Abstract
TCP transcription factors are a unique transcription family in higher plants, and play important roles in plant development, responses to environmental stresses and phytohormones. Radish is an important crop and widely cultivated worldwide. However, genome-wide identification and expression analysis of TCP family in radish has not been reported yet. In this study, 32 RsTCP genes were identified from the whole genome. Phylogenetic analysis showed that the RsTCP family can be divided into two major groups and three subgroups, namely Class I (PCF), Class II (CIN), and Class II (CYC/TB1). Chromosome mapping showed that 32 genes were distributed on all nine chromosomes of radish. Transcriptome data indicated that the transcription of RsTCP genes differed between tissues and developmental stages. Multiple phytohormone-related cis-acting elements in the promoter region of RsTCPs and several post-translational modification sites in RsTCP protein sequences were identified. The real-time quantitative PCR (RT-qPCR) analysis indicated that several RsTCP genes changed significantly in response to the treatments, including low temperature, drought, salt, gibberellin (GA3), and abscisic acid. Among them, RsTCP16 showed significantly higher levels in leaves than in other radish tissues, and the transcription of RsTCP16 was significantly upregulated under the five treatments, especially during low temperature, salt shock, and GA3 response, suggesting that RsTCP16 could be involved in the development of radish leaves and the response to stress. In summary, these results provided a basis for further exploring the molecular mechanism of RsTCP genes in radish.
1. Introduction
Plant transcription factors play key roles in regulating the initiation of transcription of specific genes, which are crucial for development and responses to environmental changes in plants [1,2]. TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR1 (TCP) transcription factors are a unique family in higher plants, because they are only detected in plants and are involved in physiological processes, including cell differentiation, seed germination, flowering, and phytohormone response in higher plants [3,4,5,6]. TCP is named using the acronyms of the three isolated members, including TEOSINTE BRANCHED 1 (TB1) in Zea mays, CYCLOIDEA (CYC) in Antirrhinum majus, PROLIFERATING CELL FACTORS 1 and 2 (PCF1 and PCF2) in Oryza sativa [7]. With the completion of genome sequencing of plant species, many TCP transcription factors have been identified and analyzed in recent decades. Up to date, a total of 24, 30, 27, 23, 39, 75, 21, and 27 TCP members have been identified in Arabidopsis thaliana [8], Solanum lycopersicum [9], Citrullus lanatus [10], Phalaenopsis equestris [11], Brassica rapa ssp. rapa [12], Gossypium barbadense [13], Oryza sativa [14], and Cucumis sativus [15], respectively.
There is a conserved TCP domain that is important for DNA binding existing in TCP transcription factors. Although the TCP domain has a N-terminal basic helix–loop–helix (bHLH) structure, there are significant differences between the TCP and bHLH transcription factors [7]. Based on homology analysis of amino acid sequences of the conserved TCP domain, TCP transcription factors can be classified into two classes, namely Class I (PCF or TCP-P) and Class II (TCP-C) [8]. Distinctly different from the Class I, which contains a non-canonical bHLH motif, the Class II has four more amino acids in the conserved domain [7,16,17]. In addition, Class II members can be further subdivided into CIN and CYC/TB1 branches based on the sequence differences in the TCP domain [18,19].
Previous studies revealed that the TCP transcription factors play important roles in plant growth, development, and responses to biotic and abiotic stresses in A. thaliana [20]. For example, AtTCP4 regulates jasmonic acid (JA) biosynthesis through interaction with the JGB gene, which mediates the pollen germination and gametophyte development [21]. MPK8 can interact with AtTCP14 in the nucleus and phosphorylate AtTCP14 outside the nucleus, promoting seed germination [3]. The TCP transcription factors (Class II) can be used in combination with the FLOWERING LOCUS T (FT)-FD complex, which further acts on the AP1 promoter and actively regulates flowering [4]. TCP transcription factors inhibit the formation of cotyledon epidermal hair by inhibiting the activation of the MYB–bHLH–WD40 (MBW) complex at both transcription and protein levels [5]. In addition, TCP transcription factors are regulated by auxin, gibberellin, brassinolide, and other hormones in A. thaliana [6,22,23]. Under adverse environments, TCP transcription factors function in plant resistance and adaptation. For example, TCP20 acts as an activator of nitrate assimilatory genes and promotes the expression of these genes under the nitrogen starvation condition in A. thaliana [24]. In response to cold stress, overexpression of DgTCP1 improved the cold tolerance of chrysanthemum, while the DgTCP1 editing line showed decreased survival ratio under cold stress [25]. OsPCF2 has been proved to promote resistance to salt and drought stress in rice by activating the expression of the OsNHX1 gene [26]. Overall, these results demonstrate a broad spectrum of TCP functions in plant growth and environmental adaptation.
Radish (Raphanus sativus L) is an important root vegetable crop [27], and the whole genome sequence of radish has been published [28]. Up to date, genome-wide identification of the TCP gene family has been performed in several species, but not in radish. In this study, 32 RsTCP genes were identified in the radish genome, and the gene structure, conserved motif, phylogenetic and evolutionary relationship were analyzed. The expression pattern of RsTCP genes was further detected in different tissues, developmental stages, and stress conditions. Overall, these results preliminarily revealed the sequence information and potential biological functions of RsTCP transcription factors in radish, contributing basic information for further exploration of the molecular mechanisms underlying radish growth and responses to environmental stresses.
2. Materials and Methods
2.1. Plant Material and Treatments
The radish variety ‘YC-FH1’ was used to test the changes in RsTCP family expression in response to low temperature, saline and drought shock, as well as to treatments with abscisic acid (ABA) and gibberellin. Radish seeds were disinfected with 5% NaClO solution for 10 min, rinsed with distilled water, and germinated at 25 °C in the dark. Next, the germinated seeds were cultivated in nursery culture pots containing sterilized culture substrate. The photoperiod was 16 h/8 h (light/dark) and the temperature was 25 °C at day time and 16 °C at night. When the fourth or fifth true leaf appeared, healthy seedlings were randomly subjected to treatments with shocks and phytohormones. For low temperature shock, the culture temperature was adjusted to 4 °C [29]. In response to salt shock, 250 mM extra NaCl was added in the culture media [30]. Drought shock was simulated by incorporating 15% PEG6000 in the culture media [31]. For tests of phytohormones, 100 µM ABA solution [32] and 100 µM GA3 [33] were sprayed on leaves, independently. All treatments were repeated three times, and at least nine seedlings were used for each treatment. The samples were collected after each treatment and immediately frozen in liquid nitrogen and stored at −80 °C for RNA isolation.
2.2. Identification of RsTCP Genes in Radish Genome
The conserved domain of TCP proteins (http://pfam.xfam.org/family/PF03634; accessed on 13 March 2020) was used as the probe to search RsTCP sequences in the Radish Genome Database (http://radish-genome.org/; accessed on 15 May 2020) using the Hidden Markov Model (HMM) with the e-value set to 0.01 [28,34]. Sequences of candidate RsTCP genes were further queried against the SMART (http://smart.embl.de/; accessed on 15 May 2020) [35,36] and InterProScan (https://www.ebi.ac.uk/interpro/; accessed on 15 May 2020) databases to verify the reliability of the results [37]. The sequences of AtTCP genes were downloaded from the A. thaliana Information Resource (TAIR, https://www.arabidopsis.org/; accessed on 15 May 2020), and the sequences of OsTCP genes were downloaded from the Rice Genome Annotation Project (RAP) database (http://rice.plantbiology.msu.edu/index.shtml; accessed on 25 April 2022).
2.3. Protein Properties and Phylogeny Analysis
The characteristics of RsTCP proteins, including the number of amino acids (AA), theoretical molecular weight (MW), instability index (II), and isoelectric point (pI), were analyzed through the online ProtParam program (https://web.expasy.org/protparam/; accessed on 3 April 2021) [38]. The Online ScanProsite tool (https://prosite.expasy.org/scanprosite/; accessed on 2 September 2020) was used to analyze the post-translational modification sites of RsTCP proteins [39]. An un-rooted phylogenetic tree of 32 predicted RsTCP proteins, 24 reported AtTCP proteins, and 26 reported OsTCP proteins were generated subsequently using MEGA X software with: 1000 bootstrap replicates (Test of phylogeny), p-distance (Model/Method), and pairwise deletion (Gaps/Missing Data treatment) [36,40]. Evolview (http://www.evolgenius.info/evolview/; accessed on 25 April 2022) was used to visualize the un-rooted phylogenetic tree [41].
2.4. Gene Structure, Conserved Motifs, and Cis-Element Analyses
According to the genome sequences and coding sequences (CDS) of the RsTCP gene family, the structure of RsTCPs were analyzed using TBtools [42]. The conservative motif analysis of RsTCP genes was performed using the MEME program. The maximum number of motifs was set to 10 and the optimal motif width was set to 6–50 bp [43]. The online software PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/; accessed on 3 July 2020) was used to investigate the cis-elements of the RsTCP genes [44].
2.5. Synteny Analysis, Chromosomal Localization, and Prediction of rsa-miR319 Target Genes
Collinearity analysis was performed according to methods described in the PlantDGD (http://pdgd.njau.edu.cn:8080/; accessed on 11 August 2020) [45]. The collinearity block in RsTCP genes was identified based on duplication events using MCScanX [46], and then Circos was used to integrate and plot the data [47]. Based on the distribution information of the putative RsTCP genes from the radish genome, the location of RsTCPs on chromosomes was identified using TBtools software [42]. Finally, psRNATarget (http://plantgrn.noble.org/psRNATarget/; accessed on 27 August 2020) was used to identify rsa-miR319 target genes.
2.6. Transcription Analysis of RsTCP Genes
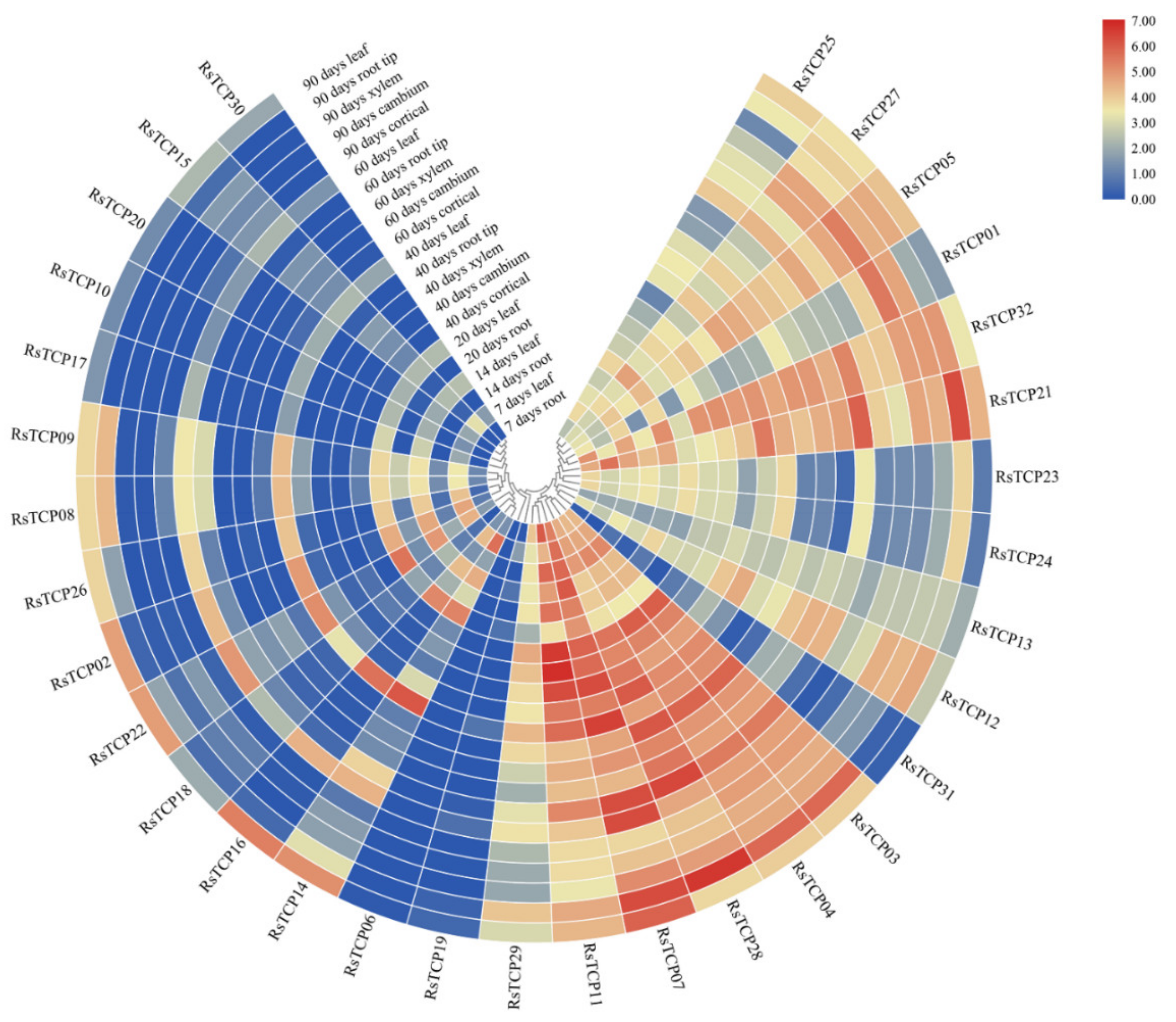
Illumina RNA sequencing data of radish leaf, root tip, cambium, cortex, and xylem tissues at six developmental stages (7, 14, 20, 40, 60, and 90 d) were downloaded from the NODAI Genome [48]. Afterwards, the transcription levels of each RsTCP gene were compared by calculating the reads per kilobase of transcript per million mapped reads (RPKM) as described in Mitsui et al. [48] and the heat map was generated using TBtools [42].
2.7. RNA Extraction and RT-qPCR Analysis
Total RNA of all samples was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer’s instruction, and the isolated RNA was subsequently treated with RNase-Free DNase I (Takara, Dalian, China). Total RNA was reverse-transcribed into cDNA using a first-strand cDNA synthesis kit (Bioer, Hangzhou, China). Specific primers (Table S1) for RsTCP genes were designed using Beacon Designer 7.7 (Premier Biosoft International, Palo Alto, CA, USA), and the SYBR Green RT-qPCR mix (Bioer, Hangzhou, China) was used for RT-qPCR assays on a LightCycler 480 Instrument (Roche, Mannheim, Germany). In addition, the RsActin gene was used as the internal control. At least three biological replicates were tested for each treatment, and three technical replicates were run for each reaction. The relative expression levels of each gene were calculated using the 2−ΔΔCT method [49]. The data used for expression profile analysis comprised the mean of three biological replicates, and the standard errors of the mean were calculated on the basis of the technical replicated data.
3. Results
3.1. Identification and Classification of RsTCP Members in Radish
In order to obtain the candidate TCP members in radish, the TCP domain (PF03634) was used to search against protein sequences of the radish genome. A total of 32 RsTCP members were identified from the radish genome and were named from RsTCP01 to RsTCP32 based on their positions on the chromosomes. Each RsTCP protein contained 109 to 460 amino acids, and their molecular weight ranged from 12.25 to 44.47 kDa. The theoretical pI ranged from 5.85 to 9.99, with a mean of 7.71, suggesting that most of RsTCPs were weakly alkaline (Table S2). The instability index of each protein ranged from 37.56 to 71.34. Among them, only one RsTCP showed an instability index less than 40.00, and thus this protein was predicted as a stable protein. The other 31 RsTCPs were structurally unstable. The aliphatic index ranged from 50.93 to 69.82, indicating that most RsTCPs contain a large number of aliphatic amino acids. The grand average of the hydropathy (GRAVY) value ranged from −1.043 to −0.204, indicating that all RsTCPs were hydrophilic (Table S2).
3.2. Phylogenetic Analysis of RsTCP Proteins
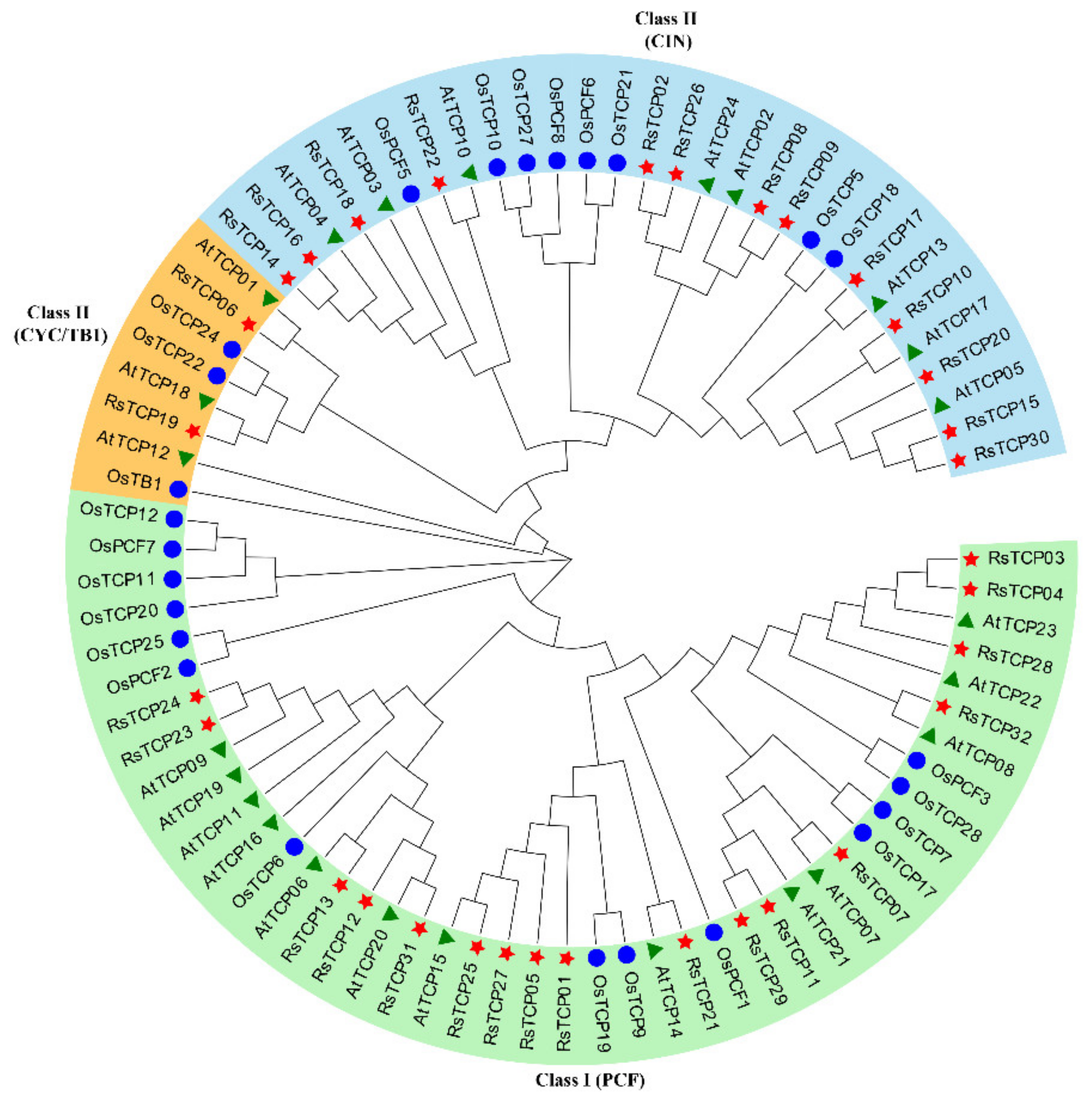
To better clarify the phylogenetic relationships among the predicted RsTCPs, the 32, 25, and 24 TCP proteins identified from radish, rice, and Arabidopsis were aligned using the ClustalW program, and then an un-rooted phylogenetic tree was constructed using MEGA X (Figure 1). The result showed that these 81 TCP proteins could be clustered into two categories (Class I and Class II) and three groups (PCFs, CINs, and CYCs). The Class I (PCF) subgroup contained 44 members (54.32%), including 17 RsTCPs, 14 OsTCPs, and 13 AtTCPs, respectively. Class II included CIN and CYC/TB1 types. Among them, 13 RsTCPs, 8 OsTCPs, and 8 AtTCPs belonged to the CIN subgroup, while 2 RsTCPs, 3 OsTCPs, and 3 AtTCPs were categorized into the CYC/TB1 type, accounting for 9.88% of the total. In addition, the result also showed that the relationship of TCPs between radish and the dicot Arabidopsis was closer to that of the monocot rice.
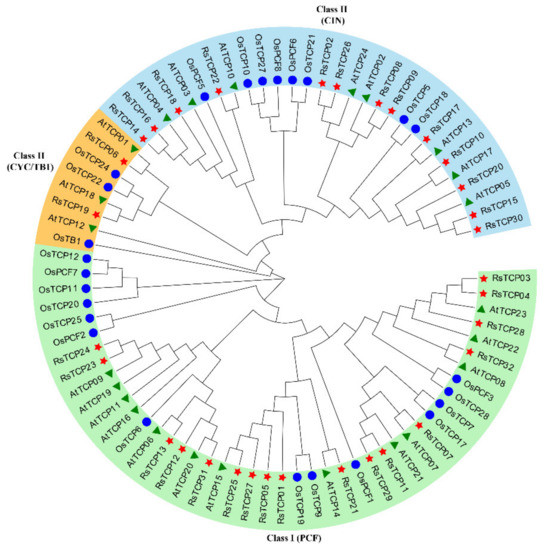
Figure 1.
Phylogenetic tree of the predicted TCP proteins from radish, rice, and Arabidopsis.
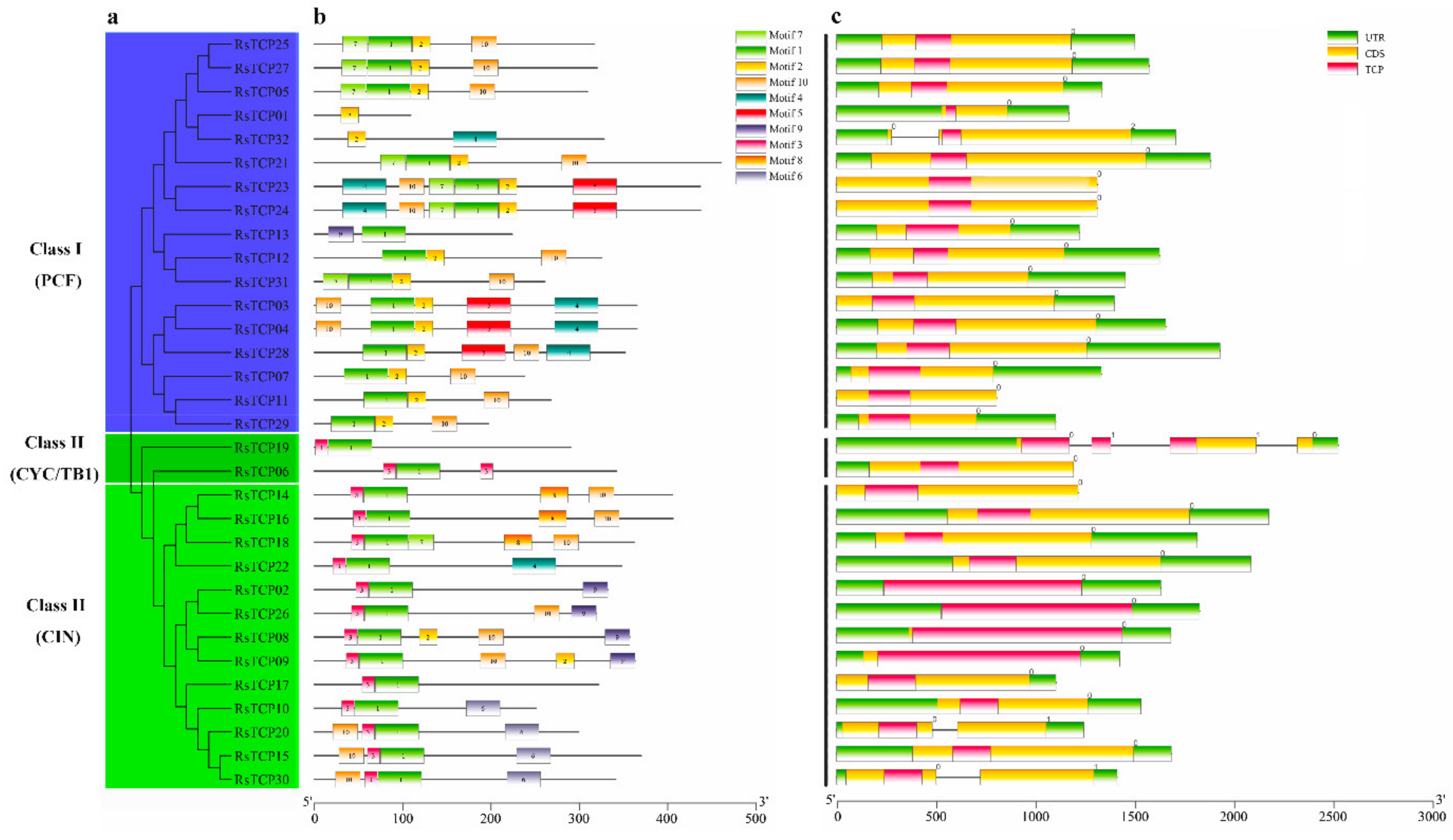
3.3. Motif and Gene Structure Analyses
In total, 10 conserved motifs were identified from 32 RsTCP proteins using the online software MEME motif search tool [50,51]. Subsequently, each of these 10 identified motifs containing 6–50aa was submitted to the InterProScan and SMART for further identification. Among them, motifs 1 and 2 were the typical characteristic domains of the plant TCP family and were present in all RsTCP members. However, the motif 3 exclusively existed in all Class II RsTCP members but not in Class I, suggesting that motif 3 may play an important role in the structural differences of RsTCP proteins (Figure 2b). Based on the phylogenetic analysis, most RsTCP proteins in the same evolutionary branch had a similar motif distribution. These results indicated that RsTCP proteins from the same evolutionary branch may have similar biological functions.
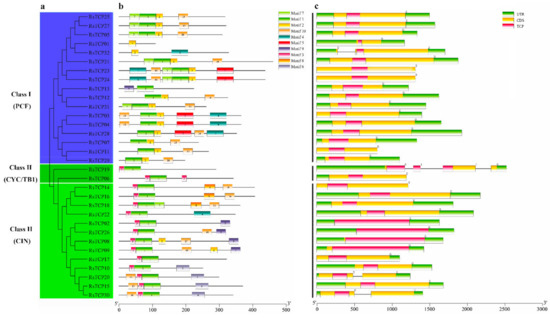
Figure 2.
Distribution of conserved motif and gene structure of RsTCP genes. (a) Phylogenetic tree of 32 RsTCP proteins; (b) distribution of the conserved motif for RsTCP proteins; (c) exon–intron structure of TCP genes in radish.
Exon–intron analysis of RsTCP genes indicated that the gene structure of the RsTCP gene family was relatively simple, and 28 out of the 32 RsTCP genes did not have any introns while RsTCP32, RsTCP06, RsTCP20, and RsTCP30 contained 1–3 introns (Figure 2c). In addition, except for RsTCP11, RsTCP14, RsTCP23, and RsTCP24, the remaining 28 RsTCP genes all had untranslated region (UTR) annotations in the radish genome.
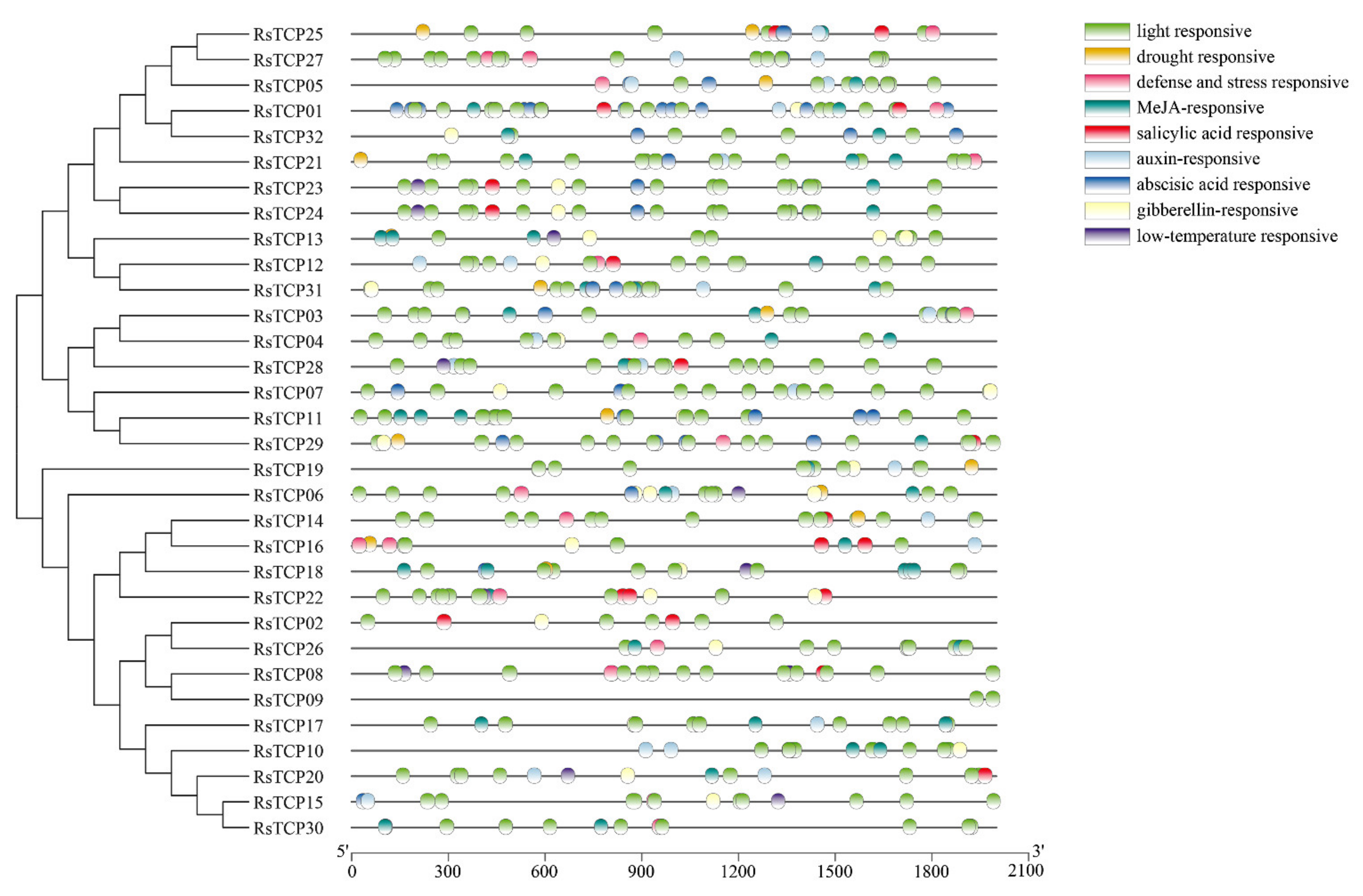
3.4. Cis-Element Analysis of RsTCP Genes
It is commonly accepted that the cis-elements play a critical role in regulation of gene transcription. In this study, the promoter sequences of all 32 RsTCP genes (within 2 kb upstream of the start codon) were analyzed using PlantCARE, and a total of 692 cis-elements were detected. In addition to some basic elements such as TATA-box and CAAT-box, nine important cis-elements potentially related to plant development and responses to various stresses were identified (Figure 3). The light responsive elements were observed in the promoter regions of all RsTCP genes. The drought responsive elements were detected in RsTCP25, RsTCP05, RsTCP21, RsTCP31, RsTCP03, RsTCP11, RsTCP29, RsTCP19, RsTCP06, RsTCP14, and RsTCP16. In addition, ABA, GA3 salicylic acid (SA), auxin (AUX), and low-temperature responsive elements presented in promoter regions of some RsTCP genes. These findings suggested that RsTCP genes might respond to changes in light, temperature, water supply, and be regulated by ABA, GA3, SA, and AUX.
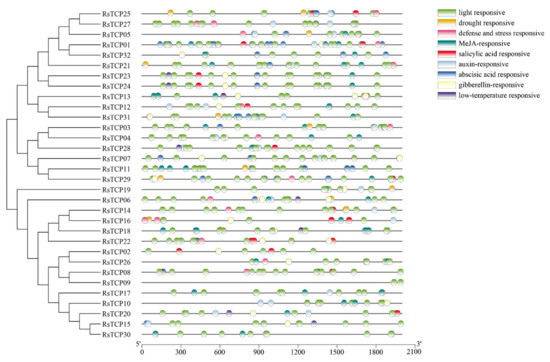
Figure 3.
Cis-acting elements on promoters of RsTCP genes.
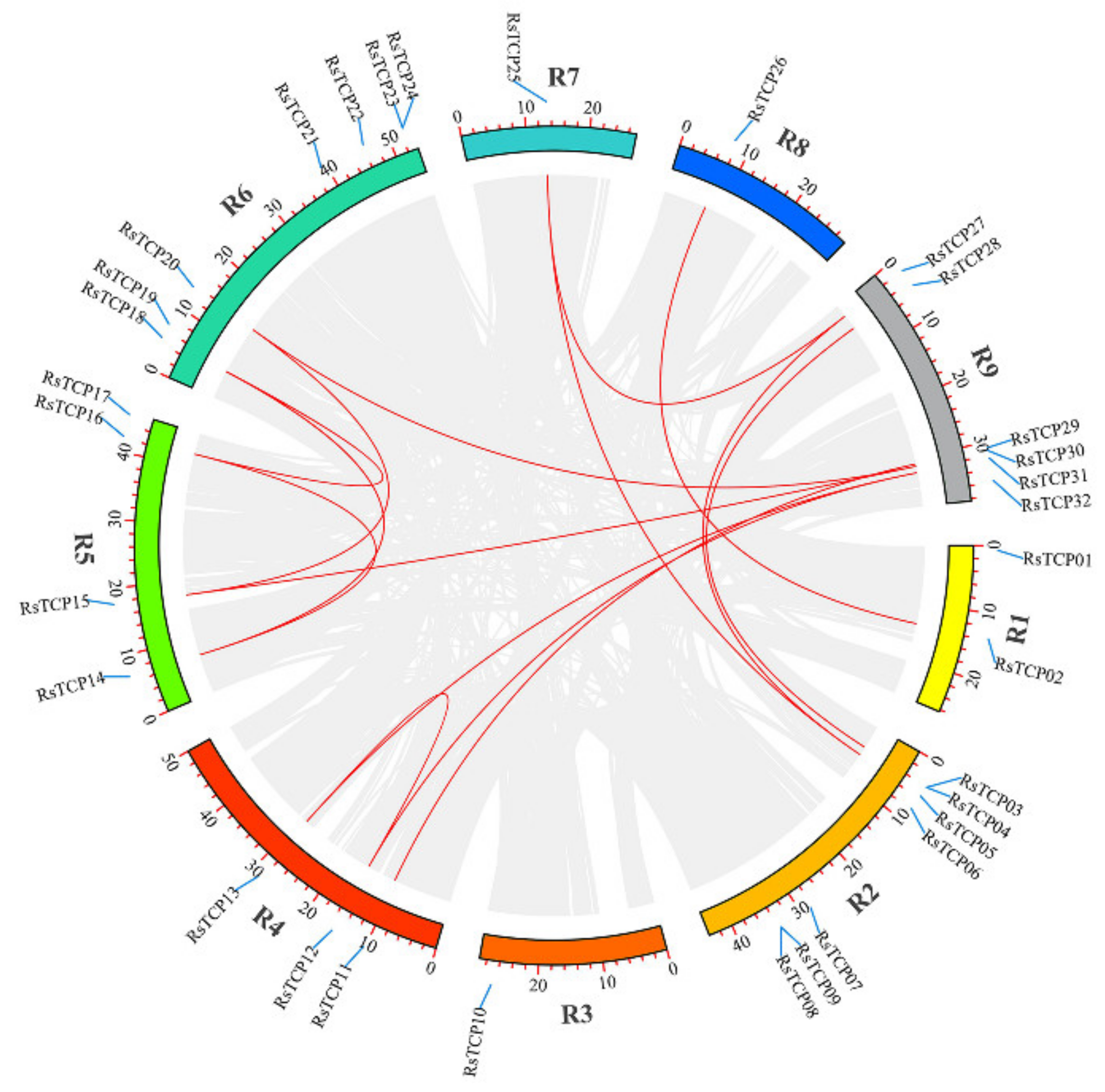
3.5. Chromosomal Localization and Gene Distribution Analysis
All these 32 RsTCP genes identified in this study were mapped to the nine chromosomes (from R1 to R9) in radish (Figure 4). The results showed that R2 and R6 contained the most RsTCP genes with seven RsTCPs (21.86%) on each chromosome, followed by R9 with six RsTCPs (18.75%). R5, R4, and R1 contained four, three, and two RsTCPs (12.5%, 9.38%, and 6.25%), respectively. In addition, each of R3, R7, and R8 contained only one RsTCP gene.
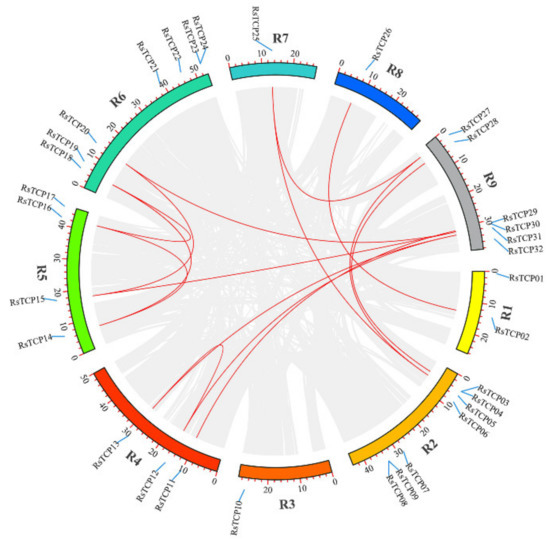
Figure 4.
The chromosomal location and synteny analysis of the RsTCP family. Red lines indicate the collinear relation.
Genome duplication events have been proven to promote the expansion of plant gene families, including whole-genome duplication (WGD)/segmental duplication (SD), transposed duplication (TRD), dispersed duplication (DD), proximal duplication (PD), and tandem duplication (TD) [52]. In this study, MCScanX analysis revealed that a total of 15 pairs of RsTCP genes were collinear, and 18 (56.25%) RsTCP genes were duplicated and retained in the WGD/SD (Figure 4). These results suggested that the WGD/SD event might play an important role in the extension of the RsTCP gene family.
3.6. Evolution Analysis of the RsTCP Genes and Target Site Analysis of rsa-miR319
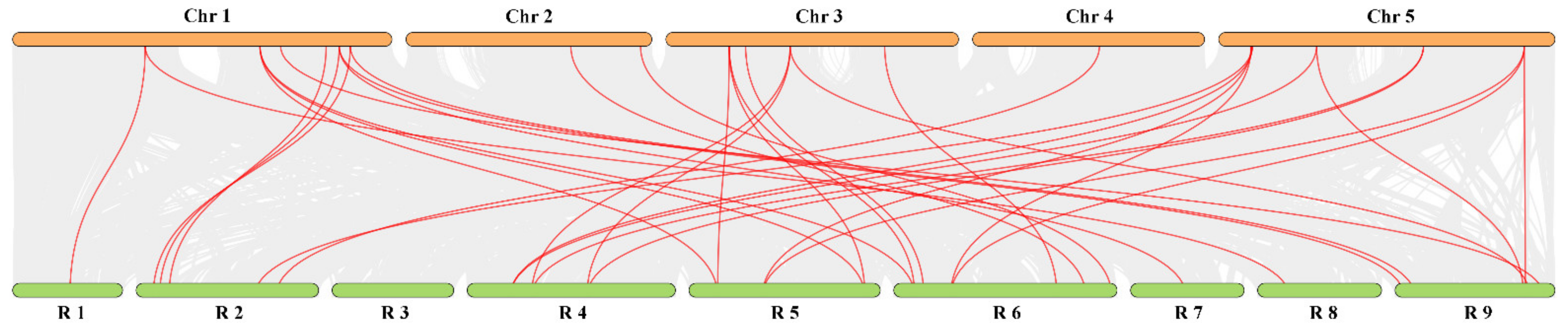
In order to explore the evolutionary history and the origin of TCP genes, syntenic regions were compared between radish and A. thaliana (Figure 5). The result demonstrated that 34 pairs of collinear genes were obtained, including 26 RsTCP genes and 18 AtTCP genes. Among them, six pairs of orthologous genes (one to one) were identified, including RsTCP06 and AtTCP01, RsTCP09 and AtTCP02, RsTCP23 and AtTCP09, RsTCP22 and AtTCP10, RsTCP21 and AtTCP14, as well as RsTCP32 and AtTCP08, suggesting that these genes might have the same common ancestors between A. thaliana and radish. In addition, one RsTCP gene may correspond to multiple A. thaliana TCP genes, for example, RsTCP11 and AtTCP21/07, RsTCP18 and AtTCP03/04. Similarly, multiple radish genes may correspond to one A. thaliana gene, such as AtTCP03 and RsTCP14/16/18, AtTCP15 and RsTCP05/25/27 (Figure 5, Table S3). These synchronous events revealed that some TCP genes evolved before the divergence of the A. thaliana and radish lineages.
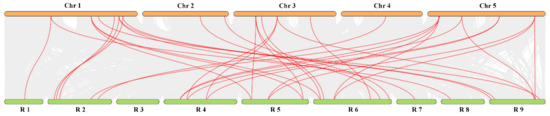
Figure 5.
Synteny analysis of TCP genes between radish and Arabidopsis. Colored lines connecting two chromosomal regions indicate syntenic regions between radish (R1–9) and Arabidopsis (Chr1–5) chromosomes.
The target sites of rsa-miR319 in the RsTCP genes were predicted by psRNATarget online software, and it was found that eight putative genes contained rsa-miR319 recognition site, namely RsTCP02, RsTCP08, RsTCP09, RsTCP14, RsTCP16, RsTCP18, RsTCP22, and RsTCP26 (Figure 6), while the other RsTCP genes did not contain a putative rsa-miR319 recognition site. This result suggested that these eight putative genes may play more critical roles in the growth and development of radish than other RsTCP members.

Figure 6.
Alignment of putative target sites for rsa-miR319. Mismatches are represented in yellow.
3.7. Characterization of Deduced RsTCP Proteins
Multiple post-translational modification sites were identified in the RsTCP protein using ScanProsite. Except RsTCP03, several putative phosphorylation sites were identified in other RsTCP proteins in the form of cAMP-cGMP kinase (cAMP-cGMP), protein kinase C (PKC), and casein kinase II (CK II). Additionally, N-Glycosylation (N-Glyc) and N-Myristoylation (N-Myr) were also found in deduced RsTCP protein sequences (Table 1). Interestingly, RsTCP proteins in the same evolutionary branch showed similar composition and number of post-translational modification sites. For example, both RsTCP23 and RsTCP24 in Class I contained nine PKC, three cAMP-cGMP, five CK II, five N-Myr, and one N-Glyc modification site. Both RsTCP08 and RsTCP09 had four PKC, seven CK II, and four N-Glyc modification sites. Similar results were also found in Class II members. These modification sites should be important for the regulation of protein activity and cell signal transduction.

Table 1.
Predicted RsTCP protein sequence features.
3.8. Transcription Profiling of RsTCP Genes in Radish
From the published database, RPKM of RsTCP genes in leaf, root tip, cambium, cortical, and xylem at different developmental stages were obtained and further analyzed (Figure 7, Table S4). Overall, the transcript levels of Class I members were significantly higher than those of Class II members in different growth periods and tissues. In the Class I, the expression of RsTCP31 was lower in all samples, while the other 16 members showed higher levels in some samples, especially RsTCP03, RsTCP04, RsTCP07, RsTCP11, and RsTCP28. The expression profiles of these RsTCP genes varied greatly among developmental stages, suggesting their potential roles in radish development. In contrast, significantly higher expression levels of RsTCP02, RsTCP08, RsTCP09, RsTCP14, RsTCP16, RsTCP18, RsTCP22, and RsTCP26 were observed in leaves at different stages, suggesting that these eight members may play crucial roles in the development of radish leaves. However, the expression profiles of other Class II members were extremely low in all samples, especially the two CYC/TB1 members in Class II, which were barely detected in all tissues at all stages. In addition, the expression of RsTCP03, RsTCP04, and RsTCP28 showed significant upregulation in cortical at 40- and 60-day-old stages and root tip at 60- and 90-day-old stages, demonstrating that these three RsTCP genes might be specifically involved in regulating specific stages of radish development. Overall, these results suggested that transcription of RsTCP genes was spatial- and temporal-specific, indicating that they may play different functions in the growth and development process.
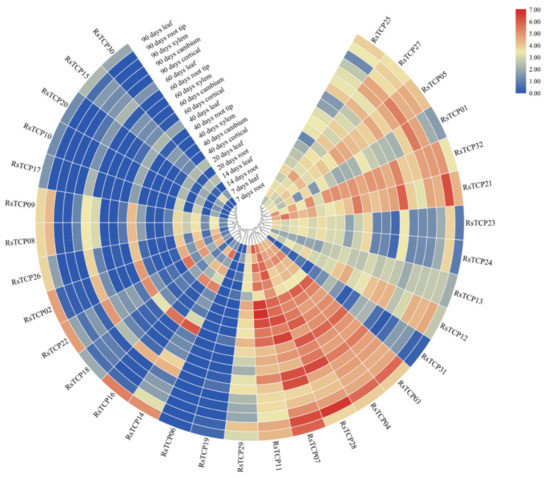
Figure 7.
Expression profile of RsTCP genes in different stages and tissues.
3.9. Transcription of RsTCP Genes in Response to Environmental Shocks and Phytohormones
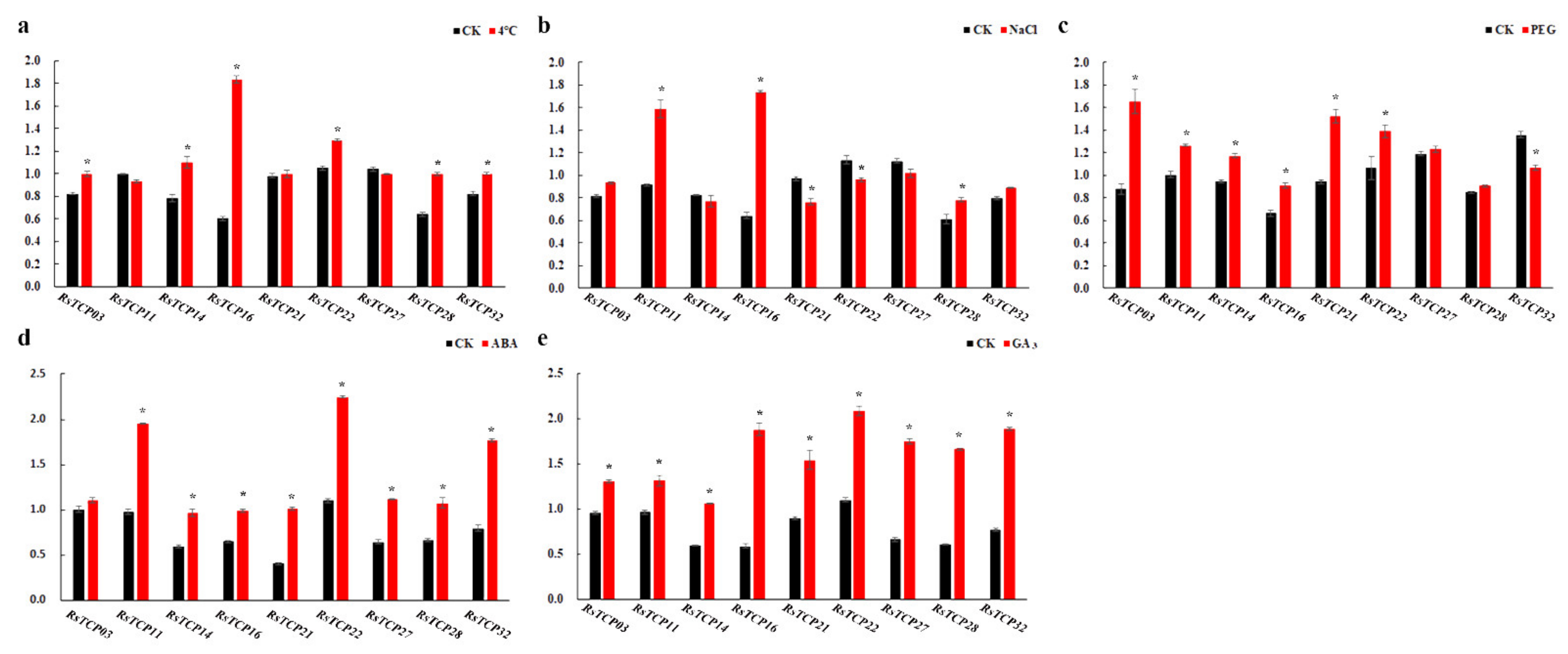
Based on the results of transcriptome data, nine RsTCP genes were selected for further investigation of the expression patterns of RsTCP genes after 4 °C, 250 mM NaCl, 15% w/v PEG6000, 100 µM ABA, and 100 µM GA3 treatments. In response to low temperature, except for RsTCP11 and RsTCP27, which showed slight downregulation, and RsTCP21 showed slight upregulation, the remaining six RsTCP genes were significantly upregulated compared with the control (Figure 8a). After NaCl treatment for 24 h, only RsTCP11, RsTCP16, and RsTCP28 showed significant upregulation, and RsTCP21 and RsTCP22 showed significant downregulation, the expression levels of the remaining four genes only showed slight changes (Figure 8b). Under drought shock, RsTCP32 exhibited a unique and significant downregulation, whereas the expression of the remaining eight RsTCP genes all showed upregulation to varying degrees (Figure 8c). Moreover, a slight increase in the transcript abundance of RsTCP03 was observed, while the expression level of the others showed significant upregulation under 100 µM ABA treatment for 24 h (Figure 8d). In addition, RsTCP genes were all highly sensitive to GA3 exposure, and all RsTCP genes were significantly upregulated after 24 h treatment of 100 µM GA3 solution (Figure 8e). The expression level of RsTCP16 showed significant upregulation under the five treatments, especially under 44 °C, NaCl and GA3. In summary, the transcript levels of RsTCP genes were differentially regulated under various treatments, indicating that the RsTCP genes might play an important role in response to abiotic stresses and phytohormones in radish.
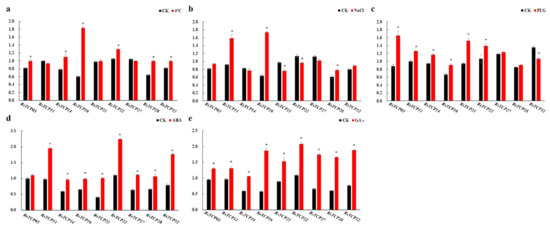
Figure 8.
The expression of RsTCP genes after 24 h treatment in response to environmental shocks and phytohormones. (a) Low temperature treatment (44 °C); (b) NaCl (250 mM); (c) PEG6000 (15% w/v); (d) ABA (100 µM); (e) GA3 (100 µM). The RsActin gene was used as the internal control, and three biological and technical replicates were conducted. The relative expression levels of each gene were calculated using the 2−ΔΔCT method. Each bar shows the mean ± SE of the triplicate assay, and * indicates significant difference at p < 0.05 according to Duncan’s multiple range test.
4. Discussion
Based on whole-genome sequences, the TCP gene family has been identified in many species [8,9,10,11,12,13,14,15]. Although previous studies have shown that some TCP genes in radish could not only participate in anthocyanin biosynthesis, but also respond to Pb, Cr, and heat stress, the members of the TCP gene family in radish have not been identified yet, and the structure, evolution, and expression patterns of these members have not been analyzed [53,54,55,56]. In this study, 32 TCP genes were identified in the radish genome according to the conserved domain of TCP proteins.
TCP genes are generally conserved in different plant species. In this study, the phylogenetic tree showed that a total of 32 RsTCP genes were divided into two major groups and three subgroups (Figure 1), consistent with the previous studies in other species [15,57,58]. As shown in Figure 1, the phylogenetic analysis showed 13 AtTCPs in Class I and 11 AtTCPs in Class II, which was consistent with Navaud et al. [8], demonstrating the reliability of the phylogenetic tree. Moreover, the expansion of the gene family was mainly caused by gene replication [59], and duplicated genes could obtain new functions to improve plant adaptability to the environment in the process of plant evolution [60]. In this study, a total of 34 pairs of collinear genes were obtained based on evolution analysis, including 26 RsTCP genes and 18 AtTCP genes (Figure 5, Table S3), while a total of 32 RsTCP genes were identified in this study, exactly 8 more than the 24 AtTCP genes identified in a previous study [8], indicating that TCP family members may have been replicated or expanded in the radish genome. In addition, similar to other plants, genes in the evolutionary branch shared similar gene structure and motif distribution [61,62], and a duplicated gene pair tended to be clustered into one branch. For examples, the pairs of RsTCP03-RsTCP04 and AtTCP15-RsTCP05/25/27 were clustered into Class I (PCF), the pairs of RsTCP02-RsTCP26 and AtTCP03-RsTCP14/16/18 were clustered into Class II (CIN) (Figure 1, Figure 4 and Figure 5). Overall, these results provided a reliable and informative basis of the TCP gene family in radish, which may be useful for further investigation of the regulatory functions of TCP genes in radish.
TCP transcription factors could affect plant growth, development and multiple physiological processes [63], including gametophyte development, seed germination, floral organ and leaf development [3,4,21,64,65]. Moreover, it is well known that microRNA (miRNA) control key biological processes in plants [66,67,68,69,70]. Among them, miR319 has been extensively studied for its roles in regulating leaf shape development due to its close relationship with TCP genes. For example, a previous study showed that BrTCP4, as an miR319 target gene, could regulate the round shape of leafy heads of Chinese cabbage [71]. Recent research revealed that AtTCP2 could play multiple roles through miR319, including regulating leaf morphogenesis and leaf senescence in A. thaliana [72]. In this study, eight putative rsa-miR319-targeted genes were identified (Figure 6). Interestingly, the phylogenetic tree showed that all these eight putative rsa-miR319-target genes belonged to the Class II (CIN) subfamily, which was similar to the result in cucumber [15]. Moreover, the expression profile analysis showed that these eight genes had relatively high expression levels in leaves at various developmental stages (Figure 7), indicating that these eight genes might play a key role in leaf development in radish. In addition, sequence characterization and phylogenetic analysis suggest that RsTCP16 is a homologue to AtTCP4 in A. thaliana (Figure 1 and Figure 5, Table S3), which not only regulates leaf growth but also inhibits cell proliferation [73,74]. Therefore, we speculated that the RsTCP16 gene might similarly regulate leaf development in radish.
Increasing evidence indicated that TCP genes could not only participate in the biosynthesis of phytohormones, but also play a key role in the regulation of phytohormone-mediated development and growth [26,75]. In this study, multiple phytohormone-related cis-acting elements were identified in the promoter regions of RsTCP genes (Figure 3), consistent with the results in cucumber [15]. Post-translational modification is also crucial for plant growth and stress tolerance [76]. In this study, multiple post-translational modification sites were predicted in RsTCPs (Table 1), indicating that the RsTCP gene family may participate in a variety of biological processes. The results of RT-qPCR suggested that only some of these nine RsTCP genes were sensitive to low temperature shock, salt shock and drought shock, and almost all RsTCP genes were significantly upregulated in response to ABA and GA3 treatments. In addition, the transcription of RsTCP16 was significantly upregulated under the five treatments, suggesting that RsTCP16 could play an important role in response to abiotic stresses and phytohormones (Figure 8). In conclusion, our results suggested that RsTCP16 might play key roles in the growth and development of radish and lay the foundation for further investigation on the mechanism underlying the abiotic stress and phytohormones response in radish.
5. Conclusions
In summary, we undertook the first identification of a total of 32 RsTCP genes from the radish genome, which localized on nine radish chromosomes. Meanwhile, several cis-elements involved in plant development and abiotic stress response were detected in the promoter regions. Moreover, post-translational modification sites were found in the RsTCP proteins and evolution analysis suggested that some TCP genes were evolved before the divergence of the radish and A. thaliana lineages. In addition, the results of this study also showed that RsTCP genes exhibited tissue-specific expression patterns and revealed that RsTCP16 might play a key role in response to abiotic stresses and phytohormones.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae8070656/s1, Table S1: Primer sequences for RT-qPCR of RsActin and RsTCP genes; Table S2: The information of the TCP gene family in radish; Table S3: Synteny blocks of TCP genes between radish and Arabidopsis genomes; Table S4: Tissue-specific expression of RsTCP genes.
Author Contributions
Conceptualization, Y.M., Z.L. and F.S.; methodology, Y.M., Z.L., J.Z., W.W., L.Z. and R.F.; software, W.W. and L.Z.; validation, Z.L.; investigation, Y.M. and J.Z.; resources, Y.Z.; data curation, Y.W.; writing—original draft preparation, Y.M.; writing—review and editing, Z.L. and F.S.; visualization, Y.M. and W.W.; supervision, Y.Z. and Y.W.; project administration, Y.M., Z.L. and F.S.; funding acquisition, Y.M. and F.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from Jiangsu Agricultural Science and Technology Innovation Fund (JASTIF, CX (20)3097).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained in the article.
Acknowledgments
Thank the Xiaobo Luo, from Guizhou Institute of Biotechnology for his technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Paul, M.V.; Iyer, S.; Amerhauser, C.; Lehmann, M.; Van Dongen, J.T.; Geigenberger, P. Oxygen Sensing via the Ethylene Response Transcription Factor RAP2.12 Affects Plant. Metabolism and Performance under Both Normoxia and Hypoxia. Plant Physiol. 2016, 172, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehti-Shiu, M.D.; Panchy, N.; Wang, P.; Uygun, S.; Shiu, S.-H. Diversity, expansion, and evolutionary novelty of plant DNA-binding transcription factor families. Biochim. Biophys. Acta Gene Reg. Mech. 2017, 1860, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Cochet, F.; Ponnaiah, M.; Lebreton, S.; Matheron, L.; Pionneau, C.; Boudsocq, M.; Resentini, F.; Huguet, S.; Blázquez, M.; et al. The MPK8-TCP14 pathway promotes seed germination in Arabidopsis. Plant J. 2019, 100, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, H.; Mou, M.; Chen, Y.; Xiang, S.; Chen, L.; Yu, D. Arabidopsis Class II TCP Transcription Factors Integrate with the FT-FD Module to Control Flowering. Plant Physiol. 2019, 181, 97–111. [Google Scholar] [CrossRef]
- Lan, J.; Zhang, J.; Yuan, R.; Yu, H.; An, F.; Sun, L.; Chen, H.; Zhou, Y.; Qian, W.; He, H.; et al. TCP transcription factors suppress cotyledon trichomes by impeding a cell differentiation-regulating complex. Plant Physiol. 2021, 186, 434–451. [Google Scholar] [CrossRef]
- Gastaldi, V.; Lucero, L.E.; Ferrero, L.V.; Ariel, F.D.; Gonzalez, D.H. Class-I TCP Transcription Factors Activate the SAUR63 Gene Subfamily in Gibberellin-Dependent Stamen Filament Elongation. Plant Physiol. 2020, 182, 2096–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: A motif found in proteins regulating plant growth and development. Plant J. 1999, 18, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Navaud, O.; Dabos, P.; Carnus, E.; Tremousaygue, D.; Hervé, C. TCP transcription factors predate the emergence of land plants. J. Mol. Evol. 2007, 65, 23–33. [Google Scholar] [CrossRef]
- Parapunova, V.; Busscher, M.; Busscher-Lange, J.; Lammers, M.; Karlova, R.; Bovy, A.G.; Angenent, G.C.; de Maagd, R.A. Identification, cloning and characterization of the tomato TCP transcription factor family. BMC Plant Biol. 2014, 14, 157. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Guy, K.M.; Wu, W.; Fang, B.; Yang, J.; Zhang, M.; Hu, Z. Genome-wide identification and expression analysis of the ClTCP transcription factors in Citrullus lanatus. BMC Plant Biol. 2016, 16, 85. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.F.; Chen, Y.Y.; Hsiao, Y.Y.; Shen, C.Y.; Hsu, J.L.; Yeh, C.M.; Mitsuda, N.; Ohme-Takagi, M.; Liu, Z.J.; Tsai, W.C. Genome-wide identification and characterization of TCP genes involved in ovule development of Phalaenopsis equestris. J. Exp. Bot. 2016, 67, 5051–5066. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Hu, S.; Yu, Q.; Wang, C.; Yang, Y.; Sun, H.; Yang, Y.; Sun, X. Genome-Wide Identification and Characterization of BrrTCP Transcription Factors in Brassica rapa ssp. rapa. Front Plant Sci. 2017, 8, 1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, K.; Ni, Z.; Qu, Y.; Cai, Y.; Yang, Z.; Sun, G.; Chen, Q. Genome-wide identification and expression analyses of TCP transcription factor genes in Gossypium barbadense. Sci. Rep. 2018, 8, 14526. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.M.; Wang, M.M.; Yang, J.; Wen, J.; Guo, P.C.; Wu, Y.W.; Ke, Y.Z.; Li, P.F.; Li, J.N.; Du, H. Evolutionary and Comparative Expression Analyses of TCP Transcription Factor Gene Family in Land Plants. Int. J. Mol. Sci. 2019, 20, 3591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, H.; Chen, Y.; Du, H.; Zhang, L.; Zhang, K.; He, H.; Pan, J.; Cai, R.; Wang, G. Genome-Wide Identification and Characterization of the TCP Gene Family in Cucumber (Cucumis sativus L.) and Their Transcriptional Responses to Different Treatments. Genes 2020, 11, 1379. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, P.; Das, G.M.; Joseph, A.P.; Chatterjee, N.; Srinivasan, N.; Nath, U. Identification of specific DNA binding residues in the TCP family of transcription factors in Arabidopsis. Plant Cell. 2010, 22, 1174–1189. [Google Scholar] [CrossRef] [Green Version]
- Danisman, S.; Wal, F.V.D.; Dhondt, S.; Waites, R.; de Folter, S.; Bimbo, A.; van Dijk, A.D.; Muino, J.M.; Cutri, L.; Dornelas, M.C.; et al. Arabidopsis class I and class II TCP transcription factors regulate jasmonic acid metabolism and leaf development antagonistically. Plant Physiol. 2012, 159, 1511–1523. [Google Scholar] [CrossRef] [Green Version]
- Howarth, D.G.; Donoghue, M.J. Phylogenetic analysis of the “ECE” (CYC/TB1) clade reveals duplications predating the core eudicots. Proc. Natl. Acad. Sci. USA 2006, 103, 9101–9106. [Google Scholar] [CrossRef] [Green Version]
- Martín-Trillo, M.; Cubas, P. TCP genes: A family snapshot ten years later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef]
- Nicolas, M.; Cubas, P. TCP factors: New kids on the signaling block. Curr. Opin. Plant Biol. 2016, 33, 33–41. [Google Scholar] [CrossRef]
- Ju, Y.; Guo, L.; Cai, Q.; Ma, F.; Zhu, Q.Y.; Zhang, Q.; Sodmergen. Arabidopsis JINGUBANG Is a Negative Regulator of Pollen Germination That Prevents Pollination in Moist Environments. Plant Cell. 2016, 28, 2131–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.Y.; Zhao, P.M.; Cheng, H.Q.; Han, L.B.; Wu, X.M.; Gao, P.; Wang, H.Y.; Yang, C.L.; Zhong, N.Q.; Zuo, J.R.; et al. The cotton transcription factor TCP14 functions in auxin-mediated epidermal cell differentiation and elongation. Plant Physiol. 2013, 162, 1669–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Zhang, D.; Li, J. TCP1 Modulates DWF4 Expression via Directly Interacting with the GGNCCC Motifs in the Promoter Region of DWF4 in Arabidopsis thaliana. J. Genet. Genom. 2015, 42, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Ripoll, J.J.; Wang, R.; Vuong, L.; Bailey-Steinitz, L.J.; Ye, D.; Crawford, N.M. Interacting TCP and NLP transcription factors control plant responses to nitrate availability. Proc. Natl. Acad. Sci. USA 2017, 114, 2419–2424. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, Q.; Liao, X.; Tian, Y.; Zhang, F.; Zhang, L.; Liu, Q. A natural antisense RNA improves chrysanthemum cold tolerance by regulating the transcription factor DgTCP1. Plant Physiol. 2022, kiac267. [Google Scholar] [CrossRef]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2017, 93, 61–77. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.; Zhai, L.; Xu, Y.; Wang, L.; Zhu, X.; Gong, Y.; Yu, R.; Limera, C.; Liu, L. Genome-wide identification and characterization of cadmium-responsive microRNAs and their target genes in radish (Raphanus sativus L.) roots. J. Exp. Bot. 2013, 64, 4271–4287. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.M.; Kim, N.; Ahn, B.O.; Oh, M.; Chung, W.H.; Chung, H.; Jeong, S.; Lim, K.B.; Hwang, Y.J.; Kim, G.B.; et al. Elucidating the triplicated ancestral genome structure of radish based on chromosome-level comparison with the Brassica genomes. Theor. Appl. Genet. 2016, 129, 1357–1372. [Google Scholar] [CrossRef]
- Li, Y.; Shan, X.; Jiang, Z.; Zhao, L.; Jin, F. Genome-wide identification and expression analysis of the GA2ox gene family in maize (Zea mays L.) under various abiotic stress conditions. Plant Physiol. Biochem. 2021, 166, 621–633. [Google Scholar] [CrossRef]
- Wang, Y.; Ying, J.; Zhang, Y.; Xu, L.; Zhang, W.; Ni, M.; Zhu, Y.; Liu, L. Genome-Wide Identification and Functional Characterization of the Cation Proton Antiporter (CPA) Family Related to Salt Stress Response in Radish (Raphanus sativus L.). Int. J. Mol. Sci. 2020, 4, 8262. [Google Scholar] [CrossRef]
- Mahesh, K.; Balaraju, P.; Ramakrishna, B.; Rao, S.S.R. Effect of Brassinosteroids on Germination and Seedling Growth of Radish (Raphanus sativus L.) under PEG-6000 Induced Water Stress. Am. J. Plant Sci. 2013, 4, 2305–2313. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Tang, Y.; Gao, S.; Su, S.; Hong, L.; Wang, W.; Fang, Z.; Li, X.; Ma, J.; Quan, W. Comprehensive analyses of the annexin gene family in wheat. BMC Genom. 2016, 17, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; He, X.; Chang, P.; Jiang, H.; Gong, D.; Sun, Q. Genome-wide identification and characterization of TCP family genes in Brassica juncea var. tumida. Peer. J. 2020, 8, e9130. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, 302–305. [Google Scholar] [CrossRef]
- Fan, L.; Xu, L.; Wang, Y.; Tang, M.; Liu, L. Genome- and Transcriptome-Wide Characterization of bZIP Gene Family Identifies Potential Members Involved in Abiotic Stress Response and Anthocyanin Biosynthesis in Radish (Raphanus sativus L.). Int. J. Mol. Sci. 2019, 20, 6334. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Ying, J.; Tang, M.; Wang, Y.; Xu, L.; Liu, M.; Liu, L. Genome-wide identification of AUX/IAA in radish and functional characterization of RsIAA33 gene during taproot thickening. Gene 2021, 795, 145782. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattike, R.A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Shen, F.; Ying, J.; Xu, L.; Sun, X.; Wang, J.; Wang, Y.; Mei, Y.; Zhu, Y.; Liu, L. Characterization of Annexin gene family and functional analysis of RsANN1a involved in heat tolerance in radish (Raphanus sativus L.). Physiol. Mol. Biol. Plants 2021, 27, 2027–2041. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Machanick, P.; Bailey, T.L. MEME-ChIP: Motif analysis of large DNA datasets. Bioinformatics 2011, 27, 1696–1697. [Google Scholar] [CrossRef] [Green Version]
- Rombauts, S.; Déhais, P.; Van Montagu, M.; Rouzé, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization-diploidization cycles in plants. Genom. Biol. 2019, 20, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, Y.; Shimomura, M.; Komatsu, K.; Namiki, N.; Shibata-Hatta, M.; Imai, M.; Katayose, Y.; Mukai, Y.; Kanamori, H.; Kurita, K.; et al. The radish genome and comprehensive gene expression profile of tuberous root formation and development. Sci. Rep. 2015, 5, 10835. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Bailey, T.L.; Gribskov, M. Combining evidence using p-values: Application to sequence homology searches. Bioinformatics 1998, 14, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Xu, L.; Wang, Y.; Cheng, W.; Luo, X.; Xie, Y.; Fan, L.; Liu, L. Genome-wide characterization and evolutionary analysis of heat shock transcription factors (HSFs) to reveal their potential role under abiotic stresses in radish (Raphanus sativus L.). BMC Genom. 2019, 20, 772. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Su, N.; Jia, L.; Tian, J.; Li, H.; Huang, L.; Shen, Z.; Cui, J. Transcriptome analysis of radish sprouts hypocotyls reveals the regulatory role of hydrogen-rich water in anthocyanin biosynthesis under UV-A. BMC Plant Biol. 2018, 18, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, W.; Shen, H.; Zhu, X.; Zhai, L.; Xu, L.; Wang, R.; Gong, Y.; Limera, C.; Liu, L. Identification of Radish (Raphanus sativus L.) miRNAs and Their Target Genes to Explore miRNA-Mediated Regulatory Networks in Lead (Pb) Stress Responses by High-Throughput Sequencing and Degradome Analysis. Plant Mol. Biol. Rep. 2015, 33, 358–376. [Google Scholar] [CrossRef]
- Yang, Z.; Li, W.; Su, X.; Ge, P.; Ge, P.; Zhou, Y.; Hao, Y.; Shu, H.; Gao, C.; Cheng, S.; et al. Early Response of Radish to Heat Stress by Strand-Specific Transcriptome and miRNA Analysis. Int. J. Mol. Sci. 2019, 20, 3321. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Xu, L.; Wang, Y.; Shen, H.; Zhu, X.; Zhang, K.; Chen, Y.; Yu, R.; Limera, C.; Liu, L. Transcriptome-wide analysis of chromium-stress responsive microRNAs to explore miRNA-mediated regulatory networks in radish (Raphanus sativus L.). Sci. Rep. 2015, 5, 14024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Wang, Q.; Sun, R.; Xie, F.; Jones, D.C.; Zhang, B. Genome-wide identification and expression analysis of TCP transcription factors in Gossypium raimondii. Sci. Rep. 2014, 4, 6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Chen, Y.Q.; Ding, A.M.; Chen, H.; Xia, F.; Wang, W.F.; Sun, Y.H. Genome-wide analysis of TCP family in tobacco. Genet. Mol. Res. 2016, 15, gmr7728. [Google Scholar] [CrossRef]
- Airoldi, C.A.; Davies, B. Gene duplication and the evolution of plant MADS-box transcription factors. J. Genet. Genom. 2012, 39, 157–165. [Google Scholar] [CrossRef]
- De Grassi, A.; Lanave, C.; Saccone, C. Genome duplication and gene-family evolution: The case of three OXPHOS gene families. Gene 2008, 421, 1–6. [Google Scholar] [CrossRef]
- Chen, P.; Li, J.; Ye, X.; Tan, B.; Zheng, X.; Cheng, J.; Wang, W.; Wang, H.; Gu, L.; Feng, J. Genome-wide identification of ziziphus jujuba TCP transcription factors and their expression in response to infection with jujube witches’ broom phytoplasma. Acta Physiol. Plant 2019, 41, 86. [Google Scholar] [CrossRef]
- Zheng, A.; Sun, F.; Cheng, T.; Wang, Y.; Xie, K.; Zhang, C.; Xi, Y. Genome-wide identification of members of the TCP gene family in switchgrass (Panicum virgatum L.) and analysis of their expression. Gene 2019, 702, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Ohashi, Y. DNA binding and dimerization specificity and potential targets for the TCP protein family. Plant J. 2002, 30, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Mitsuda, N.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. TCP transcription factors regulate the activities of ASYMMETRIC LEAVES1 and miR164, as well as the auxin response, during differentiation of leaves in Arabidopsis. Plant Cell. 2010, 22, 3574–3588. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xu, X.; Mo, X.; Zhong, L.; Zhang, J.; Mo, B.; Kuai, B. Overexpression of TCP8 delays Arabidopsis flowering through a FLOWERING LOCUS C-dependent pathway. BMC Plant Biol. 2019, 19, 534. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.E.; Schommer, C.; Palatnik, J.F. Control of cell proliferation by microRNAs in plants. Curr. Opin. Plant Biol. 2016, 34, 68–76. [Google Scholar] [CrossRef]
- Shi, G.; Fu, J.; Rong, L.; Zhang, P.; Guo, C.; Xiao, K. TaMIR1119, a miRNA family member of wheat (Triticum aestivum), is essential in the regulation of plant drought tolerance. J. Integr. Agric. 2018, 17, 2369–2378. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Prasad, A.; Maurya, J.; Prasad, M. Regulation of small RNA-mediated high temperature stress responses in crop plants. Plant. Cell Rep. 2022, 41, 765–773. [Google Scholar] [CrossRef]
- Ali, M.; Javaid, A.; Naqvi, S.H.; Batcho, A.; Kayani, W.K.; Lal, A.; Sajid, I.A.; Nwogwugwu, J.O. Biotic stress triggered small RNA and RNAi defense response in plants. Mol. Biol. Rep. 2020, 47, 5511–5522. [Google Scholar] [CrossRef]
- Zhang, L.; Song, C.; Guo, D.; Guo, L.; Hou, X.; Wang, H. Identification of differentially expressed miRNAs and their target genes in response to brassinolide treatment on flowering of tree peony (Paeonia ostii). Plant Signal Behav. 2022, 17, 2056364. [Google Scholar] [CrossRef]
- Mao, Y.; Wu, F.; Yu, X.; Bai, J.; Zhong, W.; He, Y. MicroRNA319a-targeted Brassica rapa ssp. pekinensis TCP genes modulate head shape in chinese cabbage by differential cell division arrest in leaf regions. Plant Physiol. 2014, 164, 710–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Li, S.; Ma, Q.; Wen, J.; Yan, K.; Li, Q. The Acer palmatum TCP Transcription Factor ApTCP2 Controls Leaf Morphogenesis, Accelerates Senescence, and Affects Flowering via miR319 in Arabidopsis thaliana. J. Plant Growth Regul. 2021, 41, 244–256. [Google Scholar] [CrossRef]
- Schommer, C.; Debernardi, J.M.; Bresso, E.G.; Rodriguez, R.E.; Palatnik, J.F. Repression of cell proliferation by miR319-regulated TCP4. Mol. Plant 2014, 7, 1533–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresso, E.G.; Chorostecki, U.; Rodriguez, R.E.; Palatnik, J.F.; Schommer, C. Spatial Control of Gene Expression by miR319-Regulated TCP Transcription Factors in Leaf Development. Plant Physiol. 2018, 176, 1694–1708. [Google Scholar] [CrossRef] [PubMed]
- Es, S.W.; Silveira, S.R.; Rocha, D.I.; Bimbo, A.; Martinelli, A.P.; Dornelas, M.C.; Angenent, G.C.; Immink, R.G.H. Novel functions of the Arabidopsis transcription factor TCP5 in petal development and ethylene biosynthesis. Plant J. 2018, 94, 867–879. [Google Scholar]
- Mizoi, J.; Kanazawa, N.; Kidokoro, S.; Takahashi, F.; Qin, F.; Morimoto, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arabidopsis thaliana. J. Biol. Chem. 2019, 294, 902–917. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).