
Functional Analysis of Two Terpene Synthase Genes Isolated from the Flowers of Hosta ‘So Sweet’

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. RNA Extraction and cDNA Synthesis
2.3. Cloning of HsTPS1 and HsTPS2 Genes
2.4. Sequence Analysis
2.5. Quantitative Real-Time PCR Analysis
2.6. Expression of Recombinant HsTPS1, HsTPS2 in E. coli and In Vitro Enzyme Assay
2.7. GC–MS Analysis
2.8. Subcellular Localization of HsTPS Proteins
3. Results
3.1. Phylogenetic Analysis of TPS Genes from H. ‘So Sweet’
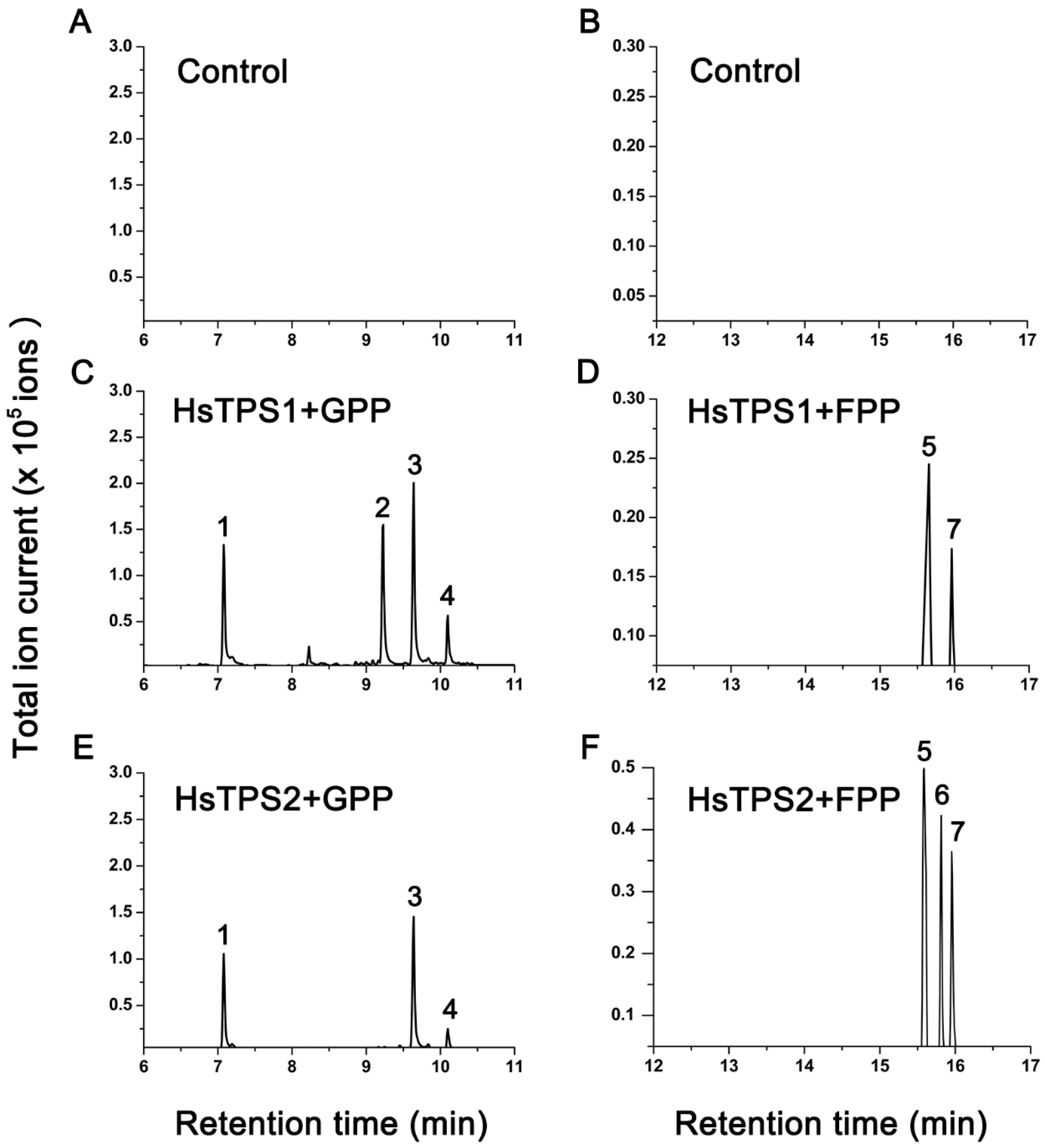
3.2. Functional Characterization of Recombinant HsTPS1 and HsTPS2
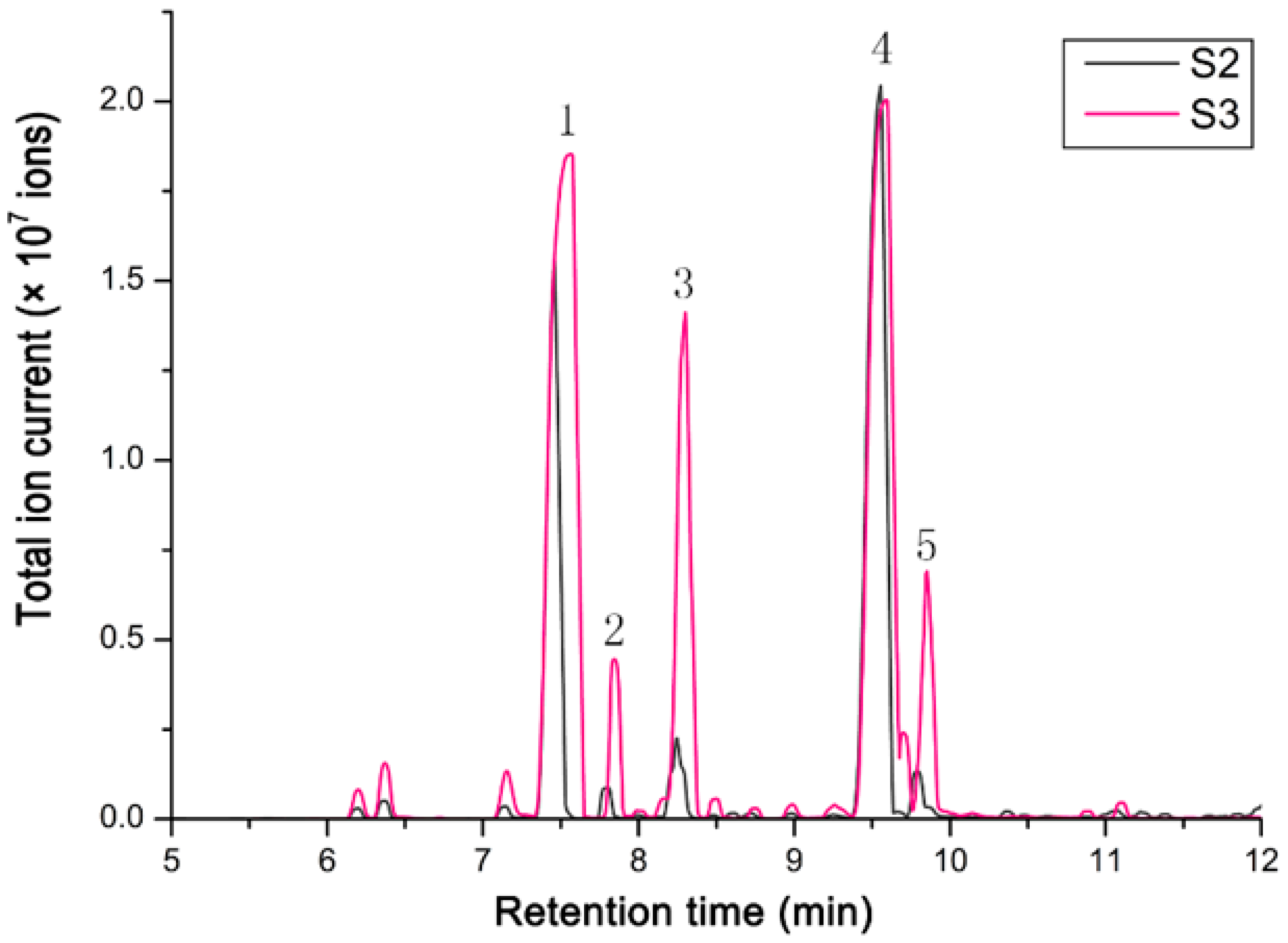
3.3. Terpenoids Production in Hosta and Expressional Analysis of HsTPS1 and HsTPS2 Genes
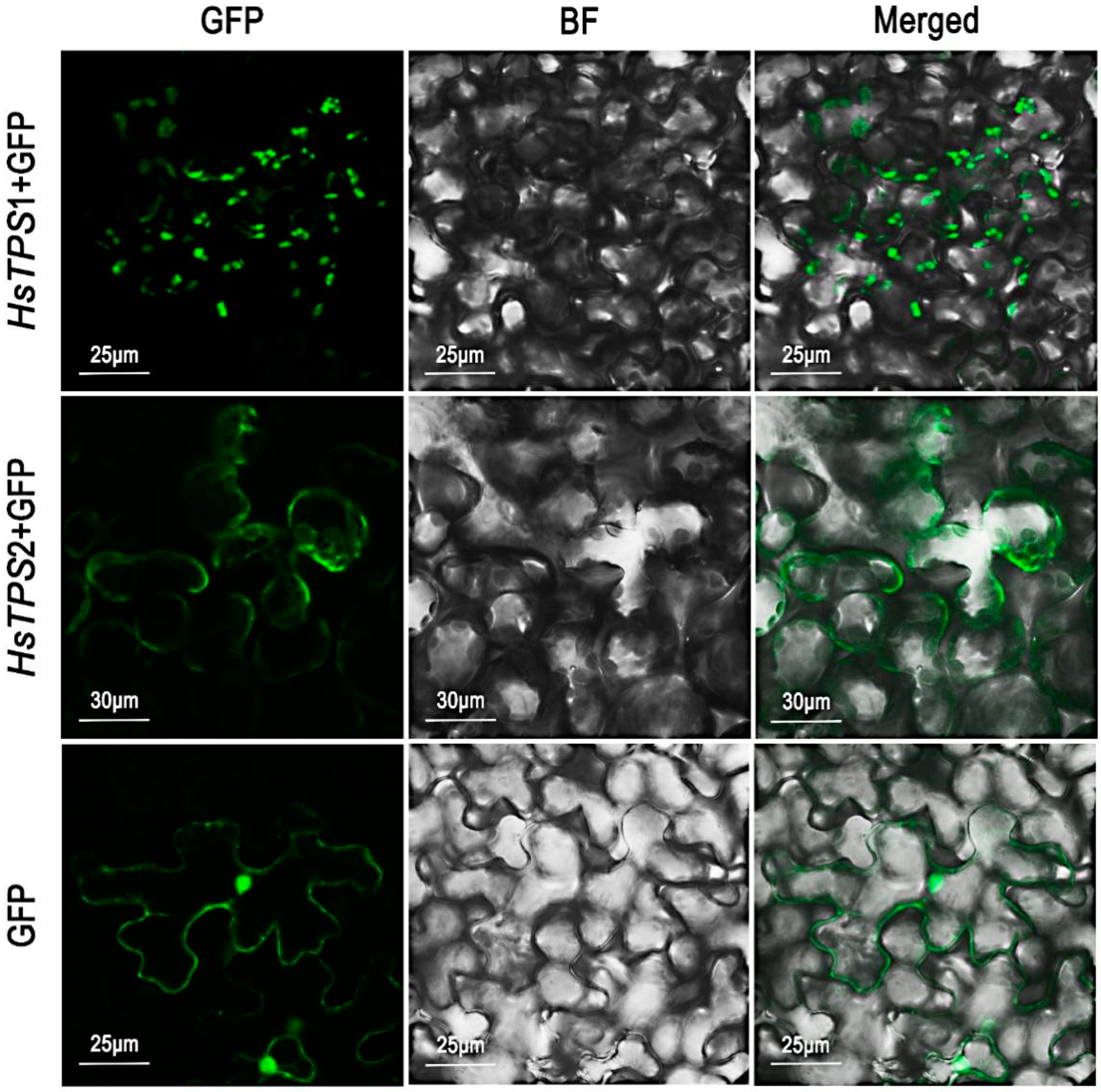
3.4. In Vivo Localization of the Two TPSs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Wang, T.; Fan, J.-M.; Liu, Z.-Z.; Zong, J.-X.; Fan, W.-X.; Han, Y.-H.; Grierson, D. Volatile composition and classification of Lilium flower aroma types and identification, polymorphisms, and alternative splicing of their monoterpene synthase genes. Hortic. Res. 2019, 6, 110. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Li, M.; Zhang, X.; Kong, W.; Bendahmane, M.; Bao, M.; Fu, X. Tissue-Specific Expression of the Terpene Synthase Family Genes in Rosa chinensis and Effect of Abiotic Stress Conditions. Genes 2022, 13, 547. [Google Scholar] [CrossRef]
- Huang, L.-M.; Huang, H.; Chuang, Y.-C.; Chen, W.-H.; Wang, C.-N.; Chen, H.-H. Evolution of Terpene Synthases in Orchidaceae. Int. J. Mol. Sci. 2021, 22, 6947. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. Cell Mol. Biol. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Nomani, M.; Noori, S.A.S.; Tohidfar, M.; Ramshini, H. Overexpression of TPS2 gene to increase thymol content using Agrobacterium tumefaciens-mediated transformation in Trachyspermum ammi (Qom ecotype)—Sciencedirect. Ind. Crop. Prod. 2018, 130, 63–70. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Stahl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Abbas, F.; Ke, Y.; Yu, R.; Yue, Y.; Amanullah, S.; Jahangir, M.M.; Fan, Y. Volatile terpenoids: Multiple functions, biosynthesis, modulation and manipulation by genetic engineering. Planta 2017, 246, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, A.; Giri, A.P.; Verstappen, F.W.; Bertea, C.M.; Sevenier, R.; Sun, Z.; Jongsma, M.A.; Schwab, W.; Bouwmeester, H.J. Gain and loss of fruit flavor compounds produced by wild and cultivated strawberry species. Plant Cell 2004, 16, 3110–3131. [Google Scholar] [CrossRef] [PubMed]
- Magnard, J.L.; Bony, A.R.; Bettini, F.; Campanaro, A.; Blerot, B.; Baudino, S.; Jullien, F. Linalool and linalool nerolidol synthases in roses, several genes for little scent. Plant Physiol. Biochem. 2018, 127, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Aros, D.; Gonzalez, V.; Allemann, R.K.; Müller, C.T.; Rosati, C.; Rogers, H.J. Volatile emissions of scented Alstroemeria genotypes are dominated by terpenes, and a myrcene synthase gene is highly expressed in scented Alstroemeria flowers. J. Exp. Bot. 2012, 63, 2739–2752. [Google Scholar] [CrossRef]
- Yue, Y.; Yu, R.; Fan, Y. Characterization of two monoterpene synthases involved in floral scent formation in Hedychium coronarium. Planta 2014, 240, 745–762. [Google Scholar] [CrossRef]
- Gao, F.; Liu, B.; Li, M.; Gao, X.; Fang, Q.; Liu, C.; Ding, H.; Wang, L.; Gao, X. Identification and characterization of terpene synthase genes accounting for volatile terpene emissions in flowers of Freesia x hybrida. J. Exp. Bot. 2018, 69, 4249–4265. [Google Scholar] [CrossRef]
- Abbas, F.; Ke, Y.; Yu, R.; Fan, Y. Functional characterization and expression analysis of two terpene synthases involved in floral scent formation in Lilium ‘Siberia’. Planta 2019, 249, 71–93. [Google Scholar] [CrossRef]
- Parachnowitsch, A.L.; Burdon, R.C.; Raguso, R.A.; Kessler, A. Natural selection on floral volatile production in Penstemon digitalis: Highlighting the role of linalool. Plant Signal. Behav. 2013, 8, e22704. [Google Scholar] [CrossRef]
- Adler, L.S.; Irwin, R.E. What you smell is more important than what you see? Natural selection on floral scent. New Phytol. 2012, 195, 510–511. [Google Scholar] [CrossRef]
- Liu, J.X.; Zhao, C.H.; Liu, X.R.; Xi, Y.Z.; Zhang, Y.L. Pollen morphology of Hosta Tratt. in China and its taxonomic signifificance. Plant Syst. Evol. 2011, 294, 99–107. [Google Scholar] [CrossRef]
- Ryu, S.H.; Walck, J.L.; Hidayati, S.N.; Rhie, Y.H.; Yang, J.C.; Lee, S.Y. Embryo growth and dormancy in seeds of six Hosta species native to Korea. Plant Species Biol. 2019, 34, 166–173. [Google Scholar] [CrossRef]
- Jo, H.; Kim, M. A new variety of Hosta (Liliaceae): Hosta clausa var. Geumgangensis. Korean J. Plant Taxon. 2016, 46, 306–313. [Google Scholar] [CrossRef][Green Version]
- He, J.W.; Yang, L.; Zhong, G.Y. Research progress in chemical constituents, pharmacological activities, clinical practices and quality control of folk medicine Hosta plantaginea. Chin. Tradit. Herb. Drugs 2016, 47, 4295–4300. [Google Scholar] [CrossRef]
- Yang, L.; He, J.W. Hosta plantaginea (Lam.) Aschers (Yuzan): An overview on its botany, traditional use, phytochemistry, quality control and pharmacology. R. Soc. Chem. Adv. 2019, 9, 35050. [Google Scholar] [CrossRef] [PubMed]
- Grenfell, D.; Shadrack, M. The Color Encyclopedia of Hostas; Timber Press: London, UK, 2004; p. 85. [Google Scholar]
- Schmid, W.G. The Genus Hosta; Timber Press: London, UK, 2009; p. 278. [Google Scholar]
- Yang, L.; Jiang, S.T.; Zhou, Q.G.; Zhong, G.Y.; He, J.W. Chemical Constituents from the Flower of Hosta plantaginea with Cyclooxygenases Inhibition and Antioxidant Activities and Their Chemotaxonomic Significance. Molecules 2017, 22, 1825. [Google Scholar] [CrossRef]
- Yang, L.; He, J. Traditional uses, phytochemistry, pharmacology and toxicological aspects of the genus Hosta (Liliaceae): A comprehensive review. J. Ethnopharmacol. 2021, 265, 113323. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. A Publ. Protein Soc. 2018, 27, 135–145. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, 320–324. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Armenteros, J.J.A.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef]
- Sindarovska, Y.; Kuchuk, M. Long-Term Potato Virus X (PVX)-Based Transient Expression of Recombinant GFP Protein in Nicotiana benthamiana Culture In Vitro. Plants 2021, 10, 2187. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.M.; Aubourg, S.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional annotation, genome organization and phylogeny of the grapevine (Vitis vinifera) terpene synthase gene family based on genome assembly, FLcDNA cloning, and enzyme assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Pichersky, E. More is better: The diversity of terpene metabolism in plants. Curr. Opin. Plant Biol. 2020, 55, 1–10. [Google Scholar] [CrossRef]
- Keilwagen, J.; Lehnert, H.; Berner, T.; Budahn, H.; Nothnagel, T.; Ulrich, D.; Dunemann, F. The Terpene Synthase Gene Family of Carrot (Daucus carota L.): Identification of QTLs and Candidate Genes Associated with Terpenoid Volatile Compounds. Front. Plant Sci. 2017, 8, 1930. [Google Scholar] [CrossRef]
- Chen, Z.; Vining, K.J.; Qi, X.; Yu, X.; Zheng, Y.; Liu, Z.; Fang, H.; Li, L.; Bai, Y.; Liang, C.; et al. Genome-Wide Analysis of Terpene Synthase Gene Family in Mentha longifolia and Catalytic Activity Analysis of a Single Terpene Synthase. Genes 2021, 12, 518. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef]
- Leferink, N.G.H.; Ranaghan, K.E.; Battye, J.; Johannissen, L.O.; Hay, S.; van der Kamp, M.; Mulholland, A.J.; Scrutton, N.S. Taming the Reactivity of Monoterpene Synthases To Guide Regioselective Product Hydroxylation. ChemBioChem 2019, 21, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Desai, J.; Liu, Y.-L.; Wei, H.; Liu, W.; Ko, T.-P.; Guo, R.-T.; Oldfield, E. Structure, Function, and Inhibition ofStaphylococcus aureusHeptaprenyl Diphosphate Synthase. ChemMedChem 2016, 11, 1915–1923. [Google Scholar] [CrossRef] [PubMed]
- Christianson, D.W. Structural and Chemical Biology of Terpenoid Cyclases. Chem. Rev. 2017, 117, 11570–11648. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral Scents and Fruit Aromas: Functions, Compositions, Biosynthesis, and Regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef]
- Green, S.A.; Chen, X.; Nieuwenhuizen, N.J.; Matich, A.J.; Wang, M.Y.; Bunn, B.J.; Yauk, Y.K.; Atkinson, R.G. Identification, functional characterization, and regulation of the enzyme responsible for floral (E)-nerolidol biosynthesis in kiwifruit (Actinidia chinensis). J. Exp. Bot. 2012, 63, 1951–1967. [Google Scholar] [CrossRef] [PubMed]
- Pazouki, L.; Niinemets, Ü. Multi-Substrate Terpene Synthases: Their Occurrence and Physiological Significance. Front. Plant Sci. 2016, 12, 1019. [Google Scholar] [CrossRef]
- Ashaari, N.S.; Ab Rahim, M.H.; Sabri, S.; Lai, K.S.; Song, A.A.; Abdul Rahim, R.; Wan Abdullah, W.M.A.N.; Ong Abdullah, J. Functional characterization of a new terpene synthase from Plectranthus amboinicus. PLoS ONE. 2020, 15, e0235416. [Google Scholar] [CrossRef]
- Zhang, X.; da Silva, J.A.T.; Niu, M.; Zhang, T.; Liu, H.; Zheng, F.; Yuan, Y.; Li, Y.; Fang, L.; Zeng, S.; et al. Functional characterization of an Indian sandalwood (Santalum album L.) dual-localized bifunctional nerolidol/linalool synthase gene involved in stress response. Phytochemistry 2021, 183, 112610. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, R.; Zhang, W.; Wei, G.; Ma, X.; Zheng, J.; Köllner, T.G.; Chen, F. Composition and Biosynthesis of Scent Compounds from Sterile Flowers of an Ornamental Plant Clematis florida cv. ‘Kaiser’. Molecules 2020, 25, 1711. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | RT 1 | Terpenoids | Relative Content 2 (%) | |
|---|---|---|---|---|
| S2 | S3 | |||
| 1 | 6.203 | α-Thujene | 0.330 ± 0.100 | 0.435 ± 0.195 |
| 2 | 6.374 | α-Pinene | 0.665 ± 0.065 | 0.990 ± 0.210 |
| 3 | 7.156 | β-Terpinene | 0.375 ± 0.135 | 0.585 ± 0.535 |
| 4 | 7.577 | β-pinene | 28.865 ± 1.095 | 37.000 ± 2.830 |
| 5 | 7.853 | α-Phellandrene | 1.060 ± 0.260 | 1.995 ± 1.335 |
| 6 | 8.011 | 2-Carene | —— | 0.095 ± 0.065 |
| 7 | 8.162 | p-Cymene | 0.050 ± 0.020 | 0.765 ± 0.345 |
| 8 | 8.287 | D-Limonene | 1.455 ± 0.415 | 8.615 ± 1.455 |
| 9 | 8.333 | β-Phellandrene | 1.575 ± 0.495 | 3.665 ± 0.305 |
| 10 | 8.491 | (E)-β-Ocimene | 0.130 ± 0.020 | 0.300 ± 0.140 |
| 11 | 8.748 | γ-Terpinene | 0.210 ± 0.040 | 0.150 ± 0.070 |
| 12 | 9.241 | Terpinolene | 0.070 ± 0.030 | 0.485 ± 0.275 |
| 13 | 9.609 | Linalool | 52.230 ± 2.210 | 40.140 ± 1.960 |
| 14 | 10.010 | 2,4(10)-Thujadiene | —— | 0.145 ± 0.065 |
| 15 | 10.885 | Terpinen-4-ol | 0.100 ± 0.030 | 0.185 ± 0.025 |
| 16 | 11.102 | α-Terpineol | 0.310 ± 0.060 | 0.235 ± 0.105 |
| 17 | 14.481 | α-Bergamotene | 0.025 ± 0.015 | —— |
| 18 | 14.652 | trans-β-Farnesene | —— | 0.085 ± 0.075 |
| 19 | 14.823 | cis-β-Farnesene | —— | 0.185 ± 0.085 |
| 20 | 15.132 | (Z,E)-πFarnesene | —— | 0.170 ± 0.080 |
| 21 | 15.316 | α-Farnesene | —— | 0.205 ± 0.065 |
| 22 | 16.053 | cis-Nerolidol | —— | 0.040 ± 0.020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, B.; Zhao, X.; Chen, G.; Qin, S.; Liu, Y.; Han, Z.; Yan, Q.; Han, L.; Liu, S.; Liu, S.; et al. Functional Analysis of Two Terpene Synthase Genes Isolated from the Flowers of Hosta ‘So Sweet’. Horticulturae 2022, 8, 447. https://doi.org/10.3390/horticulturae8050447
Cui B, Zhao X, Chen G, Qin S, Liu Y, Han Z, Yan Q, Han L, Liu S, Liu S, et al. Functional Analysis of Two Terpene Synthase Genes Isolated from the Flowers of Hosta ‘So Sweet’. Horticulturae. 2022; 8(5):447. https://doi.org/10.3390/horticulturae8050447
Chicago/Turabian StyleCui, Baiqi, Xue Zhao, Guoshuang Chen, Shijie Qin, Yitong Liu, Zhilin Han, Qian Yan, Luwen Han, Sijia Liu, Shuying Liu, and et al. 2022. "Functional Analysis of Two Terpene Synthase Genes Isolated from the Flowers of Hosta ‘So Sweet’" Horticulturae 8, no. 5: 447. https://doi.org/10.3390/horticulturae8050447
APA StyleCui, B., Zhao, X., Chen, G., Qin, S., Liu, Y., Han, Z., Yan, Q., Han, L., Liu, S., Liu, S., & Liu, H. (2022). Functional Analysis of Two Terpene Synthase Genes Isolated from the Flowers of Hosta ‘So Sweet’. Horticulturae, 8(5), 447. https://doi.org/10.3390/horticulturae8050447