
In Vitro Techniques for Shipping of Micropropagated Plant Materials

Abstract
:1. Introduction
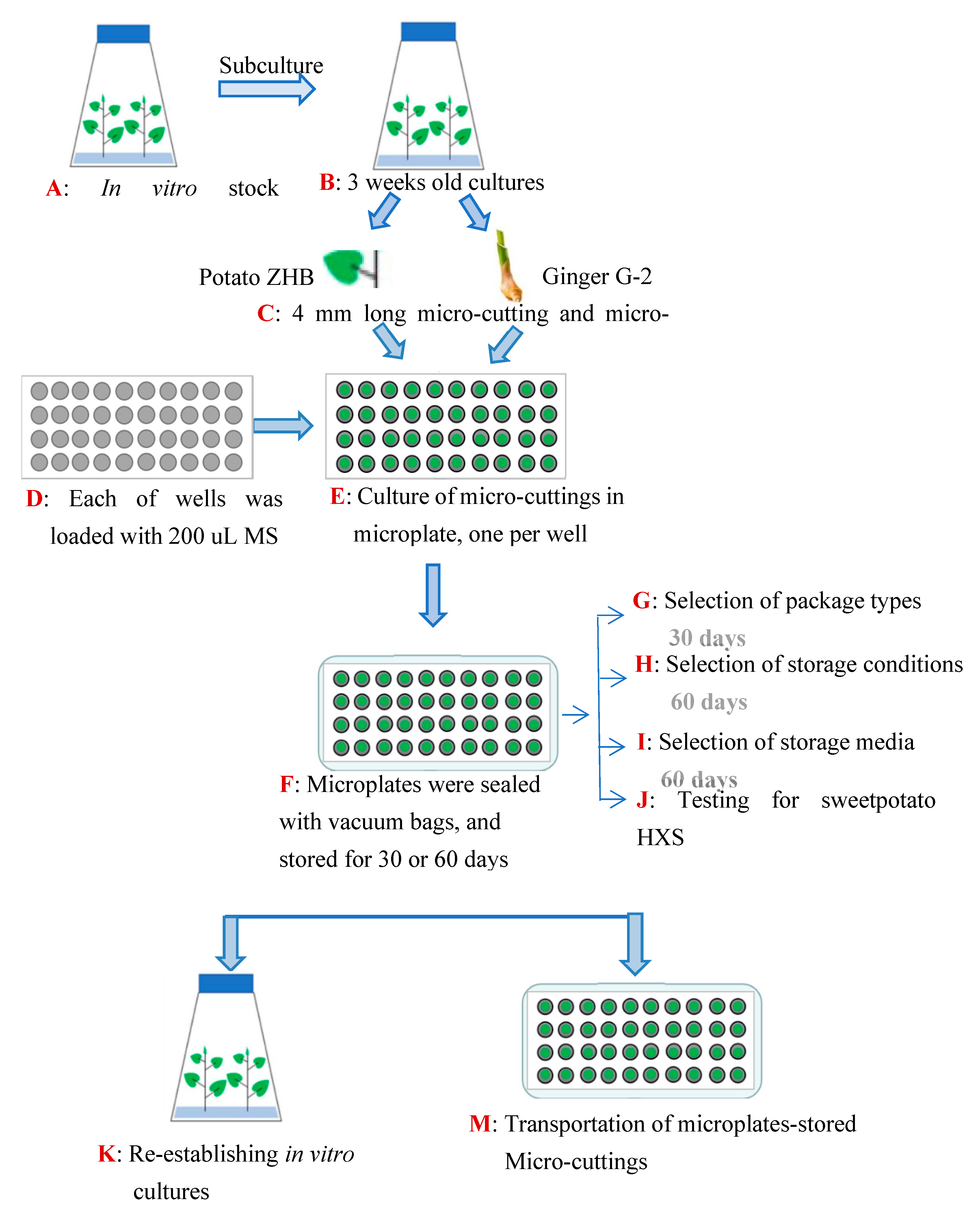
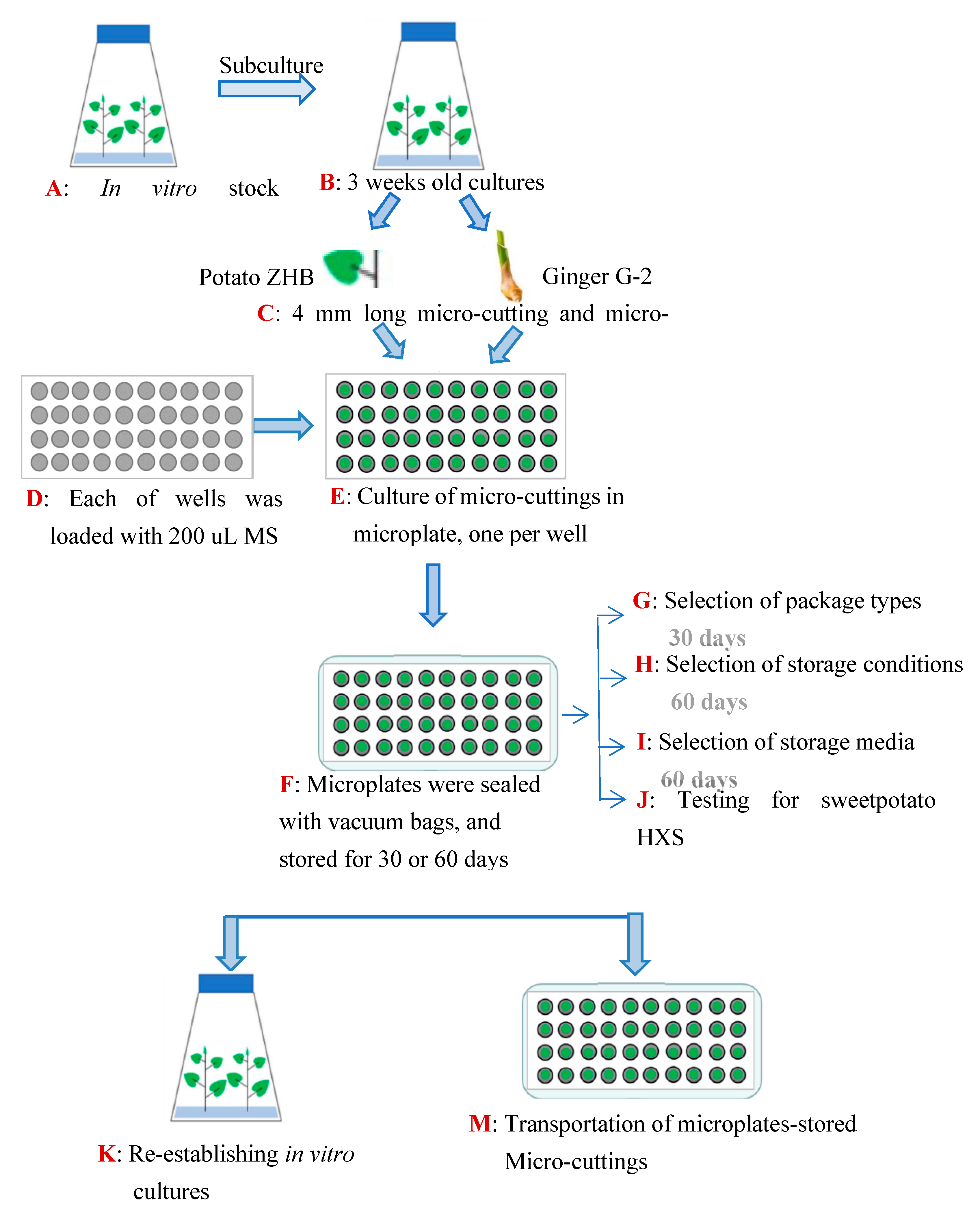
2. Materials and Methods
2.1. Plant Materials
2.2. Assessment of Packaging Type
2.3. Assessment of Incubation Condition
2.4. Assessment of Culture Medium
2.5. Regrowth Capacity Assessment of Maintained Plants
2.6. Assessment of Genetic Stability
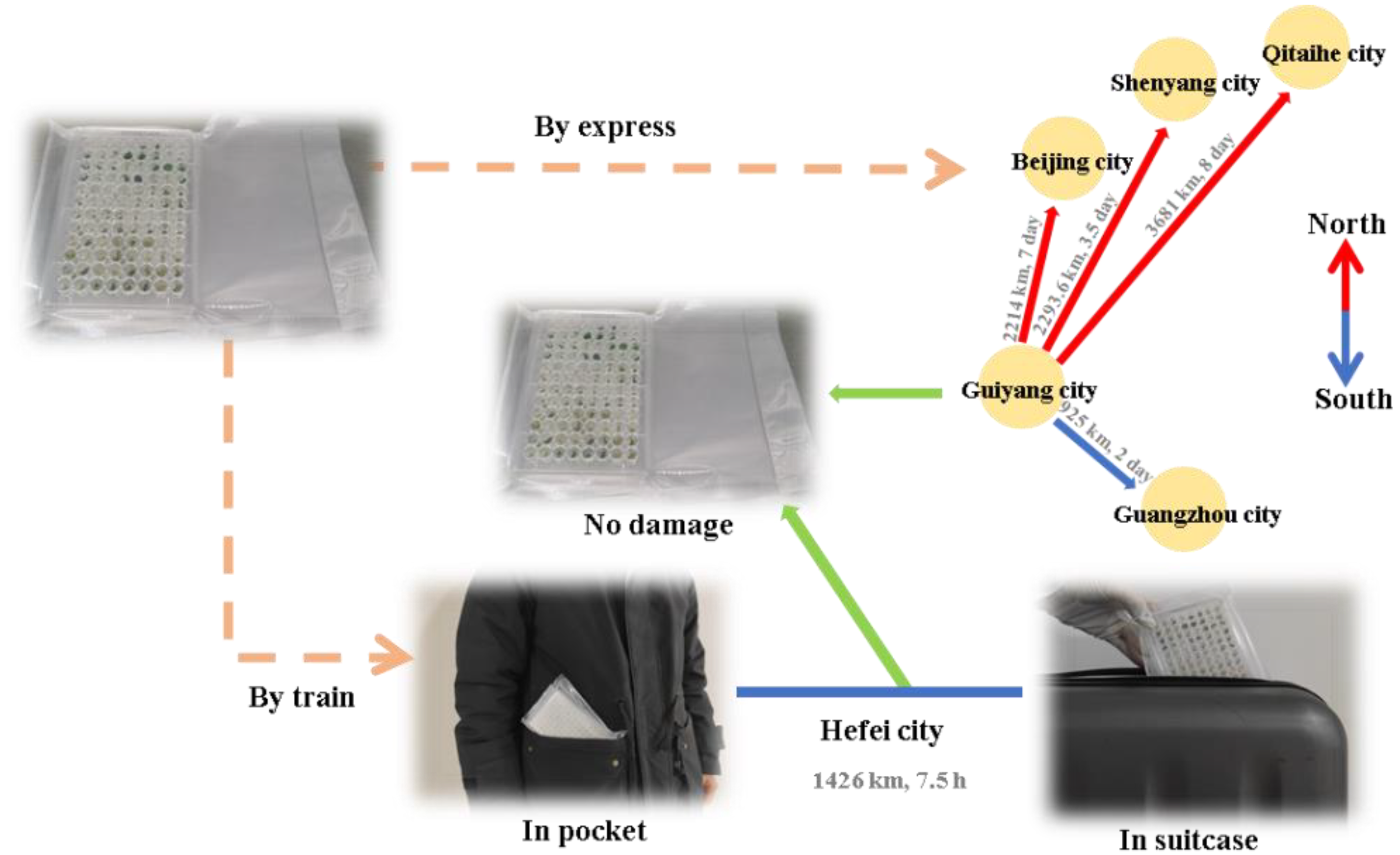
2.7. Transportation Tolerance Test
2.8. Experiment Design and Statistical Analysis
3. Results
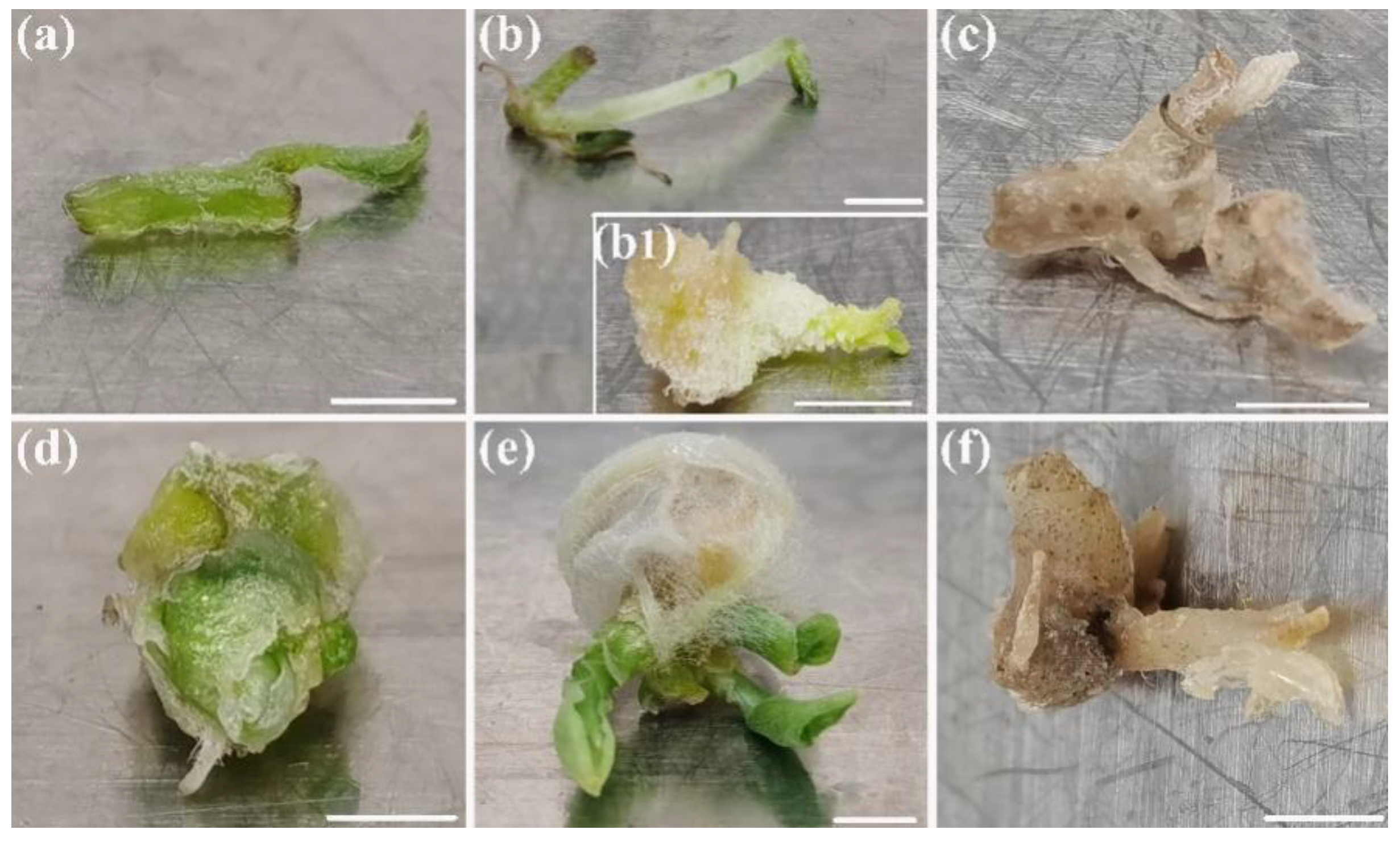
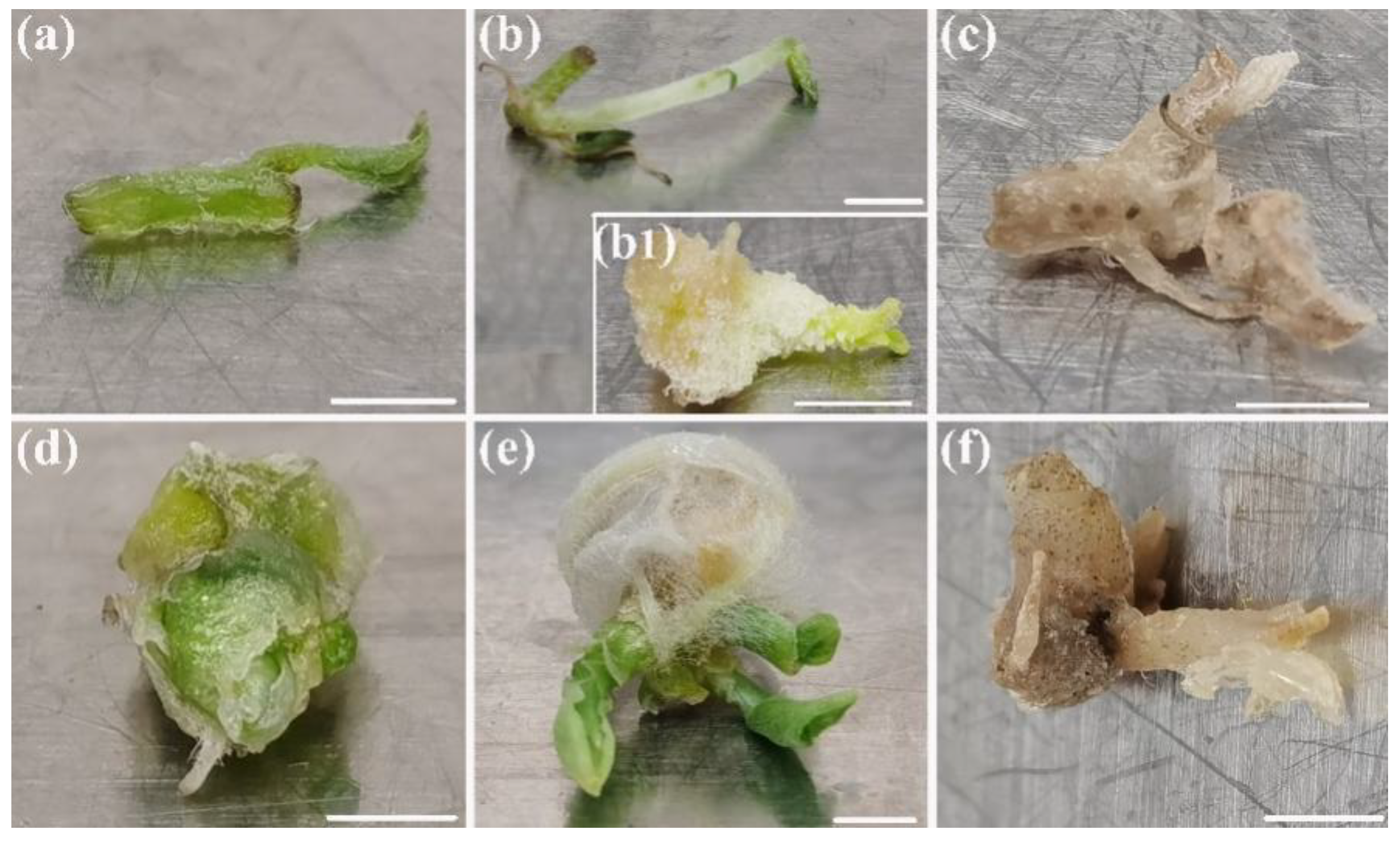
3.1. The Plant Growth and Contamination Response to Package Types
3.2. Plant Growth Response to Incubation Conditions
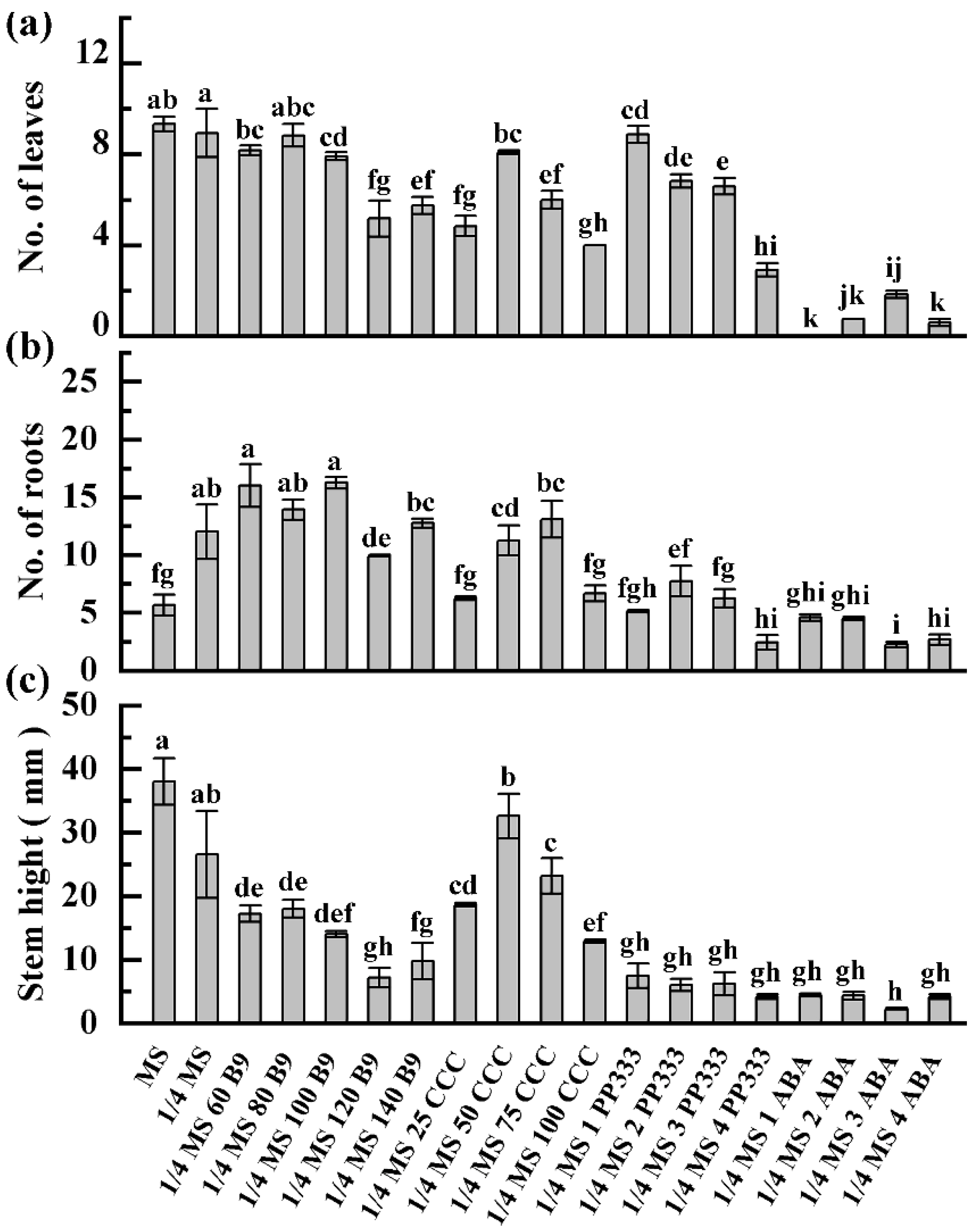
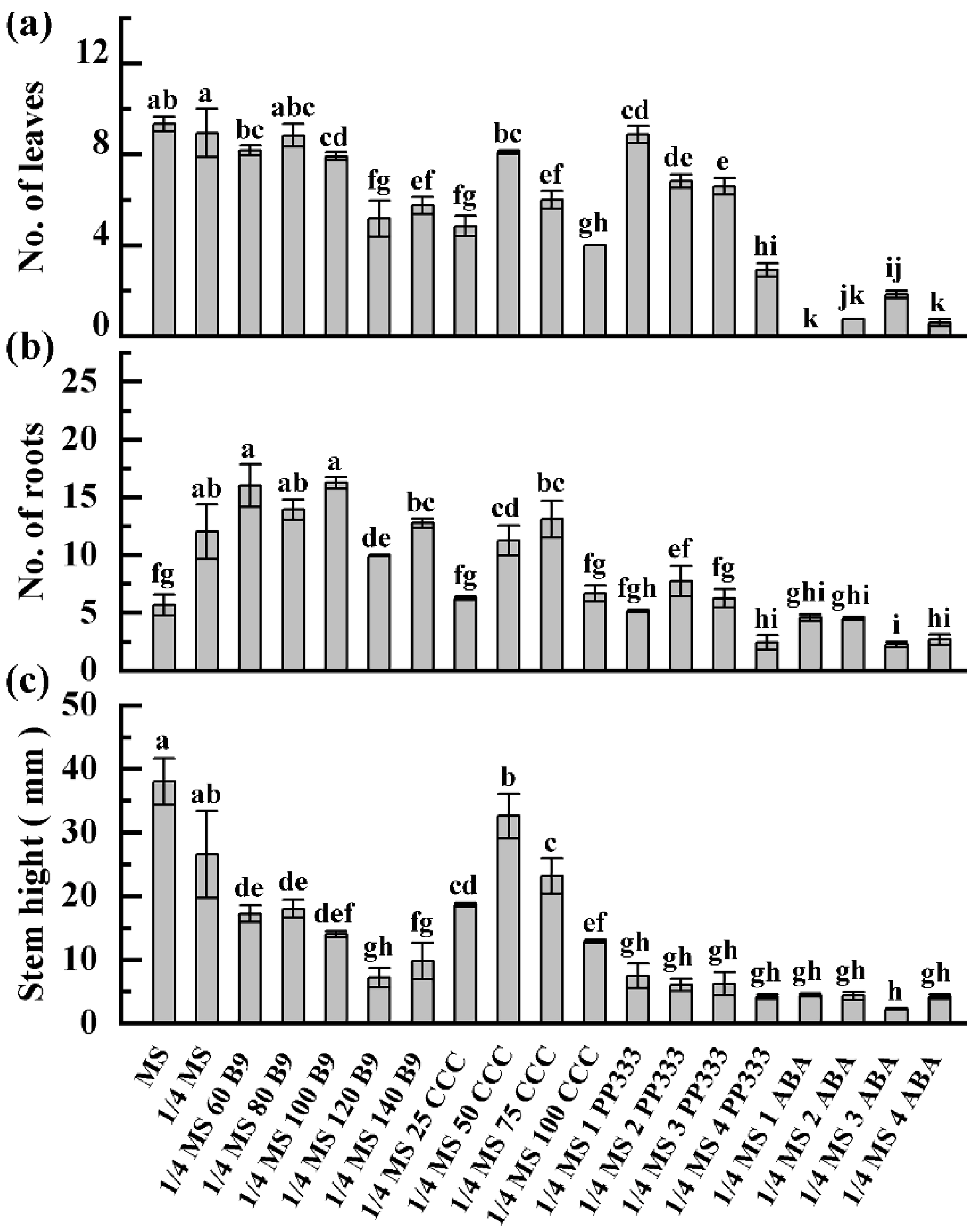
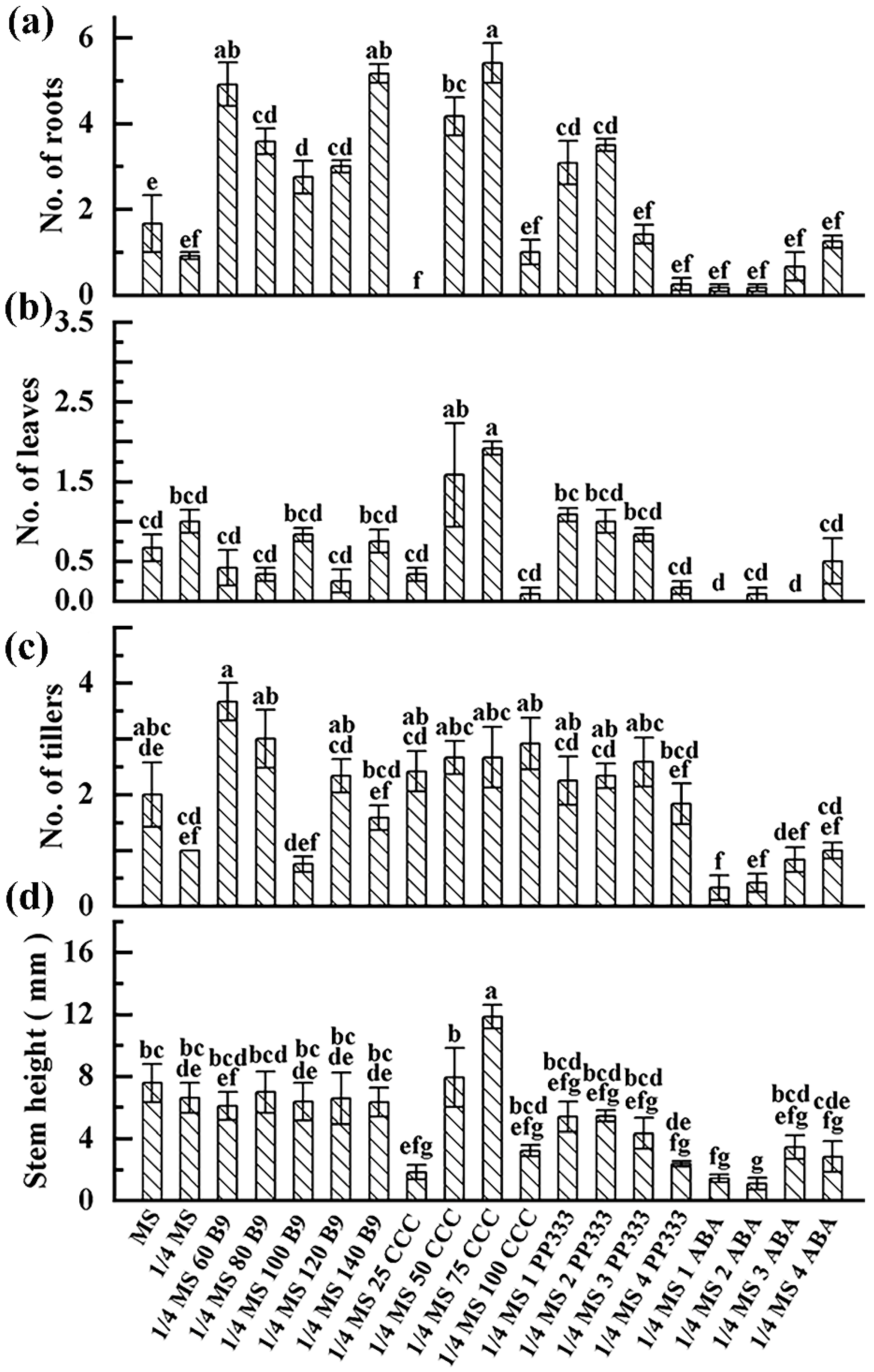
3.3. Plant Growth Response to Slow-Growth Culture Media
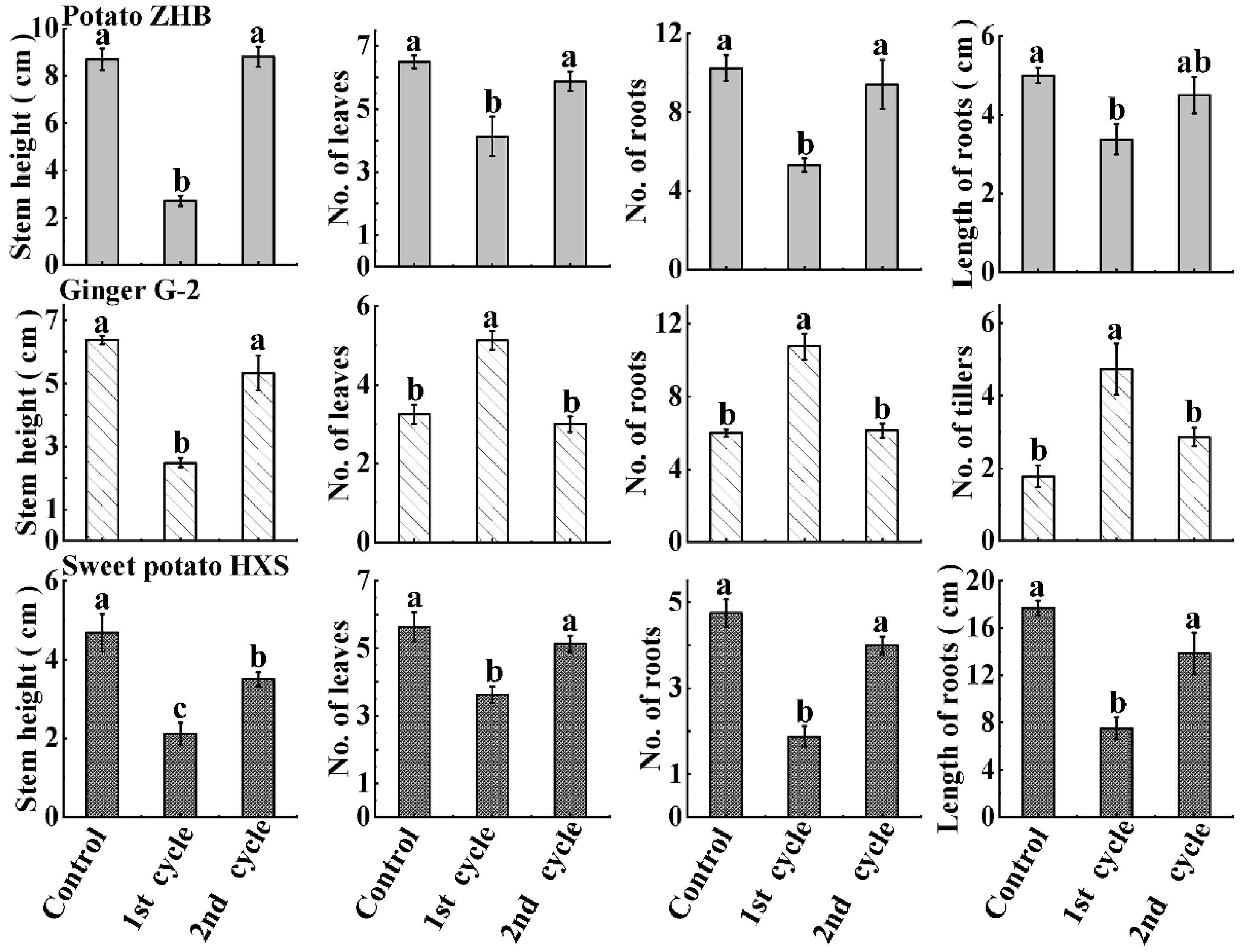
3.4. Regrowth of Packaged Micro-Cuttings
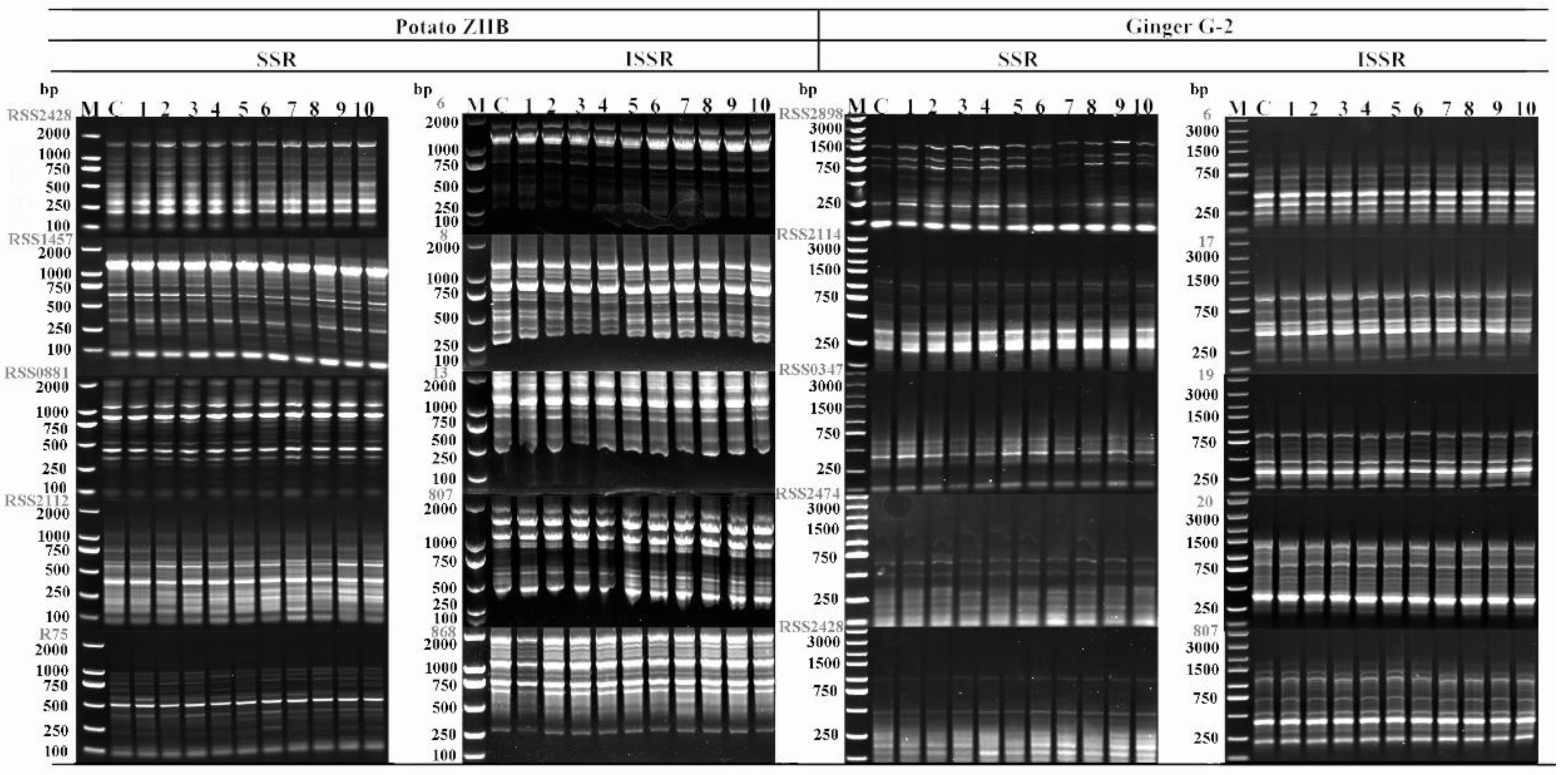
3.5. Genetic Stability
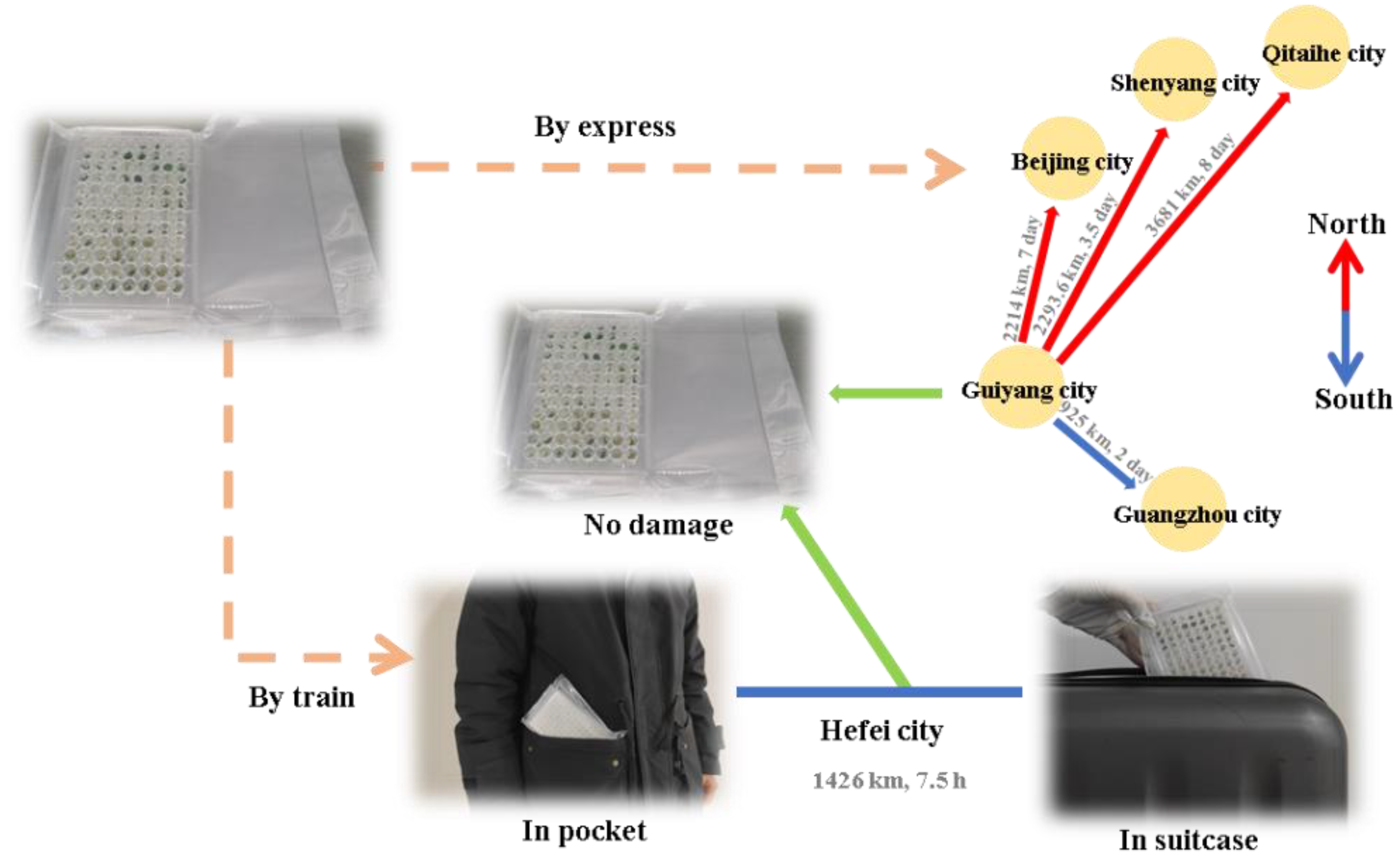
3.6. Transportation Feasibility
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Priyanka, V.; Kumar, R.; Dhaliwal, I.; Kaushik, P. Germplasm conservation: Instrumental in agricultural biodiversity—A review. Sustainability 2021, 13, 6743. [Google Scholar] [CrossRef]
- Chawla, A.; Kumar, A.; Warghat, A.; Singh, S.; Bhushan, S.; Sharma, R.K.; Bhattacharya, A.; Kumar, S. Approaches for conservation and improvement of Himalayan plant genetic resources. Adv. Crop Improv. Tech. 2020, 18, 297–317. [Google Scholar]
- Staub, J.; Chatterton, J.; Bushman, S.; Johnson, D.; Jones, T.; Larson, S.; Robins, J.; Monaco, T. A history of plant improvement by the USDA-ARS forage and range research laboratory for rehabilitation of degraded western U.S. rangelands. Rangelands 2016, 38, 233–240. [Google Scholar] [CrossRef]
- Schiva, T. Bamboo germplasm: Perspectives for ornamental purposes. Acta Hortic. 1999, 486, 255–260. [Google Scholar] [CrossRef]
- Santos, I.R.I.; Salomão, A.N. In vitro germination of zygotic embryos excised from cryopreserved endocarps of queen palm (Syagrus romanzoffiana (Cham.) Glassman). In Vitro Cell. Dev. Biol. Plant 2017, 53, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Chalam, C.V. Elimination of plant viruses by certification and quarantine for ensuring biosecurity. Appl. Plant Virol. 2020, 52, 749–762. [Google Scholar]
- Catană, R.; Mitoi, E.M.; Helepciuc, F.; Holobiuc, I. In vitro conservation under slow growth conditions of two rare plant species from Caryophyllaceae family. Electron. J. Biol. 2010, 6, 86–91. [Google Scholar]
- Reed, B.M.; Sarasan, V.; Kane, M.; Bunn, E.; Valerie, C.P. Biodiversity conservation and conservation biotechnology tools. In Vitro Cell. Dev. Biol. Plant 2011, 47, 1–4. [Google Scholar] [CrossRef]
- Waterworth, P.; Kahn, R.P. Thermotherapy and aseptic bud culture of sugarcane to facilitate the exchange of germplasm and passage through quarantine. Plant Dis. Rep. 1978, 62, 772–776. [Google Scholar]
- Kumar, C.A.; Chandel, K.P.S. Strategy for the production of virus-free plant germplasm under in-vitro conservation and exchange—A brief review. Indian J. Plant Genet. Resour. (IJPGR) 1992, 5, 67–78. [Google Scholar]
- Bekheet, S.A. In vitro culture techniques for conservation of date palm germplasmin in Arab countries. Acta Hortic. 2010, 882, 211–217. [Google Scholar] [CrossRef]
- Negahdar, N.; Hashemabadi, D.; Kaviani, B. In vitro conservation and cryopreservation of Buxus sempervirens L., a critically endangered ornamental shrub. Russ. J. Plant Physiol. 2021, 68, 661–668. [Google Scholar] [CrossRef]
- Filloux, D.; Girard, J.C. Indexing and elimination of viruses infecting yams (Dioscorea spp.) for the safe movement of germplasm. In Proceedings of the 14th Triennial Symposium of the ISTRC, Trivandrum, India, 20–26 November 2006; pp. 1–13. [Google Scholar]
- Reed, B.M.; Paynter, C.; Bartlet, B. Shipping Procudures for Plant Tissue Culture. Available online: https://www.ars.usda.gov/research/publications/publication/?seqNo115=119618# (accessed on 7 January 2022).
- El-Dawayati, M.M.; Zaid, Z.E.; Elsharabasy, S.F. Effect of conservation on steroids contents of callus explants of Date palm cv. sakkoti. Aust. J. Basic Appl. Sci. 2012, 6, 305–310. [Google Scholar]
- Ramírez-Pérez, Y.; Tapia-Campos, E.; Cruz-Gutiérrez, E.J.; Barba-Gonzalez, R. Obtaining a protocol for slow growth for in vitro conservation of eustoma cultivars (Gentianaceae). Acta Hortic. 2020, 1288, 185–188. [Google Scholar] [CrossRef]
- De Lacerda, L.F.; Gomes, H.T.; Bartos, P.M.C.; Vasconcelos, J.M.; Filho, S.C.V.; De Araújo Silva-Cardoso, I.M.; Scherwinski-Pereira, J.E. Growth, anatomy and histochemistry of fast growing species under in vitro conservation through mineral oil and low-temperature combination. Plant Cell Tissue Organ Cult. 2021, 144, 143–156. [Google Scholar] [CrossRef]
- Oseni, O.M.A. Review on plant tissue culture, a technique for propagation and conservation of endangered plant species. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 3778–3786. [Google Scholar] [CrossRef]
- Tavazza, R.; Rey, N.A.; Papacchioli, V.; Pagnotta, M.A. A validated slow-growth in vitro conservation protocol for globe artichoke germplasm: A cost-effective tool to preserve from wild to elite genotypes. Sci. Hortic. 2015, 197, 135–143. [Google Scholar] [CrossRef]
- Tahtamouni, R.; Shibli, R.; Al-Abdallat, A.; Al-Qudah, T. Turkish journal of agriculture and forestry analysis of growth, oil yield, and carvacrol in thymbra spicata l. after slow-growth conservation. Turk. J. Agric. For. 2016, 40, 213–221. [Google Scholar] [CrossRef]
- Fernando, I.; Fei, J.; Stanley, R. Measurement and analysis of vibration and mechanical damage to bananas during longdistance interstatetransport by multitrailer road trains. Postharvest Biol. Technol. 2019, 158, 11097. [Google Scholar] [CrossRef]
- Nikulina, E.O.; Ivanova, G.V.; Kolman, O.I.; Ivanova, A.N.; Perestoronin, D.Y. Research of the influence of vacuum packaging on the quality and safety of meat semi-finished products. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 667, p. 032066. [Google Scholar]
- Jiang, Q.J.; Jin, W.W.; Zhang, W.Y.; Zhang, Z.C.; You, L.F.; Bi, Y.Q.; Yuan, L.M. Analysis of vibration acceleration levels and quality deterioration of Chinese bayberry fruit in semi–vacuum package by express delivery. J. Food Process. Eng. 2021, 44, e13899. [Google Scholar] [CrossRef]
- Anca, B.; Adriana, P.V.; Rodica, Z.; Luiza, M.; Mihaela, P. The results in the field of the in vitro conservation of the cultivars when using classic and modern conservation methods. An. Univ. Oradea-Fasc. Biol. 2007, 14, 23–26. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Monticelli, S.; Gentile, A.; Forni, C.; Caboni, E. Slow growth in â Pisanaâ apricot and in â Cerinaâ. apple, two Italian cultivars. Acta Hortic. 2020, 1285, 117–124. [Google Scholar] [CrossRef]
- Bekheet, S.; Usama, I.A. In vitro conservation of globe artichoke (Cynara scolymus L.) Germplasm. Int. J. Agric. Biol. 2007, 9, 404–407. [Google Scholar]
- Benelli, C.; Carlo, A.D.; Previati, A.; Roncasaglia, R. Recenti acquisizioni sulla conservazione in vitro in crescita rallentata. In Proceedings of the IX Giornate Scientifiche SOI Italus Hortus Conference, Florence, Italy, 10–12 March 2010; p. 91. [Google Scholar]
- Arbeloaa, A.; Marí n, J.A.; Andreu, P.; Garcí a, E.; Lorente, P. In vitro conservation of fruit trees by slow growth storage. Acta Hortic. 2017, 1155, 13. [Google Scholar] [CrossRef] [Green Version]
- Žiauka, J.; Kuusienė, S. Different inhibitors of the gibberellin biosynthesis pathway elicit varied responses during in vitro culture of aspen (Populus tremula L.). Plant Cell Tissue Organ Cult. 2010, 102, 221–228. [Google Scholar]
- Ciobanu, I.B.; Constantinovici, D.; Creţu, L. Influence of genotype and cultivation conditions on vitroplantlets evolution of solanum tuberosum L. local varieties. An. Ştiinţifice Ale Univ. Al. I. Cuza Iaşi. 2011, 2, 13–20. [Google Scholar]
- Indrayanti, R.; Putri, R.E.; Sedayu, A.; Adisyahputra. Effect of paclobutrazol for in vitro medium-term storage of banana variant cv. Kepok (Musa acuminata × Balbisiana Colla). In Proceedings of the 9th International Conference on Global Resource Conservation (ICGRC 2018), Malang, Indonesia, 7–8 March 2018; pp. 020001–020009. [Google Scholar]
- Buldakov, S.A. Use of growth inhibitor chlormequat chloride in potato culture in vitro. In E3S Web of Conferences; EDP Sciences: Les Ulis, France, 2021; Volume 285, p. 03003. [Google Scholar]
- Kumar, S.; Kumari, R.; Baheti, T.; Thakur, M.; Ghani, M. Plant regeneration from axillary bud, callus and somatic embryo in carnation (Dianthus caryophyllus) and assessment of genetic fidelity using RAPD–PCR analysis. Indian J. Agric. Sci. 2016, 86, 1482–1488. [Google Scholar]
- Lizamore, D.; Bicknell, R.; Winefield, C. Elevated transcription of transposable elements is accompanied by het-siRNA-driven de novo DNA methylation in grapevine embryogenic callus. BMC Genom. 2021, 22, 676. [Google Scholar] [CrossRef]
- Kaeppler, S.M.; Kaeppler, H.F.; Rhee, Y. Epigenetic aspects of somaclonal variation in plants. Plant Mol. Biol. 2000, 43, 179–188. [Google Scholar] [CrossRef]
- Li, J.W.; Chen, H.Y.; Li, X.Y.; Zhang, Z.B.; Blystad, D.R.; Wang, Q.C. Cryopreservation and evaluations of vegetative growth, microtuber production and genetic stability in regenerants of purple-fleshed potato. Plant Cell Tissue Organ Cult. 2016, 128, 641–653. [Google Scholar] [CrossRef]
- Mendes, M.; Verde, D.; Ramos, A.; Gesteira, A.; Souza, A. In vitro conservation of citrus rootstocks using paclobutrazol and analysis of plant viability and genetic stability. Sci. Hortic. 2021, 286, 110231. [Google Scholar] [CrossRef]
- Parab, A.R.; Lynn, C.B.; Subramaniam, S. Assessment of genetic stability on in vitro and ex vitro plants of Ficus carica var. black jack using issr and damd markers. Mol. Biol. Rep. 2021, 48, 7223–7231. [Google Scholar] [CrossRef] [PubMed]
- Kandiannan, K.; Prasath, D.; Sasikumar, B. Biennial harvest reduces rhizome multiplication rate and provides no yield advantage in ginger (zingiber officinale roscoe.). J. Spices Aromat. Crops 2016, 25, 79–83. [Google Scholar]
- Marsh, L.; Hashem, F.; Smith, B. Organic ginger (Zingiber officinale Rosc.) development in a short temperate growing season: Effect of seedling transplant type and mycorrhiza application. Am. J. Plant Sci. 2021, 12, 14. [Google Scholar] [CrossRef]
- Liu, X.M.; Yang, G.C. Adventitious shoot regeneration of oriental lily (Lilium orientalis) and genetic stability evaluation based on ISSR marker variation. In Vitro Cell. Dev. Biol. Plant 2012, 48, 172–179. [Google Scholar] [CrossRef]
- Yin, Z.F.; Zhao, B.; Bi, W.L.; Chen, L.; Wang, Q.C. Direct shoot regeneration from basal leaf segments of Lilium and assessment of genetic stability in regenerants by ISSR and AFLP markers. In Vitro Cell. Dev. Biol. Plant 2013, 49, 333–342. [Google Scholar] [CrossRef]
- Yin, Z.F.; Bi, W.L.; Chen, L.; Zhao, B.; Volk, G.M.; Wang, Q.C. An efficient, widely applicable cryopreservation of Lilium shoot tips by droplet vitrification. Acta Physiol. Plant 2014, 36, 1683–1692. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Yaakob, Z.; Anuar, N. Factors affecting in vitro regeneration of Ficus carica L. and genetic fidelity studies using molecular marker. J. Plant Biochem. Biotechnol. 2020, 30, 304–316. [Google Scholar] [CrossRef]
- Gautam, N.; Bhattacharya, A. Molecular marker based assessment of genetic homogeneity within the in vitro regenerated plants of Crocus sativus L.–a globally important high value spice crop. S. Afr. J. Bot. 2021, 140, 461–467. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Vacuum Package | Package Sterilization | Survival (%) | New Tissue Regrowth (%) | Contamination |
|---|---|---|---|---|---|
| Potato ZHB | Vacuumized | + | 87.7 ± 8.4 b | 61.8 ± 6.3 z | − |
| − | 85.8 ± 7.1 b | 62.5 ± 8.8 z | − | ||
| Non- vacuumized | + | 100.0 ± 0.0 a | 100.0 ± 0.0 x | − | |
| − | 100.0 ± 0.0 a | 94.0 ± 3.5 y | − | ||
| Ginger G-2 | Vacuumized | + | 30.0 ± 0.0 b | 0.0 ± 0.0 z | − |
| − | 20.0 ± 0.0 c | 0.0 ± 0.0 z | − | ||
| Non- vacuumized | + | 83.3 ± 3.3 a | 83.3 ± 3.3 x | − | |
| − | 72.4 ± 10.5 a | 67. 0 ± 15.4 y | − |
| Conditions | Potato ZHB | Ginger G-2 | ||
|---|---|---|---|---|
| Survival Rate (%) | New Tissue Regrowth (%) | Survival Rate (%) | New Tissue Regrowth (%) | |
| 25 °C + 16 h light | 35.7 ± 17.1 c | 20.8 ± 8.8 y | 0.0 ± 0.0 d | 0.0 ± 0.0 y |
| 25 °C + dark | 18.0 ± 10.5 c | 0.0 ± 0.0 z | 0.0 ± 0.0 d | 0.0 ± 0.0 y |
| 10 °C + 16 h light | 100.0 ± 0.0 a | 83.1 ± 8.4 x | 100.0 ± 0.0 a | 2.6 ± 0.2 x |
| 10 °C + dark | 100.0 ± 0.0 a | 27.7 ± 11.6 y | 83.3 ± 16.7 a | 2.0 ± 0.2 x |
| 4 °C + 16 h light | 81.4 ± 8.2 b | 14.3 ± 5.0 y | 14.6 ± 3.3 c | 0.0 ± 0.0 y |
| 4 °C + dark | 100.0 ± 0.0 a | 0.0 ± 0.0 z | 35.0 ± 8.3 b | 0.0 ± 0.0 y |
| Mediums | Potato ZHB | Ginger G-2 | ||||
|---|---|---|---|---|---|---|
| Survival Rate (%) | Regrowth (%) | Malformation | Survival Rate (%) | Regrowth (%) | Malformation | |
| MS | 100.0 ± 0.0 a | 100.0 ± 0.0 a | − | 100.0 ± 0.0 a | 100.0 ± 0.0 a | − |
| 1/4 MS | 100.0 ± 0.0 a | 100.0 ± 0.0 a | − | 100.0 ± 0.0 a | 100.0 ± 0.0 a | − |
| 1/4 MS 60 B9 | 91.7 ± 8.3 ab | 91.7 ± 8.3 ab | + | 91.7 ± 8.3 a | 91.7 ± 8.3 ab | + |
| 1/4 MS 80 B9 | 91.7 ± 8.3 ab | 91.7 ± 8.3 ab | + | 100.0 ± 0.0 a | 91.7 ± 8.3 ab | + |
| 1/4 MS 100 B9 | 75.0 ± 14.4 ab | 75.0 ± 14.4 abc | + | 75.0 ± 14.4 ab | 41.7 ± 30.5 abc | + |
| 1/4 MS 120 B9 | 58.3 ± 8.3 cd | 58.3 ± 8.3 abc | + | 91.7 ± 8.3 a | 91.7 ± 8.3 ab | + |
| 1/4 MS 140 B9 | 83.3 ± 8.3 ab | 83.3 ± 16.7 ab | + | 83.3 ± 8.3 a | 83.3 ± 8.3 ab | + |
| 1/4 MS 25 CCC | 100.0 ± 0.0 a | 75.0 ± 25.0 abc | + | 100.0 ± 0.0 a | 91.7 ± 8.3 ab | − |
| 1/4 MS 50 CCC | 75.0 ± 0.0 ab | 83.3 ± 16.7 ab | + | 75.0 ± 14.4 ab | 50.0 ± 0.0 abc | − |
| 1/4 MS 75 CCC | 100.0 ± 0.0 a | 100.0 ± 0.0 a | − | 50.0 ± 0.0 c | 41.7 ± 8.3 abc | − |
| 1/4 MS 100 CCC | 100.0 ± 0.0 a | 100.0 ± 0.0 a | − | 100.0 ± 0.0 a | 91.7 ± 8.3 ab | − |
| 1/4 MS 1 PP333 | 66.7 ± 8.3 ab | 62.5 ± 12.5 abc | + | 91.7 ± 8.3 a | 83.3 ± 8.3 ab | − |
| 1/4 MS 2 PP333 | 66.7 ± 8.3 ab | 75.0 ± 14.4 abc | + | 91.7 ± 8.3 a | 83.3 ± 8.3 ab | + |
| 1/4 MS 3 PP333 | 83.3 ± 8.3 ab | 83.3 ± 8.3 ab | + | 83.3 ± 8.3 a | 58.3 ± 16.7 abc | + |
| 1/4 MS 4 PP333 | 58.3 ± 8.3 cd | 58.3 ± 22.1 abc | + | 91.7 ± 8.3 a | 83.3 ± 8.3 ab | + |
| 1/4 MS 1 ABA | 41.7 ± 8.3 cd | 41.7 ± 8.3 bc | + | 33.3 ± 8.3 c | 16.7 ± 8.3 c | + |
| 1/4 MS 2 ABA | 91.7 ± 8.3 ab | 91.7 ± 8.3 ab | + | 41.7 ± 8.3 c | 25.0 ± 0.0 bc | + |
| 1/4 MS 3 ABA | 58.3 ± 8.3 cd | 58.3 ± 30.1 abc | + | 52.2 ± 4.2 c | 50.0 ± 0.0 abc | + |
| 1/4 MS 4 ABA | 58.3 ± 8.3 cd | 50.0 ± 14.4 abc | + | 41.7 ± 8.3 c | 25.0 ± 0.0 bc | + |
| Sample | Survival (%) | New Tissue Regrowth (%) | Multiplication | |
|---|---|---|---|---|
| 1st Cycle | 2nd Cycle | |||
| Potato ZHB | 100.0 ± 0.0 | 0.0 ± 0.0 | 4.4 ± 0.2 b | 6.9 ± 0.1 a |
| Ginger G-2 | 75.3 ± 6.7 | 0.0 ± 0.0 | 4.3 ± 0.1 a | 2.5 ± 0.1 b |
| Sweet potato HXS | 100.0 ± 0.0 | 6.7 ± 0.7 | 2.4 ± 0.2 b | 3.6 ± 0.4 a |
| Primer Name | No. of Bands | No. of Polymorphic Bands | Primer Name | No. of Bands | No. of Polymorphic Bands | ||
|---|---|---|---|---|---|---|---|
| Potato ZHB | |||||||
| SSR | RSS2428 | 11 | 0 | ISSR | 6 | 9 | 0 |
| RSS1457 | 12 | 0 | 8 | 12 | 0 | ||
| RSS0881 | 14 | 0 | 13 | 8 | 1 | ||
| RSS2112 | 13 | 0 | 807 | 9 | 2 | ||
| RSS75 | 12 | 0 | 868 | 11 | 0 | ||
| Total | 62 | 0 | Total | 49 | 3 | ||
| Ginger G-2 | |||||||
| SSR | RSS2898 | 9 | 0 | ISSR | 6 | 8 | 0 |
| RSS2114 | 9 | 0 | 17 | 11 | 0 | ||
| RSS0347 | 6 | 0 | 19 | 9 | 0 | ||
| RSS2474 | 7 | 0 | 20 | 9 | 0 | ||
| RSS2428 | 6 | 1 | 807 | 10 | 0 | ||
| Total | 37 | 1 | Total | 47 | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; He, M.; Xu, X.; Huang, T.; Tian, H.; Zhang, W. In Vitro Techniques for Shipping of Micropropagated Plant Materials. Horticulturae 2022, 8, 609. https://doi.org/10.3390/horticulturae8070609
Li J, He M, Xu X, Huang T, Tian H, Zhang W. In Vitro Techniques for Shipping of Micropropagated Plant Materials. Horticulturae. 2022; 8(7):609. https://doi.org/10.3390/horticulturae8070609
Chicago/Turabian StyleLi, Jingwei, Min He, Xiuhong Xu, Tingmin Huang, Huan Tian, and Wanping Zhang. 2022. "In Vitro Techniques for Shipping of Micropropagated Plant Materials" Horticulturae 8, no. 7: 609. https://doi.org/10.3390/horticulturae8070609
APA StyleLi, J., He, M., Xu, X., Huang, T., Tian, H., & Zhang, W. (2022). In Vitro Techniques for Shipping of Micropropagated Plant Materials. Horticulturae, 8(7), 609. https://doi.org/10.3390/horticulturae8070609