


Analysis of Antioxidant Capacity Variation among Thai Holy Basil Cultivars (Ocimum tenuiflorum L.) Using Density-Based Clustering Algorithm

Abstract
:1. Introduction
2. Materials and Methods
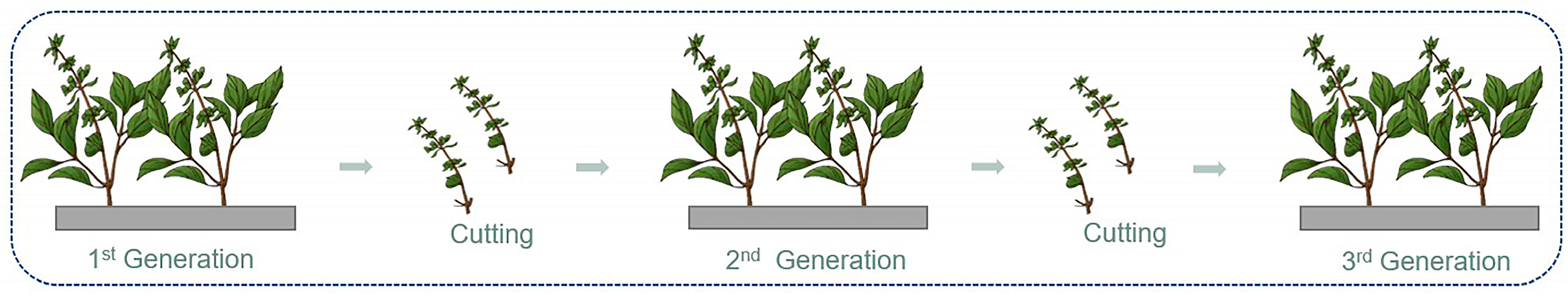
2.1. Plant Material and Growth Condition
2.2. Measurements of Biochemical Compounds
2.2.1. Sample Extraction
2.2.2. Anthocyanin Content
2.2.3. DPPH Free Radical Scavenging Activity
2.2.4. Flavonoid Content
2.2.5. Phenolics Content
2.2.6. Terpenoid Content
2.3. Statistical Analyses
2.4. OPTICS Clustering and Evaluation
3. Results
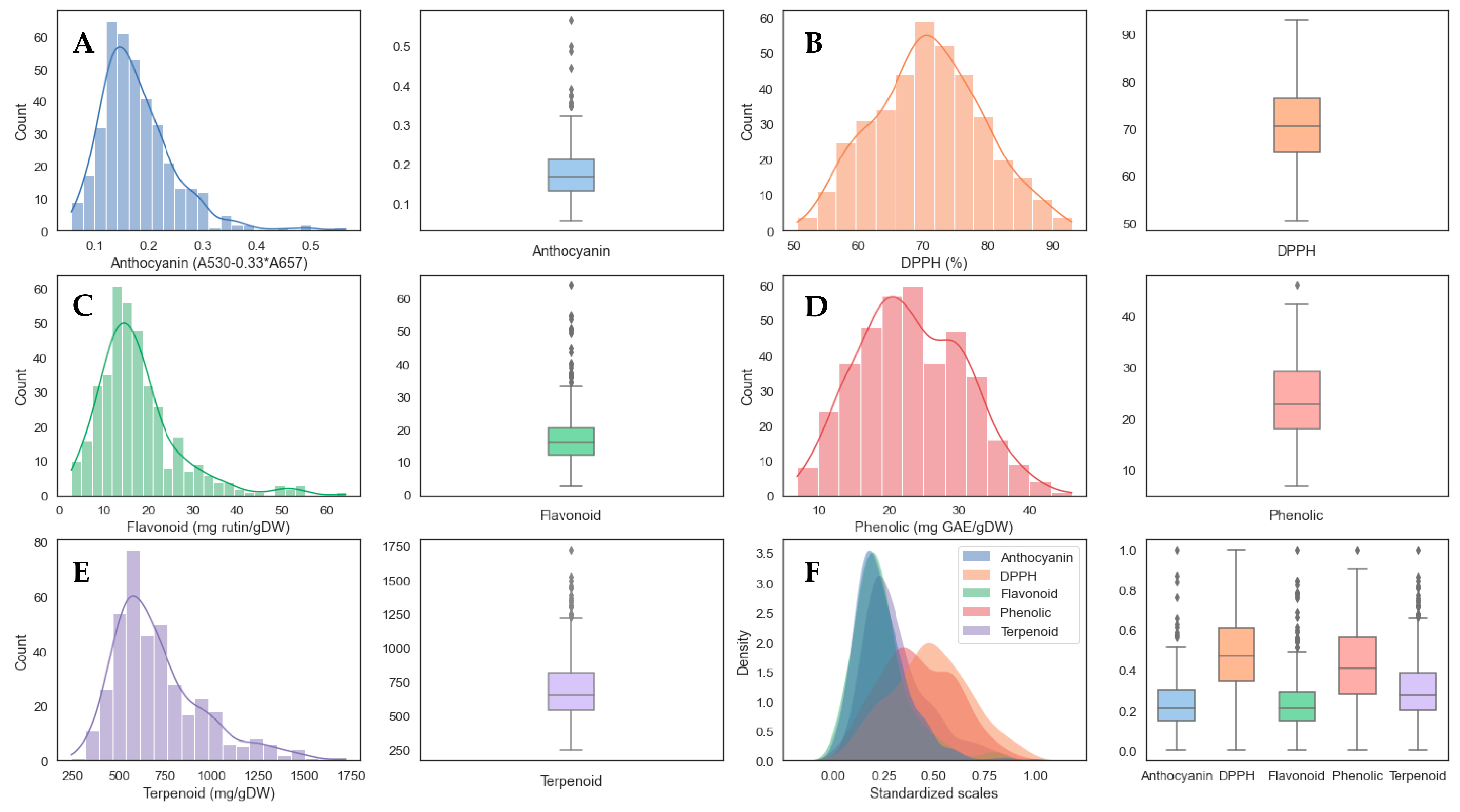
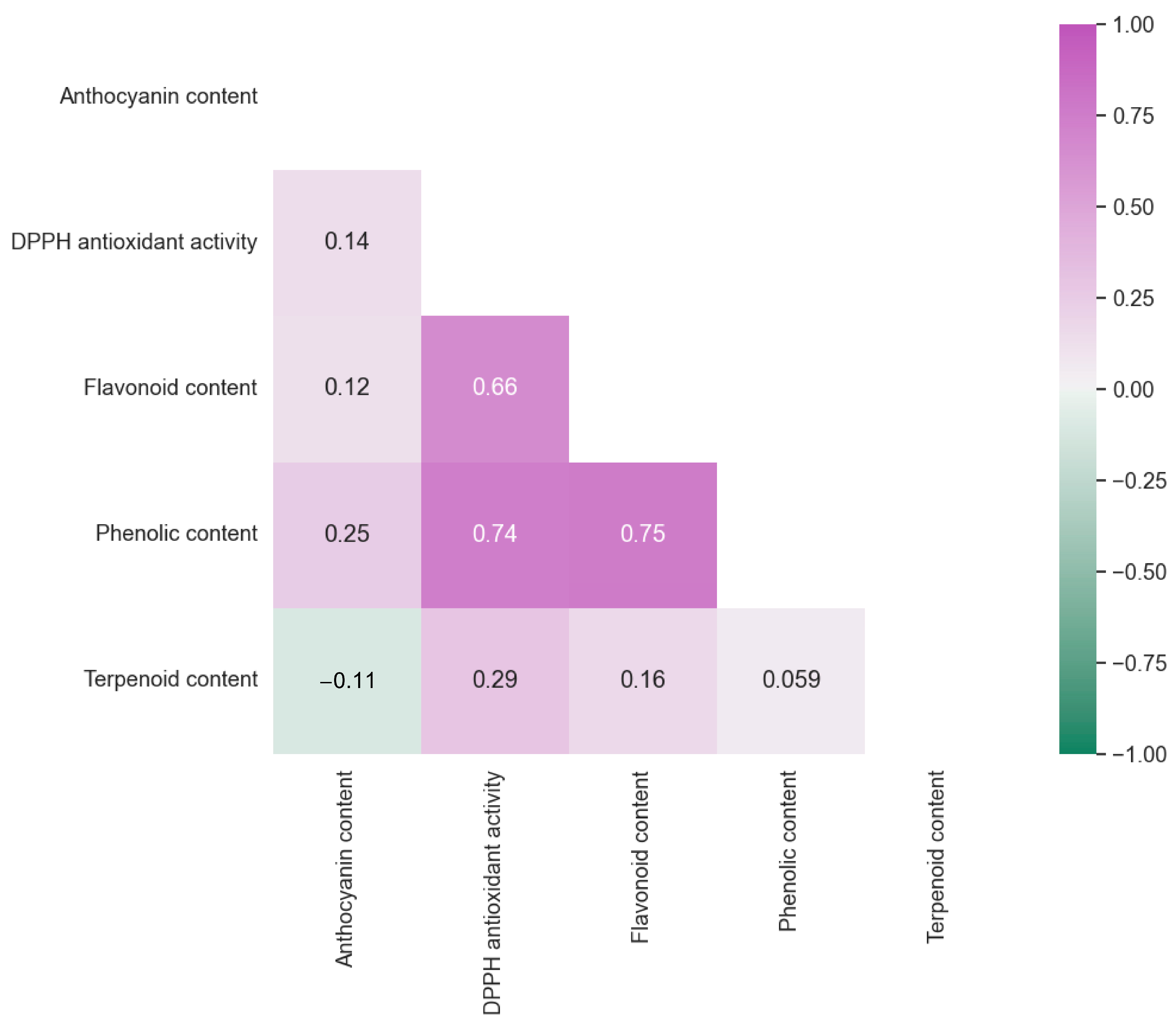
3.1. Exploration of Antioxidant Capacities and Distributions
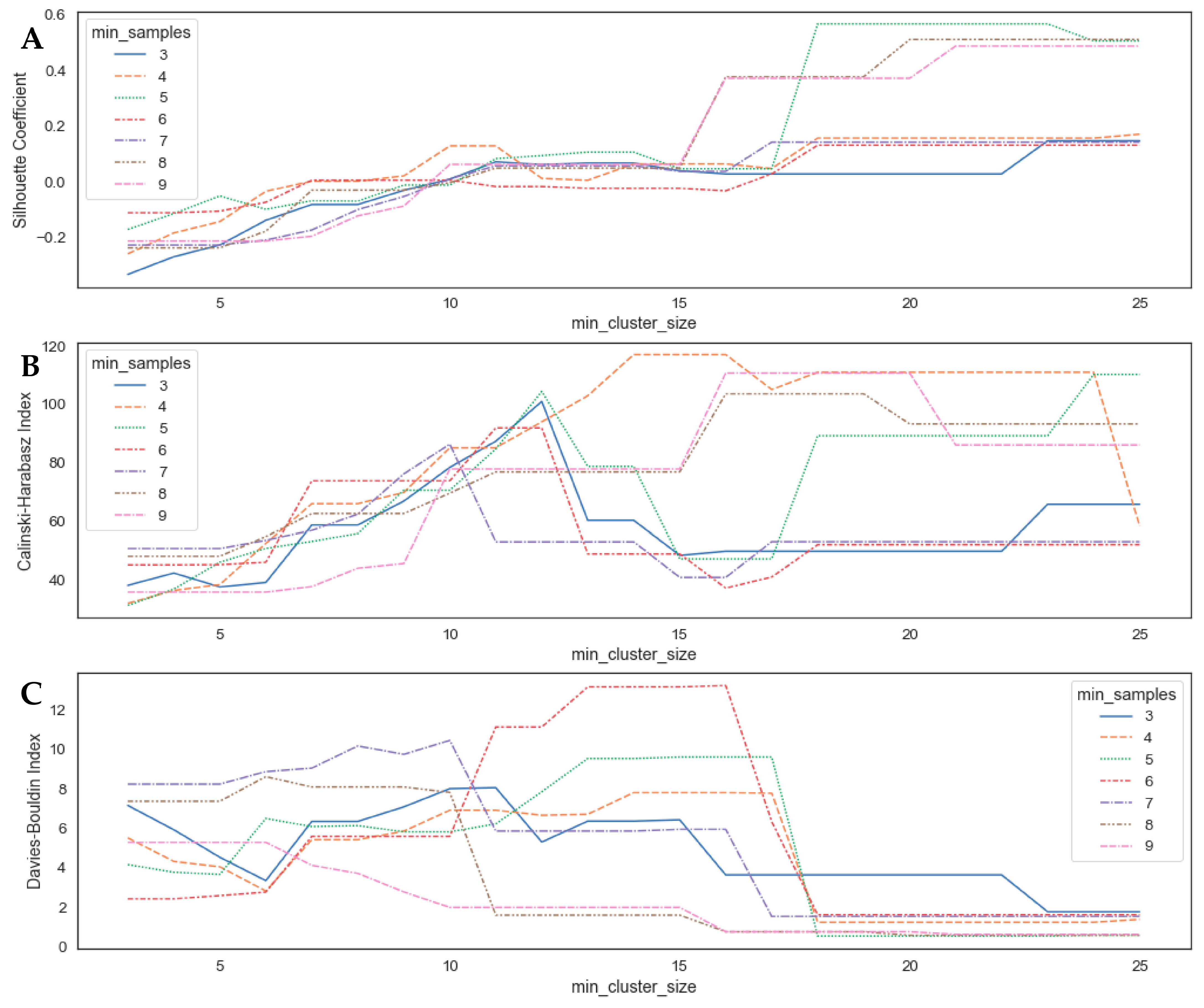
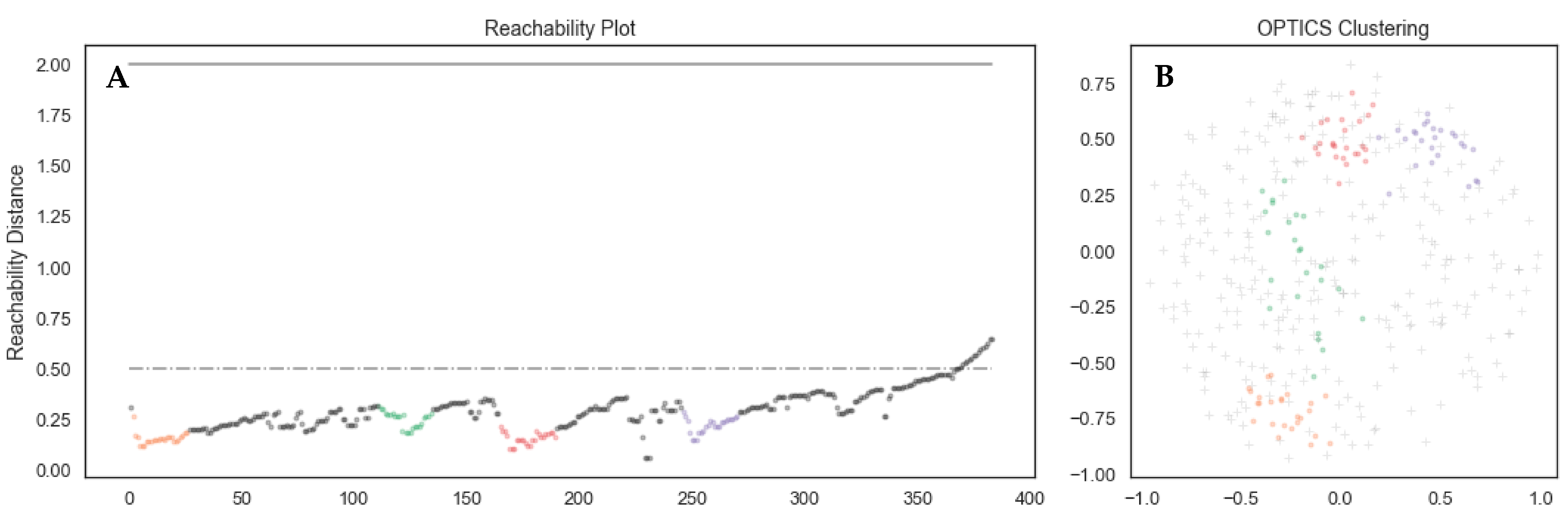
3.2. Parameter Optimization for the OPTICS Clustering Algorithm
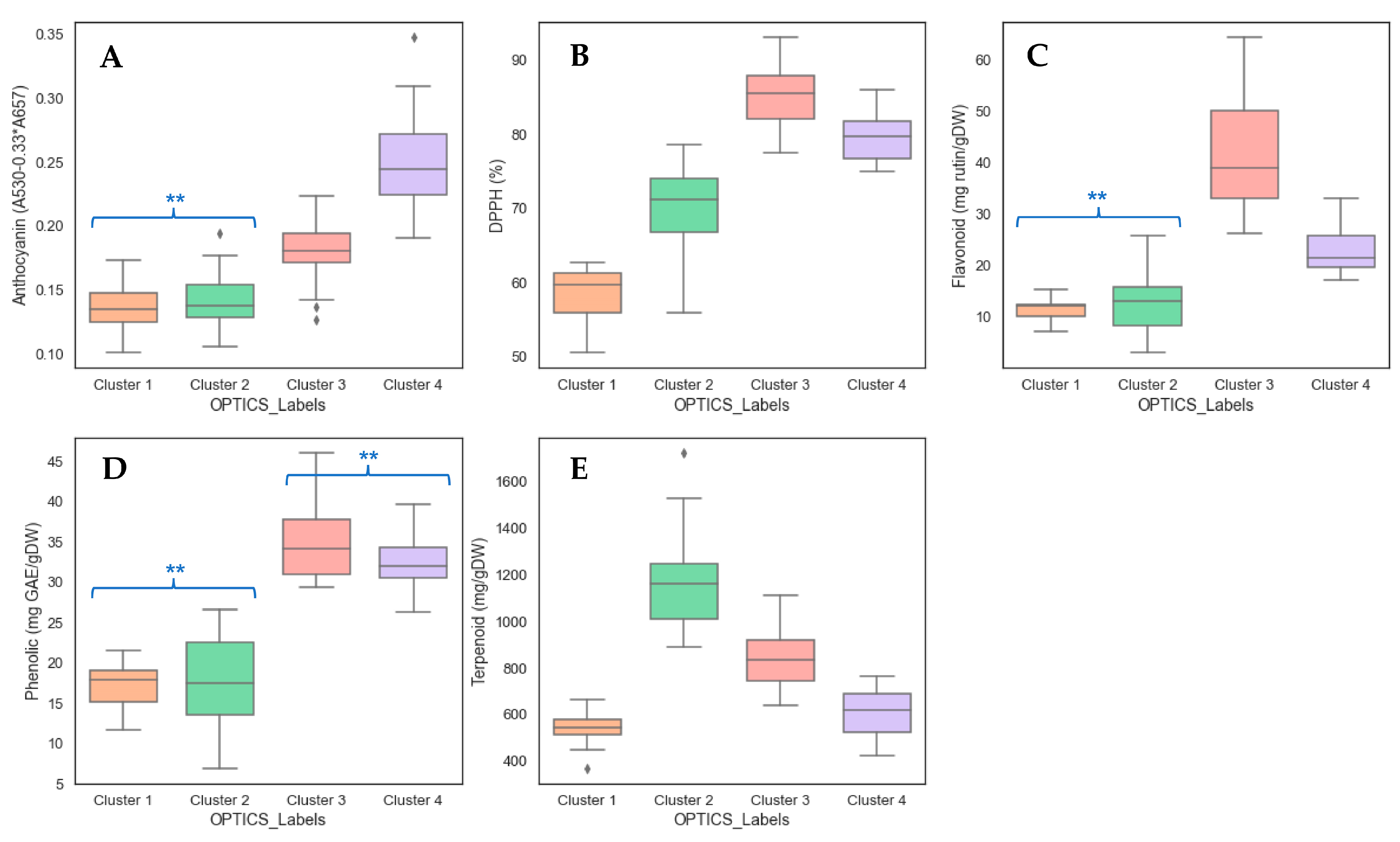
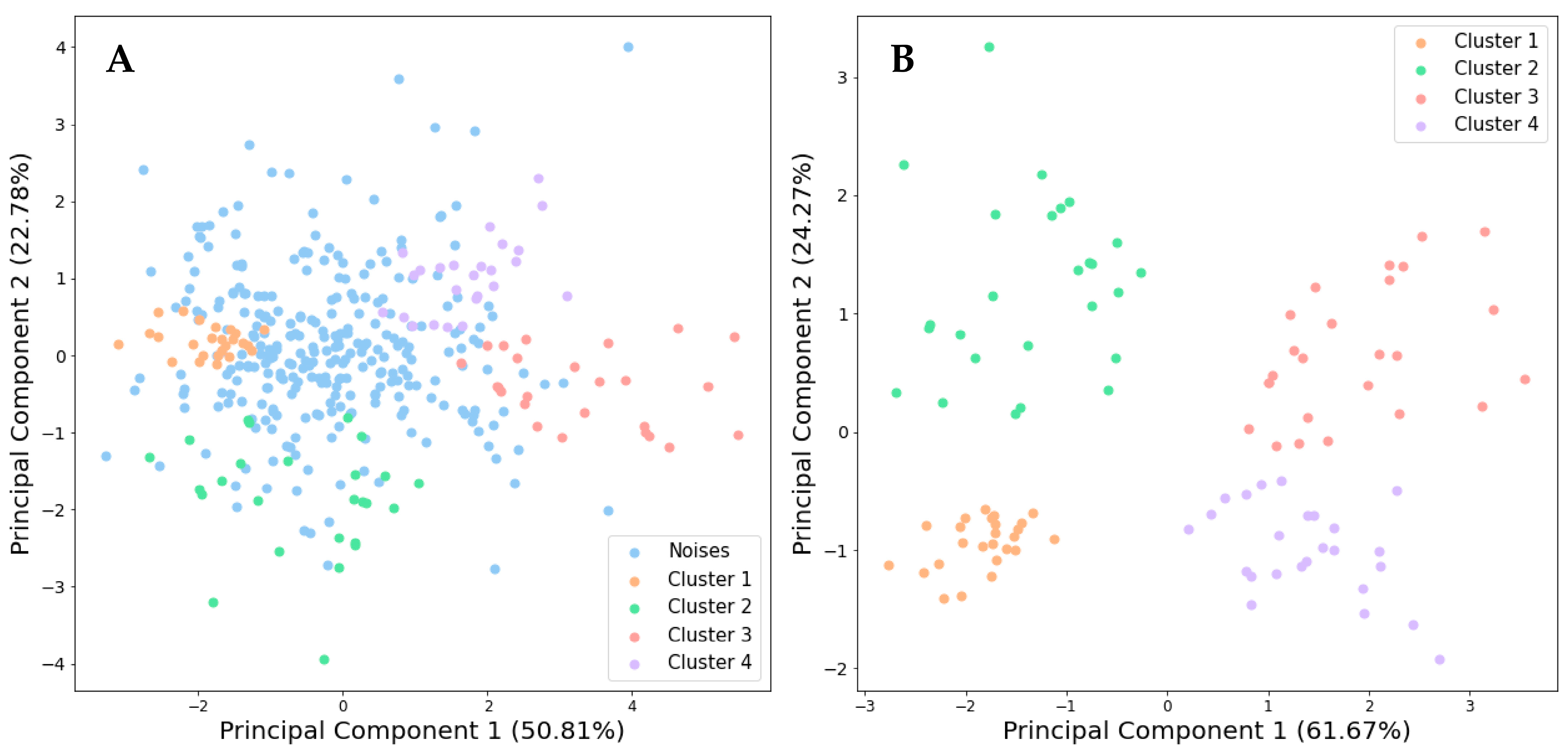
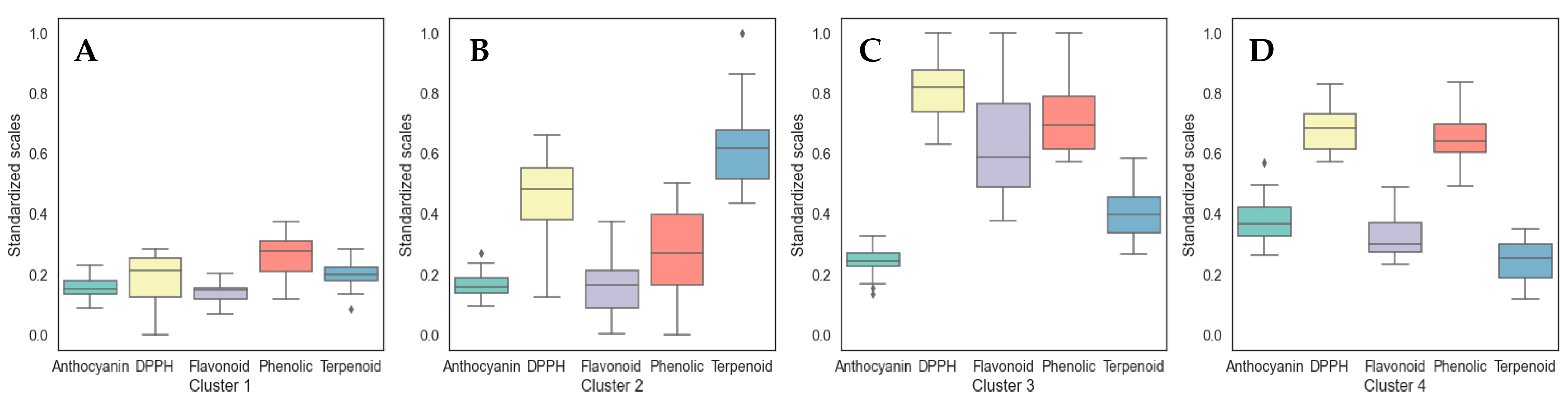
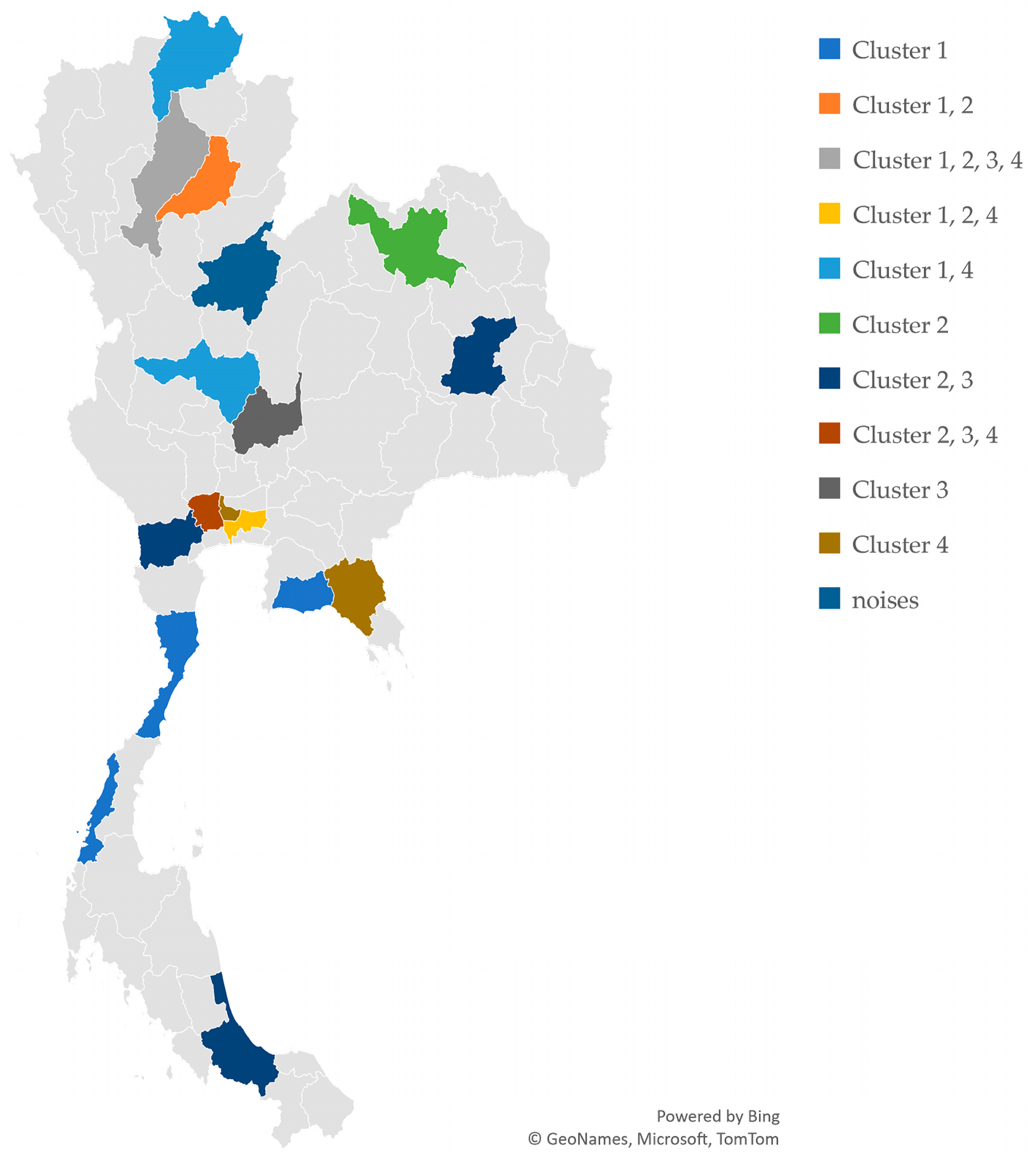
3.3. Clustering Analysis and Identification of Cultivar Groups in Holy Basil
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patwardhan, B.; Warude, D.; Pushpangadan, P.; Bhatt, N. Ayurveda and traditional Chinese medicine: A comparative overview. Evid.-Based Complement. Altern. Med. 2005, 2, 629537. [Google Scholar] [CrossRef]
- Singh, N.; Hoette, Y.; Miller, D.R. Tulsi: The Mother Medicine of Nature; International Institute of Herbal Medicine: Lucknow, India, 2002. [Google Scholar]
- Singh, S.; Taneja, M.; Majumdar, D.K. Biological activities of Ocimum sanctum L. fixed oil-an overview. Indian J. Exp. Biol. 2007, 45, 403–412. [Google Scholar] [PubMed]
- Deme, P.; Aluganti Narasimhulu, C.; Parthasarathy, S. Evaluation of anti-inflammatory properties of herbal aqueous extracts and their chemical characterization. J. Med. Food 2019, 22, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Chukwuma, I.F.; Uchendu, N.O.; Asomadu, R.O.; Ezeorba, W.F.C.; Ezeorba, T.P.C. African and Holy Basil—A review of ethnobotany, phytochemistry, and toxicity of their essential oil: Current trends and prospects for antimicrobial/anti-parasitic pharmacology. Arab. J. Chem. 2023, 16, 104870. [Google Scholar] [CrossRef]
- Mallikarjun, S.; Rao, A.; Rajesh, G.; Shenoy, R.; Pai, M. Antimicrobial efficacy of Tulsi leaf (Ocimum sanctum) extract on periodontal pathogens: An in vitro study. J. Indian Soc. Periodontol. 2016, 20, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Baliga, M.S.; Jimmy, R.; Thilakchand, K.R.; Sunitha, V.; Bhat, N.R.; Saldanha, E.; Rao, S.; Rao, P.; Arora, R.; Palatty, P.L. Ocimum sanctum L (holy basil or Tulsi) and Its phytochemicals in the prevention and treatment of cancer. Nutr. Cancer 2013, 65, 26–35. [Google Scholar] [CrossRef]
- Giridharan, V.V.; Thandavarayan, R.A.; Konishi, T. Chapter 98—Ocimum sanctum Linn. (Holy Basil) to Improve Cognition. In Diet and Nutrition in Dementia and Cognitive Decline; Martin, C.R., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 1049–1058. [Google Scholar]
- Joshi, H.; Parle, M. Cholinergic basis of memory improving effect of Ocimum tenuiflorum Linn. Indian J. Pharm. Sci. 2006, 68, p364–p365. [Google Scholar] [CrossRef]
- Singh, D.; Chaudhuri, P.K. A review on phytochemical and pharmacological properties of Holy basil (Ocimum sanctum L.). Ind. Crops Prod. 2018, 118, 367–382. [Google Scholar] [CrossRef]
- Al-Owaisi, M.; Al-Hadiwi, N.; Khan, S.A. GC-MS analysis, determination of total phenolics, flavonoid content and free radical scavenging activities of various crude extracts of Moringa peregrina (Forssk.) Fiori leaves. Asian Pac. J. Trop. Biomed. 2014, 4, 964–970. [Google Scholar] [CrossRef]
- Mahajan, N.; Singh, J.; Sinha, S. Comparison of total flavonoid, phenolic content and antioxidant capacity in leaf and seed extracts from white holy basil (Ocimum sanctum). Int. J. Appl. Biol. Pharm. Technol. 2014, 5, 34–42. [Google Scholar]
- Chutimanukul, P.; Wanichananan, P.; Janta, S.; Toojinda, T.; Darwell, C.T.; Mosaleeyanon, K. The influence of different light spectra on physiological responses, antioxidant capacity and chemical compositions in two holy basil cultivars. Sci. Rep. 2022, 12, 588. [Google Scholar] [CrossRef] [PubMed]
- Raina, A.P.; Kumar, A.; Dutta, M. Chemical characterization of aroma compounds in essential oil isolated from “Holy Basil” (Ocimum tenuiflorum L.) grown in India. Genet. Resour. Crop Evol. 2013, 60, 1727–1735. [Google Scholar] [CrossRef]
- Malav, P.; Pandey, A.; Bhatt, K.C.; Gopala Krishnan, S.; Bisht, I.S. Morphological variability in holy basil (Ocimum tenuiflorum L.) from India. Genet. Resour. Crop Evol. 2015, 62, 1245–1256. [Google Scholar] [CrossRef]
- Tangpao, T.; Chung, H.-H.; Sommano, S.R. Aromatic Profiles of Essential Oils from Five Commonly Used Thai Basils. Foods 2018, 7, 175. [Google Scholar] [CrossRef]
- Wangcharoen, W.; Morasuk, W. Antioxidant capacity and phenolic content of holy basil. Songklanakarin J. Sci. Technol. 2007, 29, 1407–1415. [Google Scholar]
- Chutimanukul, P.; Jindamol, H.; Thongtip, A.; Korinsak, S.; Romyanon, K.; Toojinda, T.; Darwell, C.T.; Wanichananan, P.; Panya, A.; Kaewsri, W.; et al. Physiological responses and variation in secondary metabolite content among Thai holy basil cultivars (Ocimum tenuiflorum L.) grown under controlled environmental conditions in a plant factory. Front. Plant Sci. 2022, 13, 1008917. [Google Scholar] [CrossRef]
- Wongpraneekul, A.; Havananda, T.; Luengwilai, K. Variation in aroma level of holy basil (Ocimum tenuiflorum L.) leaves is related to volatile composition, but not trichome characteristics. J. Appl. Res. Med. Aromat. Plants 2022, 27, 100347. [Google Scholar] [CrossRef]
- Saran, P.L.; Tripathy, V.; Saha, A.; Kalariya, K.A.; Suthar, M.K.; Kumar, J. Selection of superior Ocimum sanctum L. accessions for industrial application. Ind. Crops Prod. 2017, 108, 700–707. [Google Scholar] [CrossRef]
- Saran, P.L.; Patel, R.B. Field marker character for essential oil content in green herbage through leaf colour intensity in holy basil (Ocimum sanctum L.). Vegetos 2021, 34, 889–897. [Google Scholar] [CrossRef]
- Wang, M.; Cantrell, C.L.; Mathews, S.T.; Paudel, P.; Lee, J.; Mentreddy, S.R. Agronomy, chemical analysis, and antidiabetic activity of basil (Ocimum Species). ACS Food Sci. Technol. 2022, 2, 1243–1256. [Google Scholar] [CrossRef]
- Day, W.H.E.; Edelsbrunner, H. Efficient algorithms for agglomerative hierarchical clustering methods. J. Classif. 1984, 1, 7–24. [Google Scholar] [CrossRef]
- Hartigan, J.A.; Wong, M.A. Algorithm AS 136: A K-means clustering algorithm. J. R. Stat. Society. Ser. C Appl. Stat. 1979, 28, 100–108. [Google Scholar] [CrossRef]
- Park, H.-S.; Jun, C.-H. A simple and fast algorithm for K-medoids clustering. Expert Syst. Appl. 2009, 36, 3336–3341. [Google Scholar] [CrossRef]
- Guorong, X.; Wei, Z.; Peiqi, C. EM algorithms of Gaussian mixture model and hidden Markov model. In Proceedings of the Proceedings 2001 International Conference on Image Processing (Cat. No.01CH37205), Thessaloniki, Greece, 7–10 October 2001; Volume 141, pp. 145–148. [Google Scholar]
- Ester, M.; Kriegel, H.-P.; Sander, J.; Xu, X. A density-based algorithm for discovering clusters in large spatial databases with noise. In Proceedings of the KDD-96 Proceedings, Portland OR, USA, 2–4 August 1996; pp. 226–231. [Google Scholar]
- Zhao, L.; Zaki, M.J. TRICLUSTER: An effective algorithm for mining coherent clusters in 3D microarray data. In Proceedings of the Proceedings of the 2005 ACM SIGMOD International Conference on Management of Data, Baltimore, MD, USA, 14–16 June 2005; pp. 694–705. [Google Scholar]
- Hahsler, M.; Piekenbrock, M.; Doran, D. dbscan: Fast density-based clustering with R. J. Stat. Softw. 2019, 91, 1–30. [Google Scholar] [CrossRef]
- Ankerst, M.; Breunig, M.M.; Kriegel, H.-P.; Sander, J. OPTICS: Ordering points to identify the clustering structure. SIGMOD Rec. 1999, 28, 49–60. [Google Scholar] [CrossRef]
- Wu, C.; Chen, Y.; Dong, Y.; Zhou, F.; Zhao, Y.; Liang, C.J. VizOPTICS: Getting insights into OPTICS via interactive visual analysis. Comput. Electr. Eng. 2023, 107, 108624. [Google Scholar] [CrossRef]
- Prieto Santamaría, L.; García del Valle, E.P.; Zanin, M.; Hernández Chan, G.S.; Pérez Gallardo, Y.; Rodríguez-González, A. Classifying diseases by using biological features to identify potential nosological models. Sci. Rep. 2021, 11, 21096. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xiao, G.; Yang, B.; Ye, J.; Zhang, X.; Zheng, Y.; Chen, B. Instar determination for the tomato leafminer Tuta absoluta (Lepidoptera: Gelechiidae) using the density-based OPTICS clustering algorithm. Environ. Entomol. 2022, 51, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef]
- Alkhalifah, Y.; Phillips, I.; Soltoggio, A.; Darnley, K.; Nailon, W.H.; McLaren, D.; Eddleston, M.; Thomas, C.L.P.; Salman, D. VOCCluster: Untargeted metabolomics feature clustering approach for clinical breath gas chromatography/mass spectrometry data. Anal. Chem. 2020, 92, 2937–2945. [Google Scholar] [CrossRef]
- Thongtip, A.; Mosaleeyanon, K.; Korinsak, S.; Toojinda, T.; Darwell, C.T.; Chutimanukul, P.; Chutimanukul, P. Promotion of seed germination and early plant growth by KNO3 and light spectra in Ocimum tenuiflorum using a plant factory. Sci. Rep. 2022, 12, 6995. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Cai, Y.; Sun, M.; Wang, G.; Corke, H. Anthocyanins, flavonols, and free radical scavenging activity of chinese bayberry (Myrica rubra) extracts and Their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; He, J. Protective role of anthocyanins in plants under low nitrogen stress. Biochem. Biophys. Res. Commun. 2018, 498, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Sun, M.; Corke, H. Antioxidant activity of betalains from plants of the amaranthaceae. J. Agric. Food Chem. 2003, 51, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Duh, P.D. Scavenging effect of methanolic extracts of peanut hulls on free-radical and active-oxygen species. J. Agric. Food Chem. 1994, 42, 629–632. [Google Scholar] [CrossRef]
- Ghorai, N.; Chakraborty, S.; Gucchait, S.; Saha, S.K.; Biswas, S.K. Estimation of total Terpenoids concentration in plant tissues using a monoterpene, Linalool as standard reagent. Protoc. Exch. 2012, 1–5. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Caliński, T.; Harabasz, J. A dendrite method for cluster analysis. Commun. Stat. 1974, 3, 1–27. [Google Scholar] [CrossRef]
- Davies, D.L.; Bouldin, D.W. A cluster separation measure. IEEE Trans. Pattern Anal. Mach. Intell. 1979, PAMI-1, 224–227. [Google Scholar] [CrossRef]
- Patel, F.; Modi, N.R. Estimation of total phenolic content in selected varieties of Ocimum species grown in different environmental condition. J. Pharmacogn. Phytochem. 2018, 7, 144–148. [Google Scholar]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.Z.; Comin, C.H.; Casanova, D.; Bruno, O.M.; Amancio, D.R.; Costa, L.d.F.; Rodrigues, F.A. Clustering algorithms: A comparative approach. PLoS ONE 2019, 14, e0210236. [Google Scholar] [CrossRef]
- Gomez-Casati, D.F.; Zanor, M.I.; Busi, M.V. Metabolomics in Plants and Humans: Applications in the Prevention and Diagnosis of Diseases. BioMed Res. Int. 2013, 2013, 792527. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Liu, W.; Pan, Z.; Ling, K. Comparative Study of Hydrochemical Classification Based on Different Hierarchical Cluster Analysis Methods. Int. J. Environ. Res. Public Health 2020, 17, 9515. [Google Scholar] [CrossRef]
- Bhattacharjee, P.; Mitra, P. A survey of density based clustering algorithms. Front. Comput. Sci. 2020, 15, 151308. [Google Scholar] [CrossRef]
- Chen, X.; Shang, S.; Yan, F.; Jiang, H.; Zhao, G.; Tian, S.; Chen, R.; Chen, D.; Dang, Y. Antioxidant Activities of Essential Oils and Their Major Components in Scavenging Free Radicals, Inhibiting Lipid Oxidation and Reducing Cellular Oxidative Stress. Molecules 2023, 28, 4559. [Google Scholar] [CrossRef]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2005, 579, 200–213. [Google Scholar] [CrossRef]
- Kumar, V.; Roy, B.K. Population authentication of the traditional medicinal plant Cassia tora L. based on ISSR markers and FTIR analysis. Sci. Rep. 2018, 8, 10714. [Google Scholar] [CrossRef]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial plant compounds, extracts and essential oils: An updated review on their effects and putative mechanisms of action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential Oils: Sources of Antimicrobials and Food Preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Name | Location |
|---|---|---|
| 1 | Green | Chia Tai Co., Ltd., Bangkok, Thailand |
| 2 | OC057 | Damnoen Saduak, Ratchaburi, Thailand |
| 3 | OC059 | Kamphaeng Saen, Nakhon Pathom, Thailand |
| 4 | OC063 | Chai Badan, Lopburi, Thailand |
| 5 | OC064 | Roi Et, Thailand |
| 6 | OC072 | Wat Phleng, Ratchaburi, Thailand |
| 7 | OC081 | Bang Klam, Songkhla, Thailand |
| 8 | OC095 | Krabure, Ranong, Thailand |
| 9 | OC097 | Bangsaphan, Prachuap Khiri Khan, Thailand |
| 10 | OC099 | Bangsaphan, Prachuap Khiri Khan, Thailand |
| 11 | OC101 | Bangsaphan, Prachuap Khiri Khan, Thailand |
| 12 | OC102 | Bangsaphan, Prachuap Khiri Khan, Thailand |
| 13 | OC104 | Mueang Rayong, Rayong, Thailand |
| 14 | OC105 | Mueang Rayong, Rayong, Thailand |
| 15 | OC106 | Mueang Chanthaburi, Chanthaburi, Thailand |
| 16 | OC108 | Leam Sing, Chanthaburi, Thailand |
| 17 | OC113 | Mueang Rayong, Rayong, Thailand |
| 18 | OC130 | Mueang Lampang, Lampang, Thailand |
| 19 | OC133 | Den Chai, Phrae, Thailand |
| 20 | OC135 | Mueang Phitsanulok, Phitsanulok, Thailand |
| 21 | OC139 | Banphot Phisai, Nakhon Sawan, Thailand |
| 22 | OC141 | Mueang Phrae, Phrae, Thailand |
| 23 | OC148 | Mae Lao, Chiang Rai, Thailand |
| 24 | OC194 | Udon Thani, Thailand |
| 25 | OC195 | Kumphawapi, Udon Thani, Thailand |
| 26 | Red | BENJAMITR ENTERPRISE (1991) CO., LTD., Nonthaburi, Thailand |
| Antioxidant Capacities | Min | Max | Mean | Median | Standard Deviation |
|---|---|---|---|---|---|
| Anthocyanin A530 − (0.33 A657) | 0.056 | 0.565 | 0.178 | 0.165 | 0.07 |
| DPPH (%) | 50.568 | 92.974 | 70.961 | 70.598 | 8.361 |
| Flavonoid (mg rutin/gDW) | 2.764 | 64.184 | 17.583 | 15.807 | 9.333 |
| Phenolic (mg GAE/gDW) | 6.907 | 45.955 | 23.191 | 22.816 | 7.464 |
| Terpenoid (mg/gDW) | 243.259 | 1721.481 | 707.343 | 652.778 | 240.498 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saelao, T.; Chutimanukul, P.; Suratanee, A.; Plaimas, K. Analysis of Antioxidant Capacity Variation among Thai Holy Basil Cultivars (Ocimum tenuiflorum L.) Using Density-Based Clustering Algorithm. Horticulturae 2023, 9, 1094. https://doi.org/10.3390/horticulturae9101094
Saelao T, Chutimanukul P, Suratanee A, Plaimas K. Analysis of Antioxidant Capacity Variation among Thai Holy Basil Cultivars (Ocimum tenuiflorum L.) Using Density-Based Clustering Algorithm. Horticulturae. 2023; 9(10):1094. https://doi.org/10.3390/horticulturae9101094
Chicago/Turabian StyleSaelao, Tanapon, Panita Chutimanukul, Apichat Suratanee, and Kitiporn Plaimas. 2023. "Analysis of Antioxidant Capacity Variation among Thai Holy Basil Cultivars (Ocimum tenuiflorum L.) Using Density-Based Clustering Algorithm" Horticulturae 9, no. 10: 1094. https://doi.org/10.3390/horticulturae9101094
APA StyleSaelao, T., Chutimanukul, P., Suratanee, A., & Plaimas, K. (2023). Analysis of Antioxidant Capacity Variation among Thai Holy Basil Cultivars (Ocimum tenuiflorum L.) Using Density-Based Clustering Algorithm. Horticulturae, 9(10), 1094. https://doi.org/10.3390/horticulturae9101094