468, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Dyussembayev, K.A.; Kussainova, A.A.; Kulutaeva, M.S.; Alikulov, Z. Nitrate- and Nitrite Reducing Activity of Xanthine Oxidase in Goat, Camel and Horse Milk. Eur. J. Nat. Hist. 2015, 5, 6–10. [Google Scholar]
- Bowman, D.C.; Paul, J.L. Foliar Absorption of Urea, Ammonium, and Nitrate by Perennial Ryegrass Turf. J. Am. Soc. Hortic. Sci. Jashs 1992, 117, 75–79. [Google Scholar] [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Sanz-Luque, E.; Tejada-Jiménez, M.; Llamas, Á.; Plouviez, M.; Guieysse, B.; Fernández, E.; Galván, A. Chlamydomonas Reinhardtii, an Algal Model in the Nitrogen Cycle. Plants 2020, 9, 903. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.M.; Huber, S.C. Post-translational Regulation of Nitrate Reductase: Mechanism, Physiological Relevance and Environmental Triggers. J. Exp. Bot. 2001, 52, 1981–1989. [Google Scholar] [CrossRef]
- Pattanayak, D.; Chatterjee, S.R. Light-Mediated Regulation of Nitrate Reductase in Higher Plants. J. Plant Biochem. Biotechnol. 1998, 7, 73–78. [Google Scholar] [CrossRef]
- Allègre, A.; Silvestre, J.; Morard, P.; Kallerhoff, J.; Pinelli, E. Nitrate Reductase Regulation in Tomato Roots by Exogenous Nitrate: A Possible Role in Tolerance to Long-Term Root Anoxia. J. Exp. Bot. 2004, 55, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Kurmanbayeva, A.; Bekturova, A.; Soltabayeva, A.; Oshanova, D.; Nurbekova, Z.; Srivastava, S.; Tiwari, P.; Dubey, A.K.; Sagi, M. Active O-Acetylserine-(Thiol) Lyase A and B Confer Improved Selenium Resistance and Degrade l-Cys and l-SeCys in Arabidopsis. J. Exp. Bot. 2022, 73, 2525–2539. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Hu, C.; Hussain, S.; Rana, M.S.; Riaz, M.; Afzal, J.; Aziz, O.; Elyamine, A.M.; Farag Ismael, M.A.; Sun, X. Molybdenum-Induced Effects on Photosynthetic Efficacy of Winter Wheat (Triticum Aestivum L.) under Different Nitrogen Sources Are Associated with Nitrogen Assimilation. Plant Physiol. Biochem. 2019, 141, 154–163. [Google Scholar] [CrossRef]
- Popova, E.; Titova, M.; Tynykulov, M.; Zakirova, R.P.; Kulichenko, I.; Prudnikova, O.; Nosov, A. Sustainable Production of Ajuga Bioactive Metabolites Using Cell Culture Technologies: A Review. Nutrients 2023, 15, 1246. [Google Scholar] [CrossRef]
- Sutula, M.Y.; Akbassova, A.Z.; Yergaliev, T.M.; Nurbekova, Z.A.; Mukiyanova, G.S.; Omarov, R.T. Endowing Plants with Tolerance to Virus Infection by Their Preliminary Treatment with Short Interfering RNAs. Russ. J. Plant Physiol. 2017, 64, 939–945. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aitlessov, K.; Zhumabekova, B.; Sagyndykov, U.; Tuyakbayeva, A.; Bitkeyeva, A.; Bazarbaeva, K.Z.; Mukhtarov, A.; Nurbekova, Z.; Satkanov, M.; Kulatayeva, M.; et al. Foliar Fertilization with Molybdate and Nitrate Up-Regulated Activity of Nitrate Reductase in Lemon Balm Leaves. Horticulturae 2023, 9, 1325. https://doi.org/10.3390/horticulturae9121325
Aitlessov K, Zhumabekova B, Sagyndykov U, Tuyakbayeva A, Bitkeyeva A, Bazarbaeva KZ, Mukhtarov A, Nurbekova Z, Satkanov M, Kulatayeva M, et al. Foliar Fertilization with Molybdate and Nitrate Up-Regulated Activity of Nitrate Reductase in Lemon Balm Leaves. Horticulturae. 2023; 9(12):1325. https://doi.org/10.3390/horticulturae9121325
Chicago/Turabian StyleAitlessov, Kurmet, Bibigul Zhumabekova, Utemurat Sagyndykov, Akmaral Tuyakbayeva, Aliya Bitkeyeva, Karlygash Zh. Bazarbaeva, Abilkhas Mukhtarov, Zhadyrassyn Nurbekova, Mereke Satkanov, Maral Kulatayeva, and et al. 2023. "Foliar Fertilization with Molybdate and Nitrate Up-Regulated Activity of Nitrate Reductase in Lemon Balm Leaves" Horticulturae 9, no. 12: 1325. https://doi.org/10.3390/horticulturae9121325
APA StyleAitlessov, K., Zhumabekova, B., Sagyndykov, U., Tuyakbayeva, A., Bitkeyeva, A., Bazarbaeva, K. Z., Mukhtarov, A., Nurbekova, Z., Satkanov, M., Kulatayeva, M., Aubakirova, K. M., Nurysheva, A., & Alikulov, Z. (2023). Foliar Fertilization with Molybdate and Nitrate Up-Regulated Activity of Nitrate Reductase in Lemon Balm Leaves. Horticulturae, 9(12), 1325. https://doi.org/10.3390/horticulturae9121325