
The Xenia Effect Promotes Fruit Quality and Assists in Optimizing Cross Combinations in ‘O’Neal’ and ‘Emerald’ Blueberry

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
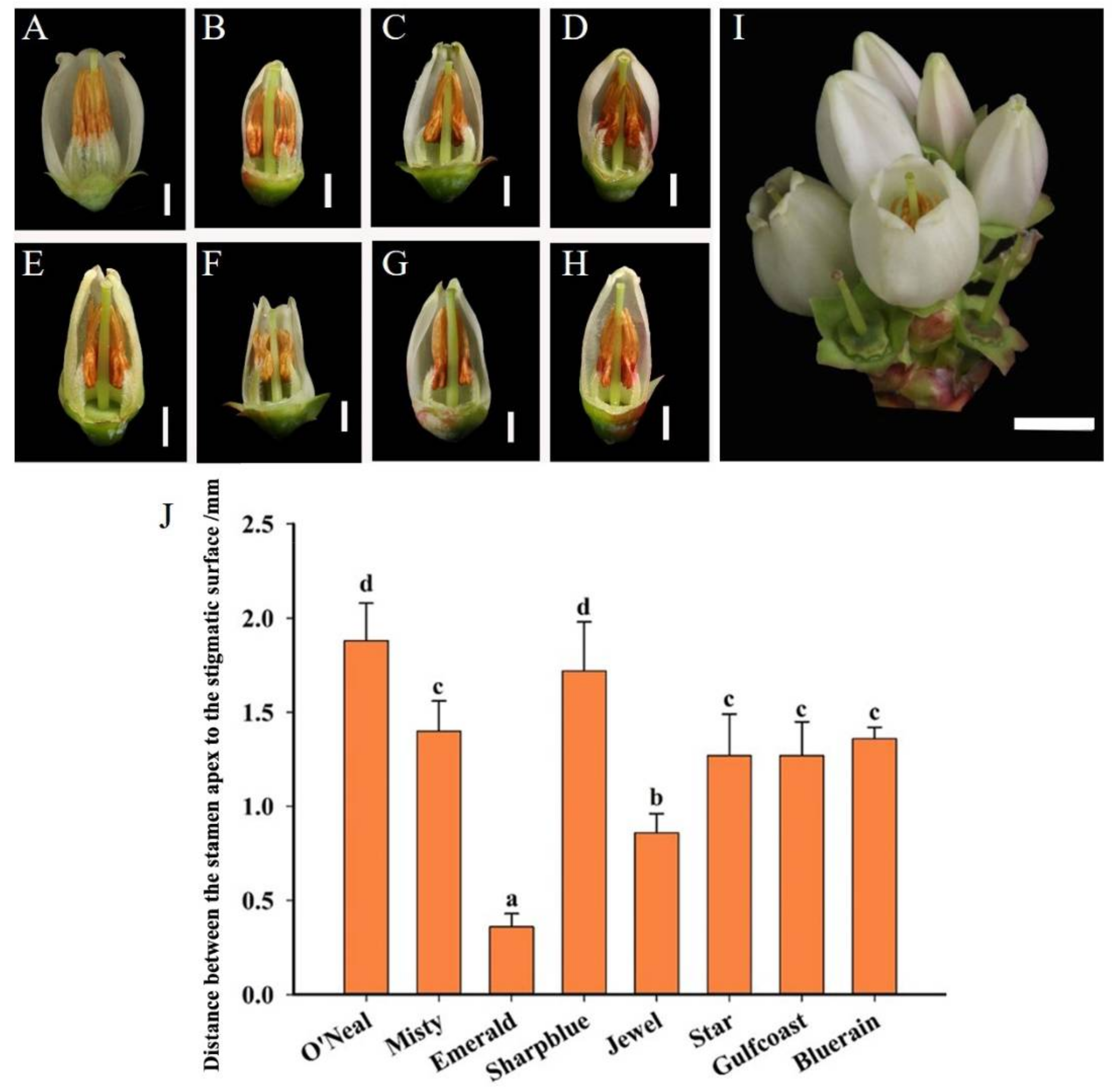
2.2. Morphological Observation of Floral Organs
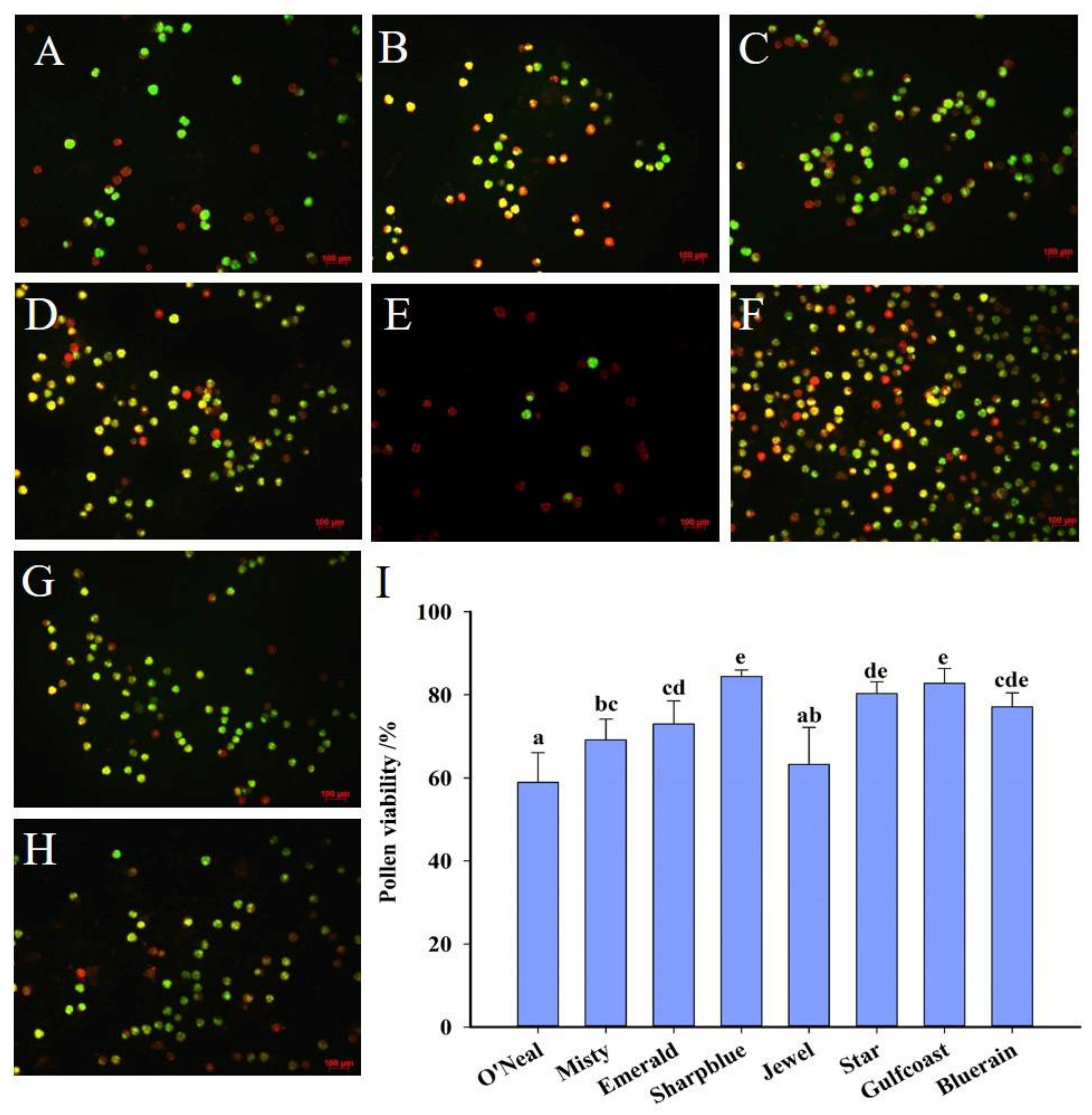
2.3. Pollen Viability and Stigma Receptivity Tests
2.4. Experimental Design and Pollination Treatments
2.5. Evaluation of the Fruit Quality of ‘O’Neal’ and ‘Emerald’
2.6. Seed Number Determination
2.7. DNA Extraction
2.8. SSR
2.9. Statistical Analysis
3. Results
3.1. Flower Morphology
3.2. Pollen Viability
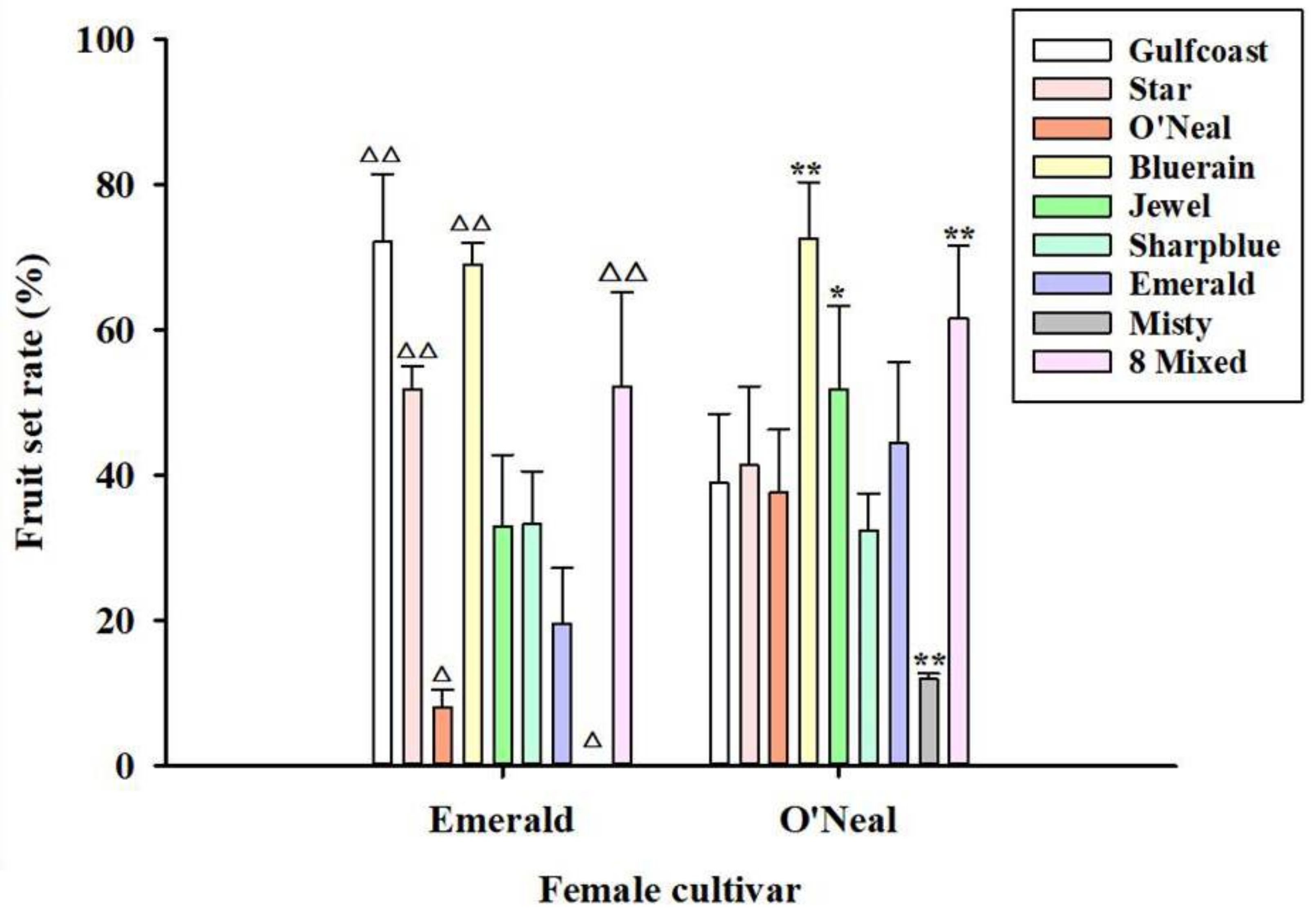
3.3. Xenia Effect on Fruit Set
3.4. Pollen Xenia Effects on the Fruit Quality
3.4.1. External Appearance
3.4.2. Interior Fruit Quality
3.4.3. Correlation Analysis between Fruit Set Rate and Fruit Quality
3.4.4. Principal Component Analysis of the Blueberry Fruit Quality
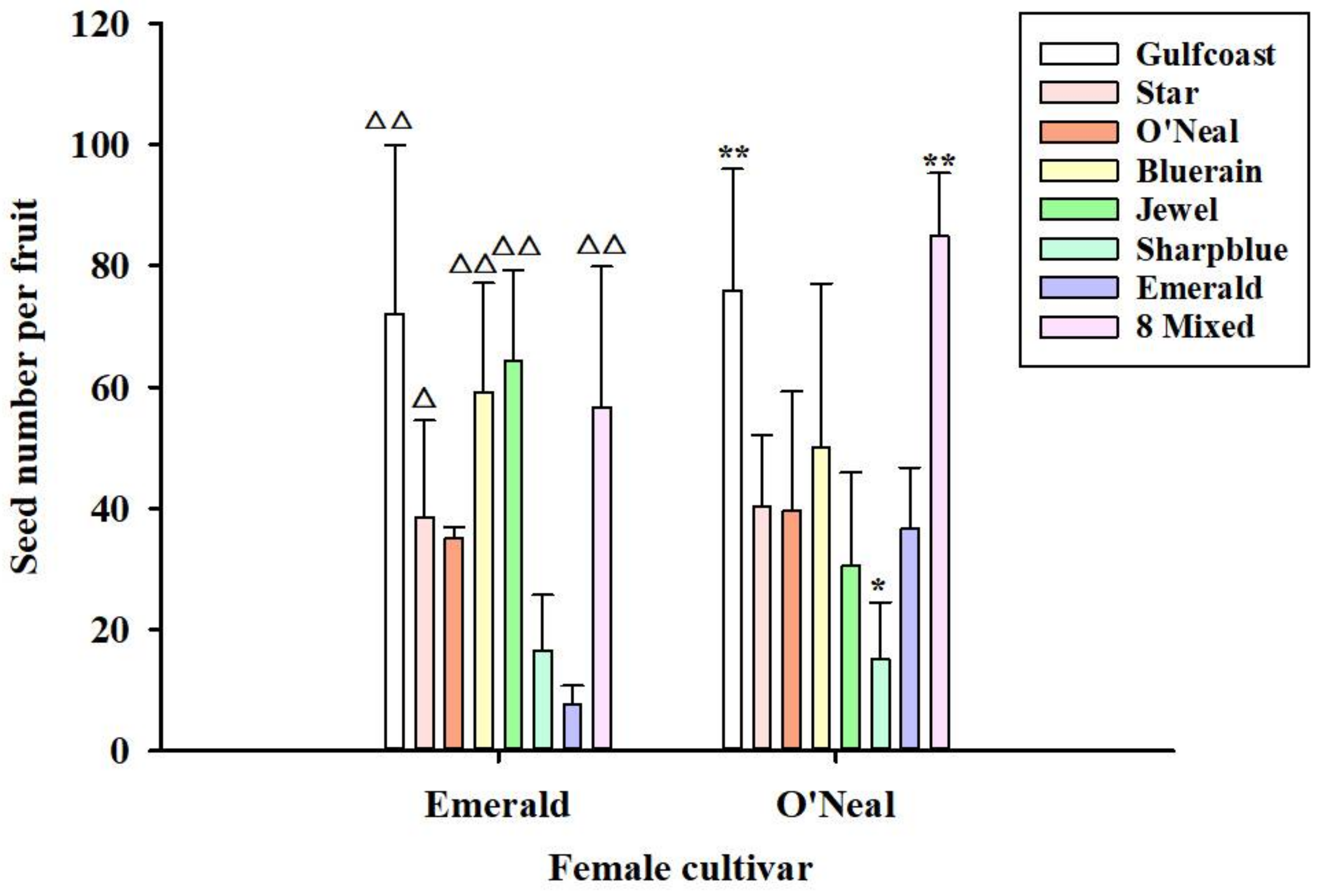
3.4.5. Seed Number
3.5. Paternal Preference Based on the SSR Analysis of Seedlings
4. Discussion
4.1. SHB Attains Higher Fruit Set, Better Fruit Quality, and More Seedlings with Optimal Cross Combination
4.2. SSR Can Help to Find the Most Efficient Hybrid Combination after Mixed Pollination
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACY | Anthocyanin Content |
| SHB | Southern Highbush Blueberry |
| FDA–PI | Fluorescein Diacetate–Propidium Iodide |
| SS/TA | Soluble Sugar Content/Titratable Acidity |
| FAM | Carboxyfluorescein |
| HEX | Hexachlorofluorescein |
| PCA | Principal Component Analysis |
References
- Qiu, Y.P.; Dai, H.F.; Li, Z.Q.; Ou, L.X.; Xiang, X.; Chen, J.Z.; Wang, B.X. Effects of pollinator on fruit quality of Guiwei litchi cultivar. J. Fruit Sci. 2006, 23, 703–706. [Google Scholar]
- Xie, H.; Gao, Q.M.; Liu, Y.J.; Nan, X.; Zhang, W.; Li, S.Q.; Li, J. Effects of xenia on fruit quality of Korla fragrant pear cultivar. Acta Agric. Boreali-Occident. Sin. 2013, 22, 93–96. [Google Scholar]
- Yu, L.Y.; Zuo, L.H.; Zhang, J.; Yang, M.S. Effect of xenia on fruit quality of 4 Malus sieversii clones. Mol. Plant Breed. 2017, 15, 3667–3675. [Google Scholar] [CrossRef]
- Zhang, X.J.; Yu, D.; Chen, Z.F.; Xu, L.; Wei, X.Q.; Xu, J.H. A preliminary discussion on pollen xenia of mango. Southeast Hortic. 2017, 5, 20–22. [Google Scholar]
- He, X.Y.; Tao, L.; Ni, S.B.; Chen, L.H.; Zhang, H.W.; Kong, G.H. Effects of pollen xenia on nut morphological characteristics and quality of ‘O.C’ cultivar in Macadamia spp. Nonwood For. Res. 2016, 34, 76–82. [Google Scholar] [CrossRef]
- Denney, J.O. Xenia includes metaxenia. HortScience 1992, 27, 722–728. [Google Scholar] [CrossRef]
- Pahlavani, M.H.; Abolhasani, K. Xenia effect on seed and embryo size in cotton (Gossypium hirsutum L.). J. Appl. Genet. 2006, 47, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Niu, L.; Zhang, Y.; Jin, M.; Ji, D.; Zhang, X. Pollen Sources Influence the Traits of Seed and Seed Oil in Paeonia ostii ‘Feng Dan’. HortScience 2017, 52, 700–705. [Google Scholar] [CrossRef]
- Yang, Q.; Fu, Y.; Liu, Y.; Zhang, T.; Peng, S.; Deng, J. Novel Classification Forms for Xenia. HortScience 2020, 55, 980–987. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. 2019, 11, 224–236. [Google Scholar] [CrossRef]
- Lang, G.A.; Danka, R.G. The influence of self-and cross-pollination on fruiting in southern highbush blueberries. HortScience 1991, 26, 486g. [Google Scholar] [CrossRef]
- Ehlenfeldt, M.K.; Kramer, M. Self-fertility Evaluations of Northern-adapted Rabbiteye Blueberry Hybrids. HortScience 2012, 47, 1837–1842. [Google Scholar] [CrossRef]
- Miller, S.; Alspach, P.; Scalzo, J.; Meekings, J. Pollination of ‘Hortblue Petite’ Blueberry: Evidence of Metaxenia in a New Ornamental Home-garden Cultivar. HortScience 2011, 46, 1468–1471. [Google Scholar] [CrossRef]
- Taber, S.K.; Olmstead, J.W. Impact of cross- and self-pollination on fruit set, fruit size, seed number, and harvest timing among 13 southern highbush blueberry cultivars. HortTechnology 2016, 26, 213–219. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, J.L.; Luo, Y.; Liu, Y.L.; Zhang, T.T.; Peng, S. Effect of xenia on formation of key quality traits of premier rabbiteye. South China Fruits 2020, 49, 103–106. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, C.-T.; Sciarappa, W.; Wang, C.Y.; Camp, M.J. Fruit quality, antioxidant capacity, and flavonoid content of organically and conventionally grown blueberries. J. Agric. Food Chem. 2008, 56, 5788–5794. [Google Scholar] [CrossRef]
- Cho, H.Y.; Kadowaki, M.; Che, J.; Horiuchi, N.; Ogiwara, I. Plant morphological characteristics for year-round production and its fruit quality in response to different light wavelengths in blueberry. In Proceedings of the XIII International Symposium on Plant Bioregulators in Fruit Production, Chiba, Japan, 27–31 August 2018; pp. 201–210. [Google Scholar]
- Wang, S.Y.; Zhou, Q.; Zhou, X.; Wei, B.D.; Ji, S.J. The effect of ethylene absorbent treatment on the softening of blueberry fruit. Food Chem. 2018, 246, 286–294. [Google Scholar] [CrossRef]
- Zhang, X.C.; Zhu, Y.Q.; Wang, Y.N.; Luo, C.; Wang, X. Effects of different plant growth regulators on blueberry fruit quality. IOP Conf. Ser. Earth Environ. Sci. 2017, 81, 012038. [Google Scholar] [CrossRef]
- Moyer, R.A.; Hummer, K.E.; Finn, C.E.; Frei, B.; Wrolstad, R.E. Anthocyanins, phenolics, and antioxidant capacity in diverse small fruits: Vaccinium, rubus, and ribes. J. Agric. Food Chem. 2002, 50, 519–525. [Google Scholar] [CrossRef]
- Hancock, J.F.; Olmstead, J.W.; Itle, R.A.; Callow, P.W.; Neils-Kraft, S.; Wheeler, E.J.; Mangandi, J.; Sooriyapathirana, S.S.; Rowland, L.J.; Mackey, T.A. Performance of an elite, hybrid family of a northern × southern highbush cross (‘Draper’ × ‘Jewel’). Euphytica 2018, 214, 95. [Google Scholar] [CrossRef]
- Nagasaka, K.; Yamane, H.; Tao, R. Evaluation of the effects of pollination on fruit size and quality of highbush blueberry. Acta Hortic. 2021, 1312, 25–30. [Google Scholar] [CrossRef]
- Yang, B.; Ballington, J.; Raja, A.; Brouwer, C.; Reid, R.; Burke, M.; Wang, X.G.; Rowland, L.J.; Bassil, N.; Brown, A. Patterns of simple sequence repeats in cultivated blueberries (Vaccinium section Cyanococcus spp.) and their use in revealing genetic diversity and population structure. Mol. Breed. 2014, 34, 675–689. [Google Scholar] [CrossRef]
- Bassil, N.; Bidani, A.; Hummer, K.; Rowland, L.J.; Olmstead, J.; Lyrene, P.; Richards, C. Assessing genetic diversity of wild southeastern North American Vaccinium species using microsatellite markers. Genet. Resour. Crop Evol. 2018, 65, 939–950. [Google Scholar] [CrossRef]
- Bhatt, D.S.; Debnath, S.C. Genetic Diversity of Blueberry Genotypes Estimated by Antioxidant Properties and Molecular Markers. Antioxidants 2021, 10, 458. [Google Scholar] [CrossRef]
- Prodorutti, D.; Pertot, I.; Giongo, L.; Gessler, C. Highbush Blueberry: Cultivation, Protection, Breeding and Biotechnology. Eur. J. Plant Sci. Biotechnol. 2007, 1, 44–56. [Google Scholar]
- Shen, L.B.; Liang, W.Y.; Qu, J.H.; Xie, M.S.; Lei, P.J.; Liu, H.J. The viability determination of cyanobacteria double staining with fluorescein diacetate and propidium iodide. Environ. Chem. 2005, 24, 554–557. [Google Scholar]
- Dafni, A.; Maués, M.M. A rapid and simple procedure to determine stigma receptivity. Sex. Plant Reprod. 1998, 11, 177–180. [Google Scholar] [CrossRef]
- Wang, Y.S.; Luo, Z.S.; Du, R.X. Nitric oxide delays chlorophyll degradation and enhances antioxidant activity in banana fruits after cold storage. Acta Physiol. Plant. 2015, 37, 74–83. [Google Scholar] [CrossRef]
- Mtenga, A.B.; Kassim, N.; Lee, W.-G.; Shim, W.-B.; Yoon, Y.; Chung, D.-H. Resistance of Bacillus cereus and its enterotoxin genes in ready-to-eat foods to γ -irradiation. Food Sci. Biotechnol. 2012, 21, 443–452. [Google Scholar] [CrossRef]
- Guan, A.Q.; Feng, W.M.; Lu, Y.Y.; Chen, G.; Han, Y.Q. Research on related factors influencing content of souble solids in tomato. Acta Agric. Jiangxi 2017, 29, 27–31. [Google Scholar] [CrossRef]
- Dogterom, M.H.; Winston, M.L.; Mukai, A. Effect of pollen load size and source (self, outcross) on seed and fruit production in highbush blueberry cv. ‘Bluecrop’ (Vaccinium Corymboum; Ericaceae). Am. J. Bot. 2000, 87, 1584–1591. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.S.; Li, F.; Xu, C.J.; Zhang, S.L.; Fu, C.X. An Efficient Macro-method of Genomic DNA Isolation from Actinidia chinensis Leaves. Hereditas 2004, 26, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Wang, Y.; Zhu, Y.Y.; Shao, X.; Li, Y.Q.; Guo, W.D. Development and validation of SSR markers based on transcriptomic data of Chinese cherry (Prunus pseudocerasus). Acta Hortic. Sin. 2016, 43, 1566–1576. [Google Scholar] [CrossRef]
- Rêgo, E.R.d.; Rêgo, M.M.d.; Cruz, C.D.; Finger, F.L.; Casali, V.W.D. Phenotypic diversity, correlation and importance of variables for fruit quality and yield traits in Brazilian peppers (Capsicum baccatum). Genet. Resour. Crop Evol. 2011, 58, 909–918. [Google Scholar] [CrossRef]
- Cadima, J.; Jolliffe, I.T. Loading and correlations in the interpretation of principle compenents. J. Appl. Stat. 1995, 22, 203–214. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Jones, A.G.; Small, C.M.; Paczolt, K.A.; Ratterman, N.L. A practical guide to methods of parentage analysis. Mol. Ecol. Resour. 2010, 10, 6–30. [Google Scholar] [CrossRef]
- Gündüz, K.; Serçe, S.; Hancock, J.F. Variation among highbush and rabbiteye cultivars of blueberry for fruit quality and phytochemical characteristics. J. Food Compos. Anal. 2015, 38, 69–79. [Google Scholar] [CrossRef]
- Silveira, T.M.T.d.; Raseira, M.d.C.B.; Nava, D.E.; Couto, M. Blueberry pollination in southern Brazil and their influence on fruit quality. Rev. Bras. Frutic. 2011, 33, 081–088. [Google Scholar] [CrossRef][Green Version]
- Courcelles, D.M.M.; Button, L.; Elle, E. Bee visit rates vary with floral morphology among highbush blueberry cultivars (Vaccinium corymbosum L.). J. Appl. Entomol. 2013, 137, 693–701. [Google Scholar] [CrossRef]
- Wang, Q.; Zheng, Y.F.; Yu, Y.B.; Gao, H.Y.; Lai, C.C.; Luo, X.L.; Huang, X.G. Effects of cross-pollination by ‘Murcott’ tangor on the physicochemical properties, bioactive compounds and antioxidant capacities of ‘Qicheng 52′ navel orange. Food Chem. 2019, 270, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Björkman, T. The effect of pollen load and pollen grain competition on fertilization success and progeny performance in Fagopyrum esculentum. Euphytica 1995, 83, 47–52. [Google Scholar] [CrossRef]
- Mulcahy, D.L.; Mulcahy, G.B. The influence of gametophytic competition on sporophytic quality in Dianthus chinensis. Theor. Appl. Genet. 1975, 46, 277–280. [Google Scholar] [CrossRef]
- Schlichting, C.D.; Stephenson, A.G.; Small, L.E.; Winsor, J.A. Pollen loads and progeny vigor in cucurbita pepo: The next generation. Evolution 1990, 44, 1358–1372. [Google Scholar] [CrossRef]
- Hormaza, J.I.; Herrero, M. Male gametophytic selection as a plant breeding tool. Sci. Hortic. 1996, 65, 321–333. [Google Scholar] [CrossRef]
- Hormaza, J.I.; Herrero, M. Dynamics of pollen tube growth under different competition regimes. Sex. Plant Reprod. 1996, 9, 153–160. [Google Scholar] [CrossRef]
- Tabbener, H.E.; Cottrell, J.E. The use of PCR based DNA markers to study the paternity of poplar seedlings. For. Ecol. Manag. 2003, 179, 363–376. [Google Scholar] [CrossRef]
- Han, Z.Q.; Gao, P.; Geng, X.N.; Du, K.; Kang, X.Y. Identification of the male parent of superior half-sib Populus tomentosa individuals based on SSR markers. Mol. Breed. 2017, 37, 155. [Google Scholar] [CrossRef]
- Lyrene, P.M.; Sherman, W.B. ‘Star’ Southern Highbush Blueberry. Hortscience 2000, 35, 956–957. [Google Scholar] [CrossRef]
- Ehlenfeldt, M.K. The genetic composition and tetrasomic inbreeding coefficients of highbush blueberry cultivars. HortScienc 1994, 29, 1342–1345. [Google Scholar] [CrossRef]
- Brevis, P.A.; Bassil, N.V.; Ballington, J.R.; Hancock, J.F. Impact of wide hybridization on highbush blueberry breeding. J. Am. Soc. Hortic. Sci. 2008, 133, 427–437. [Google Scholar] [CrossRef]
- Wu, X.W.; Cui, G.F.; Wu, L.F.; Zhang, Y.P.; Ming, J.; Wang, J.; Wang, J.H. Identification of ISSR in lily hybrids. Acta Hortic. Sin. 2009, 36, 749–754. [Google Scholar] [CrossRef]
- Fang, J.G.; Zhang, Z.; Ma, Z.Q.; Liu, D.J.; Wang, S.H.; Lavi, U. The polymorphism and segregation patterns of AFLP markers in the F1 progenies from the cross of two mango cultivars. Sci. Agric. Sin. 2000, 33, 19–24. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
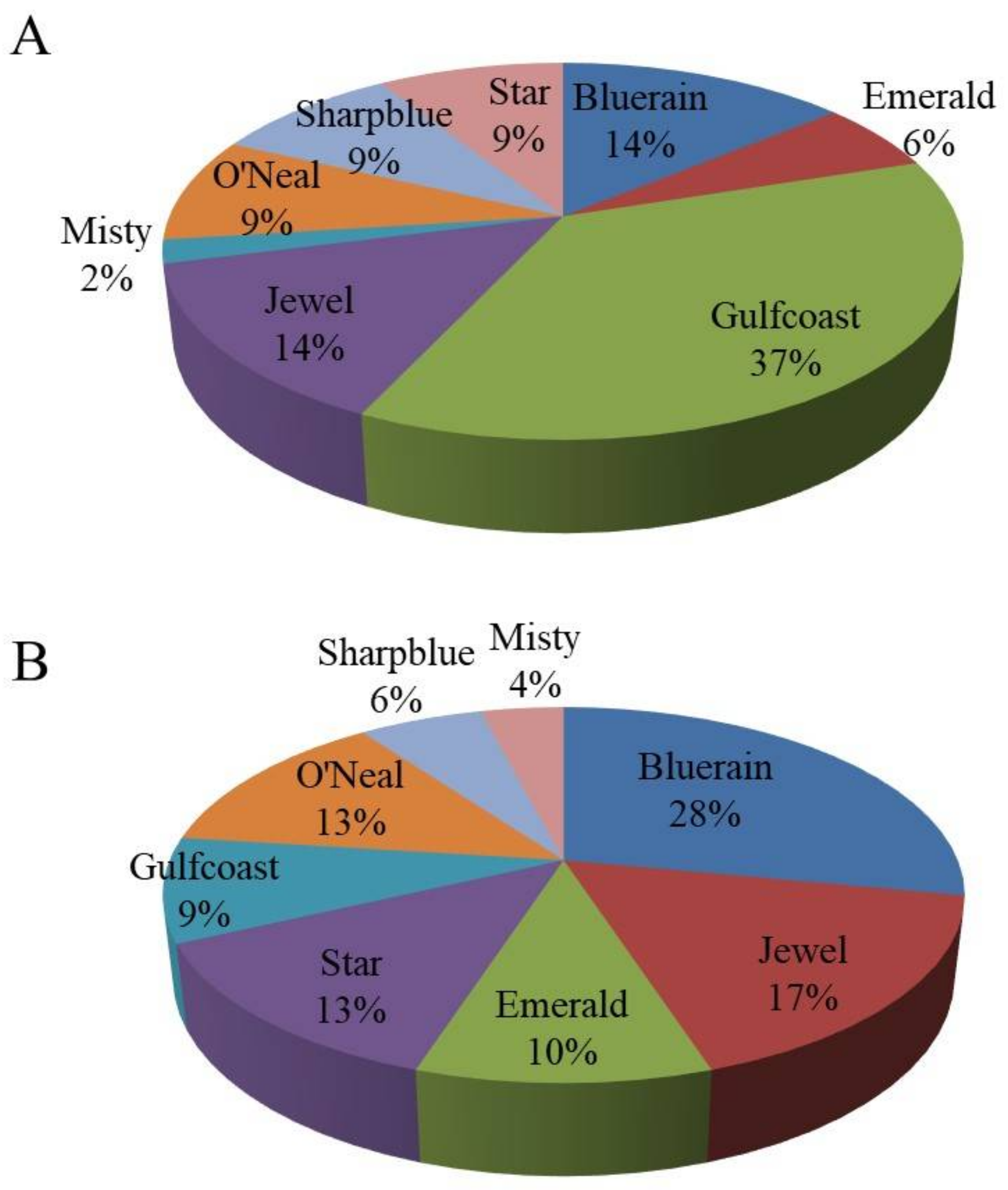{kind=link}
| Maternal Cultivar | Paternal Cultivar | Fruit Shape | Transverse Diameter (mm) | Longitudinal Diameter (mm) | Fruit Shape Index * | Fruit Weight (g/per Berry) | Firmness (×105 Pa) |
| Emerald | Emerald | oval | 13.8 ± 1.3 cd | 11.4 ± 0.8 bc | 0.8 ± 0.1 cd | 1.3 ± 0.4 cd | 4.4 ± 0.5 abc |
| Gulfcoast | oblateness | 17.1 ± 2.4 a | 13.0 ± 1.7 a | 0.8 ± 0.1 ab | 2.5 ± 0.8 a | 3.7 ± 0.5 c | |
| Star | oval | 14.3 ± 2.0 bc | 11.3 ± 1.6 bc | 0.8 ± 0.2 bc | 1.5 ± 0.5 bcd | 4.5 ± 0.7 ab | |
| O’Neal | subround | 12.3 ± 2.1 d | 10.4 ± 1.6 c | 0.9 ± 0.1 d | 0.9 ± 0.3 d | 4.3 ± 0.4 abc | |
| Bluerain | oval | 15.4 ± 2.0 bc | 11.9 ± 1.5 ab | 0.8 ± 0.1 bc | 1.9 ± 0.6 b | 4.0 ± 1.0 bc | |
| Jewel | oblateness | 14.4 ± 2.0 bd | 10.8 ± 1.2 bc | 0.8 ± 0.1 ab | 1.5 ± 0.5 bcd | 4.8 ± 0.6 a | |
| Sharpblue | oblateness | 15.8 ± 2.5 ab | 11.2 ± 1.3 bc | 0.7 ± 0.1 a | 1.8 ± 0.8 bc | 4.1 ± 0.9 abc | |
| 8 Mixed | oblateness | 15.2 ± 2.0 ab | 11.2 ± 1.6 ab | 0.7 ± 0.1 cd | 1.8 ± 0.6 bc | 4.4 ± 1.0 abc | |
| Maternal cultivar | Paternal cultivar | Fruit shape | Transverse diameter (mm) | Longitudinal diameter (mm) | Fruit shape index * | Fruit weight (g/per berry) | Firmness |
| O’Neal | O’Neal | oval | 11.0 ± 1.3 d | 8.9 ± 1.3 b | 0.8 ± 0.1 b | 0.8 ± 0.3 d | 2.0 ± 0.5 d |
| Jewel | oblateness | 14.8 ± 2.4 a | 11.1 ± 1.4 a | 0.8 ± 0.1 a | 1.7 ± 0.7 a | 2.2 ± 0.8 cd | |
| Emerald | oval | 14.2 ± 1.3 ab | 11.2 ± 1.2 a | 0.8 ± 0.1 ab | 1.6 ± 0.4 ab | 2.3 ± 0.6 cd | |
| Sharpblue | oval | 12.8 ± 1.6 d | 10.5 ± 1.0 a | 0.8 ± 0.1 b | 1.1 ± 0.4 cd | 2.8 ± 0.5 abc | |
| Bluerain | oval | 13.7 ± 2.1 bc | 10.5 ± 1.3 a | 0.8 ± 0.2 ab | 1.3 ± 0.5 abc | 2.8 ± 0.5 abc | |
| Star | oval | 13.8 ± 1.6 bc | 11.0 ± 0.8 a | 0.8 ± 0.1 ab | 1.4 ± 0.5 abc | 2.5 ± 0.6 bcd | |
| Misty | oval | 13.9 ± 1.2 bc | 10.8 ± 0.7 a | 0.8 ± 0.0 ab | 1.6 ± 0.4 ab | 2.6 ± 0.3 bcd | |
| Gulfcoast | oval | 14.3 ± 2.3 bc | 11.2 ± 1.2 a | 0.8 ± 0.1 ab | 1.6 ± 0.6 a | 2.8 ± 0.9 abc | |
| 8 Mixed | oval | 13.7 ± 2.3 bc | 10.8 ± 1.5 b | 0.8 ± 0.1 ab | 1.3 ± 0.6 bcd | 3.2 ± 1.0 ab |
| Maternal Cultivar | Paternal Cultivar | Titratable Acidity (TA) (mg/g) | Anthocyanin Ccntent (mg/g) | Soluble Sugar (SS) (mg/g) | Soluble Solids (%) | SS/TA |
| Emerald | Emerald | 1.1 ± 0 bcd | 0.3 ± 0.2 d | 12.3 ± 2.2 c | 10.4 ± 2.0 a | 11.0 |
| Gulfcoast | 1.3 ± 0.2 a | 0.5 ± 0.0 cd | 64.2 ± 6.4 a | 11.9 ± 1.9 a | 50.9 | |
| Star | 1.1 ± 0.1 bcd | 0.7 ± 0.2 bc | 38.6 ± 2.7 b | 10.2 ± 2.0 a | 35.1 | |
| O’Neal | 1.2 ± 0.8 ab | 0.5 ± 0.2 cd | 10.8 ± 0.4 c | 11.1 ± 1.7 a | 9.2 | |
| Bluerain | 0.6 ± 0.0 d | 1.1 ± 0.0 a | 59.8 ± 3.8 a | 10.9 ± 1.9 a | 98.0 | |
| Jewel | 0.8 ± 0.1 bcd | 1.0 ± 0.2 bc | 16.0 ± 0.9 c | 10.9 ± 2.8 a | 19.5 | |
| Sharpblue | 0.7 ± 0.0 cd | 0.4 ± 0.0 d | 35.8 ± 4.5 b | 8.2 ± 0.7 b | 52.7 | |
| 8 Mixed | 0.9 ± 0.0 bcd | 1.3 ± 0.2 a | 33.0 ± 0.4 b | 10.4 ± 2.2 a | 37.1 | |
| Maternal cultivar | Paternal cultivar | Titratable acidity (TA) (mg/g) | Anthocyanin Content (mg/g) | Soluble sugar (SS) (mg/g) | Soluble solids (%) | SS/TA |
| O’Neal | O’Neal | 0.6 ± 0.2 a | 0.6 ± 0.0 e | 33.7 ± 14.0 bc | 15.1 ± 1.2 a | 53.5 |
| Jewel | 0.6 ± 0.0 a | 1.2 ± 0.2 cd | 41.3 ± 5.3 b | 12.2 ± 1.5 cd | 73.8 | |
| Emerald | 0.3 ± 0.1 b | 1.9 ± 0.1 ab | 24.8 ± 6.5 bc | 11.4 ± 1.9 bcd | 79.9 | |
| Sharpblue | 0.4 ± 0.1 b | 1.0 ± 0.1 d | 24.5 ± 5.4 bc | 11.7 ± 1.8 bc | 66.1 | |
| Bluerain | 0.4 ± 0.1 b | 1.2 ± 0.0 de | 51.7 ± 4.0 a | 12.8 ± 2.2 bc | 132.5 | |
| Star | 0.4 ± 0.1 b | 1.4 ± 0.2 c | 17.7 ± 6.4 c | 12.2 ± 2.1 bc | 44.3 | |
| Gulfcoast | 0.6 ± 0.1 a | 1.0 ± 0.1 de | 38.6 ± 4.8 ab | 11.1 ± 1.7 cd | 68.9 | |
| Misty | - | 1.3 ± 0.0 cd | - | 12.8 ± 0.6 bc | - | |
| 8 Mixed | 0.4 ± 0.0 b | 2.0 ± 0.3 a | 52.2 ± 5.6 a | 13.2 ± 2.4 a | 145.0 |
| Maternal Cultivar: Emerald | Jewel | Emerald | Sharpblue | Bluerain | Star | Gulfcoast | O’Neal | Misty |
| FAC1-1 | −0.52 | −0.77 | 0.48 | 1.04 | −0.29 | 1.40 | −1.33 | - |
| FAC2-1 | −0.88 | 0.60 | −1.20 | −0.72 | −0.04 | 1.57 | 0.67 | - |
| FAC3-1 | 0.69 | −0.59 | −1.74 | 1.41 | 0.13 | −0.20 | 0.30 | - |
| Comprehensive assessment index | −0.38 | −0.36 | −0.28 | 0.59 | −0.15 | 1.08 | −0.51 | - |
| Maternal cultivar: O’Neal | Jewel | Emerald | Sharpblue | Bluerain | Star | Gulfcoast | O’Neal | Misty |
| FAC1-1 | 0.60 | 0.77 | −0.46 | 0.06 | 0.25 | 0.54 | −2.28 | 0.52 |
| FAC2-1 | 0.80 | −0.16 | −0.21 | 1.32 | −0.51 | 0.76 | −0.08 | −1.93 |
| FAC3-1 | −1.71 | −0.15 | 1.66 | 0.61 | −0.07 | 0.59 | −0.74 | −0.19 |
| Comprehensive assessment index | 0.29 | 0.33 | −0.08 | 0.42 | −0.01 | 0.53 | −1.24 | −0.23 |
| SSR Locus | Number of Alleles | Size Length (bp) | Observed Heterozygosity | Expected Heterozygosity | Polymorphic Information |
|---|---|---|---|---|---|
| VcSSR11 | 10 | 308–348 | 0.69 | 0.62 | 0.55 |
| VcSSR14 | 4 | 184–206 | 0.74 | 0.57 | 0.51 |
| VcSSR19 | 5 | 364–430 | 0.14 | 0.19 | 0.17 |
| VcSSR28 | 7 | 279–315 | 0.40 | 0.68 | 0.62 |
| VcSSR35 | 4 | 203–212 | 0.54 | 0.60 | 0.53 |
| VcSSR47 | 4 | 229–244 | 0.88 | 0.56 | 0.48 |
| Average | 5.6 | - | 0.57 | 0.54 | 0.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Xu, J.; Wang, Y.; Li, K.; Zong, Y.; Yang, L.; Chen, W.; Liao, F.; Guo, W. The Xenia Effect Promotes Fruit Quality and Assists in Optimizing Cross Combinations in ‘O’Neal’ and ‘Emerald’ Blueberry. Horticulturae 2022, 8, 659. https://doi.org/10.3390/horticulturae8070659
Liu J, Xu J, Wang Y, Li K, Zong Y, Yang L, Chen W, Liao F, Guo W. The Xenia Effect Promotes Fruit Quality and Assists in Optimizing Cross Combinations in ‘O’Neal’ and ‘Emerald’ Blueberry. Horticulturae. 2022; 8(7):659. https://doi.org/10.3390/horticulturae8070659
Chicago/Turabian StyleLiu, Jinlian, Jinjian Xu, Yujing Wang, Ke Li, Yu Zong, Li Yang, Wenrong Chen, Fanglei Liao, and Weidong Guo. 2022. "The Xenia Effect Promotes Fruit Quality and Assists in Optimizing Cross Combinations in ‘O’Neal’ and ‘Emerald’ Blueberry" Horticulturae 8, no. 7: 659. https://doi.org/10.3390/horticulturae8070659
APA StyleLiu, J., Xu, J., Wang, Y., Li, K., Zong, Y., Yang, L., Chen, W., Liao, F., & Guo, W. (2022). The Xenia Effect Promotes Fruit Quality and Assists in Optimizing Cross Combinations in ‘O’Neal’ and ‘Emerald’ Blueberry. Horticulturae, 8(7), 659. https://doi.org/10.3390/horticulturae8070659