Abstract
Hoya R.Br. is a genus of ornamental plants with an attractive flower shape and unique scents. Anatomical studies are required to understand their structure that produces, and stores, the metabolites released by plants. The present study was conducted to determine the type and position of floral glands in three Hoya species: Hoya cagayanensis C.M Burton, Hoya lacunosa Blume, and Hoya coriacea Blume. The investigations were carried out using light microscopy (LM), scanning electron microscopy (SEM), and histochemical staining tests. Secondary nectaries were present in the corona lobe portion, while simple trichomes (unicellular) were found in the petals’ adaxial epidermis of all the studied species. Conical trichomes were found in H. cagayanensis, cylindrical trichomes in H. lacunosa, and falcate trichomes in H. coriacea. In the cells of secondary nectaries, this study revealed proteins, lipids, polysaccharides, and starch grains; however, only lipids and proteins were observed in simple trichomes. Secondary nectaries and simple trichomes were described for the first time in all the studied species, contributing new insight into Hoya’s anatomical and micromorphological floral glands.
1. Introduction
Hoya is the largest genus of Marsdenieae, a subfamily Asclepiadoideae, Apocynaceae, with 300 species found in tropical and subtropical Asia, Oceania, and the Pacific islands [1,2,3]. Species of Hoya R.Br are ornamental plants with unique, attractive flowers and distinctive scents [4]. The stellate flowers come in various colours, including white, bright yellow, pink, orange, green, and blackish purple [5]. In Peninsular Malaysia, 75 Hoya species have been recorded [6,7,8,9,10]. Because of its waxy flower, Hoya spp. is known as a waxflower. They secrete latex after wounding. [11,12].
Like most other flowering plants, Hoya emits unique and various scents that act as a medium of communication to insects, between plants, and in response to pathogens [12]. The scent released results from the biosynthesis of the chemical compounds in the plant tissues and cells [13]. The production and secretion of chemical compounds in plants are often associated with specialized floral glands: osmophores, nectaries, and glandular trichomes [14,15,16,17]. Studies of floral glands in the Asclepiadoideae have identified 13 types of floral glands: colleters, glandular trichomes, laticifers, idioblasts, nectary, osmophore, stylar canal, obturator, style head, staminal wing glands, extragynoecial compitum, and tapetum [18,19,20,21,22,23,24,25,26]. Floral glands in the Asclepiadoideae have been associated with the production of various chemical compounds and categorized as protective glands that act for defensive purposes, or as nuptial glands, which aid in pollination [18].
As a type of nuptial gland, nectaries produce nectar and store sweet substances [27,28]. Nectar consists of sugars (sucrose, glucose, and fructose), proteins, amino acids, and lipids [28,29,30,31], which play an essential role as a reward to pollinators [32]. In Asclepiadoide, two types of nectaries were reported based on their position and function: primary nectaries located in stigmatic chambers and secondary nectaries in the corona lobe [18,19,20,21,33,34,35,36,37,38,39,40,41,42]. The primary nectaries in the stigmatic chambers could serve as a reward for pollinators and a pollen germination inducer. On the other hand, flowering with two types of nectaries may divide these functions: the primary nectary acts as a pollen inducer for germination. Therefore, there is a question of whether this structure can be considered as a nectary, if it does not provide nectar for pollinators. Possibly, it is rather a secretory structure that belongs to the transmitting tracts, contributing to the stigmatic fluid for interactions of the sporophyte with male gametophytes [18]. In contrast, the secondary nectary provides nectar for pollinators [18,20,42].
Trichomes were discovered for the first time in the Asclepiadoideae in 1969, in Matelea denticulata [18,43]. Those studies found multicellular glandular trichomes with stinging-type trichomes on the flower’s pedicel and abaxial sepal portions. Simple trichomes protect plants from various environmental stresses [44], while glandular trichomes are the main biosynthesis area for producing and releasing chemical compounds in plants [45,46]. Therefore, knowledge about the secretion cell structures, chemical compounds, and their function in flowering plants is essential.
No studies on Hoya have ever been reported comprehensively to understand the anatomical and micromorphological structures that are possibly related to the secretion of chemical compounds. Our previous study found that three Hoya species exhibited varied floral scent profiles, with β-ocimene and methyl salicylate compounds dominating the aroma in H. cagayanensis. H. lacunosa had the highest levels of 1-octane-3-ol, whereas H. coriacea had the highest levels of (Z)-acid butyric, 3-hexenyl ester [47]. In plants, aroma and scent release are often associated with the floral glands, chemical compounds produced in specific cells, and genes involved in metabolite biosynthesis [45,47]. Furthermore, these chemical compounds are believed to act as one of the attractants of pollination agents to assist in the pollination process. Therefore, our study aimed to identify the floral glands and their position in flowers of three Hoya species.
2. Materials and Methods
2.1. Plant Materials
The three flowering Hoya species were collected due to their diverse morphologies, floral aroma, and virtually matching colour groups (whitish to yellow) [47]. Flower samples were collected from wild-growing plants at the anthesis stage in October 2020. Three flowers from three individual plants were collected and studied for each species. All voucher specimens were deposited in the Herbarium of Universiti Kebangsaan Malaysia (UKMB) (Table 1).

Table 1.
List of flower species, vouchers, and collection locality.
2.2. Light Microscopy (LM)
The fresh flower samples were fixed in Carnoy’s fixative (Sigma-Aldrich, St. Louis, MO, USA) (acetic acid solution and 70% ethanol in a 1:3 ratio) [48] for 48 h at room temperature. The samples were then rinsed with an ethanol series and distilled water before sectioning. For LM, the sections (10–15-µm thick) were obtained using a sliding microtome (Leica SM 2000 R, Leica Biosystems, Nussloch, Germany) equipped with microtome knives (Leica 818, Leica Biosystems, Nussloch, Germany). The sections were stained in Safranin (Sigma-Aldrich, St. Louis, MO, USA) and Alcian Blue (Sigma-Aldrich, St. Louis, MO, USA) for 15–20 min. This was followed by dehydration in a graded ethanol series (50%, 70%, 90%, and 100%) and mounting in Euparal (Sigma-Aldrich, St. Louis, MO, USA) [49]. Observation of anatomical features was carried out using an Olympus BX43 light microscope, connected to an Olympus DP72 camera (Olympus Optical Co., Ltd., Tokyo, Japan) and Canon EOS 700D (Canon, Tokyo, Japan) using the Analysis Docu and EOS Utility 2 software. The images were saved in tagged image file format (TIFF). The same procedure described above was applied to all three replicates, and the representative photos were selected.
2.3. Scanning Electron Microscopy (SEM)
The samples of flowers (corolla petals and corona lobes) were cut on a wax plate (Cavex Holland BV, Haarlem, The Netherlands) into 0.5 cm × 0.5 cm using a Gillette knife blade for SEM observation. The samples were rinsed in phosphate buffer solution (PBS) (Sigma-Aldrich, St. Louis, MO, USA) (0.1 M, pH 7.4) for 10 min three times. Next, the dehydration was performed in a graded ethanol series (35%, 50%, 70%, 80%, 90%, 10 min per change, and finally in three changes of 99% ethanol) [16,50]. The samples were then dried through the critical point drying (CPD) (Leica® EM CPD 300) (Leica, Wetzlar, Germany) using liquid carbon dioxide (CO2) and coated with gold using a sputter coating device (Polaron SC 7680) (Polaron, London, UK) for 10 min. Finally, the micromorphology features were examined using a field emission scanning electron microscope (FESEM, Carl Zeiss Group, Oberkochen, Germany).
2.4. Histochemical Tests
Sudan Black B (SBB) (Sigma Aldrich, St. Louis, MO, USA) was used to detect the presence of lipids in the floral glands. First, the sections were immersed with SBB (20 min). Then, the sections were rinsed with an 80% ethanol solution and distilled water to remove any excess colour; SBB stained lipids black.
The periodic acid-Schiff reaction (PAS) (Sigma Aldrich, St. Louis, MO, USA) was used to identify the polysaccharides. First, the sections were immersed in a periodic acid solution (R&M Chemicals, London, UK) for 30 min. Next, the sections were rinsed with running water for 10 min and then immersed in Schiff’s reagent (Sigma Aldrich, St. Louis, MO, USA) for 20 min. The sections were then rinsed with distilled water before being immersed for 2 min in a 2% sodium bisulfite solution (R&M Chemicals, London, UK). Subsequently, the sections were rinsed again using distilled water for 10 min and 50% ethanol. As a result, the floral gland sections that contained carbohydrates were stained pink.
The presence of starch grains in the floral glands was detected using Lugol’s reagent (IKI) (Sigma Aldrich, St. Louis, MO, USA). The sections were immersed in an IKI solution for 10 min and then rinsed with distilled water. After IKI treatment, starch was stained violet-black, and proteins yellow.
Coomassie Blue (CB) (Sigma Aldrich, St. Louis, MO, USA) was used to detect proteins. First, the sections were immersed in a 0.25% CB solution for 15 min. After that, the sections were immersed in 7% acetic acid and rinsed with distilled water before being transferred on glass slides. Proteins were stained blue.
3. Results
3.1. Morphological Study
H. cagayanensis consisted of a white corolla (cl), corona lobe (co) with inner (ico) and outer (oco), while the guide rail (gr) was yellow and the pollinia (po) was dark chocolate; the gr and po were in between co (Figure 1A). While for H. lacunosa, the corolla (cl) was entirely white; the corona lobe (co) with inner (ico) and outer (oco) were light yellow; the pollinia (po) was bright yellow and between the co (Figure 2A). H. coriacea flowers had a bright yellow corolla (cl) and pollinia (po); the corona lobe (co) with inner (ico) and outer (oco) were both light yellow; the guide rail (gr) was purple; and gr and po were in between the co (Figure 3A).
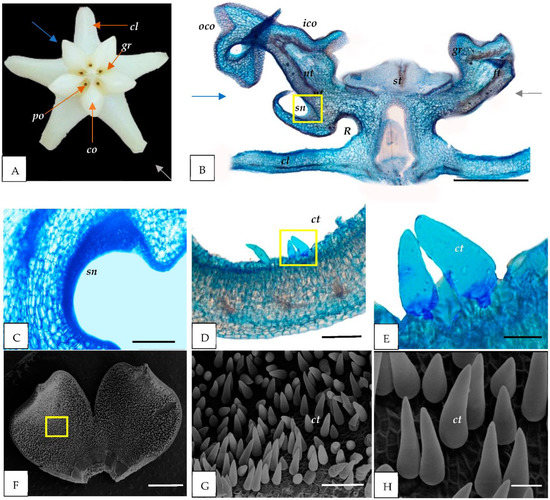
Figure 1.
Anatomical and micromorphological structure of the H. cagayanensis flower. (A) Flower of H. cagayanensis (front view) with five corolla petals (cl), five corona lobes (co) with inner (ico) and outer (oco), and five pollinia (po) and five guide rails (gr) in between each corona lobe. (B–E) LM images. (B) The longitudinal section of flower cut in a plane indicated by gray and blue arrows in (A); (C) secondary nectaries (sn) in the corona lobe (co); and (D,E) cross-sections of corolla petals with unicellular conical trichomes (ct) (yellow square). (F–H) SEM image of the petals (cl); (G,H) enlargement in (F) (yellow square) indicate the presence of ct. cl = corolla; co = corona; ft = filament tube; gr = guide rail; sn = secondary nectaries; po = pollinia R = nectar pool; nt = nectar tube; st = style head. Scale: (B) = 1000 µm; (C–E,H) = 200 µm; (F) = 1 mm; (G) =100 µm.
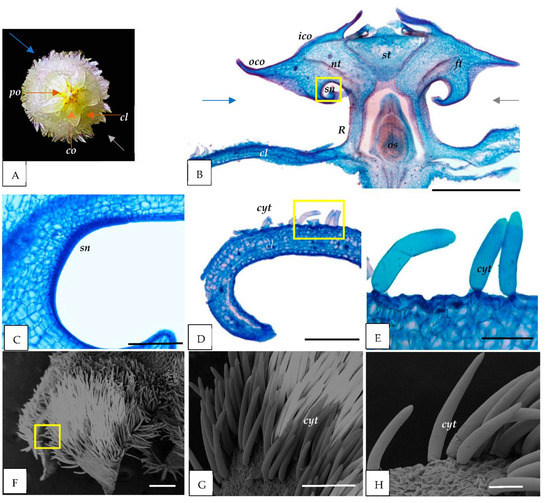
Figure 2.
Anatomical and micromorphological structure of H. lacunosa flower parts. (A) The flower of H. lacunosa (front view) consisted of five corolla petals (cl), five corona lobes (co) with inner (ico) and outer (oco), and five pollinia (po) in between each corona lobe. (B–E) LM images. (B) Longitudinal section of the flower cut in a plane indicated by gray and blue arrows in (A); (C) secondary nectaries (sn) observed in the corona lobe (co); and (D,E) cross-sections of corolla petals with unicellular cylindrical trichomes (cyt) (yellow square). (F–H) SEM image of the petals (cl); (G,H) enlargement in (F) (yellow square) show the presence of cyt. cl = corolla; co = corona; ft = filament tube; sn = secondary nectaries; R = nectar pool; nt = nectar tube; os = ovarian space; st = style head; po = pollinia. Scale: (B) = 1000 µm; (D) = 500 µm; (E,H) = 100 µm; (F) = 300 µm; (C,G) = 200 µm.
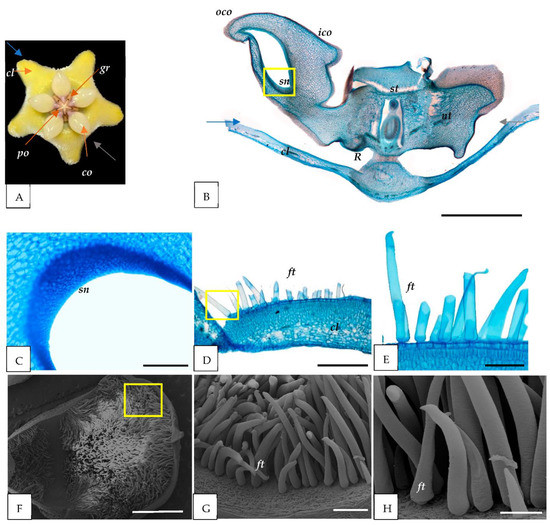
Figure 3.
Anatomical and micromorphological structure of the H. coriacea flower parts. (A) Flower of H. coriacea (front view) with five corolla petals (cl), five corona lobes (co) with inner (ico) and outer (oco), five pollinia (po), and five guide rail (gr) between each corona lobe. (B–E) LM images. (B) Longitudinal section of the flower cut in a plane indicated by gray and blue arrows in (A); (C) secondary nectaries (sn) cells observed in the corona lobe (co); and (D,E) cross-sections of corolla petals with unicellular falcate trichomes (ft) (yellow square) on the abaxial surface. (F–H) SEM image of the petals (cl); (G,H) enlargement in (F) (yellow square) show the presence of ft. cl = corolla; co = corona; gr = guide rail; sn = secondary nectaries; R = nectar pool; nt = nectar tube; st = style head. Scale: (B) = 2000 µm; (C) = 1000 µm; (D) = 500 µm; (E,H) = 200 µm; (F) = 1 mm; and (G) = 100 µm.
3.2. Light and Scanning Electron Microscopy
Our anatomical observations indicated the presence of secondary nectaries (sn) composed of a secretory epidermis and subepidermal layers of parenchyma cells of the corona lobe for all species studied (Figure 1C, Figure 2C and Figure 3C). Nearby, a nectar tube was located (Figure 1B, Figure 2B and Figure 3B). In addition, the nectar pool (reward area) (R) was discovered to be near the secondary nectaries and corolla portions of the flower (Figure 1B, Figure 2B and Figure 3B).
In this study, the corolla’s adaxial epidermis is covered with simple trichomes for all species (Figure 1D–H, Figure 2D–H and Figure 3D–H). H. cagayanensi’s trichomes are unicellular, conical (ct) with non-uniform width and height (average: width 30 μm and height 76 μm) (Figure 1E,H). In comparison, H. lacunosa’s trichomes are unicellular, cylindrical (cyt) with an average width of 31 μm and an average height of 233 μm (Figure 2E,H). For H. coriacea, the trichomes are unicellular, falcate (ft) with various sizes with an average width of 57 μm and height of 440 μm (Figure 3E,H). All trichomes in this study were observed to have a smooth surface.
3.3. Histochemical Tests
Histochemical tests were used to identify the secretion compounds in the secondary nectaries and trichomes of H. cagayanensis, H. lacunosa, and H. coriacea. The reaction with PAS reagents showed nectariferous tissue composed of polysaccharides with a pink colour in sn (Figure 4A, Figure 5A and Figure 6A). In Lugol’s solution, tiny starch grains were detected in sn with dark blue (Figure 4B, Figure 5B and Figure 6B). Coomassie blue detected proteins in sn, ct, cyt, and ft (Figure 4C,E, Figure 5C,E and Figure 6C,E). Sudan Black B-stained lipid droplets in sn, ct, cyt and ft with black (Figure 4D,F, Figure 5D,F and Figure 6D,F). The accumulation of proteins, lipids, polysaccharides, and starch grains in the secondary nectaries and trichomes of all studied species is shown in Table 2.
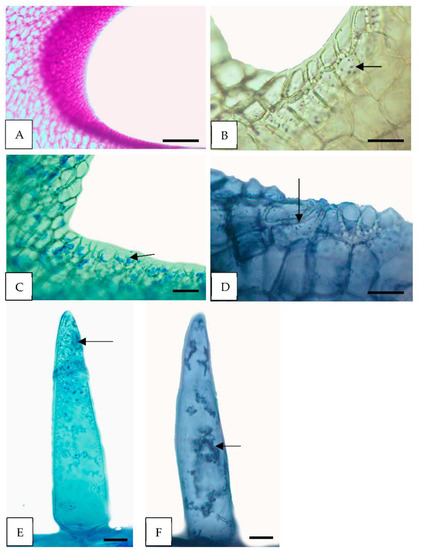
Figure 4.
Histochemical tests of the floral glands of H. cagayanensis; (A–D) Secondary nectaries (sn); (E,F) Conical trihomes (ct). (A) Polysaccharides stained pink in nectariferous tissue of sn (PAS); (B) tiny starch grains (arrows, IKI); (C,E) protein stains with CB; (D,F) few lipid droplets (arrows, SBB). PAS, periodic acid-Schiff reaction; IKI, Lugol’s reagent; CB, Commassie Blue; SBB, Sudan Black B. Scale: (A) =200 μm, (C) =100 μm; (B,D–F) =50 μm.
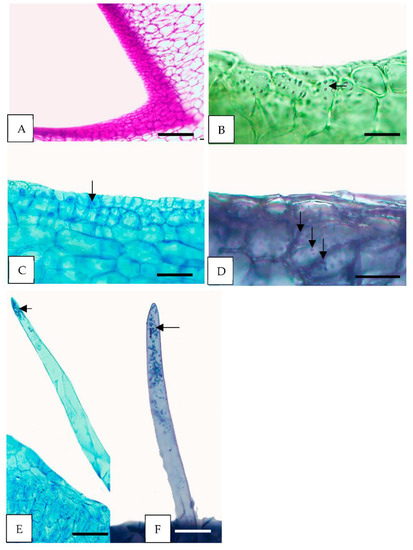
Figure 5.
Histochemical tests of the floral glands of H. lacunosa. (A–D) Fragments of secondary nectaries tissue (sn). (E,F) Cylindrical trichomes (cyt). (A) Polysaccharides in nectariferous tissue stained pink (PAS); (B) few starch grains (arrows, IKI); (C,E) protein stained with CB; (D,F) numerous blue-black lipid droplets in the cells (arrows, SBB). PAS = periodic acid-Schiff reaction; IKI = Lugol’s reagent; CB = Coomassie Blue; SBB = Sudan Black B. Scale: (A) = 200 µm, (D,F) = 100 µm, (B,C) = 50 µm.
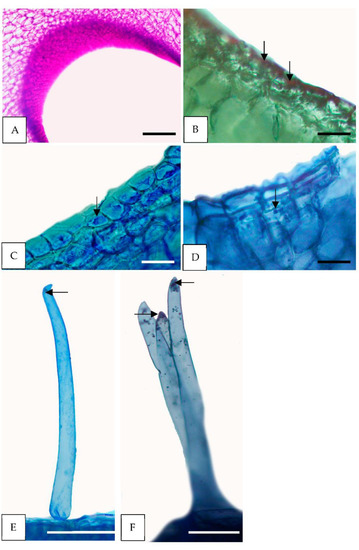
Figure 6.
Histochemical tests of the floral glands of H. coriacea. (A–D) Secondary nectaries (sn). (E,F) Falcate trichomes (ft). (A) Polysaccharides stained pink in the nectariferous tissue composed of epidermis and parenchyma of sn (PAS); (B) few starch grains (arrows, IKI). (C,E) the epidermal and subepidermal layers of sn and ft with proteins (arrows, CB); (D,F) note numerous blue-black lipid droplets (arrows, SBB). PAS = periodic acid-Schiff reaction; IKI = Lugol’s reagent; CB = Commassie Blue; SBB = Sudan Black B. Scale: (A,E,F) = 200 µm; (B–D) = 50 µm.
Table 2.
Histochemical analysis of the trichomes (t) and secondary nectaries (sn) in flowers of the three Hoya species.
4. Discussion
4.1. Nectary
Floral gland structures have been identified as the synthesis sites of various chemical compounds [51,52]. In the flowers of the Marsdenieae tribe, nine types of floral glands have been investigated: colleter, glandular trichome, laticifer, nectary (primary and secondary), osmophores, stile head, tapetum, and stylar canal [18]. In three Hoya species, we observed nectariferous tissue in the corona lobe, which signifies the presence of secondary nectaries. The secondary nectary features of these species resemble those described for some genera in the Asclepiadoideae [19,22,35,37,40,41,42]. Moreover, secondary nectaries in this subfamily were found to consist of the epidermis in the staminal corona lobe or several layers of epidermal and parenchymal nectarifers in the circle space of the corona lobe [42].
In histochemical tests, positive results were obtained for secondary nectaries for proteins, lipids, polysaccharides, and starch grains. The presence of starch grains in floral glands is a frequent feature of nectaries and osmophores, indicating that starch is used as a source of energy for the production of nectar and scent, respectively [52,53,54]. In addition, according to various studies, floral nectaries are attractive and provide rewards to the pollinators, and also may be used as plant defence mechanisms [18,55].
There are discrepancies regarding the presence of nectaries in asclepiads since this family has a complex flower structure, and different terms are used in describing the flower structure [18]. In most asclepiads, nectar is stored in the guide rail area. Still, the evolution of the corona lobe in most Hoya species, including H. carnosa, has resulted in the partial transfer of nectar from the guide rail area to secondary nectaries in the corona lobe [21,33,56,57]. Secondary nectaries are reported to play a role in releasing nectar compounds in the pollination process [42]. The presence of secondary nectaries on the cavity part of the corona lobe and nectar tubes on the side of the corona lobe are unique features of Hoya plants. However, the nectar tube on Hoya was discovered not to produce nectar and lost its function because it lacks nectariferous cells [56,57].
A study by Monteiro and Demarco [42] on nectaries found that the position of the nectar is significant in determining the mechanisms of pollinaria transfer in most asclepiadoids. Nectar is found to be produced in the corona lobe. In that case, the pollinaria will be moved by the pollinator’s legs, whereas if nectar is produced under the guide rail, the pollinaria will be moved by the pollinator’s proboscis [42,58,59,60]. The mechanism of pollinaria transfer is performed by the pollinator legs of Erebus ephesperis species in H. carnosa [61].
4.2. Trichomes
In this study, three different morphologies of simple trichomes were reported: conical trichomes (ct) in H. cagayanensis, cylindrical trichomes (cyt) in H. lacunosa, and falcate trichomes (ft) in H. coriacea. For the first time, these trichomes were identified to cover the corolla petal’s entire surface on the adaxial epidermis in all studied species. Interestingly, simple trichomes (long, multicellular, and uniseriate) have only been reported on the stem and leaves of Fischeria and Matelea in the subtribe Gonolobinae (Asclepiadoideae) [62]. Furthermore, the simple trichomes in our study demonstrated positive results for proteins and lipids in histochemical tests.
In this study, unicellular simple trichomes are abundant on the adaxial epidermis of the corolla petals of the three studied Hoya species. The unicellular simple trichome is a long single cell with a smooth cell wall surface that develops from protoderm cells without cell division [63]. Simple trichomes may act as a defence mechanism in the plant from various external factors, such as protecting animals, pathogen invasion, protecting plants tissues from extreme UV-light exposure, reducing excess heat on the plants, and increasing freezing tolerance and water absorption [44,64,65,66].
The existence, type, shape, size, structure, and position of trichomes also provide diagnostic features essential in plant taxonomic classification to distinguish closely related species [67,68]. Therefore, the identification of trichomes in this study could potentially be used as a reference in the classification of species in the genus Hoya as this is the first discovery of trichomes in Hoya.
5. Conclusions
For the first time, secondary nectaries and simple trichomes were discovered in the three Hoya species in this study. The discovery of trichomes in this study was also the first discovery in the genus Hoya. These findings give new information and a better understanding of the floral anatomy and micromorphology structures of Hoya.
Author Contributions
Conceptualisation, H.B.; methodology, S.B.; formal analysis, S.B., H.B.; investigation, M.F.M.S., M.R.A.R.; writing-draft preparation, review, and edition, S.B., H.B., N.T. and S.N.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Universiti Kebangsaan Malaysia, grant number DIP-2016-014 and GP-2021-K020959.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available in the paper.
Acknowledgments
We would like to thank the Faculty of Science and Technology, Universiti Kebangsaan Malaysia (UKM), for the instruments and facilities provided for this research. We also thank Scanning Electron Microscopy Unit FST, UKM.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rodda, M.; Simonsson, N. Hoya medinillifolia (Apocynaceae Asclepiadoideae), A New Species from Lowland Forests of Sarawak, Borneo. Webbia 2011, 66, 149–154. [Google Scholar] [CrossRef]
- Rodda, M.; Ercole, E. Hoya papaschonii (Apocynaceae: Asclepiadoideae), A New Species from Southern Thailand with a Peculiar Corona. Phytotaxa 2014, 175, 97–106. [Google Scholar] [CrossRef][Green Version]
- Rodda, M.; Omlor, R. The Taxonomy of Oreosparte (Apocynaceae: Asclepiadoideae). Webbia 2014, 68, 91–95. [Google Scholar] [CrossRef]
- Lamb, A.; Rodda, M.; Gokulsing, L.; Bosuang, S.; Rahayu, S. A Guide to Hoyas of Borneo; Natural History Publications: Kota Kinabalu, Malaysia, 2016. [Google Scholar]
- Rahayu. Hoya as a Medical Plant. Warta Kebun Raya 2011, 11, 15–21. [Google Scholar]
- Rabbani, A.; Rahmad, Z.; Rosazlina, R.; Akomolafe, G.; Edzham, S.M.; Azmi, F. The Diversity of Hoya (Apocynaceae: Asclepiadoideae) in Some Parts of Kedah and Perak, Peninsular Malaysia. Trop. Agric. Sci. 2021, 44, 193–203. [Google Scholar] [CrossRef]
- Rahmad, Z.; Rabbani, A.; Hamzah, S.M.E.S.; Hamzah, Z.; Faizal, M. Notes on the Hoya (Apocynaceae, Asclepiadoideae) from Pergau Forest Reserve, Kelantan, Peninsular Malaysia. Conf. Ser. Earth Environ. Sci. 2020, 549, 012031. [Google Scholar]
- Rodda, M.; Juhonewe, N.S.; Middleton, D.J. The Taxonomic Status of the Presumed Extinct Singaporean Hoya wallichii (Apocynaceae: Asclepiadoideae). Gard. Bull. Singap. 2016, 68, 75–187. [Google Scholar] [CrossRef]
- Rodda, M.; Rahmad, Z. Hoya peninsularis (Apocynaceae, Asclepiadoideae), a New Species from Peninsular Malaysia, and Notes on Hoya maingayi and Gongronema wrayi. Nord. J. Bot. 2020, 38, 42–52. [Google Scholar] [CrossRef]
- Salim, J.M.; Nikong, D. Notes on Hoya of Terengganu, Peninsular Malaysia. Malay. Nat. J. 2020, 72, 43–51. [Google Scholar]
- Lakshmi, S.R.; Benjamin, J.F.; Kumar, T.S.; Murthy, G.V.S.; Rao, M.V. In Vitro Propagation of Hoya wightii ssp. Palniensis KT Mathew, A Highly Vulnerable and Endemic Species of Western Ghats of Tamil Nadu, India. Afr. J. Biotechnol. 2009, 9, 84–92. [Google Scholar]
- Das, A.; Lee, S.H.; Hyun, T.K.; Kim, S.W.; Kim, J.Y. Plant Volatiles as Method of Communication. Plant Biotechnol. Rep. 2013, 7, 9–26. [Google Scholar] [CrossRef]
- Ramya, H.G.; Palanimuthu, V.; Rachna, S. An Introduction to Patchouli (Pogostemon cablin Benth.). A Medicinal and Aromatic Plant: It’s Importance to Mankind. Agric. Eng. Int. CIGR J. 2013, 15, 243–250. [Google Scholar]
- Effmert, U.; Grobe, J.; Rose, U.S.R.; Ehrig, F.; Kagi, R.; Piechulla, B. Volatile Composition, Emission Pattern, and Localization of Floral Scent Emission in Mirabilis jalapa (nyctaginaceae). Am. J. Bot. 2005, 92, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Pansarin, L.M.; Castro, M.D.M.; Sazima, M. Osmophore and Elaiophores of Grobya amherstiae (Catasetinae, Orchidaceae) and Their Relation to Pollination. Bot. J. Linn. Soc. 2009, 159, 408–415. [Google Scholar] [CrossRef]
- Wiemer, A.P.; More, M.; Benitez-Vieyra, S.; Cocucci, A.A.; Raguso, R.A.; Sérsic, A.N. A Simple Floral Fragrance and Unusual Osmophore Structure in Cyclopogon elatus (Orchidaceae). Plant Biol. 2009, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Stpiczyńska, M.; Davies, K.L.; Świątek, P.; de Miranda, V.F.O. Floral Ultrastructure of Two Brazilian Aquatic–Epiphytic Bladderworts: Utricularia cornigera Studnička and U. nelumbifolia Gardner (Lentibulariaceae). Protoplasma 2017, 254, 353–366. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Demarco, D. Floral Glands in Asclepiads: Structure, Diversity and Evolution. Acta Bot. Bras. 2017, 31, 477–502. [Google Scholar] [CrossRef]
- Rao, V.S.; Ganguli, A. The Floral Anatomy of Some Asclepiadaceae. Proc. Indian Acad. Sci. 1963, 57, 15–44. [Google Scholar] [CrossRef]
- Kunze, H. Structure and Function in Asclepiad Pollination. Plant Syst. Evol. 1991, 176, 227–253. [Google Scholar] [CrossRef]
- Kunze, H. Corona and Nectar System in Asclepiadinae (Asclepiadaceae). Flora 1997, 192, 175–183. [Google Scholar] [CrossRef]
- Valente, M.C. Matelea maritima subsp. ganglinosa (Vell.) Font.—Anatomia e Vascularização Floral (Asclepiadaceae). Arq. Jard. Botânico Rio De Jan. 1995, 33, 75–98. [Google Scholar]
- Valente, M.C.; Costa, C.G. Estudo Anatômico da Flor de Marsdenia loniceroides E. Fournier (Asclepiadoideae—Apocynaceae). Rodriguésia 2005, 56, 51–66. [Google Scholar] [CrossRef]
- Castro, M.D.; Demarco, D. Phenolic Compounds Produced by Secretory Structures in Plants: A Brief Review. Nat. Prod. Commun. 2008, 3, 1273–1284. [Google Scholar]
- Wiemer, A.P.; Sérsic, A.N.; Marino, S.; Simões, A.O.; Cocucci, A.A. Functional Morphology and Wasp Pollination of Two South American Asclepiads (Asclepiadoideae—Apocynaceae). Ann. Bot. 2012, 109, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Demarco, D. Secretory Tissues, and the Morphogenesis and Histochemistry of Pollinarium in Flowers of Asclepiadeae (Apocynaceae). Int. J. Plant Sci. 2014, 175, 1042–1053. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Biology of Floral Scent; CRC Press: London, UK, 2006. [Google Scholar]
- Poinar, G.; Poinar, G. The Antiquity of Floral Secretory Tissues That Provide Today’s Fragrances. Hist. Biol. 2020, 32, 494–499. [Google Scholar] [CrossRef]
- Fahn, A. Secretory Tissues in Plants; Academic Press: London, UK, 1979. [Google Scholar]
- Chwil, M.; Kostryco, M.; Matraszek–Gawron, R. Comparative Studies on Structure of the Floral Nectaries and the Abundance of Nectar Produc-Tion of Prunus laurocerasus L. Protoplasma 2019, 256, 1705–1726. [Google Scholar] [CrossRef]
- Konarska, A.; Masierowska, M. Structure of Floral Nectaries and Female–Biased Nectar Production in Protandrous Species Geranium macrorrhizum and Geranium phaeum. Protoplasma 2020, 257, 501–523. [Google Scholar] [CrossRef]
- Faegri, K.; van der Pijl, L. The Principles of Pollination Ecology; Pergamon Press: New York, NY, USA, 1979. [Google Scholar]
- Galil, J.; Zeroni, M. Nectar System of Asclepias curassavica. Bot. Gaz. 1965, 126, 144–148. [Google Scholar] [CrossRef]
- Christ, P.; Schnepf, E. The Nectaries of Cynanchum vincetoxicum (Asclepiadaceae). Isr. J. Plant Sci. 1985, 34, 79–90. [Google Scholar]
- Kunze, H. Floral Morphology of some Gonolobeae (Asclepiadaceae). Bot. Jahrb. 1995, 117, 211–238. [Google Scholar]
- Kunze, H.; Liede, S. Observations on Pollination in Sarcostemma (Asclepiadaceae). Plant Syst. Evol. 1991, 178, 95–105. [Google Scholar] [CrossRef]
- Kunze, H. Pollination Ecology in Two Species of Gonolobus (Asclepiadaceae). Flora 1999, 194, 309–316. [Google Scholar] [CrossRef]
- Endress, M.E.; Bruyns, P.V. A Revised Classification of the Apocynaceae sl. Bot. Rev. 2000, 66, 1–56. [Google Scholar] [CrossRef]
- Vieira, M.F.; Shepherd, G.J. Oxypetalum banksii subsp. Banksii: A Taxon of Asclepiadaceae with an Extragynoecial Compitum. Plant Syst. Evol. 2002, 233, 199–206. [Google Scholar] [CrossRef]
- Valente, M.D.C.; Silva, N.M.F.D. Anatomia Floral de Barjonia erecta (Vell.) Schum. (Asclepiadaceae). Rodriguésia 1984, 36, 95–106. [Google Scholar] [CrossRef]
- Bruyns, P.V. A Revision of Hoodia and Lavrania (Asclepiadaceae-Stapelieae). Master’s Thesis, University of Cape Town, Cape Town, South Africa, 1994. [Google Scholar]
- Monteiro, M.M.; Demarco, D. Corona Development and Floral Nectaries of Asclepiadeae (Asclepiadoideae, Apocynaceae). Acta Bot. Bras. 2017, 31, 420–432. [Google Scholar] [CrossRef]
- Spence, J. Plant Histology. In Plant Cell Biology; Hawes, C., Satiat-Jeunemaitre, B., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 189–206. [Google Scholar]
- Zhao, Q.; Chen, X.Y. Development: A New Function of Plant Trichomes. Nat. Plants 2016, 2, 16096. [Google Scholar] [CrossRef]
- Kundan, M.; Gani, U.; Nautiyal, A.K.; Misra, P. Molecular Biology of Glandular Trichomes and Their Functions in Environmental Stresses. In Molecular Approaches in Plant Biology and Environmental Challenge; Singh, S.P., Upadhyay, S.K., Pandey, A., Kumar, S., Eds.; Springer: Singapore, 2019; pp. 365–393. [Google Scholar]
- Gershenzon, J.; Dudareva, N. The Function of Terpene Natural Products in the Natural World. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Basir, S.; Akbar, M.A.; Talip, N.; Baharum, S.N.; Bunawan, H. An Integrative Volatile Terpenoid Profiling and Transcriptomics Analysis in Hoya cagayanensis, Hoya lacunosa and Hoya coriacea (Apocynaceae, Marsdenieae). Horticulturae 2022, 8, 224. [Google Scholar] [CrossRef]
- Puchtler, H.; Sweat Waldrop, F.; Conner, H.M.; Terry, M.S. Carnoy Fixation: Practical and Theoretical Considerations. Histochemie 1968, 16, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Noraini, M.A.A.; Ruzi, M. Plant Anatomy and Microscopy; Penerbit Universiti Kebangsaan Malaysia: Bangi, Malaysia, 2019. [Google Scholar]
- Kowalkowska, A.K.; Kozieradzka–Kiszkurno, M.; Turzynski, S. Morphological, Histological and Ultrastructural Features of Osmophores and Nectary of Bulbophyllum wendlandianum (Kraenzl.) Dammer (B. section Cirrhopetalum Lindl., Bulbophyllinae Schltr., Orchidaceae). Plant Syst. Evol. 2015, 301, 609–622. [Google Scholar] [CrossRef]
- Stern, W.L.; Curry, K.J.; Whitten, W.M. Staining Fragrance Glands in Orchid Flowers. Bull. Torrey Bot. Club 1986, 113, 288–297. [Google Scholar] [CrossRef]
- Vogel, S. The Role of Scent Glands Pollination; The National Science Foundation: Washington, DC, USA, 1990. [Google Scholar]
- Nepi, M. Nectary Structure and Ultrastructure. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 129–166. [Google Scholar]
- Pacini, E.; Nepi, M. Nectar production and presentation. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 167–214. [Google Scholar]
- Galetto, L.; Bernardello, G.; Sosa, C.A. The Relationship between Floral Nectar Composition and Visitors in Lycium (Solanaceae) from Argentina and Chile: What Does It Reflect? Flora 1998, 193, 303–314. [Google Scholar] [CrossRef]
- Kunze, H.; Wanntorp, L. The Gynostegium of Hoya spartioides (Apocynaceae–Asclepiadoideae): A Striking Case of Incongruence between Molecular and Phenotypic Evolution. Org. Divers. Evol. 2008, 8, 346–357. [Google Scholar] [CrossRef][Green Version]
- Wanntorp, L.; Kunze, H. Identifying Synapomorphies in the Flowers of Hoya and Dischidia, toward Phylogenetic Understanding. Int. J. Plant Sci. 2009, 170, 331–342. [Google Scholar] [CrossRef]
- Eisikowitch, D. Morpho–Ecological Aspects on the Pollination of Calotropis procera (Asclepiadaceae) in Israel. Plant Syst. Evol. 1986, 152, 185–194. [Google Scholar] [CrossRef]
- Lumer, C.; Yost, S.E. The Reproductive Biology of Vincetoxicum nigrum (L.) Moench (Asclepiadaceae), a Mediterranean Weed in New York State. Bull. Torrey Bot. Club 1995, 122, 15–23. [Google Scholar] [CrossRef]
- Vieira, M.F.; Shepherd, G.J. Pollinators of Oxypetalum (Asclepiadaceae) in Southeastern Brazil. Rev. Bras. Biol. 1999, 59, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, K.; Furukawa, S.; Kawakita, A. Pollinia Transfer on Moth Legs in Hoya carnosa (Apocynaceae). Am. J. Bot. 2017, 104, 953–960. [Google Scholar] [CrossRef]
- Medina, M.C.; Sousa-Baena, M.S.; Capelli, N.D.V.; Koch, R.; Demarco, D. Stinging Trichomes in Apocynaceae and Their Evolution in Angiosperms. Plants 2021, 10, 2324. [Google Scholar] [CrossRef] [PubMed]
- Ramayya, N. Classification, and Phylogeny of the Trichomes of Angiosperms. In Research Trends in Plant Anatomy; Ghouse, A.K.M., Yunus, M., Eds.; Tata McGraw-Hill Publishing Company Ltd.: New Delhi, India, 1972. [Google Scholar]
- Werker, E. Trichome Diversity and Development. In Advances in Botanical Research. Plant Trichomes; Hallahan, D.L., Gray, J.C., Eds.; Academic Press: New York, NY, USA, 2000; pp. 1–35. [Google Scholar]
- Bickford, C.P. Ecophysiology of Leaf Trichomes. Funct. Plant Biol. 2016, 43, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Dalin, P.; Ågren, J.; Björkman, C.; Huttunen, P.; Kärkkäinen, K. Leaf Trichome Formation and Plant Resistance to Herbivory. In Induced Plant Resistance to Herbivory; Springer: Dordrecht, The Netherlands, 2008; pp. 89–105. [Google Scholar]
- Moraes, T.M.S.; Guilherme, R.R.; Camilla, R.A.; Sebastião, J.S.N.; Maura, D.C. Comparative Leaf Anatomy and Micromorphology of Psychotria Species (Rubiaceae) from the Atlantic Rainforest. Acta Bot. Bras. 2011, 25, 178–190. [Google Scholar] [CrossRef]
- Spring, O. Chemotaxonomy Based on Metabolites from Glandular Trichomes. Adv. Bot. Res. 2000, 31, 153–174. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).