Abstract
Trunk injection is a targeted delivery of pesticides, insecticides, nutrients, or other plant protection materials into the stem or trunk of woody plants as an alternative to spraying or soil drenching. Trunk injection has historically been used for disease and pest management of high-value forest tree species or ornamental plants when aerial applications are problematic due to spatial problems and health-related concerns. An interest in using the injection technique for protection of agricultural crops in commercial production systems has emerged more recently, where foliar applications and soil drenches have proven ineffective or pose environmental hazards. This review provides an overview of the basic principles of trunk injection and the plant physiological implications, its current use in commercial agriculture and other plant systems, and associated risks.
1. Introduction
Trunk injection is a technique for applying plant protection materials that offers an alternative to foliar sprays or soil drenches. Among the main advantages that trunk injection provides over conventional methods are a higher efficiency of product delivery, reduced risk for worker exposure, reduced risk to the environment, reduced harm to non-target organisms, and the possibility for use in populated areas where other methods are not an option [1,2]. Trunk injection techniques have not been optimized for use in commercial crop production; however, there is a long history of using the method in a variety of crop and non-crop species.
According to the dictionary, the term “injection” is the act or process of forcing a liquid medicine or drug into someone or something, usually by using a special needle. In botany, this term is used in a wider sense and includes any introduction of materials into a plant organ by cutting or through holes with or without force. In this sense, the earliest evidence for plant injection is from the 12th century, when Arabic horticulturists applied perfumes, spices, dyes, and other substances through wounds in plants to affect the smell, color, or other attributes of flowers and fruits [3]. The first documented experimentation on trunk injections occurred in the 15th century by Leonardo da Vinci [3], who injected arsenic and other poisonous solutions through bore holes into apple trees to render the fruit poisonous. Other experimentation until the early 1900s included injection of different nutrient solutions such as ferrous sulphate and ferric chloride as remedies for nutritional deficiencies [3]. This was followed by injection of other inorganic substances such as iron pyrophosphate, potassium cyanide, and aluminum sulphate to control insects and plant diseases [3]. In addition to inorganic materials, organic substances including salicylic acid and plant- and microorganism-derived liquids were injected by various methods for the same purpose. An excellent review of these early studies is presented by Roach (1939) [3], the first comprehensive review on plant injection detailing history, injection methods, and physiology of the injection method known at the time.
Modern research on the use of injection to deliver liquid materials for plant protection and different techniques of injection ignited with the devastating spread of Dutch elm disease (DED) in the 1960s [4]. Many of the experiments during this first wave of renewed interest investigated the injection of fungicides and potassium salts for fungal management [5,6,7,8,9]. In 1978, Michigan State University held the first Symposium on Systemic Chemical Treatments in Tree Culture, followed by a second symposium in 1987. From the 1970s through the 1990s, researchers continued to explore the efficacy of different compounds and conducted mechanism-based research on pressurized injection systems [10], patterns of water and dye movement [11,12,13], wounding, compartmentalization and decay of wood [14,15,16,17,18,19], and different wood characteristics [20,21].
More recent mechanism-based experiments explored wounding and decay [22,23], movement of water and other compounds [24,25], and the use of more effective or less injurious injection methods [26,27]. In 2015, the Tree Care Industry Association held a series of symposia on tree injection, and the following year the International Society of Arboriculture published the Tree Injection Best Management Practices Guide.
Between 2016 and 2021, more than 60 research studies were published describing tree injection-based experiments. Most of these studies tested the efficacy of different chemicals against specific pests or diseases, while few focused on the mechanisms and theories behind trunk injection. The latter included a study by Aćimović et al. (2016) [23], who investigated injection port damage and wound closure in apple trees, Dalakouras et al. (2018) [28], who examined the movement of hairpin and small-interfering RNAs in apple and grape, Killiny et al. (2019) [29], who investigated the uptake and translocation of antibiotics after stem infusion, and Kuroda et al. (2018 and 2020) [30,31], who studied the radial movement of trunk-injected minerals.
A recent review by Berger and Laurent (2019) [32] focuses on modern injection technologies, factors influencing their efficacy, and risks. In this review, we expand on these concepts to include trunk injection as a method to deliver protection materials to agricultural crops to manage pests and diseases, while also discussing the underlying physiological principles and concerns associated with this method of delivery.
2. Trunk Injection Methods
The earliest reported studies on trunk injection involved wounding of roots, branches, and trunks followed by application of different solid substances. Throughout the 19th and beginning of the 20th century, applications of liquids through leaves, branches, roots, and stems through wounds and bore holes in combination with syringes, different tubing systems, or other specially designed devices were explored in many countries throughout the world [3].
More advanced technologies are now available that allow the consistent and rapid injection or infusion of plant protection materials of trees, palms, and woody shrubs. Most of these technologies require the drilling of relatively large-sized holes (4.5–9.5 mm) using pressures of 100 psi (689 kPa) or more. High-pressure injection usually occurs through injectors that are attached via tubing to a portable canister filled with compressed gas (air or nitrogen). Fewer technologies are based on low pressure or passive infusion and do not require a compressed air canister or other peripheral features. Berger and Laurent (2019) [32] provided a tabular overview of some of the available technologies, and few other technologies have been introduced since then. Figure 1 summarizes some of the currently available injection systems.
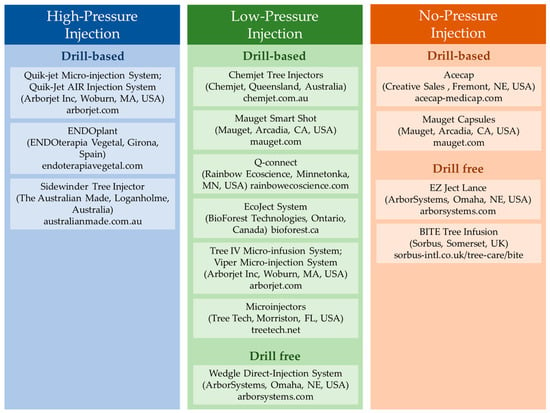
Figure 1.
Some of the currently available trunk injection devices (links accessed 14 June 2022).
2.1. High-Pressure Injection
The Arborjet Quik-Jet Air (Arborjet Inc., Woburn, MA, USA), is one example of a high-pressure application device. Arborjet and similar high-pressure systems, such as ENDOplant (ENDOterapia Vegetal, Girona, Spain), use 7.15 mm or larger diameter plastic plugs as injection ports which are inserted into the tree after drilling of a hole. This creates a tight seal for injection, prevents leaking, and protects the wound from pathogens and insects. Injection of the compounds occurs though specialized metal injection tips at pressures of 60–100 psi (413–689 kPa) created using compressed gas. Although the plastic plugs enable the rapid injection of large volumes of material, they can cause more damage to trees than no-plug methods, as they increase the size of the injection hole, increase the probability of injury from bark cracking, and may interfere with full wound closure [23]. Dendrology research has shown that plugging tree core wounds does not provide any benefit and can even interfere with the natural healing capabilities of the tree [33]. Another high-pressure system is the Sidewinder tree injector (The Australian Made, Loganholme, Australia).
2.2. Low-Pressure Injection
An alternative to high-pressure injection are syringe or needle-based methods sold by Chemjet (Kerrville, TX, USA), Mauget (Arcadia, CA, USA), Rainbow Ecoscience (Minnetonka, MN, USA), and other manufacturers, which allow the plug-free injection of materials. Injection using these devices occurs at relatively low pressures (<60 psi) by manual squeezing or use of a spring-loaded syringe system. Most of these systems rely on drilled holes prior to application; however, ArborSystems (Omaha, NE, USA) has developed a low-pressure drill-free system, the Wedgle® Direct-Inject. With this device, a shallow stem core is removed, and liquid is manually pushed into the trunk. The rate of liquid uptake associated with lower-pressure devices is slower than that of higher-pressure systems but eliminates the additional step of placing an injection port prior to injection. This may make these techniques more efficient at the commercial scale and allows injection in smaller trees.
2.3. No-Pressure Injection
No-pressure systems can also be either drill-based or drill-free. Implanted capsules, such as the Acecap (Creative Sales Inc., Fremont, NE, USA) or Mauget capsules (Mauget, Arcadia, CA, USA), are inserted into a drilled hole. Uptake of materials is through the transpiration-driven natural transport of sap in the tree. A similar drill-free system is the EZ Ject Lance (ArborSystems, Omaha, NE, USA), which pushes shells that contain the chemical underneath the bark of trees via a spring-based mechanism, and the chemical is then dissolved into the trunk through the water-soluble cap. However, this system was not designed for tree protection but rather for the targeted delivery of herbicides. A trial using this system to control apple scab did not demonstrate improved efficacy over aerial applications of salicylic acid or Actigard [34].
The BITE Tree Infusion tool (Sorbus, Frome, Somerset UK) creates a narrow lenticular-shaped hole designed to minimize tree damage. The injector is mounted horizontally in the tree using a built-in sliding hammer. Injection occurs through a syringe placed on top of the device, and liquid can be injected with manual pressure or by gravity-based infusion in combination with the natural vascular movement of sap in the tree. One of the expected benefits of this injection system is a faster closure of the narrow lenticular-shaped injection wound than the larger drill-induced wounds inflicted with other technologies [27].
Although external wounding may be minimized, the use of no-pressure injection and gravity-based infusion is likely too slow to be of any practical value in large-scale commercial crop production systems [35]; however, these devices may be practical on a smaller scale. Aćimović et al. (2016) [23] documented wound closure and healing after application of different injection technologies, including the BITE system and the Arborjet Quik-Jet Air (Arborjet Inc., Woburn, MA, USA), in apple trees. Apart from this study, little research has been conducted to study wounding effects associated with trunk injections.
3. Physiological Principles of Trunk Injection
3.1. Tree Anatomy
The movement of crop protection materials after trunk injection varies depending on environmental conditions and the physiological status of the tree. In general, conditions where the vapor pressure deficit is high and trees are metabolically active have a positive effect on the speed by which injected materials are taken up and the maximum injectable volume is achieved [36,37]. In addition, different tree anatomical features, such as the size and arrangement of xylem vessels, tracheids, and vessel parenchyma cells, determine the path and efficiency by which materials are being distributed throughout the tree [38]. Recent research suggests that the radial movement of minerals after injection occurs through a combination of active transport by parenchyma cells and diffusion through cell walls [30].
Hardwood species, which includes most agricultural tree crops, can be classified into diffuse-porous and ring-porous species, although these terms are not absolute. In ring-porous species, the largest pores or xylem vessels are in the earlywood because most cambial growth occurs during the earlier period of the growing season [39]. Smaller pores are found in the latewood formed in the later period of the growing season when cambial activity is reduced [39]. In diffuse-porous wood, xylem vessels are more evenly distributed and more uniform in size, with less distinction in the growth rings typical for ring-porous species. The arrangement and size of the vessels in the trunk determines the patterns of compound uptake and distribution, as well as wound response and compartmentalization. Sinclair and Larsen (1981) [20] created an index for “injectability” by determining the relative frequency and integration of xylem vessels in a cross-section divided by the specific gravity of the wood in that tissue.
Differences in vascular integration and sectoriality among species can be useful in predicting the potential efficacy of injections [40,41,42,43,44]. Highly sectorialized species have a more limited capacity to move liquids laterally across the trunk, preventing their even distribution throughout the tree [45]. In contrast, highly integrated species have a greater capacity to distribute injected materials rapidly and evenly. Some trees show a spiral distribution of liquid materials along the tree trunk [12], but the patterns of spiral movement can vary considerably across species and even between individuals of the same species [12]. Determining the vascular integration of crop species and individual cultivars is therefore necessary for predicting the distribution of injected compounds and developing recommendations on the spacing and number of injection ports required for the most effective application.
The level of vascular integration also determines the ability of different species to recover after wounding. Highly integrated trees can create transport pathways around blocked or embolized vessels more effectively than less integrated trees; however, these species may also be more likely to introduce decay-causing organisms in new areas of the trunk [46,47].
In addition to differences in vascular anatomy, differences in the site of injection on the tree can affect the rate of uptake and distribution of materials [45]. For example, root injection of dye in elm trees resulted in movement of dye to the xylem vessels of the current year’s growth, while stem injection resulted in the distribution across multiple xylem layers [11]. In contrast, in a study on nine different forest tree species, dyes injected into the lower trunk of the trees moved radially throughout the entire root tissue, while in the stem the dye was confined to the most recent growth [24]. Injections into the root flare caused a significant amount of dye to move downward and across the entire root xylem, while injection into the stem caused most of the dye to move upward, although this response was seasonal. Injection in the fall resulted in more downward movement compared to injection in other seasons [24]. Additional information on the uptake and distribution of compounds as a function of the injection location on the tree is required for individual crop species before recommendations for best injection practices can be developed.
3.2. Cohesion-Tension Theory
A fundamental concept for understanding the mechanisms of trunk injection is the cohesion-tension theory. The theory was first documented by Dixon and Joly (1894) [48], who suggested that sap will rise in a plant due to a suction-like force caused by the transpiration of water on the leaf surface. The resulting tension on the water in the tracheary and/or vessel elements of the xylem continues into the roots as a functionally continuous column. The cohesion of the dipolar water molecules creates the high tensile strength necessary to allow formation of continuous water columns from roots to leaves [49,50].
Since xylem sap is physically in a metastable state [51,52], the introduction of air into the vessels causes cavitation [53]. Cavitation can occur naturally, for example during periods of drought, when the water tension in an individual vessel becomes so high that air is introduced in the water column. This breaks the continuity of the water, and air expands to fill the length or a portion of the column [53], embolizing the xylem vessel and rendering it non-functional. To reduce the risk of rampant cavitation, xylem vessels do not span the length of the tree as one individual column [54]; rather, vessel elements are stacked end-to-end and are separated by a perforated endplate. The perforation plates joining vessel elements do not cause resistance to water movement but act as barriers to contain cavitation when water is in a negative-pressure (high-tension) state. The strong surface tension created by the cohesion of water blocks incoming air and prevents it from nucleating adjoining vessels [55].
Understanding water hydraulics is important for understanding the mechanism of uptake and movement of liquids when using trunk injection to apply crop protection materials. Drilling or boring holes into the trunk for injection causes xylem vessels to embolize, rendering them non-functional. Additional research is necessary to determine the mechanisms by which trunk-injected liquids are taken up and distributed through the tree using high-, medium-, or no-pressure systems, and whether injection-induced embolization will reduce hydraulic functioning over time.
3.3. CODIT
The concept of CODIT Compartmentalization Of Decay In Trees) was developed by Shigo and Marx (1977) [14] and is based on the idea that trees are highly sectorialized organisms able to compartmentalize injuries or wounds after they occur to prevent large-scale damage. Understanding how different crop species compartmentalize wounds is important to assess the feasibility of using trunk injections for systemic delivery of therapeutic materials to manage pests and diseases.
Based on the CODIT concept, after an injury, trees respond by blocking the decay-causing organisms, mostly fungi and bacteria, from moving horizontally or vertically through the formation of four walls. Walls 1–3 comprise the reaction zone, where lignin formation protects against further oxidative degradation [56]. Wall 4 is the barrier zone. The walls are ordered numerically by their order of effectiveness, with wall 1 being the weakest and wall 4 the strongest. Wall 1 prevents the vertical (axial) spread of decay. Axial decay is the most difficult to contain because bacteria and fungi introduced into the vascular system can be transported rapidly along the transpiration stream. After wounding, compounds such as tyloses and gels accumulate in affected vessel elements to limit the spread of vertical decay [57]. The hydrophobic nature of these materials prevents fluid diffusion, forming the upper and lower boundaries (wall 1) of decay [58]. The speed at which vessel elements plug determines the length of the occluded wood. This varies by species and vascular anatomy [59], size and severity of the wound, time of year, position of the wound, and environmental conditions [14].
Wall 2 prevents radial decay (towards the pith). Spread of decay towards the pith is blocked by the suberization of xylem parenchyma cells, occlusion of xylem vessels, and, in ring-porous species, a static barrier formed by the highly lignified cell walls of fibers found at the growth-ring boundary [14,60]. The compounds formed by vessel-associated parenchyma cells to cause occlusion of xylem vessels vary across species but may include suberized tyloses or other fibrillar or amorphous deposits, gum secretions, and phenolic compounds [61,62,63,64]. In ring-porous species, such as most temperate perennial crops, the latewood vessels located along the outer edges of growth rings are both smaller and have thicker walls than earlywood vessels, typically resulting in more effective occlusion and blockage of decay [39,65].
Wall 3 prevents the tangential (left and right) movement of decay and is formed by the ray parenchyma cells that are arranged radially in the tree trunk [60]. The ray parenchyma cells are the first to respond to wounding and thus have the highest capacity to compartmentalize and prevent decay from girdling the tree [66]. In response to wounding, these cells can relatively quickly lignify and suberize to become impervious to microbial penetration [60].
Cambium cells surrounding the wounding site are responsible for generating the strongest wall, wall 4, also called the barrier zone, which separates the wood formed before wounding from the new wood formed after wounding [14]. Cambium injury initiates suberization and lignification to prevent decay from spreading into the newly created wound wood [67]. Avoiding injury to the cambium is therefore important for preventing spread of injury and decay. Nevertheless, some injection devices were designed with the specific purpose of delivering liquids just beneath the bark and using the separation zone between bark and wood as a reservoir for the injected liquid [65].
Understanding CODIT and how different tree species respond to wounding is essential for developing trunk injection methods suitable for commercial agricultural systems. This is especially important in systems where treatments may need to be applied at least annually. Most published studies that investigated wounding and compartmentalization, however, were conducted on forest tree species and tree species growing in temperate climates [15,17,19,23,68,69]. When applying injection technologies to agricultural crops, the anatomical and physiological responses of the crop of interest need to be established before application on a commercial scale.
4. Physiological Implications of Trunk Injection
Using large injection ports and/or higher pressure to force liquids into a tree can increase the rate of uptake during injection compared to small ports and lower pressure. Both factors will also influence the physiological effects and damage inflicted on the tree.
4.1. Wound Size: Drilled-Based vs. No-Drill Injection
The principles of CODIT can be applied to predict the possible consequences of different types of injection. A sharp drill will cut perpendicularly through the xylem vessels and remove a core of wood [23], which exposes a relatively large area of the xylem, therefore improving the uptake and distribution of injected materials. Although uptake may be improved with deep, drilled holes, the inflicted damage is more extensive than the damage inflicted by needle- or blade-like injection devices [23]. Deeply drilled wounds will cause more damage to the inner wood, which is metabolically less active, and therefore, compartmentalization of decay is slower than in the outer wood [70]. This results in greater discoloration and more extensive decay towards the center of the tree compared to the periphery [71].
Blade- or “lenticular”-shaped devices are drill-free options that are inserted into the trunk so that their longest edge is oriented axially and parallel to the fibers of the xylem vessels [27]. These injection devices have been developed to take advantage of the strong compartmentalization capacity of ray parenchyma cells [60] and to minimize damage and decay in the tangential direction and reduce the risk of girdling. These devices are inserted shallower into the trunk than drills, which also reduces the extent of radial damage [27].
Needle-based devices are another alternative to drill-based devices which may accelerate wound closure by reducing the extent of physical damage to bark, cambium, and xylem tissue. These devices were designed to deposit the injected liquid at the xylem/bark interface near the cambium for a gradual and controlled uptake [65,72]. However, this methodology can cause the separation of the bark from the xylem, which may promote cambial decay across the separation zone [65]. Most of the drill-based devices that have been developed are specifically designed to reduce the occurrence of bark separation and minimize cambial dieback.
In large forest trees, annual trunk injections through ports less than 25 mm in diameter are unlikely to impact tree growth, and wounds under 12 mm in diameter generally heal over in less than one year [18]. In agricultural tree crops, the risks associated with large drill-induced wounds are considerably greater as they may lead to partial or full girdling of the trunk or branches, especially after multiple applications [4]. Smaller-diameter injection devices are therefore preferable to reduce the risks associated with injection. Although drills cause the most wounding, they have been most widely adopted because liquids can be delivered more rapidly and in larger quantities than when using blades or needles [23].
4.2. Pressure: High-Pressure vs. Low- or No-Pressure Injection
Pressurized devices allow for relatively large quantities of product to be injected into a tree in a short period of time. These devices also distribute products more evenly throughout the tree compared to the non-pressurized devices [73]. However, the extent of the physiological damage inflicted on the tree when forcing liquids through embolized or inactive vessels needs to be established for each crop system.
Implant-based systems are no-pressure devices inserted into a drilled hole and left in place while they slowly release compounds into the tree. Implants reduce the amount of time and labor needed for injection because devices can be left in the tree indefinitely. Potential phytotoxic effects of chemicals delivered with these methods may be more severe because of the extended contact time between plant tissue and the injected chemical, which can lead to an increased likelihood of damage at the injection sites [7].
No-pressure systems increase the risk of phytotoxicity and limit the amount of compound that can be applied, while high-pressure systems increase the risk of cavitation and usually require relatively large injection ports. Technologies that reduce the size of injection ports while still effectively delivering plant therapies may be necessary before trunk injection can be widely adopted in crop production systems. Moreover, the labor and associated costs of injection relative to the potential benefits need to be established for each production system before pursuing its use on a large commercial scale.
5. Trunk Injection to Control Pests and Diseases
Historically, trunk injection has been explored as a curiosity and to establish the physiological principles of sap movement in plants, and later to remedy nutrient deficiencies and manage pests and diseases, including fungal and bacterial pathogens, insects, and nematodes. Modern research has generally followed a path that coincides with outbreaks of catastrophic tree diseases or pests.
5.1. Fungal and Oomycete Diseases
Modern research on trunk injection began in response to the catastrophic spread of Dutch elm disease (DED) [36]. DED is a vascular wilt disease that can affect many species of elm, but native European elms (Ulmus glabra, U. laevis, and U. minor) and American elm (U. americana) are especially susceptible [74]. The disease is caused by fungal species in the genus Ophiostoma, which occupy roots and stems of living elm trees. The spores of the fungus are vectored by beetles that feed on the bark. These spores germinate in the xylem and produce mycelia which spread systemically to the stems and roots tissues [75]. The proliferation of the fungus causes the xylem to clog, rendering it non-functional. Vulnerable trees may die within one year of becoming infested [75]. Injecting fungicides into the trees can be performed as preventative measure to create an environment that is inhospitable for fungal growth, or as a curative measure to stop fungal growth after infestation [8].
Although tolerant elm cultivars are now available [76], DED remains a problem, especially in older trees. Different injectable protection materials have been developed that offer some protection [77]. Registered fungicides include propiconazole (e.g., Arborsystems ‘Sheperd’ or Rainbow Treecare ‘Alamo’), thiabendazole hypophosphite (Rainbow Treecare ’Arbotect 20S’), or a biocontrol product from the conidiospores of a nonpathogenic strain of the fungus Verticillium albo-atrum (Dutch Trig®, PJ Apeldoorn, The Netherlands). Protection and application of the latter has shown efficacy at reducing the impact and occurrence of DED [78]. However, most materials will not provide retroactive control if more than 20% of the tree canopy is affected [79].
As seen with DED management, trunk injection for management of diseases is still primarily utilized for forest and ornamental trees. Commercial avocado (Persea americana) production is one example of an agricultural crop production system that has relied on trunk injection to manage fungal diseases for decades [80]. Initially, fungicide injections were conducted to control Phytophthora root rot caused by Phytophthora cinnamoni. Darvas et al. (1984) [6] found that fosetyl-Al injected twice per season significantly reduced the pathogen in the roots and greatly improved tree health, though some phytotoxicity was seen in foliage. There was little improvement of tree health after the first year of injection, but dramatic improvement occurred in years 2 and 3 [6]. More recent experiments in avocado have studied a variety of new products to expand management options for Phytophthora root rot. For example, injection of potassium phosphite in combination with soil-applied fungicides reduced P. cinnamomi severity and improved tree health [81]; in contrast, another study found no effect of potassium silicate injections [82].
Trunk injection has been used in Australia and South Africa for avocado production since the 1980s [64] but was initially not practiced much in the United States. The spread of laurel wilt (LW) across much of the Southeastern USA, however, has generated interest in trunk injection as an alternative management practice [83]. Laurel wilt is caused by the fungus Harringtonia lauricola (basionym: Raffaelea lauricola) [84,85,86] and vectored by a number of ambrosia beetle species, including Xyleborus glabratus and X. bispinatus [87,88,89]. Many species in the Lauraceae family are susceptible, including redbay (P. borbonia) and sassafras (Sassafras albidum), as well as avocado [90,91]. Propiconazole has been used for DED and LW management. Others such as thiabendazole have been used in DED management and registration of tebuconazole for LW management is in progress for H. lauricola [92]. Since 2014, approximately 20% of the commercial avocado acreage in Florida has been injected with propiconazole prophylactically on a 12- to 24-month basis to prevent (suppress) the laurel wilt pathogen [91].
The efficacy of trunk injection for management of a diverse number of fungal pathogens in both crop and non-crop species continues to be investigated. These include: potassium phosphite and micronutrient mixes to treat Chestnut ink disease [93], thiabendazole, prochloraz, allicin, prochloraz + tetraconazole, and abamectin (insecticide) to treat thousand cankers disease in walnut [94], fosetyl-aluminum to treat root rot causing oomycetes and phytophthora in oak [95,96], the fungicide pyraclostrobin to treat the grapevine trunk disease Esca [97], triazoles for protection of grape vines against powdery mildew [98], and phosphonate fungicides and phosphites to treat phytophthora in avocado [99,100]. For management of Verticillium wilt in olives, researchers also investigated the use of fosetyl-aluminum, benomyl, thiophanate methyl, carbendazim, copper sulphate, and dodine, though results were inconsistent between experiments [101,102].
5.2. Insect Pests and Nematodes
The devastating outbreak of the emerald ash borer (EAB), Agrilus planipennis, was one impetus for the advancement of injection technologies to systemically target insect pests in the early 2000s [103,104,105,106]. The EAB affects multiple species of North American ash trees (Fraxinus spp.). The insect is native to parts of Asia and was first discovered in 2002 in North America, where it has rapidly spread across the eastern half of the United States and multiple provinces in Canada (http://www.emeraldashborer.info/documents/MultiState_EABpos.pdf; accessed on 14 June 2022). The insects burrow and feed beneath the bark and cause close to 100% mortality of ash trees when left untreated; however, several insecticide formulations have proven effective at reducing infestation.
Emamectin benzoate (e.g., ‘Tree-age’ by Arborjet, or ‘Mectinite’ by Rainbow Tree care) and imidacloprid (e.g., ‘Pointer’ by Arborsystems, or ‘Imicide’ by Mauget) are two insecticidal products that reduce larval activity of EAB when applied as a foliar spray and have since proven effective when applied through trunk injection [105]. In one trial, imidacloprid injections provided good control of EAB beetles in the first year after injection, but the efficacy of the product decreased in the second year [105]. In contrast, emamectin benzoate showed efficacy against the EAB larva for up to three years [107]. Due to the multi-year efficacy of some insecticidal products, economic analyses have indicated that the costs associated with injections to protect against EAB are ultimately less than the removal of infected trees [108].
In addition to reducing environmental impacts, such as leaching or damage to off-target organisms, one benefit associated with trunk injection over soil drenches or spray applications is the more rapid uptake and distribution of compounds [1]. While soil drenches may require 4–6 weeks for the chemical to be taken up and distributed, trunk-injected compounds can be applied 1–2 weeks before the expected onset of beetle emergence [109]. Like conventional insecticide applications, the timing of trunk injections will significantly impact their efficacy [109]. Protection against EAB using trunk injection has proven to be an effective option for managing both larvae and adult insects. One challenge with managing EAB, however, is that there are hundreds of millions of ash trees growing in forests across the United States and eradicating the insect or protecting a large percentage of ash trees is therefore difficult.
The pinewood nematode (Bursaphelenchus xylophilus) vectored by the pine sawyer beetle (Monochamus spp.) is the causal agent of the devastating pine wilt disease, which although of global impact, has become a highly destructive pest in many Asian countries [110]. Emamectin benzoate emerged as a potential candidate for management of the disease because it has strong nematocidal activity against B. xylophilus and can also diminish M. gallo-provincialis populations [110,111]. Preventative, though not curative, injections of abamectin, cyhalodiamide, and some naphthoquinones have also been shown to be effective for nematode management [72,112,113].
The successful injection of insecticides for EAB management and of nematicides for pinewood nematode management spurred studies on commercial crop species to investigate the potential for insecticide injection to manage pests and diseases. The efficacy of insecticide injections in reducing a variety of insect pests has been studied most extensively in apple trees [2,23,114]. Like the success of the compounds for EAB management, emamectin benzoate and imidacloprid both controlled the oblique-banded leafroller and potato leafhopper at certain concentrations [2], and other compounds such as dinotefuran and abamectin also showed some efficacy [115]. Limitations to widespread adoption of injection for insect management in commercial crops remain due to the time and labor costs associated with this delivery method.
The efficacy of trunk injection for management of a diverse number of insect pests continues to be investigated. Examples are the woolly adelgid in eastern hemlock [116], the oak borer [117], the leaf gall wasp in Chinese banyan [118], the Asian long-horned beetle in willows [119], lobate lac scale in banyan [120], the nettle caterpillar in sweet olive [121], eriophyid mite in oxhorn bucida [122], horse chestnut leaf miners [123,124], and the shot-hole borer in sycamore [125,126].
Exotic species continue to be introduced to new environments where they face few natural predators and can spread rapidly. The use of trunk injection for preventative or curative management has shown potential for protecting high-value forest, ornamental, and landscape trees from pests and diseases, perhaps justifying its use in crop production systems. Recent research on insecticide injection in tree crops includes emamectin benzoate for shot-hole borers in avocado [127], azadirachtin and abamectin for pear psylla in pear trees [128], abamectin for tiger longicorn beetle in mulberry [129], imidacloprid, oxydemeton-methyl, thiamethoxam, and fertilizers for leopard moth control in Persian walnut [130], abamectin to manage walnut husk fly in walnut trees [131], imidacloprid and spirotetramat for banana thrips [132], emamectin benzoate, imidacloprid, and thiamethoxam to control the red palm weevil in date palms [133,134], and thiamethoxam to manage apple aphids in apple trees [135].
5.3. Bacterial and Phytoplasma Diseases
Bacterial diseases can be especially difficult to control because bacteria reproduce rapidly, can be spread by wind, rain, insects, or other animals, and often reside within plant tissues such as leaves, the inner bark, or the vascular system. Injections of antibiotics to control bacterial and phytoplasma pathogens as an alternative to aerial applications have been attempted [136,137,138] and are preferable as less product is needed for effective management and the risk to human health is diminished [2].
Fire blight is a devastating bacterial disease of apples, pears, and other pome fruits caused by Erwinia amylovora. The bacteria infect blossoms, fruits, or vegetative tissues and spread systemically through the rest of the tree [139], while movement between trees by insects or rain is also possible. The most effective method for management during the blossom phase of the tree is with the use of antibiotics, and spray applications of streptomycin became the most widely used method to protect the flowers [140]. However, the continued use of streptomycin led to a significant increase in resistance of E. amylovora in different orchards in the USA [141,142]. In the 1980s, growers began using oxytetracycline to combat the development of resistance to streptomycin. However, oxytetracycline is bacteriostatic and thus generally less effective compared to the bactericidal streptomycin when used on sensitive strains of E. amylovora [139]. When used as foliar sprays, both streptomycin and oxytetracycline are most effective at preventing the growth of bacterial populations when they are applied prior to a rain event. However, oxytetracycline is very sensitive to degradation by sunlight and can be easily washed off the leaf surfaces during rain events [143]. Due to these limitations, researchers have begun exploring the efficacy of trunk-injected antibiotics for fire blight management [144,145].
Currently, the most widespread use of bactericide delivery by trunk injection is for control of lethal-yellowing-type phytoplasmas in palms [146]. Lethal yellowing (LY), and more recently, lethal bronzing (LB), are two genetically distinct phytoplasmas with detrimental impacts on a variety of palm species. The phytoplasma Candidatus Phytoplasma palmae is a phloem-limited pathogen and is transmitted most extensively by the planthopper Haplaxius crudus [147]. LY was first identified in Florida in the 1970s and the original strain was most devastating for coconut palms, whereas it generally did not affect native species [148]. Oxytetracycline hydrochloride has been used both preventatively and curatively for management of LY in palms and has only shown efficacy when applied through injection as opposed to soil drenches and foliar sprays [149]. The more recently identified phytoplasma causing LB affects a broader range of palm species, including many that are native to the Southeastern USA [146]. Oxytetracycline injections prior to infection have proven effective for LB management, though it is unclear if oxytetracycline is effective against LB once symptoms have developed [146].
Although the use of trunk injections for widespread containment and management of LY-type phytoplasmas in palms can be used as a model for the potential utility of this delivery method, the benefits associated with injection in palms may not be the same for woody trees. Palms are monocotyledonous plants in the Aracaceae family. The structure and arrangement of the xylem and phloem vessels in the trunk of palms are therefore fundamentally different from true tree species, likely affecting the distribution of injected chemicals to reach the target tissue, i.e., the phloem, to effectively manage phytoplasma diseases.
Another phloem-limited disease which gave rise to investigations of trunk injection as a targeted delivery method [150] is huanglongbing (HLB), or citrus greening. HLB is a devastating disease associated with phloem-limited bacteria that has wreaked havoc to citrus industries in the USA and worldwide [151,152,153]. In the USA and most of Asia and South America, the disease is associated with Candidatus Liberibacter asiaticus and is transmitted by the Asian citrus psyllid Diaphorina citri. Studies in the 1970s and 1980s demonstrated the efficacy of using oxytetracycline and other bactericides to alleviate HLB disease symptoms [154,155]. Recent experiments using the bactericides penicillin [156,157,158] and streptomycin [159] have shown mixed results regarding their efficacy and the potential for widescale adoption. The efficacy of oxytetracycline in reducing pathogen levels and improving tree health was demonstrated more recently [136,160,161,162]. However, whether oxytetracycline injections are cost-effective on a commercial scale has not yet been demonstrated. Moreover, antibiotic residue levels in harvested citrus fruits will need to be established before implementation of this methodology to mitigate HLB.
Although injection as a management technique needs evaluation for a large commercial scale, at the experimental level, bactericidal products have been tested for bacterial management in a variety of crop species (Table 1). ‘Tree Tech® OTC’ is an oxytetracycline calcium complex registered for use in crop-bearing peaches and pears for bacterial blast, blossom blight, and fire blight management, and X-disease of peaches; however, there are few other bactericides registered for injection in crop-bearing trees.

Table 1.
Published studies regarding the efficacy of trunk injections for bacterial and phytoplasma disease management in crop species.
Most recently, injection experiments have extended beyond fungicides, insecticides, and bactericides to include other compounds such as plant defense activators [114,161,174], RNA [28,175], plant growth regulators [176], nanoparticles [177,178,179], novel antimicrobial molecules [180], essential oils [181], biofertilizers [182], nutrients [183], nano-carrier treatments [97], and bacteria [157,184,185,186]. However, none are currently used on a commercial basis.
6. Other Considerations
6.1. Location of Injection
There are different recommendations regarding the best placement of injections, which are based on the organism or pathogen being targeted and its location in the tree, the tree species, and the environmental conditions. A finding by Roach (1939) [3] that is still relevant today is that the “uniform permeation of all the branches depends on the proper placing of the hole or holes in relation to the branches”. Early research suggested that injection into the roots allows for a more uniform distribution of injected materials in branches compared to injection into the trunk [187], while others suggested injecting low on trunk ridges (sometimes called flare roots) but not on the roots or depressions between roots [17,188]. The proposed rationale for either of these suggestions is an increased rate of uptake and a quicker wound closure. However, injecting into the root flares may increase the risk for decay caused by soil-borne pathogens [4]. Additional research is necessary in tree crop species to determine where to perform injections to be most effective while preventing injury.
6.2. Xenobiotic Mobility
Xenobiotics, chemical substances found within an organism that are not naturally produced, can be transported through both apoplastic and symplastic routes. Trunk injection relies on the distribution of the chemicals within the vessel elements of the xylem [127,189]; therefore, to be effective, injected compounds will need to be mobile mostly within this tissue. For phloem-limited diseases such as phytoplasmas or Candidatus Liberibacter spp., the mobility of injected compounds in the phloem and between the xylem and phloem also needs to be considered.
Vascular movement and distribution across tissues depend on the chemical properties of the injected chemical [190,191]. Apoplastic transport depends on water solubility, formulation type, and the adsorption coefficient of the compound [36]. Compounds with a high adsorption coefficient are bound to the negative charge of the xylem cell walls and will not move as readily over long distances. Studies by Byrne et al. (2020) [127] suggest that more diluted concentrations of some formulations may improve compound uptake and distribution.
Movement into the phloem is enhanced when compounds have a high dissociation constant (pKa); in contrast, movement within the phloem is enhanced when compounds have a low pKa [192]. This means that compounds are not optimized for both transport into the phloem and movement within the phloem. Compounds that can readily move back and forth across the plasma membrane of sieve tubes will preferentially be transported in the xylem because of the greater flow volume in the xylem compared to the phloem [193].
Understanding the molecular structure and chemical properties of injected materials is critical for predicting the efficacy of injections and developing formulations. In addition to the molecular structure, movement of a xenobiotic depends on various tree physiological characteristics, such as tree height, the radius of the vessel elements, the arrangement of vessels and parenchyma cells, the length of the vessel elements, and the sink strength of roots or other plant organs [194]. Finally, environmental conditions just prior to (e.g., very dry, or high rainfall days) and during (e.g., cloud cover) injection, such as temperature, vapor pressure deficit, and soil moisture, are important factors determining sap flow and therefore efficacy of uptake and distribution of materials within the tree [36]. Understanding these principles is critical for developing effective management methods to protect crops from pests and disease using trunk injection.
Another risk associated with xenobiotics is the potential phytotoxicity of the active ingredient or other components in the injected formulation [7]. The chemicals applied via trunk injection may generate more extensive damage inside the trunk than the damage caused by the physical injury.
6.3. Timing of Injection
The appropriate time (seasonality) for injection has been established for some trees and some products; however, more extensive testing is required for each product and each tree species.
It is expected that the speed of compound uptake and distribution after trunk injection increases in rapidly transpiring trees [36], or rapidly transpiring parts of trees [37,195]. Conditions that lead to increased transpiration are thus recommended for quick uptake and even distribution of materials. For deciduous trees in temperate climates, injections in late spring and summer are most ideal for upward movement, while injections in fall and winter may result in more downward movement [24]. This can be related to the higher rates of transpiration during the active growing seasons. Fully watered trees, warm temperatures, and slightly windy conditions can also enhance transpiration and therefore increase the uptake rate of injected materials. However, timing of injection must also consider the stage of the disease and/or of the pest lifecycle at which management is expected to be most effective [36].
Although seasonality and weather may dictate the rate of uptake and movement of compounds within a tree, regulatory restrictions for compound residues in the fruits may ultimately be the determinant in the timing for injections. Recent studies that have conducted residue analyses have demonstrated variable results depending on the compound and the timing of injections. For example, emamectin benzoate was not detected in the nectar or pollen of apple trees when injected in the fall but was detected in both when injected in the spring [115]. Imidacloprid, on the other hand, was detected in pollen after injection in the fall, but was not detected in either nectar or pollen when injected in the spring [115]. In palms, emamectin benzoate was not detected in fruits 60 days after injection [134], and in avocado, one trial comparing foliar sprays with trunk injection of phosphites found that residues in the fruits after spraying were significantly higher than after injection [100]. Ultimately, compound residues in crops may be the determining factor in the timing and feasibility of injections in commercial orchard systems.
6.4. Environmental and Human Health Concerns
Trunk injection is an effective method for delivering plant protection products into tree trunks, especially compared to foliar sprays or soil drenches. However, despite multiple advantages of this method, such as higher efficacy and less environmental runoff or drift [1], there are some concerns related to its use. For example, when applied via trunk injection, the neonicotinoid insecticide imidacloprid may affect target and non-target species for multiple years due to its persistence in plant tissues [196]. Imidacloprid use has been banned in the European Union since 2018 due to growing evidence of its harmful effects on bees and other beneficial insects.
Another concern is the impact on human health when using antibiotics in plant protection [197,198]. Recently, Taylor and Reeder expressed a growing concern about the use of antibiotics in plant protection due to the potential spread of resistance to medically important bacteria [199]. Moreover, use and regulations differ considerably between different regions around the world and between countries in the same region.
7. Conclusions and Future Perspectives
Trunk injection offers numerous benefits compared to soil drenches or foliar sprays of plant protection compounds [1,2]. Although historically the use of trunk injection has focused on application in large or high-value forest and ornamental species, the technique could be optimized to reduce environmental and human health threats in commercial crop systems. Currently, the cost of manual injection at the orchard scale is not economically feasible for most growers and is a major factor for the limited adoption of the practice in agriculture. Development of techniques and equipment which would allow for more efficient injection are necessary for widespread use.
In addition to the equipment and methods required for injection at the commercial scale, research on best practices for injection in specific species is necessary to develop recommendations. The long-term effects on tree physiology and wounding also need to be examined for each crop system to determine the economic feasibility of trunk injections. Nevertheless, this delivery method has the potential for managing diseases that cannot be managed with traditional methods, and more research is warranted to establish the strengths and limitations for the crop system of interest.
Author Contributions
Conceptualization, U.A. and L.A.; resources, U.A.; writing—original draft preparation, L.A. and U.A.; writing—review and editing, J.H.C.; visualization, L.A. and U.A.; supervision, U.A.; project administration, U.A.; funding acquisition, U.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by USDA-NIFA-SCRI/CDRE, project 2019-70016-29096.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sánchez Zamora, M.A.; Fernández Escobar, R. Injector-Size and the Time of Application Affects Uptake of Tree Trunk-Injected Solutions. Sci. Hortic. 2000, 84, 163–177. [Google Scholar] [CrossRef]
- Wise, J.C.; VanWoerkom, A.H.; Acimovic, S.G.; Sundin, G.W.; Cregg, B.M.; Vandervoort, C.V. Trunk Injection: A Discriminating Delivering System for Horticulture Crop IPM. Entomol. Ornithol. Herpetol. Curr. Res. 2014, 3, 1. [Google Scholar] [CrossRef]
- Roach, W.A. Plant Injection as a Physiological Method. Ann. Bot. 1939, 3, 155–277. [Google Scholar] [CrossRef]
- Perry, T.; Santamour, F.; Stipes, R.; Shear, T.; Shigo, A. Exploring Alternatives To Tree Injection. J. Arboric. 1991, 17, 217–226. [Google Scholar] [CrossRef]
- Osterbauer, N.K.; French, D.W. Propiconazole as a Treatment for Oak Wilt in Quercus rubra and Q. ellipsoidalis. J. Agric. Sci. Technol. A 1992, 18, 221–226. [Google Scholar] [CrossRef]
- Darvas, J.M.; Torien, J.C.; Milne, D.L. Injection of Established Avocado Trees for the Effective Control of Phytophthora Root Rot. Calif. Avocado Soc. 1983, 67, 76–77. [Google Scholar]
- Stennes, M.A.; French, D.W. Distribution and Retention of Thiabendazole Hypophosphite and Carbendazim Phosphate Injected into Mature American Elms. Phytopathology 1987, 77, 707–712. [Google Scholar] [CrossRef]
- Haugen, B.L.; Stennes, M. Fungicide Injection to Control Dutch Elm Disease: Understanding the Options. Plant Dis. Q. 1999, 20, 29–38. [Google Scholar]
- Fernandez-Escobar, R.; Gallego, F.J.; Benlloch, M.; Membrillo, J.; Infante, J.; Perez De Algaba, A. Treatment of Oak Decline Using Pressurized Injection Capsules of Antifungal Materials. Eur. J. For. Pathol. 1999, 29, 29–38. [Google Scholar] [CrossRef]
- Filer, T.H. Pressure Apparatus for Injecting Chemicals Into Trees. Plant Dis. Rep. 1973, 57, 338–341. [Google Scholar]
- Holmes, F.W. Distribution of Dye in Elms after Trunk or Root Injection. J. Arboric. 1982, 8, 250–252. [Google Scholar]
- Kozlowski, T.T.; Winget, C.H. Patterns of Water Movement in Forest Trees. Bot. Gaz. 1963, 124, 301–311. [Google Scholar] [CrossRef]
- Sachs, R.M.; Nyland, G.; Hackett, W.P.; Coffelt, J.; Debie, J.; Giannini, G. Pressurized Injection of Aqueous Solutions into Tree Trunks. Sci. Hortic. 1977, 6, 297–310. [Google Scholar] [CrossRef]
- Shigo, A.L.; Marx, H.G. Compartmentalization of Decay in Trees; USDA Forest Service Agency Information Bulletin; USDA: Washington, DC, USA, 1977; pp. 1–73.
- Santamour, F.S. Wound Compartmentalization in Cultivars of Acer, Gleditsia, and Other Genera. J. Environ. Hortic. 1984, 2, 123–125. [Google Scholar] [CrossRef]
- Stack, R.W. Effect of Tree Size, Hole Location and Wetwood Fluxing on Healing of Injection Wounds in American Elms. J. Arboric. 1985, 11, 45–47. [Google Scholar]
- Smith, K.T. Wounding, Compartmentalization, and Treatment Tradeoffs. J. Arboric. 1988, 14, 226–229. [Google Scholar]
- Neely, D. Tree Wound Closure. J. Arboric. 1988, 14, 148–152. [Google Scholar]
- Costinis, A.C. The Wounding Effects of Mauget and Creative Sales Injections. J. Arboric. 1980, 6, 204–208. [Google Scholar]
- Sinclair, W.A.; Larsen, A.O. Wood Characteristics Related to “Injectability” of Trees. J. Arboric. 1980, 7, 6–10. [Google Scholar]
- Chaney, W.R. Anatomy and Physiology Related to Chemical Movement in Trees. J. Arboric. 1985, 12, 85–91. [Google Scholar]
- Doccola, J.J.; Smitley, D.R.; Davis, T.W.; Aiken, J.J.; Wild, P.M. Tree Wound Responses Following Systemic Insecticide Trunk Injection Treatments in Green Ash (Fraxinus pennsylvanica Marsh.) as Determined by Destructive Autopsy. Arboric. Urban For. 2011, 37, 6–12. [Google Scholar] [CrossRef]
- Aćimović, S.G.; Cregg, B.M.; Sundin, G.W.; Wise, J.C. Comparison of Drill- and Needle-Based Tree Injection Technologies in Healing of Trunk Injection Ports on Apple Trees. Urban For. Urban Green. 2016, 19, 151–157. [Google Scholar] [CrossRef]
- Tattar, T.; Tattar, S. Evidence for the Downward Movement of Materials Injected into Trees. Arboric. Urban For. 1999, 25, 325–332. [Google Scholar] [CrossRef]
- Ford, C.R.; Vose, J.M.; Daley, M.; Phillips, N. Use of Water by Eastern Hemlock: Implications for Systemic Insecticide Application. Arboric. Urban For. 2007, 33, 421–427. [Google Scholar] [CrossRef]
- Shang, Q.; Liao, K.; Liu, H.; Zhao, B. Study on Structure of Needle Head and Seal Mechanism of Tree Trunk Injection. In Proceedings of the 2011 International Conference on Transportation, Mechanical, and Electrical Engineering, TMEE 2011, Changchun, China, 16–18 December 2011; pp. 813–816. [Google Scholar]
- Montecchio, L. A Venturi Effect Can Help Cure Our Trees. J. Vis. Exp. 2013, 80, e51199. [Google Scholar] [CrossRef]
- Dalakouras, A.; Jarausch, W.; Buchholz, G.; Bassler, A.; Braun, M.; Manthey, T.; Krczal, G.; Wassenegger, M. Delivery of Hairpin Rnas and Small Rnas into Woody and Herbaceous Plants by Trunk Injection and Petiole Absorption. Front. Plant Sci. 2018, 9, 1253. [Google Scholar] [CrossRef]
- Killiny, N.; Gonzalez-Blanco, P.; Santos-Ortega, Y.; Al-Rimawi, F.; Levy, A.; Hijaz, F.; Albrecht, U.; Batuman, O. Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin. Antibiotics 2019, 8, 262. [Google Scholar] [CrossRef]
- Kuroda, K.; Yamane, K.; Itoh, Y. In Planta Analysis of the Radial Movement of Minerals from inside to Outside in the Trunks of Standing Japanese Cedar (Cryptomeria japonica D. Don) Trees at the Cellular Level. Forests 2021, 12, 251. [Google Scholar] [CrossRef]
- Kuroda, K.; Yamane, K.; Itoh, Y. Cellular Level in Planta Analysis of Radial Movement of Artificially Injected Caesium in Cryptomeria japonica Xylem. Trees Struct. Funct. 2018, 32, 1505–1517. [Google Scholar] [CrossRef]
- Berger, C.; Laurent, F. Trunk Injection of Plant Protection Products to Protect Trees from Pests and Diseases. Crop Prot. 2019, 124, 104831. [Google Scholar] [CrossRef]
- Tsen, E.W.J.; Sitzia, T.; Webber, B.L. To Core, or Not to Core: The Impact of Coring on Tree Health and a Best-Practice Framework for Collecting Dendrochronological Information from Living Trees. Biol. Rev. 2016, 91, 899–924. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, P.A.; Ali, S.; Braun, G.; Bevis, E.; Fillmore, S. Reducing Apple Scab and Frogeye or Black Rot Infections with Salicylic Acid or Its Analogue on Field-Established Apple Trees. Can. J. Plant Pathol. 2019, 41, 345–354. [Google Scholar] [CrossRef]
- Pegg, K.G. Trunk Injection Methodology. Australas. Plant Pathol. 1990, 19, 142–143. [Google Scholar] [CrossRef]
- Aćimović, S.G.; Martin, D.K.H.; Turcotte, R.M.; Meredith, C.L.; Munck, I.A. Choosing an Adequate Pesticide Delivery System for Managing Pathogens with Difficult Biologies: Case Studies on Diplodia corticola, Venturia inaequalis and Erwinia amylovora. In Plant Diseases—Current Threats and Management Trends; IntechOpen: London, UK, 2020. [Google Scholar]
- Hunt, P.; Dabek, A.J.; Schuilin, M. Remission of Symptoms Following Tetracycline Treatment of Lethal Yellowing-Infected Coconut Palms. Phytopathology 1974, 64, 307–312. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Mencuccini, M.; Álvarez, X.; Camacho, J.; Loepfe, L.; Piñol, J. Spatial Distribution and Packing of Xylem Conduits. Am. J. Bot. 2012, 99, 1189–1196. [Google Scholar] [CrossRef]
- Cartenì, F.; Deslauriers, A.; Rossi, S.; Morin, H.; de Micco, V.; Mazzoleni, S.; Giannino, F. The Physiological Mechanisms behind the Earlywood-to-Latewood Transition: A Process-Based Modeling Approach. Front. Plant Sci. 2018, 9, 1053. [Google Scholar] [CrossRef]
- Ellmore, G.S.; Zanne, A.E.; Orians, C.M. Comparative Sectoriality in Temperate Hardwoods: Hydraulics and Xylem Anatomy. Bot. J. Linn. Soc. 2006, 150, 61–71. [Google Scholar] [CrossRef]
- Zanne, A.E.; Sweeney, K.; Sharma, M.; Orians, C.M. Patterns and Consequences of Differential Vascular Sectoriality in 18 Temperate Tree and Shrub Species. Funct. Ecol. 2006, 20, 200–206. [Google Scholar] [CrossRef]
- Zanne, A.E.; Westoby, M.; Falster, D.S.; Ackerly, D.D.; Loarie, S.R.; Arnold, S.E.J.; Coomes, D.A. Angiosperm Wood Structure: Global Patterns in Vessel Anatomy and Their Relation to Wood Density and Potential Conductivity. Am. J. Bot. 2010, 97, 207–215. [Google Scholar] [CrossRef]
- Nadezhdina, N. Integration of Water Transport Pathways in a Maple Tree: Responses of Sap Flow to Branch Severing. Ann. For. Sci. 2010, 67, 107. [Google Scholar] [CrossRef]
- Thorn, A.M.; Orians, C.M. Modeling the Influence of Differential Sectoriality on the Photosynthetic Responses of Understory Saplings to Patchy Light and Water Availability. Trees Struct. Funct. 2011, 25, 833. [Google Scholar] [CrossRef]
- Tanis, S.R.; Cregg, B.M.; Mota-Sanchez, D.; McCullough, D.G.; Poland, T.M. Spatial and Temporal Distribution of Trunk-Injected 14C-Imidacloprid in Fraxinus Trees. Pest Manag. Sci. 2012, 68, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Ewers, F.W.; Ewers, J.M.; Jacobsen, A.L.; López-Portillo, J. Vessel Redundancy: Modeling Safety in Numbers. IAWA J. 2007, 28, 373–388. [Google Scholar] [CrossRef]
- Schenk, H.J.; Espino, S.; Goedhart, C.M.; Nordenstahl, M.; Cabrera, H.I.M.; Jones, C.S. Hydraulic Integration and Shrub Growth Form Linked across Continental Aridity Gradients. Proc. Natl. Acad. Sci. USA 2008, 105, 11248–11253. [Google Scholar] [CrossRef]
- Dixon, H.H.; Joly, J. XII. On the Ascent of Sap. Philos. Trans. R. Soc. Lond. 1895, 186, 563–576. [Google Scholar] [CrossRef]
- Pickard, W.F. The Ascent of Sap in Plants. Prog. Biophys. Mol. Biol. 1981, 37, 181–229. [Google Scholar] [CrossRef]
- Kim, H.K.; Park, J.; Hwang, I. Investigating Water Transport through the Xylem Network in Vascular Plants. J. Exp. Bot. 2014, 65, 1895–1904. [Google Scholar] [CrossRef]
- Zimmermann, U.; Schneider, H.; Wegner, L.H.; Haase, A. Water Ascent in Tall Trees: Does Evolution of Land Plants Rely on a Highly Metastable State? New Phytol. 2004, 162, 575–615. [Google Scholar] [CrossRef]
- Pereira, L.; Jansen, S.; Miranda, M.T.; Pacheco, V.S.; Kaack, L.; Pires, G.S.; Guan, X.; Mayer, J.L.S.; Machado, E.C.; Schenk, H.J.; et al. Dynamic Changes in Gas Solubility of Xylem Sap Reiterate the Enigma of Plant Water Transport under Negative Pressure. bioRxiv 2022. [Google Scholar]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant Xylem Hydraulics: What We Understand, Current Research, and Future Challenges. J. Integr. Plant Biol. 2017, 59, 356–389. [Google Scholar] [CrossRef]
- Jacobsen, A.L.; Pratt, B.R.; Tobin, M.F.; Hacke, U.G.; Ewers, F.W. A Global Analysis of Xylem Vessel Length in Woody Plants. Am. J. Bot. 2012, 99, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Stroock, A.D.; Pagay, V.V.; Zwieniecki, M.A.; Holbrook, N.M. The Physicochemical Hydrodynamics of Vascular Plants. Annu. Rev. Fluid Mech. 2014, 46, 615–642. [Google Scholar] [CrossRef]
- Nagy, N.E.; Norli, H.R.; Fongen, M.; Østby, R.B.; Heldal, I.M.; Davik, J.; Hietala, A.M. Patterns and Roles of Lignan and Terpenoid Accumulation in the Reaction Zone Compartmentalizing Pathogen-Infected Heartwood of Norway Spruce. Planta 2022, 255, 63. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, A.; Planas-Marquès, M.; Capellades, M.; Valls, M.; Coll, N.S. Blocking Intruders: Inducible Physico-Chemical Barriers against Plant Vascular Wilt Pathogens. J. Exp. Bot. 2021, 72, 184–198. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Roddy, A.B.; Wason, J.W.; McElrone, A.J. Functional Status of Xylem Through Time. Annu. Rev. Plant Biol. 2019, 70, 407–433. [Google Scholar] [CrossRef]
- Pouzoulet, J.; Rolshausen, P.E.; Charbois, R.; Chen, J.; Guillaumie, S.; Ollat, N.; Gambetta, G.A.; Delmas, C.E.L. Behind the Curtain of the Compartmentalization Process: Exploring How Xylem Vessel Diameter Impacts Vascular Pathogen Resistance. Plant Cell Environ. 2020, 43, 2782–2796. [Google Scholar] [CrossRef]
- Morris, H.; Brodersen, C.; Schwarze, F.W.M.R.; Jansen, S. The Parenchyma of Secondary Xylem and Its Critical Role in Tree Defense against Fungal Decay in Relation to the CODIT Model. Front. Plant Sci. 2016, 7, 1665. [Google Scholar] [CrossRef]
- Morris, H.; Hietala, A.M.; Jansen, S.; Ribera, J.; Rosner, S.; Salmeia, K.A.; Schwarze, F.W.M.R. Using the CODIT Model to Explain Secondary Metabolites of Xylem in Defence Systems of Temperate Trees against Decay Fungi. Ann. Bot. 2020, 125, 701–720. [Google Scholar] [CrossRef]
- Pearce, R.B. Reaction Zone Relics and the Dynamics of Fungal Spread in the Xylem of Woody Angiosperms. Physiol. Mol. Plant Pathol. 1991, 39, 41–55. [Google Scholar] [CrossRef]
- Schmitt, U.; Liese, W. Response of Xylem Parenchyma by Suberization in Some Hardwoods after Mechanical Injury. Trees 1993, 8, 23–30. [Google Scholar] [CrossRef]
- Robbertse, P.J.; Duvenhage, J.A. Reaction of Avocado Wood on Injections into the Trunk. South Afr. Avacado Grow. Assoc. Yearb. 1999, 22, 39–47. [Google Scholar]
- Smith, K.T.; Lewis, P.A. Potential Concerns for Tree Wound Response from Stem Injection. In Proceedings of the 3rd Symposium on Hemlock Wooley Adelgid Conference, Asheville, NC, USA, 1–3 February 2005; pp. 173–178. [Google Scholar]
- Biggs, A.R. Suberized Boundary Zones and the Chronology of Wound Response in Tree Bark. Phytopathology 1985, 75, 1191–1195. [Google Scholar] [CrossRef]
- Eyles, A.; Davies, N.W.; Mohammed, C. Novel Detection of Formylated Phloroglucinol Compounds (FPCs) in the Wound Wood of Eucalyptus Globulus and E. Nitens. J. Chem. Ecol. 2003, 29, 881–898. [Google Scholar] [CrossRef] [PubMed]
- Shigo, A.L.; Money, W.E.; Dodds, D.I. Some Internal Effects of Mauget Tree Injections. J. Arboric. 1977, 3, 213–220. [Google Scholar] [CrossRef]
- Shigo, A.L.; Shortle, W.C. Compartmentalization of Discolored Wood in Heartwood of Red Oak. Phytopathology 1979, 69, 710–711. [Google Scholar] [CrossRef]
- Vasconcellos, L.A.B.C.; Castle, W.S. Trunk Xylem Anatomy of Mature Healthy and Blighted Grapefruit Trees on Several Rootstocks. J. Am. Soc. Hortic. Sci. 1994, 119, 185–194. [Google Scholar] [CrossRef]
- Dujesiefken, D.; Liese, W. The CODIT Principle: Implications for Best Practices; Martin One Source: Champaign, IL, USA, 2015. [Google Scholar]
- James, R.; Tisserat, N.; Todd, T. Prevention of Pine Wilt of Scots Pine (Pinus sylvestris) with Systemic Abamectin Injections. Arboric. Urban For. 2006, 32, 195–201. [Google Scholar] [CrossRef]
- Reil, W. Pressure-Injecting Chemicals in to Trees. Calif. Agric. 1979, 33, 16–19. [Google Scholar]
- Martín, J.A.; Sobrino-Plata, J.; Rodríguez-Calcerrada, J.; Collada, C.; Gil, L. Breeding and Scientific Advances in the Fight against Dutch Elm Disease: Will They Allow the Use of Elms in Forest Restoration? New For. 2019, 50, 183–215. [Google Scholar] [CrossRef]
- D’Arcy, C.J. Dutch elm disease. Am. Phytopathol. Soc. 1981, 245, 56–67. [Google Scholar] [CrossRef]
- Marcotrigiano, M. Elms Revisited. Arboric. Urban For. 2017, 43, 217–241. [Google Scholar] [CrossRef]
- Brazee, N.J.; Marra, R.E. Incidence of Internal Decay in American Elms (Ulmus americana) under Regular Fungicide Injection to Manage Dutch Elm Disease. Arboric. Urban For. 2020, 46, 1–11. [Google Scholar] [CrossRef]
- Postma, J.; Goossen-van de Geijn, H. Twenty-Four Years of Dutch Trig® Application to Control Dutch Elm Disease. BioControl 2016, 61, 305–312. [Google Scholar] [CrossRef]
- Stipes, R.J. The Management of Dutch Elm Disease. In The Elms; Springer: Boston, MA, USA, 2000; pp. 157–172. [Google Scholar]
- Schutte, G.C.; Botha, T.; Bezuidenhout, J.J.; Kotze, J.M. Distribution of Phosphite in Avocado Trees after Trunk Injection with Phosphorous Acid and Its Possible Response to Phytophthora cinnamomi. South Afr. Avocado Grow. Assoc. Yearb. 1988, 12, 32–34. [Google Scholar]
- Ramírez-Gil, J.G.; Castañeda-Sánchez, D.A.; Morales-Osorio, J.G. Production of Avocado Trees Infected with Phytophthora cinnamomi under Different Management Regimes. Plant Pathol. 2017, 66, 623–632. [Google Scholar] [CrossRef]
- Dann, E.K.; Le, D.P. Effects of Silicon Amendment on Soilborne and Fruit Diseases of Avocado. Plants 2017, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.H.; Montas, W.; Evans, E.A.; Olszack, R. How to Make a Simple and Inexpensive Passive and Pressurized Infusion System for Systemically Applied Pest Control Substances to Fruit Trees. Proc. Fla. State Hortic. Soc. 2014, 127, 6–9. [Google Scholar]
- De Beer, Z.W.; Procter, M.; Wingfield, M.J.; Marincowitz, S.; Duong, T.A. Generic Boundaries in the Ophiostomatales Reconsidered and Revised. In Studies in Mycology; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2022; pp. 57–120. [Google Scholar] [CrossRef]
- Fraedrich, S.W.; Harrington, T.C.; Rabaglia, R.J.; Ulyshen, M.D.; Mayfield, A.E.; Hanula, J.L.; Eickwort, J.M.; Miller, D.R. A Fungal Symbiont of the Redbay Ambrosia Beetle Causes a Lethal Wilt in Redbay and Other Lauraceae in the Southeastern United States. Plant Dis. 2008, 92, 215–224. [Google Scholar] [CrossRef]
- Harrington, T.C.; Fraedrich, S.W.; Achayeva, D.N. Raffaelea Lauricola, a New Ambrosia Beetle Symbiont and Pathogen on the Lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Carrillo, D.; Duncan, R.E.; Ploetz, J.N.; Campbell, A.F.; Ploetz, R.C.; Peña, J.E. Lateral Transfer of a Phytopathogenic Symbiont among Native and Exotic Ambrosia Beetles. Plant Pathol. 2014, 63, 54–62. [Google Scholar] [CrossRef]
- Cruz, L.F.; Menocal, O.; Kendra, P.E.; Carrillo, D. Phoretic and Internal Transport of Raffaelea lauricola by Different Species of Ambrosia Beetle Associated with Avocado Trees. Symbiosis 2021, 84, 151–161. [Google Scholar] [CrossRef]
- Dann, E.K.; Ploetz, R.C.; Coates, L.M.; Pegg, K.G. Foliar, Fruit and Soilborne Diseases. In The Avocado: Botany, Production and Uses; CABI: Wallingford, UK, 2013; pp. 380–422. [Google Scholar]
- Crane, J.H.; Wasielweski, J.; Carrillo, D.; Gazis, R.; Schaffer, B.; Ballen, F.; Evans, E. Recommendations for the Detection and Mitigation of Laurel Wilt Disease in Avocado and Related Tree Species in the Home Landscape: HS1358, 2/2020; EDIS, University of Florida: Gainesville, FL, USA, 2020. [Google Scholar] [CrossRef]
- Crane, J.H.; Carrillo, D.; Evans, E.A.; Gazis, R.; Schaffer, B.; Ballen, F.; Wasielweski, J. Current Recommendations for Control of Laurel Wilt and Its Ambrosia Beetle Vectors in Florida’s Commercial Avocado Groves. Proc. Fla. State Hortic. Soc. 2020, 133, 4–6. [Google Scholar]
- Crane, J.H.; Ploetz, R.C.; White, T.; Krogstad, G.C.; Prosser, T.; Konkol, J.; Wideman, R. Efficacy of Three Microinfused Fungicides to Control Laurel Wilt on Avocado in Martin and Brevard Counties. Proc. Fla. State Hortic. Soc. 2015, 128, 58–60. [Google Scholar]
- Dal Maso, E.; Cocking, J.; Montecchio, L. An Enhanced Trunk Injection Formulation of Potassium Phosphite against Chestnut Ink Disease. Arboric. J. 2017, 39, 124–141. [Google Scholar] [CrossRef]
- Dal Maso, E.; Linaldeddu, B.T.; Fanchin, G.; Faccoli, M.; Montecchio, L. The Potential for Pesticide Trunk Injections for Control of Thousand Cankers Disease of Walnut. Phytopathol. Mediterr. 2019, 58, 73–79. [Google Scholar] [CrossRef]
- González, M.; Romero, M.Á.; Serrano, M.S.; Sánchez, M.E. Fosetyl-Aluminium Injection Controls Root Rot Disease Affecting Quercus Suber in Southern Spain. Eur. J. Plant Pathol. 2020, 156, 101–109. [Google Scholar] [CrossRef]
- Romero, M.A.; González, M.; Serrano, M.S.; Sánchez, M.E. Trunk Injection of Fosetyl-Aluminium Controls the Root Disease Caused by Phytophthora cinnamomi on Quercus ilex Woodlands. Ann. Appl. Biol. 2019, 174, 313–318. [Google Scholar] [CrossRef]
- Fischer, J.; Beckers, S.J.; Yiamsawas, D.; Thines, E.; Landfester, K.; Wurm, F.R. Targeted Drug Delivery in Plants: Enzyme-Responsive Lignin Nanocarriers for the Curative Treatment of the Worldwide Grapevine Trunk Disease Esca. Adv. Sci. 2019, 6, 1802315. [Google Scholar] [CrossRef]
- Düker, A.; Kubiak, R. Stem Injection of Triazoles for the Protection of Vitis vinifera L. (‘Riesling’) against Powdery Mildew (Uncinula necator). Vitis J. Grapevine Res. 2011, 50, 73–79. [Google Scholar]
- Masikane, S.L.; Novela, P.; Mohale, P.; McLeod, A. Effect of Phosphonate Application Timing and -Strategy on Phosphite Fruit and Root Residues of Avocado. Crop Prot. 2020, 128, 105008. [Google Scholar] [CrossRef]
- McLeod, A.; Masikane, S.L.; Novela, P.; Ma, J.; Mohale, P.; Nyoni, M.; Stander, M.; Wessels, J.P.B.; Pieterse, P. Quantification of Root Phosphite Concentrations for Evaluating the Potential of Foliar Phosphonate Sprays for the Management of Avocado Root Rot. Crop Prot. 2018, 103, 87–97. [Google Scholar] [CrossRef]
- Mule, R.; Fodale, A.S.; Tucci, A. Control of Olive Verticillium Wilt by Trunk Injection with Different Doses of Fosetyl Al and Benomyl. Acta Hortic. 2002, 586, 761–764. [Google Scholar] [CrossRef]
- Tsror, L.; Bar-On, N.; Tugendhaft, Y.; Lavee, S. Attempts to Control Verticillium Wilt on Olives in Israel. In Proceedings of the Olivebioteq, Marsala, Italy, 5–10 November 2006; pp. 235–238. [Google Scholar]
- Tattar, T.A.; Dotson, J.A.; Ruizzo, M.S.; Steward, V.B. Translocation of Imidacloprid in Three Tree Species When Trunk- and Soil-Injected. J. Arboric. 1998, 24, 54–56. [Google Scholar] [CrossRef]
- Cowles, R.S.; Montgomery, M.E.; Cheah, C.A.S.-J. Activity and Residues of Imidacloprid Applied to Soil and Tree Trunks to Control Hemlock Woolly Adelgid (Hemiptera: Adelgidae) in Forests. J. Econ. Entomol. 2006, 99, 1258–1267. [Google Scholar] [CrossRef]
- Mota-Sanchez, D.; Cregg, B.M.; McCullough, D.G.; Poland, T.M.; Hollingworth, R.M. Distribution of Trunk-Injected 14C-Imidacloprid in Ash Trees and Effects on Emerald Ash Borer (Coleoptera: Buprestidae) Adults. Crop Prot. 2009, 28, 655–661. [Google Scholar] [CrossRef]
- Doccola, J.J.; Wild, P.M. Tree Injection as an Alternative Method of Insecticide Application. In Insecticides—Basic and Other Applications; Soloneski, S., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Smitley, D.R.; Doccola, J.J.; Cox, D.L. Multiple-Year Protection of Ash Trees from Emerald Ash Borer with a Single Trunk Injection of Emamectin Benzoate, and Single-Year Protection with an Imidacloprid Basal Drench. Arboric. Urban For. 2010, 36, 206–211. [Google Scholar] [CrossRef]
- McCullough, D.G.; Poland, T.M.; Tluczek, A.R.; Anulewicz, A.; Wieferich, J.; Siegert, N.W. Emerald Ash Borer (Coleoptera: Buprestidae) Densities over a 6-Yr Period on Untreated Trees and Trees Treated with Systemic Insecticides at 1-, 2-, and 3-Yr Intervals in a Central Michigan Forest. J. Econ. Entomol. 2019, 112, 201–212. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer Invasion of North America: History, Biology, Ecology, Impacts, and Management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef]
- Lee, S.C.; Lee, H.R.; Kim, D.S.; Kwon, J.H.; Huh, M.J.; Park, I.K. Emamectin Benzoate 9.7% SL as a New Formulation for a Trunk-Injections against Pine Wood Nematode, Bursaphelenchus xylophilus. J. For. Res. 2020, 31, 1399–1403. [Google Scholar] [CrossRef]
- Sousa, E.; Naves, P.; Vieira, M. Prevention of Pine Wilt Disease Induced by Bursaphelenchus xylophilus and Monochamus galloprovincialis by Trunk Injection of Emamectin Benzoate. Phytoparasitica 2012, 41, 143–148. [Google Scholar] [CrossRef]
- Yang, Z.; ZhengYun, L.; DingChao, Y.; FeiFei, H.; LinPing, Z.; Dong, L. Screening and Prevention Effect of New and Effective Pesticides against Pine Wilt Nematode Bursaphelenchus xylophilus. J. Plant Prot. 2017, 44, 856–862. [Google Scholar]
- Cha, D.J.; Kim, J.; Kim, D.S. Nematicidal Activities of Three Naphthoquinones against the Pine Wood Nematode, Bursaphelenchus xylophilus. Molecules 2019, 24, 3634. [Google Scholar] [CrossRef] [PubMed]
- Coslor, C.C.; Sundin, G.W.; Wise, J.C. The Efficacy of Trunk Injections of Emamectin Benzoate and Phosphorous Acid for Control of Obliquebanded Leafroller and Apple Scab on Semi-Dwarf Apple. Crop Prot. 2019, 118, 44–49. [Google Scholar] [CrossRef]
- Coslor, C.C.; Vandervoort, C.; Wise, J.C. Insecticide Dose and Seasonal Timing of Trunk Injection in Apples Influence Efficacy and Residues in Nectar and Plant Parts. Pest Manag. Sci. 2019, 75, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, R.M.; Lagalante, A.; Jones, J.; Cook, F.; Elliott, T.; Billings, A.A.; Park, Y.L. Spatial and Temporal Distribution of Imidacloprid within the Crown of Eastern Hemlock. J. Insect Sci. 2017, 17, 1–7. [Google Scholar] [CrossRef]
- Coleman, T.W.; Smith, S.L.; Jones, M.I.; Graves, A.D.; Strom, B.L. Efficacy of Systemic Insecticides for Control of the Invasive Goldspotted Oak Borer (Coleoptera: Buprestidae) in California. J. Econ. Entomol. 2017, 110, 2129–2139. [Google Scholar] [CrossRef]
- Bhandari, B.P.; Cheng, Z. Trunk Injection of Systemic Insecticides to Control Stem and Leaf Gall Wasps, Josephiella Species (Hymenoptera: Agaonidae), on Chinese Banyan (Rosales: Moraceae) in Hawaii. Fla. Entomol. 2016, 99, 172–177. [Google Scholar] [CrossRef]
- Wang, J.H.; Che, S.C.; Qiu, L.F.; Li, G.; Shao, J.L.; Zhong, L.; Zhang, G.F.; Xu, H. Efficacy of Emamectin Benzoate Trunk Injection against the Asian Long-Horned Beetle [Anoplophora glabripennis (Coleoptera: Cerambycidae)]. J. Econ. Entomol. 2020, 113, 340–347. [Google Scholar] [CrossRef]
- Bhandari, B.P.; Cheng, Z. Lobate Lac Scale, Paratachardina Pseudolobata (Hemiptera: Keriidae), in Hawaii’s Urban Landscape: Hosts and Management. Int. J. Trop. Insect Sci. 2018, 38, 71–76. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, J.; Li, Y.; Li, J.; Shi, X.H. Evaluation of the Effectiveness of Insecticide Trunk Injections for Control of Latoia Lepida (Cramer) in the Sweet Olive Tree Osmanthus fragrans. PeerJ 2016, 4, e2480. [Google Scholar] [CrossRef]
- Ali, A.D.; Caldwell, D.L. Management of Staining and Galling Associated with Oxhorn Bucida Trees in Florida. Fla. Entomol. 2017, 100, 602–606. [Google Scholar] [CrossRef]
- Jagiełło, R.; Walczak, U.; Iszkuło, G.; Karolewski, P.; Baraniak, E.; Giertych, M.J. Impact of Cameraria Ohridella on Aesculus hippocastanum Growth and Long-Term Effects of Trunk Injection with Pesticides. Int. J. Pest Manag. 2019, 65, 33–43. [Google Scholar] [CrossRef]
- Gubka, A.; Zubrik, M.; Rell, S.; Gareau, N.; Goble, T.; Nikolov, C.; Galko, J.; Vakula, J.; Kunca, A.; Dejonge, R. The Effectiveness of the Neem Product TreeAzin® in Controlling Cameraria ohridella (Lepidoptera: Gracillariidae: Lithocolletinae). Eur. J. Entomol. 2020, 117, 463–473. [Google Scholar] [CrossRef]
- Grosman, D.M.; Eskalen, A.; Brownie, C. Evaluation of Emamectin Benzoate and Propiconazole for Management of a New Invasive Shot Hole Borer (Euwallacea Nr. fornicatus, Coleoptera: Curculionidae) and Symbiotic Fungi in California Sycamores. J. Econ. Entomol. 2019, 112, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E.; Kabashima, J.; Eskalen, A.; Dimson, M.; Mayorquin, J.S.; Carrillo, J.D.; Hanlon, C.C.; Paine, T.D. Evaluations of Insecticides and Fungicides for Reducing Attack Rates of a New Invasive Ambrosia Beetle (Euwallacea Sp., Coleoptera: Curculionidae: Scolytinae) in Infested Landscape Trees in California. J. Econ. Entomol. 2017, 110, 1611–1618. [Google Scholar] [CrossRef]
- Byrne, F.J.; Almanzor, J.; Tellez, I.; Eskalen, A.; Grosman, D.M.; Morse, J.G. Evaluation of Trunk-Injected Emamectin Benzoate as a Potential Management Strategy for Kuroshio Shot Hole Borer in Avocado Trees. Crop Prot. 2020, 132, 105136. [Google Scholar] [CrossRef]
- Wheeler, C.E.; Vandervoort, C.; Wise, J.C. Organic Control of Pear Psylla in Pear with Trunk Injection. Insects 2020, 11, 650. [Google Scholar] [CrossRef]
- Monteys, V.I.S.; Ribes, A.C.; Savin, I. The Invasive Longhorn Beetle Xylotrechus chinensis, Pest of Mulberries, in Europe: Study on Its Local Spread and Efficacy of Abamectin Control. PLoS ONE 2021, 16, e0245527. [Google Scholar] [CrossRef]
- Mokhtaryan, A.; Sheikhigarjan, A.; Arbab, A.; Mohammadipour, A.; Ardestanirostami, H. The Efficiency of Systemic Insecticides and Complete Fertilizer by Trunk Injection Method against Leopard Moth in Infested Walnut Trees. J. Basic Appl. Zool. 2021, 82, 55. [Google Scholar] [CrossRef]
- Kiss, M.; Hachoumi, I.; Nagy, V.; Ladányi, M.; Gutermuth, Á.; Szabó, Á.; Sörös, C. Preliminary Results about the Efficacy of Abamectin Trunk Injection against the Walnut Husk Fly (Rhagoletis completa). J. Plant Dis. Prot. 2021, 128, 333–338. [Google Scholar] [CrossRef]
- Fu, B.; Qiu, H.; Li, Q.; Tang, L.; Zeng, D.; Liu, K.; Gao, Y. Flower Injection of Imidacloprid and Spirotetramat: A Novel Tool for the Management of Banana Thrips Thrips hawaiiensis. J. Pest Sci. 2020, 93, 1073–1084. [Google Scholar] [CrossRef]
- Chihaoui-Meridja, S.; Harbi, A.; Abbes, K.; Chaabane, H.; la Pergola, A.; Chermiti, B.; Suma, P. Systematicity, Persistence and Efficacy of Selected Insecticides Used in Endotherapy to Control the Red Palm Weevil Rhynchophorus ferrugineus (Olivier, 1790) on Phoenix canariensis. Phytoparasitica 2020, 48, 75–85. [Google Scholar] [CrossRef]
- Mashal, M.M.; Obeidat, B.F. The Efficacy Assessment of Emamectin Benzoate Using Micro Injection System to Control Red Palm Weevil. Heliyon 2019, 5, e01833. [Google Scholar] [CrossRef] [PubMed]
- Kou, H.; Sun, Y.; Dong, Z.; Zhang, Z. Comparison between Sustained Effects of Spray and Injection Thiamethoxam on Apple Aphids and Non-Target Insects in Apple Orchard. Ecotoxicol. Environ. Saf. 2021, 207, 111307. [Google Scholar] [CrossRef]
- Hu, J.; Wang, N. Evaluation of the Spatiotemporal Dynamics of Oxytetracycline and Its Control Effect against Citrus Huanglongbing via Trunk Injection. Phytopathology 2016, 106, 1495–1503. [Google Scholar] [CrossRef]
- Amanifar, N.; Taghavi, M.; Salehi, M. Xylella Fastidiosa from Almond in Iran: Overwinter Recovery and Effects of Antibiotics. Phytopathol. Mediterr. 2016, 55, 337–345. [Google Scholar] [CrossRef]
- Shin, K.; Ascunce, M.S.; Narouei-Khandan, H.A.; Sun, X.; Jones, D.; Kolawole, O.O.; Goss, E.M.; van Bruggen, A.H.C. Effects and Side Effects of Penicillin Injection in Huanglongbing Affected Grapefruit Trees. Crop Prot. 2016, 90, 106–116. [Google Scholar] [CrossRef]
- Norelli, J.L.; Jones, A.L.; Aldwinckle, H.S. Fire Blight Management in the Twenty-First Century: Using New Technologies That Enhance Host Resistance in Apple. Plant Dis. 2003, 87, 756–765. [Google Scholar] [CrossRef]
- Sundin, G.W.; Werner, N.A.; Yoder, K.S.; Aldwinckle, H.S. Field Evaluation of Biological Control of Fire Blight in the Eastern United States. Plant Dis. 2009, 93, 386–394. [Google Scholar] [CrossRef]
- McManus, P.S.; Jones, A.L. Epidemiology and Genetic Analysis of Streptomycin-Resistant Erwinia amylovora from Michigan and Evaluation of Oxytetracycline for Control. Phytopathology 1994, 84, 627–633. [Google Scholar] [CrossRef]
- Loper, J.E.; Henkels, M.D. Evaluation of Streptomycin, Oxytetracycline, and Copper Resistance of Erwinia amylovora Isolated from Pear Orchards in Washington State. Plant Dis. 1991, 75, 287–290. [Google Scholar] [CrossRef]
- Christiano, R.S.C.; Reilly, C.C.; Miller, W.P.; Scherm, H. Oxytetracycline Dynamics on Peach Leaves in Relation to Temperature, Sunlight, and Simulated Rain. Plant Dis. 2010, 94, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Aćimović, S.G.; Mcghee, G.C.; Sundin, G.W.; Wise, J.C. Evaluation of Trunk-Injected Bactericides and Prohexadione-Calcium for Environmentally Friendly Control of Fire Blight (Erwinia amylovora) in Apples. In Proceedings of the 7th Congress on Plant Protection, Zlatibor, Serbia, 24–28 November 2014; Marčić, D.M., Glavendekić, P.N., Eds.; Plant Protection Society of Serbia: Serbia, Belgrade, 2015; pp. 129–134. [Google Scholar]
- Aćimović, S.G.; Zeng, Q.; McGhee, G.C.; Sundin, G.W.; Wise, J.C. Control of Fire Blight (Erwinia amylovora) on Apple Trees with Trunk-Injected Plant Resistance Inducers and Antibiotics and Assessment of Induction of Pathogenesis-Related Protein Genes. Front. Plant Sci. 2015, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Bahder, B.W.; Helmick, E.E.; Chakrabarti, S.; Osorio, S.; Soto, N.; Chouvenc, T.; Harrison, N.A. Disease Progression of a Lethal Decline Caused by the 16SrIV-D Phytoplasma in Florida Palms. Plant Pathol. 2018, 67, 1821–1828. [Google Scholar] [CrossRef]
- Gurr, G.M.; Johnson, A.C.; Ash, G.J.; Wilson, B.A.L.; Ero, M.M.; Pilotti, C.A.; Dewhurst, C.F.; You, M.S. Coconut Lethal Yellowing Diseases: A Phytoplasma Threat to Palms of Global Economic and Social Significance. Front. Plant Sci. 2016, 7, 1521. [Google Scholar] [CrossRef]
- Elliott, M.L. Emerging Palm Diseases in Florida. Horttechnology 2009, 19, 717–718. [Google Scholar] [CrossRef]
- McCoy, R.E.; Norris, R.C.; Vieyra, G.; Delgado, S. Lethal Yellowing Disease of Coconut Palms. FAO Plant Prot. Bull. 1982, 30, 79–80. [Google Scholar]
- Li, M.; Nangong, Z. Precision Trunk Injection Technology for Treatment of Huanglongbing (HLB)-Affected Citrus Trees—A Review. J. Plant Dis. Prot. 2022, 129, 15–34. [Google Scholar] [CrossRef]
- Gottwald, T.R.; da Graça, J.V.; Bassanezi, R.B. Citrus Huanglongbing: The Pathogen and Its Impact. Plant Health Prog. 2007, 8, 31. [Google Scholar] [CrossRef]
- Da Graça, J.V.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G.; Zhao, H. Huanglongbing: An Overview of a Complex Pathosystem Ravaging the World’s Citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar] [CrossRef]
- Bassanezi, R.B.; Primiano, I.V.; Vescove, H.V. Effect of Enhanced Nutritional Programs and Exogenous Auxin Spraying on Huanglongbing Severity, Fruit Drop, Yield and Economic Profitability of Orange Orchards. Crop Prot. 2021, 145, 105609. [Google Scholar] [CrossRef]
- Graca, J.V. Citrus Greening Disease. Annu. Rev. Phytopathol. 1991, 29, 109–136. [Google Scholar] [CrossRef]
- Aubert, B.; Bové, J.M. Effect of Penicillin or Tetracycline Injections of Citrus Trees Affected by Greening Disease Under Field Conditions in Reunion Island. Int. Organ. Citrus Virol. Conf. Proc. 1980, 8, 103–108. [Google Scholar] [CrossRef]
- McVay, J.; Sun, X.; Jones, D.; Urbina, H.; Aldeek, F.; Cook, J.M.; Jeyaprakash, A.; Hodges, G.; Smith, T. Limited Persistence of Residues and Metabolites in Fruit and Juice Following Penicillin Trunk Infusion in Citrus Affected by Huanglongbing. Crop Prot. 2019, 125, 104753. [Google Scholar] [CrossRef]
- Ascunce, M.S.; Shin, K.; Huguet-Tapia, J.C.; Poudel, R.; Garrett, K.A.; van Bruggen, A.H.C.; Goss, E.M. Penicillin Trunk Injection Affects Bacterial Community Structure in Citrus Trees. Microb. Ecol. 2019, 78, 457–469. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, C.; Powell, C.A.; Avery, P.B.; Wang, J.; Huang, Y.; Duan, Y. Field Evaluation of Integrated Management for Mitigating Citrus Huanglongbing in Florida. Front. Plant Sci. 2019, 9, 1890. [Google Scholar] [CrossRef]
- Li, J.; Kolbasov, V.G.; Lee, D.; Pang, Z.; Huang, Y.; Collins, N.; Wang, N. Residue Dynamics of Streptomycin in Citrus Delivered by Foliar Spray and Trunk Injection and Effect on “Candidatus Liberibacter asiaticus” Titer. Phytopathology 2021, 111, 1095–1103. [Google Scholar] [CrossRef]
- Li, J.; Pang, Z.; Duan, S.; Lee, D.; Kolbasov, V.G.; Wang, N. The in Planta Effective Concentration of Oxytetracycline against ‘Candidatus Liberibacter asiaticus’ for Suppression of Citrus Huanglongbing. Phytopathology 2019, 109, 2046–2054. [Google Scholar] [CrossRef]
- Hu, J.; Jiang, J.; Wang, N. Control of Citrus Huanglongbing via Trunk Injection of Plant Defense Activators and Antibiotics. Phytopathology 2018, 108, 186–195. [Google Scholar] [CrossRef]
- Albrecht, U.; Archer, L. Principles and risks of trunk injection for delivery of crop protection materials. Citrus Ind. Mag. 2021, 102, 14–17. [Google Scholar]
- Rumbos, I.C. Control of Plum Chlorotic Leaf Roll Disease by Trunk Pressure Injection of Oxytetracycline-HCI. J. Plant Dis. Prot. 1985, 92, 581–587. [Google Scholar]
- Keil, H.L. Apricot Trunk Injection with Oxytetracycline to Control Bacterial Spot. Proc. Am. Phytopathol. Soc. 1977, 4, 216. [Google Scholar]
- Yang, C.; Powell, C.A.; Duan, Y.; Shatters, R.G.; Lin, Y.; Zhang, M. Mitigating Citrus Huanglongbing via Effective Application of Antimicrobial Compounds and Thermotherapy. Crop Prot. 2016, 84, 150–158. [Google Scholar] [CrossRef]
- Harrison, N.A.; Elliott, M.L. Lethal Yellowing (LY) of Palm. PP-222/PP146 01/2006; EDIS, University of Florida: Gainesville, FL, USA, 2019. [Google Scholar] [CrossRef]
- Cooley, D.R.; Tattar, T.A.; Schieffer, J.T. Treatment of X-Disease of Peaches Using Oxytetracycline Microinjection Capsules. HortScience 1992, 27, 235–237. [Google Scholar] [CrossRef]
- Rosenberger, D.A.; Jones, A.L. Symptom Remission in X-Diseased Peach Trees as Affected by Date, Method, and Rate of Application of Oxytetracycline-HCl. Phytopathology 1977, 77, 277–282. [Google Scholar] [CrossRef]
- Reil, W.O.; Beutel, J.A.; Hemstreet, C.L.; Seyman, W.S. Trunk Injection Corrects Iron and Zinc Deficiency in Pear Trees. Calif. Agric. 1978, 32, 22–23. [Google Scholar]
- McIntyre, J.L.; Schneider, H.; Lacy, G.H.; Dodds, J.A. Pear Decline in Connecticut and Response of Diseased Trees to Oxytetracycline Infusion. Phytopathology 1979, 69, 955–958. [Google Scholar] [CrossRef]
- Lacy, G.H.; McLntyre, J.L.; Walton, G.S.; Dodds, J.A. Rapid Method for and Effects of Infusing Trees with Concentrated Oxytetracycline-HCL Solutions for Pear Decline Control. Can. J. Plant Pathol. 1980, 2, 96–101. [Google Scholar] [CrossRef]
- Du Plessis, H.J. Control of Bacterial Spot on Plums by Sprays of Calcium Oxytetracycline and Copper and Infusion with N-Pyrrolidinomethyl Tetracycline. Phytophylactica 1987, 19, 227–229. [Google Scholar] [CrossRef]
- Puttamuk, T.; Zhang, S.; Duan, Y.; Jantasorn, A.; Thaveechai, N. Effect of Chemical Treatments on “Candidatus Liberibacter asiaticus” Infected Pomelo (Citrus maxima). Crop Prot. 2014, 65, 114–121. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Konkol, J.L.; Pérez-Martínez, J.M.; Fernandez, R. Management of Laurel Wilt of Avocado, Caused by Raffaelea lauricola. Eur. J. Plant Pathol. 2017, 149, 133–143. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi Efficiency, Systemic Properties, and Novel Delivery Methods for Pest Insect Control: What We Know so Far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; von Aderkas, P.; Zaharia, L.I. Effects of Stem-Injected Gibberellins and 6-Benzylaminopurine on Phytohormone Profiles and Cone Yield in Two Lodgepole Pine Genotypes. Trees Struct. Funct. 2018, 32, 765–775. [Google Scholar] [CrossRef]
- Stephano-Hornedo, J.L.; Torres-Gutiérrez, O.; Toledano-Magaña, Y.; Gradilla-Martínez, I.; Pestryakov, A.; Sánchez-González, A.; García-Ramos, J.C.; Bogdanchikova, N. ArgovitTM Silver Nanoparticles to Fight Huanglongbing Disease in Mexican Limes (Citrus aurantifolia Swingle). RSC Adv. 2020, 10, 6146–6155. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Ashworth, V.E.T.M.; Geitner, N.K.; Wiesner, M.R.; Ginnan, N.; Rolshausen, P.; Roper, C.; Jassby, D. Delivery, Fate, and Mobility of Silver Nanoparticles in Citrus Trees. ACS Nano 2020, 14, 2966–2981. [Google Scholar] [CrossRef]
- Beckers, S.J.; Staal, A.H.J.; Rosenauer, C.; Srinivas, M.; Landfester, K.; Wurm, F.R. Targeted Drug Delivery for Sustainable Crop Protection: Transport and Stability of Polymeric Nanocarriers in Plants. Adv. Sci. 2021, 8, 2100067. [Google Scholar] [CrossRef]
- Gardner, C.L.; da Silva, D.R.; Pagliai, F.A.; Pan, L.; Padgett-Pagliai, K.A.; Blaustein, R.A.; Merli, M.L.; Zhang, D.; Pereira, C.; Teplitski, M.; et al. Assessment of Unconventional Antimicrobial Compounds for the Control of ‘Candidatus Liberibacter asiaticus’, the Causative Agent of Citrus Greening Disease. Sci. Rep. 2020, 10, 5395. [Google Scholar] [CrossRef]
- Werrie, P.Y.; Burgeon, C.; le Goff, G.J.; Hance, T.; Fauconnier, M.L. Biopesticide Trunk Injection into Apple Trees: A Proof of Concept for the Systemic Movement of Mint and Cinnamon Essential Oils. Front. Plant Sci. 2021, 12, 650132. [Google Scholar] [CrossRef]
- Scortichini, M.; Loreti, S.; Pucci, N.; Scala, V.; Tatulli, G.; Verweire, D.; Oehl, M.; Widmer, U.; Codina, J.M.; Hertl, P.; et al. Progress towards Sustainable Control of Xylella fastidiosa subsp. pauca in Olive Groves of Salento (Apulia, Italy). Pathogens 2021, 10, 668. [Google Scholar] [CrossRef]
- Hesami, A.; Jafari, N.; Shahriari, M.H.; Zolfi, M. Yield and Physico-Chemical Composition of Date-Palm (Phoenix dactylifera) as Affected by Nitrogen and Zinc Application. Commun. Soil Sci. Plant Anal. 2017, 48, 1943–1954. [Google Scholar] [CrossRef]
- Rosmana, A.; Taufik, M.; Asman, A.; Jayanti, N.J.; Hakkar, A.A. Dynamic of Vascular Streak Dieback Disease Incidence on Susceptible Cacao Treated with Composted Plant Residues and Trichoderma asperellum in Field. Agronomy 2019, 9, 650. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Ager, K.L. Biological Control of Citrus Huanglongbing with Eb92-1, a Benign Strain of Xylella fastidiosa. Plant Dis. 2021, 105, 2914–2918. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.; Czarnocka, K.; Cochard, B.; Oszako, T.; Lefort, F. Biocontrol Endotherapy with Trichoderma spp. and Bacillus amyloliquefaciens against Phytophthora spp.: A Comparative Study with Phosphite Treatment on Quercus robur and Fagus sylvatica. J. Agric. Sci. Technol. A 2015, 5, 428–439. [Google Scholar] [CrossRef]
- Kondo, E.S. A Method for Introducing Water—Soluble Chemicals into Mature Elms; Information Report O-X-171; Canadian Forest Services Publications: Sault Ste. Marie, ON, Canada, 1972. [Google Scholar]
- Stipes, R.J. Glitches and Gaps in the Science and Technology of Tree Injection. J. Arboric. 1988, 14, 165–172. [Google Scholar]
- Aćimović, S.G.; Vanwoerkom, A.H.; Reeb, P.D.; Vandervoort, C.; Garavaglia, T.; Cregg, B.M.; Wise, J.C. Spatial and Temporal Distribution of Trunk-Injected Imidacloprid in Apple Tree Canopies. Pest Manag. Sci. 2014, 70, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.A. Physicochemical Factors Governing the Transport of Xenobiotic Chemicals in Plants: Movement into Roots and Partitioning Between Xylem and Phloem. Acta Hortic. 1989, 239, 43–54. [Google Scholar] [CrossRef]
- Al-Rimawi, F.; Hijaz, F.; Nehela, Y.; Batuman, O.; Killiny, N. Uptake, Translocation, and Stability of Oxytetracycline and Streptomycin in Citrus Plants. Antibiotics 2019, 8, 196. [Google Scholar] [CrossRef] [PubMed]
- Kleier, D.A. Phloem Mobility of Xenobiotics I. Mathematical Model Unifying the Weak Acid and Intermediate Permeability Theories. Plant Physiol. 1988, 86, 803–810. [Google Scholar] [CrossRef]
- Riederer, M. Uptake and Transport of Xenobiotics. In Plant Toxicology; Hock, B., Elstner, E.F., Eds.; Marcel Dekker: New York, NY, USA, 2004; pp. 131–150. [Google Scholar]
- Tyree, M.T.; Peterson, C.A.; Edgington, L.V. A Simple Theory Regarding Ambimobility of Xenobiotics with Special Reference to the Nematicide, Oxamyl. Plant Physiol. 1979, 63, 367–374. [Google Scholar] [CrossRef]
- McCoy, R.E. Comparative Epidemiology of the Lethal Yellowing, Kaincope, and Cadang-Cadang Diseases of Coconut Palm. Plant Dis. Report. 1976, 60, 498–502. [Google Scholar]
- VanWoerkom, A.H.; Aćimović, S.G.; Sundin, G.W.; Cregg, B.M.; Mota-Sanchez, D.; Vandervoort, C.; Wise, J.C. Trunk Injection: An Alternative Technique for Pesticide Delivery in Apples. Crop Prot. 2014, 65, 173–185. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Duffy, B. Use of Antibiotics in Plant Agriculture. OIE Rev. Sci. Tech. 2012, 31, 199–210. [Google Scholar] [CrossRef] [PubMed]
- McManus, P.S.; Stockwell, V.O.; Sundin, G.W.; Jones, A.L. Antibiotic Use in Plant Agriculture. Annu. Rev. Phytopathol. 2002, 40, 443–465. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Reeder, R. Antibiotic Use on Crops in Low and Middle-Income Countries Based on Recommendations Made by Agricultural Advisors. CABI Agric. Biosci. 2020, 1, 1. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).