
Identification, Characterization and Comparison of the Genome-Scale UTR Introns from Six Citrus Species

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Data Preparation
2.2. Intron Extraction and UI Identification
2.3. UI Characterization and Identification of Common UIs among the Six Citrus Species
2.4. Annotation and Pathway Enrichment Analysis of Common UI-Ts
3. Results
3.1. Identification of Introns in CDSs, 5′UTRs and 3′UTRs in Six Citrus Species
3.2. Size and Position Distributions of UIs in Six Citrus Species
3.3. Intron Size Comparisons among Six Citrus Species
3.4. Splice Site Conservation Analysis of UIs in Six Citrus Species
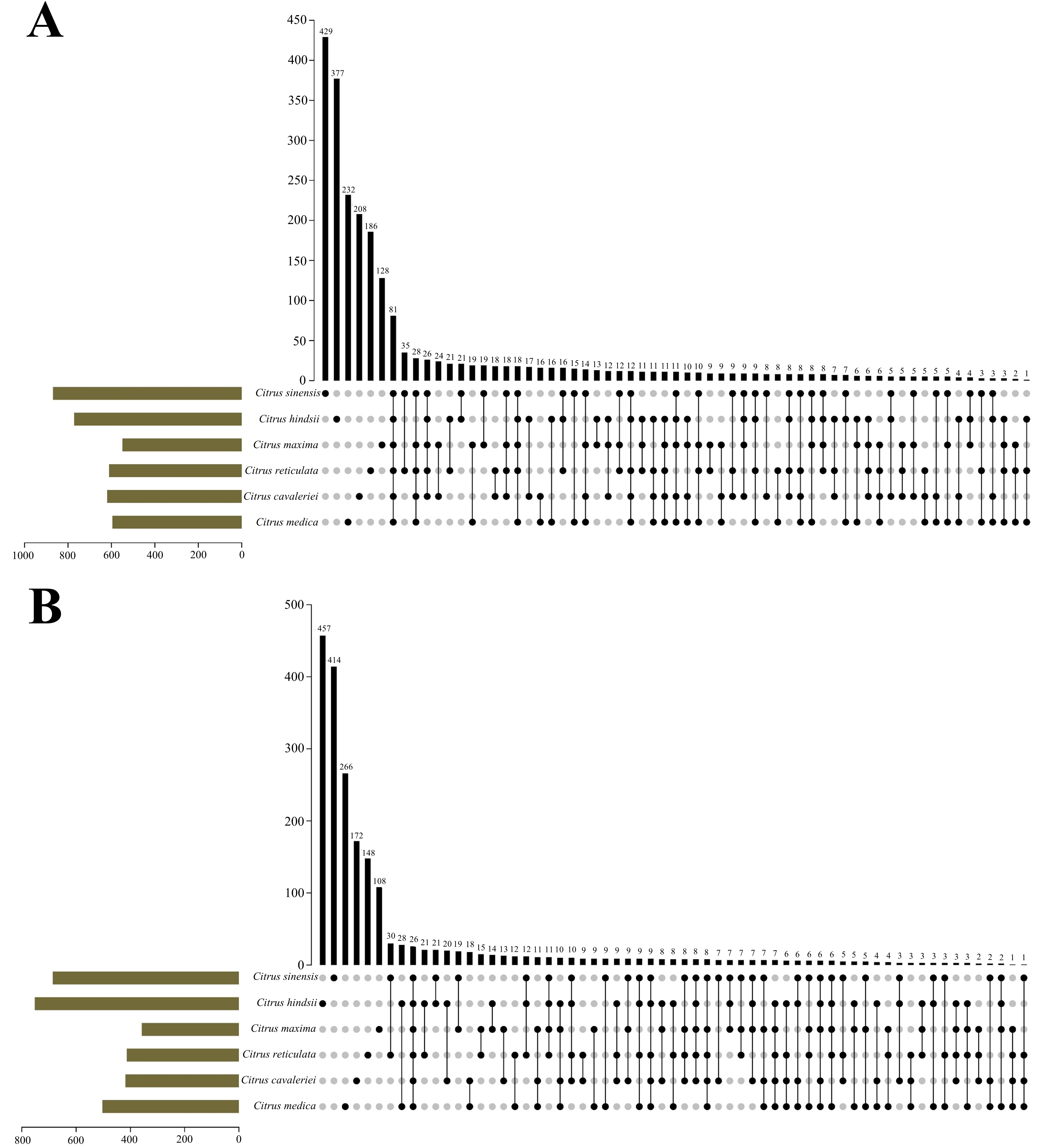
3.5. Coexistence Analysis of UIs in All the Six Citrus Species
3.6. Annotation and Enrichment Analysis of the Common UI-Containing Transcripts (UI-Ts)
4. Discussion
4.1. Characteristics of UIs in Different Citrus Species
4.2. Most Common UI-Containing Transcripts Were Involved in Gene Expression or Gene Expression Regulation
4.3. UIs Might Function in Cell Development, Stress Responses and Phytohormone Metabolism and Signaling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berget, S.M.; Moore, C.; Sharp, P.A. Spliced segments at the 5′ terminus of adenovirus 2 late mRNA. Proc. Natl. Acad. Sci. USA 1977, 74, 3171–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, L.T.; Gelinas, R.E.; Broker, T.R.; Roberts, R.J. An amazing sequence arrangement at the 5′ ends of adenovirus 2 messenger RNA. Cell 1977, 12, 1–8. [Google Scholar] [CrossRef]
- Vonk, J.; Shackelford, T. Encyclopedia of Animal Cognition and Behavior; Springer: New York, NY, USA, 2017. [Google Scholar]
- Jo, B.S.; Choi, S.S. Introns: The Functional Benefits of Introns in Genomes. Genom. Inform. 2015, 13, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, N.; Martin, W. The energetics of genome complexity. Nature 2010, 467, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M. Intron evolution as a population-genetic process. Proc. Natl. Acad. Sci. USA 2002, 99, 6118–6123. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Xia, T.; Yang, Y.; Zheng, H.; Han, X.; Jin, H.; Xiong, Z.; Qian, W.; Xia, L.; Ji, X.; Li, G.; et al. Efficient expression and function of a receptor-like kinase in wheat powdery mildew defence require an intron-located MYB binding site. Plant Biotechnol. J. 2021, 19, 897–909. [Google Scholar] [CrossRef]
- Baier, T.; Jacobebbinghaus, N.; Einhaus, A.; Lauersen, K.J.; Kruse, O. Introns mediate post-transcriptional enhancement of nuclear gene expression in the green microalga Chlamydomonas reinhardtii. PLoS Genet. 2020, 16, e1008944. [Google Scholar] [CrossRef]
- Remy, E.; Cabrito, T.R.; Batista, R.A.; Hussein, M.A.; Teixeira, M.C.; Athanasiadis, A.; Sá-Correia, I.; Duque, P. Intron retention in the 5′UTR of the novel ZIF2 transporter enhances translation to promote zinc tolerance in arabidopsis. PLoS Genet. 2014, 10, e1004375. [Google Scholar] [CrossRef] [Green Version]
- Gage, J.L.; Mali, S.; Mcloughlin, F.; Khaipho-Burch, M.; Monier, B.; Bailey-serres, J.; Vierstra, R.; Buckler, E.S. Variation in upstream open reading frames contributes to allelic diversity in protein abundance. Proc. Natl. Acad. Sci. USA 2021, 119, e2112516119. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Meshorer, E.; Ast, G. Chromatin organization marks exon-intron structure. Nat. Struct. Mol. Biol. 2009, 16, 990–995. [Google Scholar] [CrossRef]
- Lewis, B.P.; Green, R.E.; Brenner, S.E. Evidence for the widespread coupling of alternative splicing and nonsense-mediated mRNA decay in humans. Proc. Natl. Acad. Sci. USA 2003, 100, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyna, M.; Simpson, C.G.; Syed, N.H.; Lewandowska, D.; Marquez, Y.; Kusenda, B.; Marshall, J.; Fuller, J.; Cardle, L.; McNicol, J.; et al. Alternative splicing and nonsense-mediated decay modulate expression of important regulatory genes in Arabidopsis. Nucleic Acids Res. 2012, 40, 2454–2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvunis, A.R.; Rolland, T.; Wapinski, I.; Calderwood, M.A.; Yildirim, M.A.; Simonis, N.; Charloteaux, B.; Hidalgo, C.A.; Barbette, J.; Santhanam, B.; et al. Proto-genes and de novo gene birth. Nature 2012, 487, 370–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rearick, D.; Prakash, A.; McSweeny, A.; Shepard, S.S.; Fedorova, L.; Fedorov, A. Critical association of ncRNA with introns. Nucleic Acids Res. 2011, 39, 2357–2366. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.C.H.; Rasko, J.E.J. Splice and dice: Intronic microRNAs, splicing and cancer. Biomedicines 2021, 9, 1268. [Google Scholar] [CrossRef]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, H.; Nott, A.; Moore, M.J. How introns influence and enhance eukaryotic gene expression. Trends Biochem. Sci. 2003, 28, 215–220. [Google Scholar] [CrossRef]
- Oswald, A.; Oates, A.C. Control of endogenous gene expression timing by introns. Genome Biol. 2011, 12, 107. [Google Scholar] [CrossRef]
- Gottlieb, L.D.; Ford, V.S. The 5′ leader of plant PgiC has an intron: The leader shows both the loss and maintenance of constraints compared with introns and exons in the coding region. Mol. Biol. Evol. 2002, 19, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Parra, G.; Bradnam, K.; Rose, A.B.; Korf, I. Comparative and functional analysis of intron-mediated enhancement signals reveals conserved features among plants. Nucleic Acids Res. 2011, 39, 5328–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laxa, M.; Muller, K.; Lange, N.; Doering, L.; Pruscha, J.T.; Peterhansel, C. The 5′UTR Intron of Arabidopsis GGT1 Aminotransferase Enhances Promoter Activity by Recruiting RNA Polymerase II. Plant Physiol. 2016, 172, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91 Pt B, 145–155. [Google Scholar] [CrossRef]
- You, H.; Sun, B.; Li, N.; Xu, J.W. Efficient expression of heterologous genes by the introduction of the endogenous glyceraldehyde-3-phosphate dehydrogenase gene intron 1 in Ganoderma lucidum. Microb. Cell Fact. 2021, 20, 164. [Google Scholar] [CrossRef] [PubMed]
- Bicknell, A.A.; Cenik, C.; Chua, H.N.; Roth, F.P.; Moore, M.J. Introns in UTRs: Why we should stop ignoring them. Bioessays 2012, 34, 1025–1034. [Google Scholar] [CrossRef]
- Shi, X.; Wu, J.; Mensah, R.A.; Tian, N.; Liu, J.; Liu, F.; Chen, J.; Che, J.; Guo, Y.; Wu, B.; et al. Genome-wide identification and characterization of UTR-introns of Citrus sinensis. Int. J. Mol. Sci. 2020, 21, 3088. [Google Scholar] [CrossRef]
- Matsumoto, K.; Wassarman, K.M.; Wolffe, A.P. Nuclear history of a pre-mRNA determines the translational activity of cytoplasmic mRNA. EMBO J. 1998, 17, 2107–2121. [Google Scholar] [CrossRef] [Green Version]
- Furger, A.; O’Sullivan, J.M.; Binnie, A.; Lee, B.A.; Proudfoot, N.J. Promoter proximal splice sites enhance transcription. Genes Dev. 2002, 16, 2792–2799. [Google Scholar] [CrossRef] [Green Version]
- Masuda, S.; Das, R.; Cheng, H.; Hurt, E.; Dorman, N.; Reed, R. Recruitment of the human TREX complex to mRNA during splicing. Genes Dev. 2005, 19, 1512–1517. [Google Scholar] [CrossRef] [Green Version]
- Salimonti, A.; Carbone, F.; Romano, E.; Pellegrino, M.; Benincasa, C.; Micali, S.; Tondelli, A.; Conforti, F.L.; Perri, E.; Ienco, A.; et al. Association study of the 5′UTR Intron of the FAD2-2 gene with oleic and linoleic acid content in Olea europaea L. Front. Plant Sci. 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Barrou, B.M.; Gruessner, A.C.; Sutherland, D.E.; Gruessner, R.W. A potent enhancer element in the 5′-UTR intron is crucial for transcriptional regulation of the human ubiquitin C gene. Gene 2009, 448, 88–101. [Google Scholar]
- Grant, T.N.; De La Torre, C.M.; Zhang, N.; Finer, J.J. Synthetic introns help identify sequences in the 5′ UTR intron of the Glycine max polyubiquitin (Gmubi) promoter that give increased promoter activity. Planta 2017, 245, 849–860. [Google Scholar] [CrossRef] [PubMed]
- David-Assael, O.; Berezin, I.; Shoshani-Knaani, N.; Saul, H.; Mizrachy-Dagri, T.; Chen, J.X.; Brook, E.; Shaul, O. AtMHX is an auxin and ABA-regulated transporter whose expression pattern suggests a role in metal homeostasis in tissues with photosynthetic potential. Funct. Plant Biol. 2006, 33, 661–672. [Google Scholar] [CrossRef]
- Akua, T.; Shaul, O. The Arabidopsis thaliana MHX gene includes an intronic element that boosts translation when localized in a 5′ UTR intron. J. Exp. Bot. 2013, 64, 4255–4270. [Google Scholar] [CrossRef] [Green Version]
- Cenik, C.; Chua, H.N.; Zhang, H.; Tarnawsky, S.P.; Akef, A.; Derti, A.; Tasan, M.; Moore, M.J.; Palazzo, A.F.; Roth, F.P. Genome analysis reveals interplay between 5′UTR introns and nuclear mRNA export for secretory and mitochondrial genes. PLoS Genet. 2011, 7, e1001366. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.F.; Imam, J.S.; Wilkinson, M.F. The nonsense-mediated decay RNA surveillance pathway. Annu. Rev. Biochem. 2007, 76, 51–74. [Google Scholar] [CrossRef] [Green Version]
- Barrett, L.W.; Fletcher, S.; Wilton, S.D. Regulation of eukaryotic gene expression by the untranslated gene regions and other non-coding elements. Cell. Mol. Life Sci. 2012, 69, 3613–3634. [Google Scholar] [CrossRef] [Green Version]
- Kurilla, A.; Szőke, A.; Auber, A.; Káldi, K.; Silhavy, D. Expression of the translation termination factor eRF1 is autoregulated by translational readthrough and 3′UTR intron-mediated NMD in Neurospora crassa. FEBS Lett. 2020, 594, 3504–3517. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, X.; Qian, Y.; Maquat, L.E. Intron function in the nonsense-mediated decay of beta-globin mRNA: Indications that pre-mRNA splicing in the nucleus can influence mRNA translation in the cytoplasm. RNA 1998, 4, 801–815. [Google Scholar] [CrossRef]
- Weischenfeldt, J.; Damgaard, I.; Bryder, D.; Theilgaard-Monch, K.; Thoren, L.A.; Nielsen, F.C.; Jacobsen, S.E.; Nerlov, C.; Porse, B.T. NMD is essential for hematopoietic stem and progenitor cells and for eliminating by-products of programmed DNA rearrangements. Genes Dev. 2008, 22, 1381–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Saltzman, A.L.; Kim, Y.K.; Misquitta, C.; Shai, O.; Maquat, L.E.; Frey, B.J.; Blencowe, B.J. Quantitative microarray profiling provides evidence against widespread coupling of alternative splicing with nonsense-mediated mRNA decay to control gene expression. Genes Dev. 2006, 20, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltzman, A.L.; Pan, Q.; Blencowe, B.J. Regulation of alternative splicing by the core spliceosomal machinery. Genes Dev. 2011, 25, 373–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskerville, S.; Bartel, D.P. Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. RNA 2005, 11, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, B.Y.; Simons, C.; Firth, A.E.; Brown, C.M.; Hellens, R.P. Effect of 5′UTR introns on gene expression in Arabidopsis thaliana. BMC Genom. 2006, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Hong, X.; Scofield, D.G.; Lynch, M. Intron size, abundance, and distribution within untranslated regions of genes. Mol. Biol. Evol. 2006, 23, 2392–2404. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Shi, X.; Wu, J.; Zhang, Y.; Lü, P. Genome-scale computational identification and characterization of UTR introns in Atalantia buxifolia. Horticulturae 2021, 7, 556. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.B.; Hao, B.H.; Lyon, M.P.; et al. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thimm, O.; Blasing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Kruger, P.; Selbig, J.; Muller, L.A.; Rhee, S.Y.; Stitt, M. MAPMAN: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Sedehizadeh, S.; Brook, J.D.; Kozlowski, P.; Wojciechowska, M. Differential fates of introns in gene expression due to global alternative splicing. Hum. Genet. 2022, 141, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Pesole, G.; Mignone, F.; Gissi, C.; Grillo, G.; Licciulli, F.; Liuni, S. Structural and functional features of eukaryotic mRNA untranslated regions. Gene 2001, 276, 73–81. [Google Scholar] [CrossRef]
- Cenik, C.; Derti, A.; Mellor, J.C.; Berriz, G.F.; Roth, F.P. Genome-wide functional analysis of human 5′ untranslated region introns. Genome Biol. 2010, 11, R29. [Google Scholar] [CrossRef] [Green Version]
- Goguel, V.; Rosbash, M. Splice site choice and splicing efficiency are positively influenced by pre-mRNA intramolecular base pairing in yeast. Cell 1993, 72, 893–901. [Google Scholar] [CrossRef]
- Brown, J.W.; Simpson, C.G.; Thow, G.; Clark, G.P.; Jennings, S.N.; Medina-Escobar, N.; Haupt, S.; Chapman, S.C.; Oparka, K.J. Splicing signals and factors in plant intron removal. Biochem. Soc. Trans. 2002, 30, 146–149. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Jiao, X.; Tian, B.; Zhang, M.; Zhou, C.; Wang, R.; Chen, H.; Wang, B.; Li, J.; et al. Suppressor of Ty 16 promotes lung cancer malignancy and is negatively regulated by miR-1227-5p. Cancer Sci. 2020, 111, 4075–4087. [Google Scholar] [CrossRef]
- Tracy, C.; Warren, J.S.; Szulik, M.; Wang, L.; Garcia, J.; Makaju, A.; Russell, K.; Miller, M.; Franklin, S. The Smyd Family of Methyltransferases: Role in Cardiac and Skeletal Muscle Physiology and Pathology. Curr. Opin. Physiol. 2018, 1, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.C.; Tsai, Y.C.; Chou, W.L.; Liu, H.Y.; Chang, C.C.; Wu, S.J.; Lu, C.A. A CCR4-associated factor 1, OsCAF1B, confers tolerance of low-temperature stress to rice seedlings. Plant Mol. Biol. 2021, 105, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Manna, S. An overview of pentatricopeptide repeat proteins and their applications. Biochimie 2015, 113, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Pereira, A.M.; Pinto, S.C.; Silva, J.; Pereira, L.G.; Coimbra, S. In silico and expression analyses of fasciclin-like arabinogalactan proteins reveal functional conservation during embryo and seed development. Plant Reprod. 2019, 32, 353–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malumbres, M. Cyclin-dependent kinases. Genome Biol. 2014, 15, 122. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.F.; Schluttenhoffer, C.M.; Wang, P.C.; Fu, F.Y.; Thimmapuram, J.; Zhu, J.K.; Lee, S.Y.; Yun, D.J.; Mengiste, T. Cyclin-dependent kinase8 differentially regulates plant immunity to fungal pathogens through kinase-dependent and -independent functions in arabidopsis. Plant Cell 2014, 26, 4149–4170. [Google Scholar] [CrossRef] [Green Version]
- Slane, D.; Reichardt, I.; El Kasmi, F.; Bayer, M.; Jurgens, G. Evolutionarily diverse SYP1 Qa-SNAREs jointly sustain pollen tube growth in Arabidopsis. Plant J. 2017, 92, 375–385. [Google Scholar] [CrossRef]
- Giri, J.; Vij, S.; Dansana, P.K.; Tyagi, A.K. Rice A20/AN1 zinc-finger containing stress-associated proteins (SAP1/11) and a receptor-like cytoplasmic kinase (OsRLCK253) interact via A20 zinc-finger and confer abiotic stress tolerance in transgenic Arabidopsis plants. New Phytol. 2011, 191, 721–732. [Google Scholar] [CrossRef]
- Kim, G.D.; Cho, Y.H.; Yoo, S.D. Regulatory functions of evolutionarily conserved AN1/A20-like Zinc finger family proteins in Arabidopsis stress responses under high temperature. Biochem. Biophys. Res. Commun. 2015, 457, 213–220. [Google Scholar] [CrossRef]
- Stuhrwohldt, N.; Hartmann, J.; Dahlke, R.I.; Oecking, C.; Sauter, M. The PSI family of nuclear proteins is required for growth in arabidopsis. Plant Mol. Biol. 2014, 86, 289–302. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Position | No. of Sequences | Sequences with Introns | Total Bases (Genomic) | Intron/Sequence | No. of UIs | No. of Introns/Nucleotides (mRNA) |
|---|---|---|---|---|---|---|---|
| C. sinensis * | 5′UTR | 16,916 | 617 | 6.80 × 106 | 0.06 | 965 | 1.42 × 10−4 |
| CDS | 23,394 | 17,897 | 3.77 × 107 | 4.81 | - | 2.98 × 10−3 | |
| 3′UTR | 17,408 | 469 | 1.17 × 107 | 0.04 | 745 | 6.36 × 10−5 | |
| C. hindsii | 5′UTR | 17,275 | 567 | 1.32 × 107 | 0.05 | 935 | 7.07 × 10−5 |
| CDS | 32,257 | 24,326 | 4.18 × 107 | 4.16 | - | 3.21 × 10−3 | |
| 3′UTR | 17,160 | 482 | 2.61 × 107 | 0.05 | 854 | 3.27 × 10−5 | |
| C. maxima | 5′UTR | 13,330 | 408 | 4.12 × 106 | 0.05 | 604 | 1.47 × 10−4 |
| CDS | 30,123 | 22,186 | 3.42 × 107 | 3.68 | - | 3.25 × 10−3 | |
| 3′UTR | 14,144 | 244 | 8.94 × 106 | 0.03 | 385 | 4.31× 10−5 | |
| C. reticulata | 5′UTR | 13,784 | 430 | 5.32 × 106 | 0.05 | 675 | 1.27 × 10−4 |
| CDS | 28,833 | 21,372 | 3.48 × 107 | 3.87 | - | 3.20 × 10−3 | |
| 3′UTR | 14,127 | 274 | 1.08 × 107 | 0.03 | 446 | 4.14 × 10−5 | |
| C. cavaleriei | 5′UTR | 14,336 | 450 | 5.45 × 106 | 0.05 | 683 | 1.25 × 10−4 |
| CDS | 32,067 | 24,499 | 3.45 × 107 | 3.74 | - | 3.47 × 10−3 | |
| 3′UTR | 14,116 | 280 | 9.74 × 106 | 0.03 | 450 | 4.62 × 10−5 | |
| C. medica | 5′UTR | 15,240 | 444 | 6.48 × 106 | 0.04 | 682 | 1.05 × 10−4 |
| CDS | 32,579 | 24,411 | 3.56 × 107 | 3.72 | - | 3.40 × 10−3 | |
| 3′UTR | 15,502 | 349 | 1.40 × 107 | 0.04 | 572 | 4.07 × 10−5 |
| Species | Intron Type | Mean | Median | LQ | UQ |
|---|---|---|---|---|---|
| C. sinensis * | 5UI | 587.50 | 450 | 168 | 836 |
| CI | 343.20 | 171 | 102 | 441 | |
| 3UI | 563.5 | 335 | 139 | 730 | |
| C. hindsii | 5UI | 818.16 | 472 | 165 | 925 |
| CI | 463.07 | 172 | 102 | 457 | |
| 3UI | 930.25 | 459 | 141 | 981.25 | |
| C. maxima | 5UI | 665.42 | 475.5 | 180.75 | 851.5 |
| CI | 468.96 | 174 | 102 | 449 | |
| 3UI | 751.44 | 508 | 159 | 960 | |
| C. reticulata | 5UI | 1594.74 | 494 | 186.5 | 977 |
| CI | 461.40 | 176 | 102 | 459 | |
| 3UI | 858.35 | 473 | 154.5 | 955.5 | |
| C. cavaleriei | 5UI | 654.86 | 506 | 197.5 | 881.5 |
| CI | 433.85 | 177 | 102 | 463 | |
| 3UI | 691.95 | 435 | 153.25 | 928.25 | |
| C. medica | 5UI | 682.38 | 485 | 198.25 | 914.75 |
| CI | 403.59 | 177 | 102 | 462 | |
| 3UI | 742.33 | 428.5 | 150 | 861 |
| Citrus Species | UI Type | SS Pair Type | No. | Percentage |
|---|---|---|---|---|
| C. sinensis | 3UIs | GT-AG | 724 | 97.18% |
| GC-AG | 16 | 2.14% | ||
| AT-AC | 2 | 0.26% | ||
| GT-TG | 1 | 0.13% | ||
| TA-AG | 1 | 0.13% | ||
| TT-AG | 1 | 0.13% | ||
| 5UIs | GT-AG | 945 | 97.92% | |
| GC-AG | 14 | 1.45% | ||
| AT-AC | 2 | 0.20% | ||
| CT-AC | 2 | 0.20% | ||
| GT-GG | 1 | 0.10% | ||
| TG-AG | 1 | 0.10% | ||
| C. hindsii | 3UIs | GT-AG | 823 | 96.37% |
| GC-AG | 16 | 1.87% | ||
| AT-AC | 15 | 1.75% | ||
| 5UIs | GT-AG | 909 | 97.21% | |
| GC-AG | 20 | 2.13% | ||
| AT-AC | 6 | 0.64% | ||
| C. maxima | 3UIs | GT-AG | 371 | 96.36% |
| GC-AG | 10 | 2.59% | ||
| AT-AC | 4 | 1.03% | ||
| 5UIs | GT-AG | 589 | 97.51% | |
| GC-AG | 9 | 1.49% | ||
| AT-AC | 6 | 0.99% | ||
| C. reticulata | 3UIs | GT-AG | 429 | 96.18% |
| GC-AG | 15 | 3.36% | ||
| AT-AC | 2 | 0.44% | ||
| 5UIs | GT-AG | 657 | 97.33% | |
| GC-AG | 13 | 1.92% | ||
| AT-AC | 5 | 0.74% | ||
| C. cavaleriei | 3UIs | GT-AG | 434 | 96.44% |
| GC-AG | 13 | 2.88% | ||
| AT-AC | 3 | 0.66% | ||
| 5UIs | GT-AG | 666 | 97.51% | |
| GC-AG | 16 | 2.34% | ||
| AT-AC | 1 | 0.14% | ||
| C. medica | 3UIs | GT-AG | 547 | 95.62% |
| AT-AC | 12 | 2.09% | ||
| GC-AG | 9 | 1.57% | ||
| CA-AG | 1 | 0.17% | ||
| TA-CA | 1 | 0.17% | ||
| TT-AT | 1 | 0.17% | ||
| TT-CT | 1 | 0.17% | ||
| 5UIs | GT-AG | 665 | 97.50% | |
| GC-AG | 9 | 1.31% | ||
| AT-AC | 3 | 0.43% | ||
| AT-GA | 1 | 0.14% | ||
| GG-GC | 1 | 0.14% | ||
| TA-CT | 1 | 0.14% | ||
| TT-AT | 1 | 0.14% | ||
| TT-TA | 1 | 0.14% |
| BINcode | BinName | Gene ID | Description | Type |
|---|---|---|---|---|
| 3.13.3.1 | Carbohydrate metabolism. nucleotide sugar biosynthesis. UDP-D-glucuronic acid biosynthesis. UDP-D-glucose 6-dehydrogenase | cs6g22050.1 | UDP-D-glucose 6-dehydrogenase | 5UI-T |
| 7.3.1 | Coenzyme metabolism. S-adenosyl methionine (SAM) cycle. S-adenosyl methionine synthetase (MAT) | cs6g01310.2 | S-adenosyl methionine synthetase | 5UI-T |
| cs9g01410.1 | S-adenosyl methionine synthetase | 5UI-T | ||
| 10.4.2.4 | Redox homeostasis. thiol-based redox regulation. peroxiredoxin activities. type-2 peroxiredoxin (PrxII) | cs6g15550.2 | type-2 peroxiredoxin (PrxII) | 3UI-T |
| 11.1.2.1.3 | Phytohormone action. abscisic acid. perception and signalling. receptor activities. regulatory protein (EAR1) | cs8g14510.2 | regulatory protein (EAR1) of abscisic acid signaling | 5UI-T |
| 11.2.4.2 | Phytohormone action. auxin. transport. auxin efflux transporter (PILS) | cs2g13710.1 | auxin efflux transporter (PILS) | 5UI-T |
| 11.10.2.4.2 | Phytohormone action. signalling peptides. CRP (cysteine-rich-peptide) category. RALF/RALFL-peptide activity. RALF-peptide receptor (CrRLK1L) | cs6g10250.2 | RALF-peptide receptor (CrRLK1L) | 5UI-T |
| 12.2.2.1 | Chromatin organisation. histone chaperone activities. FACT histone chaperone complex. component SPT16 | cs6g01200.1 | component SPT16 of FACT histone chaperone complex | 5UI-T |
| 12.3.1.1.7 | Chromatin organisation. post-translational histone modification. histone methylation. lysine methylation. class-VI histone methyltransferase (SMYD) | cs8g12080.2 | class-VI histone methyltransferase (SMYD) | 3UI-T |
| 13.2.1.2.5 | Cell division. cell cycle organisation. cell cycle control. CYCLIN-dependent protein kinase complex. catalytic component CDKE | cs5g01900.2 | protein kinase (CDKE/CDK8) | 5UI-T |
| 13.4.2.3.1 | Cell division. cytokinesis. cell-plate formation. SNARE cell-plate vesicle fusion complex. Qa-SNARE component KNOLLE | cs5g26090.1 | SYP1-group Qa-type SNARE component | 5UI-T |
| 15.1.4.6 | RNA biosynthesis. DNA-dependent RNA polymerase complexes. RNA polymerase IV complex. subunit NRPD7 | cs9g11150.1 | subunit NRPD7 of RNA polymerase IV complex | 5UI-T |
| 15.1.5.6 | RNA biosynthesis. DNA-dependent RNA polymerase complexes. RNA polymerase V complex. subunit NRPE7 | cs9g11150.1 | subunit NRPE7 of RNA polymerase V complex | 5UI-T |
| 15.3.4.2.6 | RNA biosynthesis. RNA polymerase II-dependent transcription. transcription co-activation. TFIId complex. component TAF8 | cs6g22280.1 | component TAF8 of TFIId basal transcription regulation complex | 5UI-T |
| 15.3.4.3.3.2 | RNA biosynthesis. RNA polymerase II-dependent transcription. transcription co-activation. SAGA complex. SPT recruitment module. component ADA1 | cs1g21850.2 | component ADA1 of SAGA transcription co-activator complex | 5UI-T |
| cs7g07280.1 | component ADA1 of SAGA transcription co-activator complex | 5UI-T | ||
| 15.3.4.4.4.3 | RNA biosynthesis. RNA polymerase II-dependent transcription. transcription co-activation. MEDIATOR complex. regulatory kinase module. component CDK8 | cs5g01900.2 | protein kinase (CDKE/CDK8) | 5UI-T |
| 15.5.2.2 | RNA biosynthesis. transcriptional regulation. MYB transcription factor superfamily. transcription factor (MYB-related) | cs1g24225.1 | transcription factor (MYB-related) | 3UI-T |
| 15.5.12 | RNA biosynthesis. transcriptional regulation. transcription factor (GRAS) | cs4g12130.1 | transcription factor (GRAS) | 5UI-T |
| cs7g02550.1 | transcription factor (GRAS) | 5UI-T | ||
| 15.5.20 | RNA biosynthesis. transcriptional regulation. transcription factor (Trihelix) | cs4g16730.1 | transcription factor (Trihelix) | 3UI-T |
| 15.5.30 | RNA biosynthesis. transcriptional regulation. transcription factor (bHLH) | cs4g02590.1 | transcription factor (bHLH) | 5UI-T |
| cs5g30170.2 | transcription factor (bHLH) | 5UI-T | ||
| 15.5.32 | RNA biosynthesis. transcriptional regulation. transcription factor (BBR/BPC) | orange1.1t01638.1 | transcription factor (BBR/BPC) | 5UI-T |
| 15.6.2.2 | RNA biosynthesis. organelle machinery. transcriptional regulation. transcription factor (mTERF) | cs5g31960.1 | transcription factor (mTERF) | 5UI-T |
| cs8g01080.1 | transcription factor (mTERF) | 5UI-T | ||
| 16.1.1.2.8 | RNA processing. pre-RNA splicing. U2-type-intron-specific major spliceosome. U2 small nuclear ribonucleoprotein particle (snRNP). pre-mRNA splicing factor (SF1) | cs9g15030.1 | pre-mRNA splicing factor (SF1) | 3UI-T |
| 16.4.9.4 | RNA processing. RNA homeostasis. mRNA stress granule formation. regulatory protein (UBA1/2) of UBP1 activity | cs6g16060.1 | regulatory protein (UBA1/2) of UBP1 activity | 3UI-T |
| cs7g25330.3 | regulatory protein (UBA1/2) of UBP1 activity | 3UI-T | ||
| 16.5.2.3.3 | RNA processing. mRNA silencing. miRNA pathway. miRNA degradation. regulatory protein (HWS) | orange1.1t00443.1 | regulatory protein (HWS) of miRNA degradation | 5UI-T |
| 16.6.1.1.11 | RNA processing. organelle machinery. pre-RNA splicing. plastidial RNA splicing. splicing factor (mTERF4) | cs5g31960.1 | mTERF4 plastidial RNA splicing factor | 5UI-T |
| 16.6.2.2.4.7 | RNA processing. organelle machinery. RNA modification. C-to-U RNA editing. PPR-type RNA editing factor activities. RNA editing factor (MEF9) | cs4g13530.1 | RNA editing factor (MEF9) | 3UI-T |
| 17.1.2.2.2.5 | Protein biosynthesis. ribosome biogenesis. large ribosomal subunit (LSU). LSU processome. pre-60S ribosomal subunit nuclear export. export factor (NMD3) | cs6g17980.1 | pre-60S subunit nuclear export factor (NMD3) | 5UI-T |
| 17.1.3.2.1.3.1 | Protein biosynthesis. ribosome biogenesis. small ribosomal subunit (SSU). SSU processome. pre-40S ribosomal subunit nuclear assembly. UtpB module. assembly factor (UTP18) | cs5g30340.1 | SSU processome assembly factor (UTP18) | 5UI-T |
| 17.3.1.1.2 | Protein biosynthesis. translation initiation. Pre-Initiation Complex (PIC) module. eIF1 PIC assembly factor activity. assembly factor (eIF1A) | cs2g20280.1 | assembly factor (eIF1A) of eIF1 | 5UI-T |
| 17.3.1.2.2 | Protein biosynthesis. translation initiation. Pre-Initiation Complex (PIC) module. eIF2 Met-tRNA binding factor activity. activating factor (eIF5) of eIF2-GTP hydrolysis | cs3g18950.1 | activating factor (eIF5) of eIF2-GTP hydrolysis | 5UI-T |
| 18.2.4 | Protein modification. acetylation. N-terminal acetylase (NatD) | cs1g20790.1 | N-terminal acetylase (NatD) | 5UI-T |
| 18.3.4.1.1.2 | Protein modification. lipidation. glycophosphatidylinositol (GPI) anchor addition. GPI pre-assembly. GPI N-acetylglucosamine transferase complex. component PIG-C | cs2g11690.2 | component PIG-C of GPI N-acetylglucosamine transferase complex | 5UI-T |
| 18.4.1.16 | Protein modification. phosphorylation. TKL protein kinase superfamily. protein kinase (CrlRLK1) | cs6g10250.2 | protein kinase (CrlRLK1) | 5UI-T |
| 18.4.3.1.5 | Protein modification. phosphorylation. CMGC protein kinase superfamily. CDK protein kinase families. protein kinase (CDKE/CDK8) | cs5g01900.2 | protein kinase (CDKE/CDK8) | 5UI-T |
| 18.13.1 | Protein modification. protein folding. protein folding catalyst (Cyclophilin) | cs8g12840.1 | protein folding catalyst | 5UI-T |
| 19.1.2.3 | Protein homeostasis. protein quality control. ribosome-associated chaperone activities. co-chaperone (ZRF) | cs6g08770.1 | Hsp40-chaperone ZRF ribosome-associated chaperone complex | 5&3UI-T |
| 19.2.1.3.1.2 | Protein homeostasis. ubiquitin-proteasome system. N-degron pathways. Pro/N-degron pathway. GID ubiquitination complex. ubiquitin ligase component GID2 | cs8g03080.1 | ubiquitin ligase component GID2 of GID ubiquitination complex | 5UI-T |
| 19.2.2.1.4.3.3.2 | Protein homeostasis. ubiquitin-proteasome system. ubiquitin-fold protein conjugation. ubiquitin conjugation (ubiquitylation). ubiquitin-ligase E3 activities. RING-domain E3 ligase activities. RING-H2-class ligase activities. BTL-subclass ligase | cs6g16300.2 | RING-H2-class E3 BTL-subclass ubiquitin ligase | 5UI-T |
| 19.2.2.8.1.4.3 | Protein homeostasis. ubiquitin-proteasome system. ubiquitin-fold protein conjugation. Cullin-based ubiquitylation complexes. SKP1-CUL1-FBX (SCF) E3 ubiquitin ligase complexes. F-BOX substrate adaptor activities. substrate adaptor (FBX) | orange1.1t00443.1 | substrate adaptor FBX of SCF E3 ubiquitin ligase complex | 5UI-T |
| 19.2.5.2.2.3 | Protein homeostasis. ubiquitin-proteasome system. 26S proteasome. 19S regulatory particle. lid subcomplex. regulatory component RPN6 | cs4g04180.1 | regulatory component RPN6 of 26S proteasome | 5UI-T |
| 21.4.1.1.3 | Cell wall organisation. cell wall proteins. hydroxyproline-rich glycoprotein activities. arabinogalactan-protein activities. Fasciclin-type arabinogalactan protein (FLA) | cs2g20030.1 | arabinogalactan protein (Fasciclin) | 5UI-T |
| cs2g20030.2 | arabinogalactan protein (Fasciclin) | 5UI-T | ||
| cs2g20030.3 | arabinogalactan protein (Fasciclin) | 5UI-T | ||
| 22.1.1.1.1 | Vesicle trafficking. anterograde trafficking. Coat protein II (COPII) coatomer machinery. coat protein complex. scaffolding component Sec13 | cs2g28780.1 | scaffolding component Sec13 of coat protein complex | 5UI-T |
| 22.3.1.1.2 | Vesicle trafficking. endocytic trafficking. ESCRT-mediated sorting. ESCRT-I complex. component VPS28 | cs2g06750.1 | component VPS28 of ESCRT-I complex | 5UI-T |
| 22.5.2.4.3.2 | Vesicle trafficking. multi-pathway trafficking regulation. vesicle tethering. RAB-GTPase membrane association. RAB-GDI displacement factor (GDF) activities. B-G-class Rab-GDF protein | cs7g30390.2 | B-G-class Rab-GDF protein | 3UI-T |
| 22.5.3.1.1.1 | Vesicle trafficking. multi-pathway trafficking regulation. target membrane fusion. SNARE membrane fusion complexes. Qa-type SNARE components. SYP1-group component | cs5g26090.1 | SYP1-group Qa-type SNARE component | 5UI-T |
| 23.1.2.2 | Protein translocation. chloroplast. outer envelope TOC translocation system. receptor GTPase (Toc90/120/132/159) | cs8g12230.1 | component Toc90/120/132/159 of outer envelope TOC translocation system | 5UI-T |
| 23.5.2.3.2 | Protein translocation. nucleus. nucleocytoplasmic transport. RAN GTPase cycle. Ran-activating protein (Ran-GAP) | cs9g06440.1 | Ran-activating protein of nucleocytoplasmic transport | 5UI-T |
| 24.2.5.2.2 | Solute transport. carrier-mediated transport. BART superfamily. AEC family. auxin transporter (PILS) | cs2g13710.1 | auxin transporter (PILS) | 5UI-T |
| 26.9.2.2.4 | External stimuli response. pathogen. effector-triggered immunity (ETI) network. RIN4-RPM1 immune signalling. regulatory protein (GCN4) of RIN4 activity | cs1g13980.1 | regulatory protein (GCN4) of RIN4 activity | 5UI-T |
| 27.6.1.4.5 | Multi-process regulation. phosphatidylinositol and inositol phosphate system. biosynthesis. phosphatidylinositol kinase activities. phosphatidylinositol 4-kinase (PI4K-gamma) | cs7g12040.1 | phosphatidylinositol 4-kinase (PI4K-gamma) | 5UI-T |
| 35.1 | not assigned. annotated | cs1g11100.1 | probable CCR4-associated factor 1 homolog 6 | 5UI-T |
| cs1g11700.2 | F-box/LRR-repeat protein 14 | 5UI-T | ||
| cs2g11780.1 | pentatricopeptide repeat-containing protein | 3UI-T | ||
| cs2g18610.1 | transmembrane 9 superfamily member 12 | 5UI-T | ||
| cs3g20090.2 | pentatricopeptide repeat-containing protein | 5UI-T | ||
| cs4g03945.1 | pentatricopeptide repeat-containing protein | 3UI-T | ||
| cs5g03910.1 | putative pentatricopeptide repeat-containing protein | 5&3UI-T | ||
| cs5g09740.1 | zinc finger A20 and AN1 domain-containing stress-associated protein 1 | 5UI-T | ||
| cs6g04970.2 | serine/threonine-protein kinase ATM | 5&3UI-T | ||
| cs6g07760.1 | pentatricopeptide repeat-containing protein | 3UI-T | ||
| cs6g08820.1 | pentatricopeptide repeat-containing protein | 3UI-T | ||
| cs6g08820.2 | pentatricopeptide repeat-containing protein | 3UI-T | ||
| cs6g15060.2 | zinc finger A20 and AN1 domain-containing stress-associated protein 4 | 5UI-T | ||
| cs6g15600.2 | protein PSK SIMULATOR 1 | 5UI-T | ||
| cs7g02570.1 | calmodulin binding protein PICBP | 3UI-T | ||
| cs7g04050.2 | F-box/Kelch-repeat protein SKIP11 | 5UI-T | ||
| cs7g04620.1 | pentatricopeptide repeat-containing protein | 5&3UI-T | ||
| cs7g15390.1 | pentatricopeptide repeat-containing protein | 3UI-T | ||
| cs7g19080.2 | FT-interacting protein 3 | 5UI-T | ||
| cs8g04770.1 | F-box/Kelch-repeat protein | 3UI-T | ||
| cs8g12590.2 | pentatricopeptide repeat-containing protein | 5UI-T | ||
| cs8g16750.1 | F-box/Kelch-repeat protein | 5UI-T | ||
| cs9g04270.1 | chaperone protein dnaJ 49 | 3UI-T | ||
| cs9g04270.2 | chaperone protein dnaJ 49 | 5UI-T | ||
| orange1.1t00345.2 | FT-interacting protein 3 | 5UI-T | ||
| orange1.1t04379.2 | FT-interacting protein 3 | 5UI-T | ||
| 35.2 | not assigned. not annotated | cs1g26580.1 | unknown | 5UI-T |
| cs1g26840.2 | unknown | 5UI-T | ||
| cs2g13570.1 | unknown | 5UI-T | ||
| cs2g19060.1 | unknown | 5UI-T | ||
| cs2g21340.2 | unknown | 5UI-T | ||
| cs3g11280.1 | unknown | 5UI-T | ||
| cs3g16520.2 | unknown | 5UI-T | ||
| cs4g16820.2 | unknown | 3UI-T | ||
| cs4g17430.1 | unknown | 3UI-T | ||
| cs4g18220.2 | unknown | 5UI-T | ||
| cs5g24290.1 | unknown | 5UI-T | ||
| cs5g27890.1 | unknown | 5UI-T | ||
| cs6g13200.1 | unknown | 5UI-T | ||
| cs7g09380.2 | unknown | 5UI-T | ||
| cs7g10870.2 | unknown | 5UI-T | ||
| cs7g24520.1 | unknown | 5UI-T | ||
| cs8g16190.1 | unknown | 5UI-T | ||
| cs9g02430.1 | unknown | 5UI-T | ||
| cs9g11810.2 | unknown | 5UI-T | ||
| orange1.1t00607.1 | unknown | 5UI-T | ||
| orange1.1t01667.1 | unknown | 5UI-T | ||
| orange1.1t02210.1 | unknown | 3UI-T | ||
| orange1.1t05845.1 | unknown | 5UI-T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, C.; Shi, X.; Zhang, Y.; Wang, B.; Wu, J.; Yang, S.; Wang, S. Identification, Characterization and Comparison of the Genome-Scale UTR Introns from Six Citrus Species. Horticulturae 2022, 8, 434. https://doi.org/10.3390/horticulturae8050434
Cheng C, Shi X, Zhang Y, Wang B, Wu J, Yang S, Wang S. Identification, Characterization and Comparison of the Genome-Scale UTR Introns from Six Citrus Species. Horticulturae. 2022; 8(5):434. https://doi.org/10.3390/horticulturae8050434
Chicago/Turabian StyleCheng, Chunzhen, Xiaobao Shi, Yongyan Zhang, Bin Wang, Junwei Wu, Shizao Yang, and Shaohua Wang. 2022. "Identification, Characterization and Comparison of the Genome-Scale UTR Introns from Six Citrus Species" Horticulturae 8, no. 5: 434. https://doi.org/10.3390/horticulturae8050434
APA StyleCheng, C., Shi, X., Zhang, Y., Wang, B., Wu, J., Yang, S., & Wang, S. (2022). Identification, Characterization and Comparison of the Genome-Scale UTR Introns from Six Citrus Species. Horticulturae, 8(5), 434. https://doi.org/10.3390/horticulturae8050434